A Review of Environment Effects on Nitrate Accumulation in Leafy Vegetables Grown in Controlled Environments

 , ,
, ,
Abstract
:1. Introduction
2. Nitrate Assimilation and Accumulation in Vegetables in Response to Light Conditions
2.1. The Effects of Light Intensity
2.2. The Function of Light Duration or Photoperiod
2.3. The Function of Light Spectra
2.3.1. Red and Blue Light
2.3.2. Green Light, UV Light, and Far-Red Light
3. Nitrate Uptake, Distribution, and Assimilation in Response to Growth Temperature
4. Nitrate Assimilation and Accumulation in Response to Elevated Carbon Dioxide Concentration
5. Nitrate Assimilation and Accumulation in Response to Fertilizer Management
6. Nitrate Uptake, Distribution, and Accumulation in Response to Water Quality and Irrigation Management
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Crawford, N.M. Nitrate: Nutrient and signal for plant growth. Plant Cell 1995, 7, 859. [Google Scholar] [PubMed] [Green Version]
- Agostini, F.; Tei, F.; Silgram, M.; Farneselli, M.; Benincasa, P.; Aller, M. Decreasing nitrate leaching in vegetable crops with better N management. In Genetic Engineering, Biofertilisation, Soil Quality and Organic Farming; Springer: Berlin, Germany, 2010; pp. 147–200. [Google Scholar]
- Ju, X.T.; Xing, G.X.; Chen, X.P.; Zhang, S.L.; Zhang, L.J.; Liu, X.J.; Cui, Z.L.; Yin, B.; Christie, P.; Zhu, Z.L. Reducing environmental risk by improving N management in intensive Chinese agricultural systems. Proc. Natl. Acad. Sci. USA 2009, 106, 3041–3046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutton, M.A.; Bleeker, A.; Howard, C.M.; Bekunda, M.; Grizzetti, B.; de Vries, W.; van Grinsven, H.J.M.; Abrol, Y.P.; Adhya, T.K.; Billen, G.; et al. Our Nutrient World: The Challenge to Produce More Food and Energy with Less Pollution. Global Overview of Nutrient Management; Centre for Ecology and Hydrology on behalf of the Global Partnership on Nutrient Management and the International Nitrogen Initiative: Edinburgh, UK, 2013. [Google Scholar]
- Bian, Z.H.; Yang, Q.C.; Liu, W.K. Effects of light quality on the accumulation of phytochemicals in vegetables produced in controlled environments: A review. J. Sci. Food Agric. 2015, 95, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Bian, Z.H.; Cheng, R.F.; Yang, Q.C.; Wang, J.; Lu, C.G. Continuous light from red, blue, and green light-emitting diodes reduces nitrate concentration and enhances phytochemical concentrations and antioxidant capacity in lettuce. J. Am. Soc. Hortic. Sci. 2016, 141, 186–195. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Fan, X.; Miller, A.J. Plant nitrogen assimilation and use efficiency. Annu. Rev. Plant Biol. 2012, 63, 153–182. [Google Scholar] [CrossRef] [Green Version]
- Ashworth, A.; Mitchell, K.; Blackwell, J.R.; Vanhatalo, A.; Jones, A.M. High-nitrate vegetable diet increases plasma nitrate and nitrite concentrations and reduces blood pressure in healthy women. Public Health Nutr. 2015, 18, 2669–2678. [Google Scholar] [CrossRef] [Green Version]
- Ashworth, A.; Bescos, R. Dietary nitrate and blood pressure: Evolution of a new nutrient? Nutr. Res. Rev. 2017, 30, 208–219. [Google Scholar] [CrossRef] [Green Version]
- Chan, T.Y. Vegetable-borne nitrate and nitrite and the risk of methaemoglobinaemia. Toxicol. Lett. 2011, 200, 107–108. [Google Scholar] [CrossRef]
- Song, P.; Wu, L.; Guan, W. Dietary nitrates, nitrites, and nitrosamines intake and the risk of gastric cancer: A meta-analysis. Nutrients 2015, 7, 9872–9895. [Google Scholar] [CrossRef]
- Joint FAO/WHO Expert Committee on Food Additives (JECFA). Evaluation of Certain Food Additives and Contaminants: Eightieth Report of the Joint FAO/WHO Expert Committee on Food Additives; World Health Organization (WHO): Geneva, Switzerland, 2016; Volume 995, p. 132. [Google Scholar]
- Van Velzen, A.G.; Sips, A.J.A.M.; Schothorst, R.C.; Lambers, A.C.; Meulenbelt, J. The oral bioavailability of nitrate from nitrate-rich vegetables in humans. Toxicol. Lett. 2008, 181, 177–181. [Google Scholar] [CrossRef]
- Bian, Z.H.; Cheng, R.C.; Wang, Y.; Yang, Q.C.; Lu, C.G. Effect of green light on nitrate reduction and edible quality of hydroponically grown lettuce (Lactuca sativa L.) under short-term continuous light from red and blue light-emitting diodes. Environ. Exp. Bot. 2018, 153, 63–71. [Google Scholar] [CrossRef] [Green Version]
- Colonna, E.; Rouphael, Y.; Barbieri, G.; De Pascale, S. Nutritional quality of ten leafy vegetables harvested at two light intensities. Food Chem. 2016, 199, 702–710. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.H.; Huang, M.Y.; Huang, W.D.; Hsu, M.H.; Yang, Z.W.; Yang, C.M. The effects of red, blue, and white light-emitting diodes on the growth, development, and edible quality of hydroponically grown lettuce (Lactuca sativa L. var. capitata). Sci. Hortic. 2013, 150, 86–91. [Google Scholar] [CrossRef]
- Lam, H.M.; Coschigano, K.T.; Oliveira, I.C.; Melo-Oliveira, R.; Coruzzi, G.M. The molecular-genetics of nitrogen assimilation into amino acids in higher plants. Annu. Rev. Plant. Biol. 1996, 47, 569–593. [Google Scholar] [CrossRef] [PubMed]
- Hickman, G. International greenhouse vegetable production-statistics. In A Review of Currently Available Data on the International Production of Vegetables in Greenhouses; Cuesta roble greenhouse consultants: Mariposa, CA, USA, 2016. [Google Scholar]
- Sabir, N.; Singh, B. Protected cultivation of vegetables in global arena: A review. Indian J. Agri. Sci. 2013, 83, 123–135. [Google Scholar]
- Khan, F.A. A review on hydroponic greenhouse cultivation for sustainable agriculture. Int. J. Agric. Environ. Food Sci. 2018, 2, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Anjana, S.U.; Iqbal, M. Nitrate accumulation in plants, factors affecting the process, and human health implications. A review. Agron. Sustain. Dev. 2007, 27, 45–57. [Google Scholar] [CrossRef]
- Bindraban, P.S.; Dimkpa, C.; Nagarajan, L.; Roy, A.; Rabbinge, R. Revisiting fertilisers and fertilisation strategies for improved nutrient uptake by plants. Biol. Fertil. Soils 2015, 51, 897–911. [Google Scholar] [CrossRef] [Green Version]
- Gruda, N. Impact of environmental factors on product quality of greenhouse vegetables for fresh consumption. Crit. Rev. Plant Sci. 2005, 24, 227–247. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Tsay, Y.F. Arabidopsis nitrate transporter NRT1.9 is important in phloem nitrate transport. Plant Cell 2011, 23, 1945–1957. [Google Scholar] [CrossRef] [Green Version]
- Iglesias-Bartolomé, R.; González, C.A.; Kenis, J.D. Nitrate reductase dephosphorylation is induced by sugars and sugar-phosphates in corn leaf segments. Physiol. Plant. 2004, 122, 62–67. [Google Scholar] [CrossRef]
- Busch, F.A.; Sage, R.F.; Farquhar, G.D. Plants increase CO2 uptake by assimilating nitrogen via the photorespiratory pathway. Nat. Plants 2018, 4, 46. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.L.; Jin, Y.; Yang, Y.J.; Li, G.J.; Boyer, J.S. Sugar input, metabolism, and signaling mediated by invertase: Roles in development, yield potential, and response to drought and heat. Mol. Plant 2010, 3, 942–955. [Google Scholar] [CrossRef]
- Miller, A.J.; Fan, X.; Orsel, M.; Smith, S.J.; Wells, D.M. Nitrate transport and signalling. J. Exp. Bot. 2007, 58, 2297–2306. [Google Scholar] [CrossRef]
- Walker, B.J.; Strand, D.D.; Kramer, D.M.; Cousins, A.B. The response of cyclic electron flow around photosystem I to changes in photorespiration and nitrate assimilation. Plant Physiol. 2014, 165, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Van der Boon, J.; Steenhuizen, J.; Steingrover, E.G. Growth and nitrate concentration of lettuce as affected by total nitrogen and chloride concentration, NH4/NO3 ratio and temperature of the recirculating nutrient solution. J. Hortic. Sci. 1990, 65, 309–321. [Google Scholar] [CrossRef]
- Nieuwhof, M. Effects of temperature and light on nitrate content of radish (Raphanus sativus L.). Gartenbauwissenschaft 1995, 59, 220–224. [Google Scholar]
- Gangappa, S.N.; Botto, J.F. The multifaceted roles of HY5 in plant growth and development. Mol. Plant 2016, 9, 1353–1365. [Google Scholar] [CrossRef] [Green Version]
- Osterlund, M.T.; Hardtke, C.S.; Wei, N.; Deng, X.W. Targeted destabilization of HY5 during light-regulated development of Arabidopsis. Nature 2000, 405, 462–466. [Google Scholar] [CrossRef]
- Calatayud, Á.; Gorbe, E.; Roca, D.; Martínez, P.F. Effect of two nutrient solution temperatures on nitrate uptake, nitrate reductase activity, NH4+ concentration and chlorophyll a fluorescence in rose plants. Environ. Exp. Bot. 2008, 64, 65–74. [Google Scholar] [CrossRef]
- Gent, M.P.N. Effect of degree and duration of shade on quality of greenhouse tomato. Hortscience 2007, 42, 514–520. [Google Scholar] [CrossRef]
- Demšar, J.; Osvald, J.; Vodnik, D. The effect of light-dependent application of nitrate on the growth of aeroponically grown lettuce (Lactuca sativa L.). J. Am. Soc. Hortic. Sci. 2004, 129, 570–575. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Li, H.; Yu, J.; Liu, H.; Cao, Z.; Manukovsky, N.; Liu, H. Interaction effects of light intensity and nitrogen concentration on growth, photosynthetic characteristics and quality of lettuce (Lactuca sativa L. Var. youmaicai). Sci. Hortic. 2017, 214, 51–57. [Google Scholar] [CrossRef]
- Zheng, Y.J.; Zhang, Y.T.; Liu, H.C.; Li, Y.M.; Liu, Y.L.; Hao, Y.W.; Lei, B.F. Supplemental blue light increases growth and quality of greenhouse pak choi depending on cultivar and supplemental light intensity. J. Integr. Agric. 2018, 17, 2245–2256. [Google Scholar] [CrossRef] [Green Version]
- Samuolienė, G.; Brazaitytė, A.; Jankauskienė, J.; Viršilė, A.; Sirtautas, R.; Novičkovas, A.; Sakalauskienė, S.; Sakalauskaitė, J.; Duchovskis, P. LED irradiance level affects growth and nutritional quality of Brassica microgreens. Open Life Sci. 2013, 8, 1241–1249. [Google Scholar] [CrossRef]
- Perez-Lopez, U.; Miranda-Apodaca, J.; Munoz-Rueda, A.; Mena-Petite, A. Interacting effects of high light and elevated CO2 on the nutraceutical quality of two differently pigmented Lactuca sativa cultivars (Blonde of Paris Batavia and Oak Leaf). Sci. Hortic. 2015, 191, 38–48. [Google Scholar] [CrossRef]
- Jonassen, E.M.; Lea, U.S.; Lillo, C. HY5 and HYH are positive regulators of nitrate reductase in seedlings and rosette stage plants. Planta 2008, 227, 559–564. [Google Scholar] [CrossRef]
- Fait, A.; Sienkiewicz-Porzucek, A.; Fernie, A.R. Metabolomics approaches to advance understanding of nitrogen assimilation and carbon–nitrogen interactions. Annu. Plant Rev. Online 2018, 6, 249–268. [Google Scholar]
- An, J.P.; Qu, F.J.; Yao, J.F.; Wang, X.N.; You, C.X.; Wang, X.F.; Hao, Y.J. The bZIP transcription factor MdHY5 regulates anthocyanin accumulation and nitrate assimilation in apple. Hortic. Res. 2017, 4, 17023. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Yao, Q.; Gao, X.; Jiang, C.; Harberd, N.P.; Fu, X. Shoot-to-root mobile transcription factor HY5 coordinates plant carbon and nitrogen acquisition. Curr. Biol. 2016, 26, 640–646. [Google Scholar] [CrossRef]
- Lillo, C. Signalling cascades integrating light-enhanced nitrate metabolism. Biochem. J. 2008, 415, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Nunes-Nesi, A.; Fernie, A.R.; Stitt, M. Metabolic and signaling aspects underpinning the regulation of plant carbon nitrogen interactions. Mol. Plant 2010, 3, 973–996. [Google Scholar] [CrossRef] [PubMed]
- Burns, I.G.; Zhang, K.; Turner, M.K.; Edmondson, R. Iso-osmotic regulation of nitrate accumulation in lettuce. J. Plant Nutr. 2010, 34, 283–313. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Chang, D.; Yang, H.; Wang, J.; Shi, H. Metabolome and molecular basis for carbohydrate increase and nitrate reduction in burley tobacco seedlings by glycerol through upregulating carbon and nitrogen metabolism. Sci. Rep. 2018, 8, 13300. [Google Scholar] [CrossRef] [Green Version]
- Cheung, C.M.; Poolman, M.G.; Fell, D.A.; Ratcliffe, R.G.; Sweetlove, L.J. A diel flux balance model captures interactions between light and dark metabolism during day-night cycles in C3 and crassulacean acid metabolism leaves. Plant Physiol. 2014, 165, 917–929. [Google Scholar] [CrossRef] [Green Version]
- Tegeder, M.; Masclaux-Daubresse, C. Source and sink mechanisms of nitrogen transport and use. New Phytol. 2018, 217, 35–53. [Google Scholar] [CrossRef] [Green Version]
- He, Y.N.; Peng, J.S.; Cai, Y.; Liu, D.F.; Guan, Y.; Yi, H.Y.; Gong, J.M. Tonoplast-localized nitrate uptake transporters involved in vacuolar nitrateefflux and reallocation in Arabidopsis. Sci. Rep. 2017, 7, 6417. [Google Scholar] [CrossRef]
- Neely, H.L.; Koenig, R.T.; Miles, C.A.; Koenig, T.C.; Karlsson, M.G. Diurnal fluctuation in tissue nitrate concentration of field-grown leafy greens at two latitudes. Hort Sci. 2010, 45, 1815–1818. [Google Scholar] [CrossRef] [Green Version]
- Zhou, W.; Liu, W.; Yang, Q. Quality changes in hydroponic lettuce grown under pre-harvest short-duration continuous light of different intensities. J. Hortic. Sci. Biotechnol. 2012, 87, 429–434. [Google Scholar] [CrossRef]
- Jonassen, E.M.; Sandsmark, B.A.; Lillo, C. Unique status of NIA2 in nitrate assimilation: NIA2 expression is promoted by HY5/HYH and inhibited by PIF4. Plant Signal. Behav. 2009, 4, 1084–1086. [Google Scholar] [CrossRef] [Green Version]
- Jonassen, E.M.; Sévin, D.C.; Lillo, C. The bZIP transcription factors HY5 and HYH are positive regulators of the main nitrate reductase gene in Arabidopsis leaves, NIA2, but negative regulators of the nitrate uptake gene NRT1.1. J. Plant Physiol. 2009, 166, 2071–2076. [Google Scholar] [CrossRef]
- Meyer, C.; Lea, U.S.; Provan, F.; Kaiser, W.M.; Lillo, C. Is nitrate reductase a major player in the plant NO (nitric oxide) game? Photosynth. Res. 2005, 83, 181–189. [Google Scholar] [CrossRef]
- Krapp, A. Plant nitrogen assimilation and its regulation: A complex puzzle with missing pieces. Curr. Opin. Plant Biol. 2015, 25, 115–122. [Google Scholar] [CrossRef]
- Maevskaya, S.; Bukhov, N. Effect of light quality on nitrogen metabolism of radish plants. Russ. J. Plant Physiol. 2005, 52, 304–310. [Google Scholar] [CrossRef]
- Sakuraba, Y.; Yanagisawa, S. Light signalling-induced regulation of nutrient acquisition and utilisation in plants. Semin. Cell Dev. Biol. 2018, 83, 123–132. [Google Scholar] [CrossRef]
- Chang, C.L.; Chang, K.P. The growth response of leaf lettuce at different stages to multiple wavelength-band light-emitting diode lighting. Sci. Hortic. 2014, 179, 78–84. [Google Scholar] [CrossRef]
- Brazaitytė, A.; Viršile, A.; Jankauskiene, J.; Sakalauskiene, S.; Samuoliene, G.; Sirtautas, R.; Novičkovas, A.; Dabašinskas, L.; Miliauskiene, J.; Vaštakaite, V.; et al. Effect of supplemental UV-A irradiation in solid-state lighting on the growth and phytochemical content of microgreens. Int. Agrophysics 2015, 29, 13–22. [Google Scholar] [CrossRef]
- Chen, X.; Xue, X.Z.; Guo, W.; Wang, L.; Qiao, X. Growth and nutritional properties of lettuce affected by mixed irradiation of white and supplemental light provided by light-emitting diode. Sci. Hortic. 2016, 200, 111–118. [Google Scholar] [CrossRef]
- Samuolienė, G.; Brazaitytė, A.; Sirtautas, R.; Novičkovas, A.; Duchovskis, P. The effect of supplementary LED lighting on the antioxidant and nutritional properties of lettuce. Acta Hortic. 2012, 952, 835–841. [Google Scholar] [CrossRef]
- Bliznikas, Z.; Žukauskas, A.; Samuoliene, G.; Viršile, A.; Brazaityte, A.; Jankauskiene, J.; Duchovskis, P.; Novičkovas, A. Effect of supplementary pre-harvest LED lighting on the antioxidant and nutritional properties of green vegetables. Acta Hortic. 2012, 939, 85–91. [Google Scholar] [CrossRef]
- Johnson, C.F.; Langhans, R.W.; Albright, L.D.; Combs, G.F.; Welch, R.M.; Heller, L.; Glahn, R.P.; Wheeler, R.M.; Goins, G.D. Spinach: Nitrate analysis of an advanced life support (ALS) crop cultured under ALS candidate artificial light sources. SAE Tech. Paper 1999, 1–5. [Google Scholar] [CrossRef]
- Han, X.; Tohge, T.; Lalor, P.; Dockery, P.; Devaney, N.; Esteves-Ferreira, A.A.; Fernie, A.R.; Sulpice, R. Phytochrome A and B regulate primary metabolism in Arabidopsis leaves in response to light. Front. Plant Sci. 2017, 8, 1394. [Google Scholar] [CrossRef] [Green Version]
- Monostori, I.; Heilmann, M.; Kocsy, G.; Rakszegi, M.; Ahres, M.; Altenbach, S.; Szalai, G.; Pál, M.; Toldi, D.; Simon-Sarkadi, L. LED lighting–modification of growth, metabolism, yield and flour composition in wheat by spectral quality and intensity. Front. Plant Sci. 2018, 9, 605. [Google Scholar] [CrossRef]
- Lillo, C.; Appenroth, K.J. Light regulation of nitrate reductase in higher plants: Which photoreceptors are involved? Plant Biol. 2001, 3, 455–465. [Google Scholar] [CrossRef]
- Appenroth, K.J.; Meço, R.; Jourdan, V.; Lillo, C. Phytochrome and post-translational regulation of nitrate reductase in higher plants. Plant Sci. 2000, 159, 51–56. [Google Scholar] [CrossRef]
- Chandok, M.R.; Sopory, S.K. Phosphorylation/dephosphorylation steps are key events in the phytochrome-mediated enhancement of nitrate reductase mRNA levels and enzyme activity in maize. Mol. Gen. Genet. 1996, 251, 599–608. [Google Scholar] [CrossRef]
- Qi, L.; Liu, S.; Xu, L.; Yu, W.; Liang, Q.; Hao, S. Effects of light qualities on accumulation of oxalate, tannin and nitrate in spinach. Trans. Chin. Soc. Agric. Eng. 2007, 23, 201–205. [Google Scholar]
- Osterlund, M.T.; Wei, N.; Deng, X.W. The roles of photoreceptor systems and the COP1-targeted destabilization of HY5 in light control of Arabidopsis seedling development. Plant Physiol. 2000, 124, 1520–1524. [Google Scholar] [CrossRef] [Green Version]
- Castillon, A.; Shen, H.; Huq, E. Blue light induces degradation of the negative regulator phytochrome interacting factor 1 to promote photomorphogenic development of Arabidopsis seedlings. Genetics 2009, 182, 161–171. [Google Scholar] [CrossRef] [Green Version]
- Kaur, G.; Asha, W.; Abdin, M.Z.; Sarwat, M.; Ahmad, A.I. Molecular network of nitrogen and sulphur signaling in plants. In Stress Signaling in Plants: Genomics and Proteomics Perspective, 1st ed.; Sarwat, M., Ahmad, A., Ahmad, M.Z., Eds.; Springer: New York, NY, USA, 2013; p. 240. [Google Scholar]
- Sysoeva, M.I.; Markovskaya, E.F.; Shibaeva, T.G. Plants under continuous light: A review. Plant Stress 2010, 4, 5–17. [Google Scholar]
- O’Connor, D.J.; Wright, G.C.; Dieters, M.J.; George, D.L.; Hunter, M.N.; Tatnell, J.R.; Fleischfresser, D.B. Development and application of speed breeding technologies in a commercial peanut breeding program. Peanut Sci. 2013, 40, 107–114. [Google Scholar] [CrossRef]
- Velez-Ramirez, A.I.; van Ieperen, W.; Vreugdenhil, D.; Millenaar, F.F. Plants under continuous light. Trends Plant Sci. 2011, 16, 310–318. [Google Scholar] [CrossRef] [Green Version]
- Bian, Z.H.; Yang, Q.C.; Li, T.; Cheng, R.F.; Barnett, Y.; Lu, C.G. Study of the beneficial effects of green light on lettuce grown under short-term continuous red and blue light-emitting diodes. Physiol. Plant. 2018, 164, 226–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, Z.H.; Zhang, X.Y.; Wang, Y.; Lu, C.G. Improving drought tolerance by altering the photosynthetic rate and stomatal aperture via green light in tomato (Solanum lycopersicum L.) seedlings under drought conditions. Environ. Exp. Bot. 2019, 167, 103844. [Google Scholar] [CrossRef]
- Wargent, J.J.; Elfadly, E.M.; Moore, J.P.; Paul, N.D. Increased exposure to UV-B radiation during early development leads to enhanced photoprotection and improved long-term performance in Lactuca sativa. Plant Cell Environ. 2011, 34, 1401–1413. [Google Scholar] [CrossRef] [PubMed]
- Schreiner, M.; Martínez-Abaigar, J.; Glaab, J.; Jansen, M. UV-B induced secondary plant metabolites: Potential benefits for plant and human health. Opt. Photonik 2014, 9, 34–37. [Google Scholar] [CrossRef]
- Xu, J.; Gao, K. UV-A enhanced growth and UV-B induced positive effects in the recovery of photochemical yield in Gracilaria lemaneiformis (Rhodophyta). J. Photochem. Photobiol. B Biol. 2010, 100, 117–122. [Google Scholar] [CrossRef] [Green Version]
- Xu, K.; Qiu, B.S. Responses of superhigh-yield hybrid rice Liangyoupeijiu to enhancement of ultraviolet-B radiation. Plant Sci. 2007, 172, 139–149. [Google Scholar] [CrossRef]
- Jansen, M.A.; Gaba, V.; Greenberg, B.M. Higher plants and UV-B radiation: Balancing damage, repair and acclimation. Trends Plant Sci. 1998, 3, 131–135. [Google Scholar] [CrossRef]
- Kataria, S.; Jajoo, A.; Guruprasad, K.N. Impact of increasing Ultraviolet-B (UV-B) radiation on photosynthetic processes. J. Photochem. Photobiol. B Biol. 2014, 137, 55–66. [Google Scholar] [CrossRef]
- Krause, G.H.; Gallé, A.; Gademann, R.; Winter, K. Capacity of protection against ultraviolet radiation in sun and shade leaves of tropical forest plants. Funct. Plant Biol. 2003, 30, 533–542. [Google Scholar] [CrossRef]
- Takahashi, S.; Milward, S.E.; Yamori, W.; Evans, J.R.; Hillier, W.; Badger, M.R. The solar action spectrum of photosystem II damage. Plant Physiol. 2010, 153, 988–993. [Google Scholar] [CrossRef] [Green Version]
- Bashri, G.; Singh, M.; Mishra, R.K.; Kumar, J.; Singh, V.P.; Prasad, S.M. Kinetin regulates UV-B-induced damage to growth, photosystem II photochemistry, and nitrogen metabolism in tomato seedlings. J. Plant Growth Regul. 2018, 37, 233–245. [Google Scholar] [CrossRef]
- Huang, G.; Wang, L.; Zhou, Q. Lanthanum (III) regulates the nitrogen assimilation in soybean seedlings under ultraviolet-B radiation. Bio. Trace Elem. Res. 2013, 151, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Schwalbe, M.; Teller, S.; Oelmüller, R.; Appenroth, K.J. Influence of UVB irradiation on nitrate and ammonium assimilating enzymes in Spirodela polyrhiza. Aquat. Bot. 1999, 64, 19–34. [Google Scholar] [CrossRef]
- Ulm, R.; Baumann, A.; Oravecz, A.; Máté, Z.; Ádám, É.; Oakeley, E.J.; Schäfer, E.; Nagy, F. Genome-wide analysis of gene expression reveals function of the bZIP transcription factor HY5 in the UV-B response of Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 1397–1402. [Google Scholar] [CrossRef] [Green Version]
- Cloix, C.; Kaiserli, E.; Heilmann, M.; Baxter, K.J.; Brown, B.A.; O’Hara, A.; Smith, B.O.; Christie, J.M.; Jenkins, G.I. C-terminal region of the UV-B photoreceptor UVR8 initiates signaling through interaction with the COP1 protein. Proc. Natl. Acad. Sci. USA 2012, 109, 16366–16370. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.Y.; Cheng, Y.H.; Chen, K.E.; Tsay, Y.F. Nitrate transport, signaling, and use efficiency. Annu. Rev. Plant Biol. 2018, 69, 85–122. [Google Scholar] [CrossRef]
- He, J.; Austin, P.T.; Lee, S.K. Effects of elevated root zone CO2 and air temperature on photosynthetic gas exchange, nitrate uptake, and total reduced nitrogen concentration in aeroponically grown lettuce plants. J. Exp. Bot. 2010, 61, 3959–3969. [Google Scholar] [CrossRef] [Green Version]
- Peet, M.; Sato, S.; Clément, C.; Pressman, E. Heat stress increases sensitivity of pollen, fruit and seed production in tomatoes (Lycopersicon esculentum Mill.) to non-optimal vapor pressure deficits. Acta Hortic. 2003, 618, 209–215. [Google Scholar] [CrossRef]
- Toivonen, P.M. Postharvest physiology of vegetables. In Handbook of Vegetables and Vegetable Processing; Sinha, N., Hui, Y.H., Evranuz, E.Ö., Siddiq, M., Ahmed, J., Eds.; Wiley-Blackwell Publishing: Oxford, UK, 2010; pp. 199–220. ISBN 978-0-8138-1541-1. [Google Scholar]
- Klimenko, S.; Peshkova, A.; Dorofeev, N. Nitrate reductase activity during heat shock in winter wheat. J. Stress Physiol. Biochem. 2006, 2, 50–55. [Google Scholar]
- Shu, S.; Tang, Y.; Yuan, Y.; Sun, J.; Zhong, M.; Guo, S. The role of 24-epibrassinolide in the regulation of photosynthetic characteristics and nitrogen metabolism of tomato seedlings under a combined low temperature and weak light stress. Plant Physiol. Biochem. 2016, 107, 344–353. [Google Scholar] [CrossRef]
- Li, W.; Wang, Y.; Okamoto, M.; Crawford, N.M.; Siddiqi, M.Y.; Glass, A.D. Dissection of the AtNRT2.1:AtNRT2.2 inducible high-affinity nitrate transporter gene cluster. Plant Physiol. 2007, 143, 425–433. [Google Scholar] [CrossRef] [Green Version]
- Orsel, M.; Chopin, F.; Leleu, O.; Smith, S.J.; Krapp, A.; Daniel-Vedele, F.; Miller, A.J. Characterization of a two-component high-affinity nitrate uptake system in Arabidopsis. Physiology and protein-protein interaction. Plant Physiol. 2006, 142, 1304–1317. [Google Scholar] [CrossRef] [Green Version]
- Goel, P.; Singh, A.K. Abiotic stresses downregulate key genes involved in nitrogen uptake and assimilation in Brassica juncea L. PLoS ONE 2015, 10, e0143645. [Google Scholar] [CrossRef]
- Catalá, R.; Medina, J.; Salinas, J. Integration of low temperature and light signaling during cold acclimation response in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 16475–16480. [Google Scholar] [CrossRef] [Green Version]
- Karayekov, E.; Sellaro, R.; Legris, M.; Yanovsky, M.J.; Casal, J.J. Heat shock–induced fluctuations in clock and light signaling enhance phytochrome B–mediated arabidopsis deetiolation. Plant Cell 2013, 25, 2892–2906. [Google Scholar] [CrossRef] [Green Version]
- Bloom, A.J.; Asensio, J.S.R.; Randall, L.; Rachmilevitch, S.; Cousins, A.B.; Carlisle, E.A. CO2 enrichment inhibits shoot nitrate assimilation in C3 but not C4 plants and slows growth under nitrate in C3 plants. Ecology 2012, 93, 355–367. [Google Scholar] [CrossRef]
- Natali, S.M.; Sañudo-Wilhelmy, S.A.; Lerdau, M.T. Effects of elevated carbon dioxide and nitrogen fertilization on nitrate reductase activity in sweetgum and loblolly pine trees in two temperate forests. Plant Soil 2009, 314, 197–210. [Google Scholar] [CrossRef]
- White, A.C.; Rogers, A.; Rees, M.; Osborne, C.P. How can we make plants grow faster? A source–sink perspective on growth rate. J. Exp. Bot. 2016, 67, 31–45. [Google Scholar] [CrossRef]
- Dong, J.; Gruda, N.; Lam, S.K.; Li, X.; Duan, Z. Effects of elevated CO2 on nutritional quality of vegetables: A Review. Front. Plant Sci. 2018, 9, 9. [Google Scholar] [CrossRef]
- Piñero, M.C.; Pérez-Jiménez, M.; López-Marín, J.; del Amor, F.M. Changes in the salinity tolerance of sweet pepper plants as affected by nitrogen form and high CO2 concentration. J. Plant Physiol. 2016, 200, 18–27. [Google Scholar] [CrossRef]
- Robredo, A.; Pérez-López, U.; Miranda-Apodaca, J.; Lacuesta, M.; Mena-Petite, A.; Muñoz-Rueda, A. Elevated CO2 reduces the drought effect on nitrogen metabolism in barley plants during drought and subsequent recovery. Environ. Exp. Bot. 2011, 71, 399–408. [Google Scholar] [CrossRef]
- Larios, B.; Agüera, E.; de la Haba, P.; Pérez-Vicente, R.; Maldonado, J.M. A short-term exposure of cucumber plants to rising atmospheric CO2 increases leaf carbohydrate concentration and enhances nitrate reductase expression and activity. Planta 2001, 212, 305–312. [Google Scholar] [CrossRef]
- Stitt, M.; Krapp, A. The interaction between elevated carbon dioxide and nitrogen nutrition: The physiological and molecular background. Plant Cell Environ. 1999, 22, 583–621. [Google Scholar] [CrossRef]
- Bloom, A.J.; Burger, M.; Kimball, B.A.; Pinter, P.J., Jr. Nitrate assimilation is inhibited by elevated CO2 in field-grown wheat. Nat. Clim. Chang. 2014, 4, 477. [Google Scholar] [CrossRef]
- Stitt, M. Limitation of photosynthesis by carbon metabolism: I. Evidence for excess electron transport capacity in leaves carrying out photosynthesis in saturating light and CO2. Plant Physiol. 1986, 81, 1115–1122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloom, A.J.; Smart, D.R.; Nguyen, D.T.; Searles, P.S. Nitrogen assimilation and growth of wheat under elevated carbon dioxide. Proc. Natl. Acad. Sci. USA 2002, 99, 1730–1735. [Google Scholar] [CrossRef] [Green Version]
- Matallana González, M.; Martínez-Tomé, M.; Torija Isasa, M. Nitrate and nitrite concentration in organically cultivated vegetables. Food Addit. Contam. Part B 2010, 3, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Lal, R. Feeding 11 billion on 0.5 billion hectare of area under cereal crops. Food Energy Secur. 2016, 5, 239–251. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.K.; Yang, Q.C. Effects of short-term treatment with various light intensities and hydroponic solutions on nitrate concentration of lettuce. Acta Agric. Scand. Sect. B -Soil Plant Sci. 2012, 62, 109–113. [Google Scholar] [CrossRef]
- Liu, W.K.; Yang, Q.C.; Du, L.F. Short-term treatment with hydroponic solutions containing osmotic ions and ammonium molybdate decreased nitrate concentration in lettuce. Acta Agric. Scand. Sect. B -Soil Plant Sci. 2011, 61, 573–576. [Google Scholar] [CrossRef]
- Wang, B.; Shen, Q. NH4+-N/NO3−-N ratios on growth and NO3−-N remobilization in root vacuoles and cytoplasm of lettuce genotypes. Can. J. Plant Sci. 2011, 91, 411–417. [Google Scholar] [CrossRef]
- Lin, X.Y.; Liu, X.X.; Zhang, Y.P.; Zhou, Y.Q.; Hu, Y.; Chen, Q.H.; Zhang, Y.S.; Jin, C.W. Short-term alteration of nitrogen supply prior to harvest affects quality in hydroponic-cultivated spinach (Spinacia oleracea). J. Sci. Food Agric. 2014, 94, 1020–1025. [Google Scholar] [CrossRef] [PubMed]
- Urlić, B.; Dumičić, G.; Romić, M.; Ban, S.G. The effect of N and NaCl on growth, yield, and nitrate concentration of salad rocket (Eruca sativa Mill.). J. Plant Nutr. 2017, 40, 2611–2618. [Google Scholar] [CrossRef]
- Henriques, A.D.P.; Marcelis, L. Regulation of growth at steady-state nitrogen nutrition in lettuce (Lactuca sativa L.): Interactive effects of nitrogen and irradiance. Ann. Bot. 2000, 86, 1073–1080. [Google Scholar] [CrossRef]
- Lei, B.; Bian, Z.H.; Yang, Q.C.; Wang, J.; Cheng, R.F.; Li, K.; Liu, W.K.; Zhang, Y.; Fang, H.; Tong, Y.X. The positive function of selenium supplementation on reducing nitrate accumulation in hydroponic lettuce (Lactuca sativa L.). J. Integr. Agric. 2018, 17, 837–846. [Google Scholar] [CrossRef] [Green Version]
- Wojciechowska, R.; Kowalska, I. The effect of foliar application of urea, Mo and BA on nitrate metabolism in lettuce leaves in the spring and summer-autumn seasons. Folia Hortic. 2011, 23, 119–123. [Google Scholar] [CrossRef] [Green Version]
- Moncada, A.; Miceli, A.; Sabatino, L.; Iapichino, G.; D’Anna, F.; Vetrano, F. Effect of molybdenum rate on yield and quality of lettuce, escarole, and curly endive grown in a floating system. Agronomy 2018, 8, 171. [Google Scholar] [CrossRef] [Green Version]
- Kovács, B.; Puskás-Preszner, A.; Huzsvai, L.; Lévaib, L.; Bódi, É. Effect of molybdenum treatment on molybdenum concentration and nitrate reduction in maize seedlings. Plant Physiol. Biochem. 2015, 96, 38–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sabatino, L.; D’Anna, F.; D’Anna, F.; Iapichino, G.; Moncada, A.; D’Anna, E.; De Pasquale, C. Interactive.effects of genotype and molybdenum supply on yield and overall fruit quality of tomato. Front. Plant Sci. 2019, 9, 1922. [Google Scholar] [CrossRef] [Green Version]
- Borlotti, A.; Vigani, G.; Zocchi, G. Iron deficiency affects nitrogen metabolism in cucumber (Cucumis sativus L.) plants. BMC Plant Biol. 2012, 12, 189. [Google Scholar] [CrossRef] [Green Version]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Vernieri, P.; Ferrante, A. Biostimulants and crop responses: A review. Biol. Agric. Hortic. 2015, 31, 1–17. [Google Scholar] [CrossRef]
- Colla, G.; Rouphael, Y. Biostimulants in horticulture. Sci. Hortic. 2015, 30, 1–2. [Google Scholar] [CrossRef]
- Amanda, A.; Ferrante, A.; Valagussa, M.; Piaggesi, A. Effect of biostimulants on quality of baby leaf lettuce grown under plastic tunnel. Acta Hortic. 2019, 807, 407–412. [Google Scholar] [CrossRef]
- Kunicki, E.; Grabowska, A.; Sekara, A.; Wojciechowska, R. The effect of cultivar type, time of cultivation, and biostimulant treatment on the yield of spinach (Spinacia oleracea L.). Folia Hortic. 2010, 22, 9–13. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.Q.; Ko, K.Y.; Kim, S.H.; Lee, K.S. Effect of amino acid fertilization on nitrate assimilation of leafy radish and soil chemical properties in high nitratesoil. Commun. Soil Sci. Plant Anal. 2008, 39, 269–281. [Google Scholar] [CrossRef]
- Liu, X.Q.; Lee, K.S. Effect of mixed amino acids on crop growth. In Agricultural Science; Aflakpui, G., Ed.; Tech Europe Publisher: Rijeka, Croatia, 2012; pp. 119–158. [Google Scholar]
- Raigon, M. Comparación de parámetros de calidad en hortalizas de hoja ancha bajo sistemas de producción ecológica y convencional. Agrícola vergel Frutic. Hortic. Floric. 2002, 241, 26–32. [Google Scholar]
- Ma, B.L.; Biswas, D.K. Precision nitrogen management for sustainable corn production. In Sustainable Agriculture Reviews; Lichtfouse, E., Goyal, A., Eds.; Springer: Dordrecht, The Netherlands, 2015; pp. 33–62. [Google Scholar]
- Rogovska, N.; Laird, D.A.; Chiou, C.P.; Bond, L.J. Development of field mobile soil nitrate sensor technology to facilitate precision fertilizer management. Precis. Agric. 2019, 20, 40–55. [Google Scholar] [CrossRef]
- Schiattone, M.I.; Viggiani, R.; Di Venere, D.; Sergio, L.; Cantore, V.; Todorovic, M.; Perniola, M.; Candido, V. Impact of irrigation regime and nitrogen rate on yield, quality and water use efficiency of wild rocket under greenhouse conditions. Sci. Hortic. 2018, 229, 182–192. [Google Scholar] [CrossRef]
- Koyama, R.; Itoh, H.; Kimura, S.; Mirioka, A.; Uno, Y. Augmentation of antioxidant constituents by drought stress to roots in leafy vegetables. Hort Technol. 2012, 22, 121–125. [Google Scholar] [CrossRef] [Green Version]
- Lv, M.C.; Cai, H.J.; Chen, X.M. Effects of sewage irrigation on physiological characteristics of potato and soil environment. J. Irrig. Drain. 2007, 26, 26–29. [Google Scholar]
- Al-Lahham, O.; El Assi, N.M.; Fayyad, M. Impact of treated wastewater irrigation on quality attributes and contamination of tomato fruit. Agric. Water Manag. 2003, 61, 51–62. [Google Scholar] [CrossRef]
- Yang, B.; Kong, X.; Cui, B.J.; Jin, D.C.; Deng, Y.; Zhuang, X.L.; Zhuang, G.Q.; Bai, Z.H. Impact of rural domestic wastewater irrigation on the physicochemical and microbiological properties of pakchoi and soil. Water 2015, 7, 1825–1839. [Google Scholar] [CrossRef] [Green Version]
- Di Mola, I.; Rouphael, Y.; Colla, G.; Fagnano, M.; Paradiso, R.; Mori, M. Morphophysiological traits and nitrate concentration of greenhouse lettuce as affected by irrigation with saline water. HortScience 2017, 52, 1716–1721. [Google Scholar] [CrossRef] [Green Version]
- Borgognone, D.Y.; Rouphael, M.; Cardarelli, L.; Lucini, L.l.; Colla, G. Changes in biomass, mineral composition, and quality of cardoon in response to NO3–:Cl– ratio and nitrate deprivation from the nutrient solution. Front. Plant Sci. 2016, 7, 978. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Byrt, C.S.; Qiu, J.; Baumann, U.; Hrmova, M.; Evrard, A.; Johnson, A.A.; Birnbaum, K.D.; Mayo, G.M.; Jha, D. Identification of a stelar-localized transport protein that facilitates root-to-shoot transfer of chloride in Arabidopsis. Plant Physiol. 2016, 170, 1014–1029. [Google Scholar] [CrossRef]
- Li, B.; Qiu, J.; Jayakannan, M.; Xu, B.; Li, Y.; Mayo, G.M.; Tester, M.; Gilliham, M.; Roy, S.J. AtNPF2.5 modulates chloride (Cl−) efflux from roots of Arabidopsis thaliana. Front. Plant Sci. 2017, 7, 2013. [Google Scholar] [CrossRef] [Green Version]
- Bian, Z.H.; Bo, B.; Cheng, R.F.; Wang, Y.; Li, T.; Yang, Q.C. Selenium distribution and nitrate metabolism in hydroponic lettuce (Lactuca sativa L.): Effects of selenium forms and light spectra. J. Integr. Agric. 2020, 19, 133–144. [Google Scholar] [CrossRef]
- Ma, C.; Zhang, H.H.; Wang, X. Machine learning for big data analytics in plants. Trends Plant Sci. 2014, 19, 798–808. [Google Scholar] [CrossRef] [PubMed]
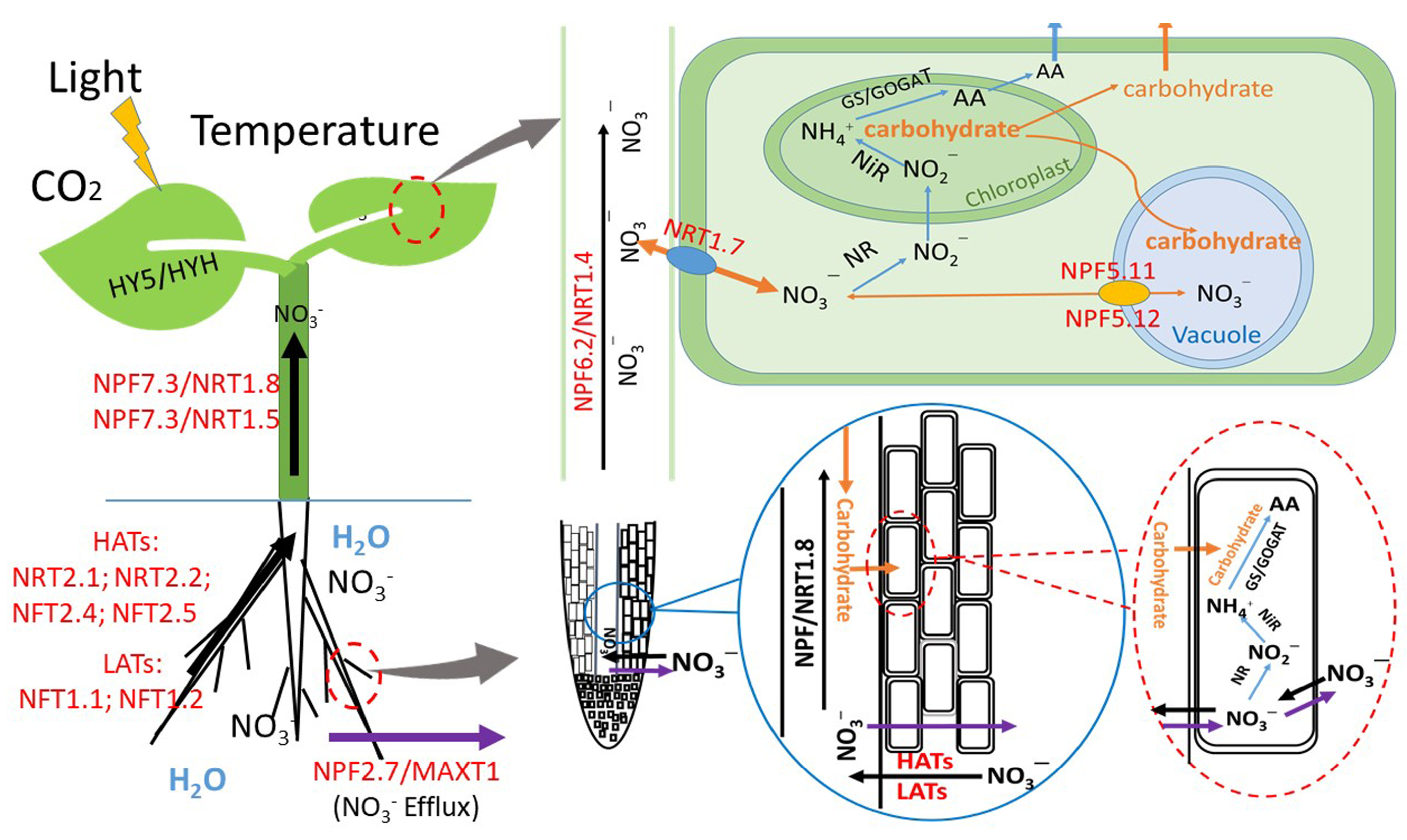
 ” represents NO3– efflux; “
” represents NO3– efflux; “  ” represents NO3– influx and transport in plant roots and stems. “
” represents NO3– influx and transport in plant roots and stems. “  ” indicates NO3– metabolism. “
” indicates NO3– metabolism. “  ” indicates carbohydrate transfer; “
” indicates carbohydrate transfer; “  ” represents NO3– transfer among plant cells.
” represents NO3– efflux; “ ” represents NO3– influx and transport in plant roots and stems. “ ” indicates NO3– metabolism. “ ” indicates carbohydrate transfer; “ ” represents NO3– transfer among plant cells.
” represents NO3– transfer among plant cells.
” represents NO3– efflux; “ ” represents NO3– influx and transport in plant roots and stems. “ ” indicates NO3– metabolism. “ ” indicates carbohydrate transfer; “ ” represents NO3– transfer among plant cells.
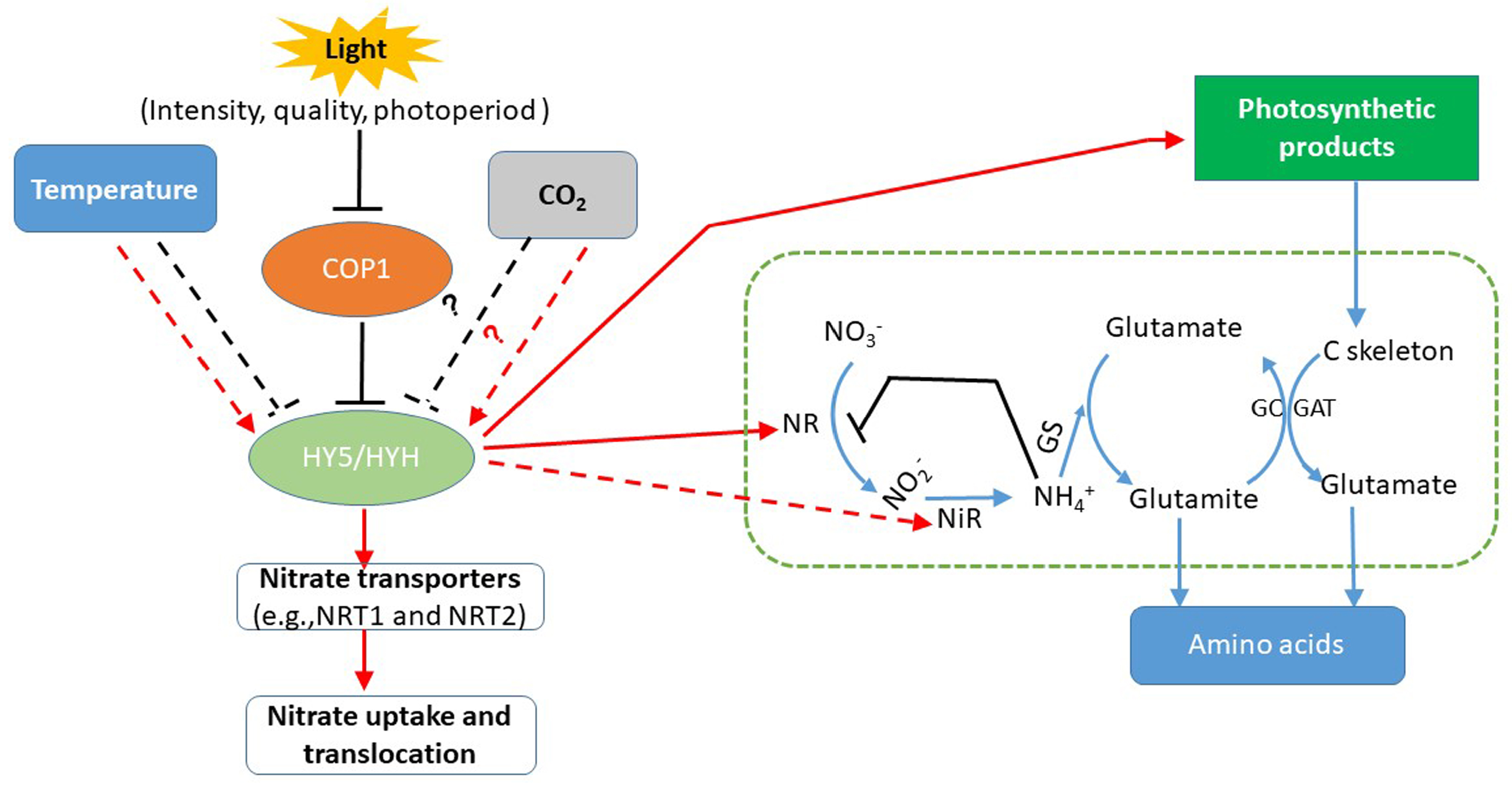
 ” represents nitrate assimilation under the reduction of nitrogen-metabolizing enzymes. “
” represents nitrate assimilation under the reduction of nitrogen-metabolizing enzymes. “  and
and  ” indicates defined and presumed positive regulation. “
” indicates defined and presumed positive regulation. “  and
and  ” represents defined and presumed inhibition, whereas “?” indicates a speculation.
” represents nitrate assimilation under the reduction of nitrogen-metabolizing enzymes. “ and ” indicates defined and presumed positive regulation. “ and ” represents defined and presumed inhibition, whereas “?” indicates a speculation.
” represents defined and presumed inhibition, whereas “?” indicates a speculation.
” represents nitrate assimilation under the reduction of nitrogen-metabolizing enzymes. “ and ” indicates defined and presumed positive regulation. “ and ” represents defined and presumed inhibition, whereas “?” indicates a speculation.

{kind=link}
{kind=link}
| Lighting Condition | Vegetable Species or Varieties | Nitrate Metabolic Effects | Reference | |||
|---|---|---|---|---|---|---|
| Light Spectra | Light Intensity (μmol m–2 s–1) | Light Duration | ||||
| UV—A light (315–402 nm) | UV-A (383–426 nm) + red (599–644 nm) and blue (435–489 nm) light | UV-A LEDs: 30.3 Red LEDs: 111 Blue LEDs: 158.7 | 18 h light/6 h dark | Lettuce (var. crispa) | Nitrate decreased by 300 mg kg–1 | [60] |
| UV-A (366, 390, and 402 nm), + combination of red (638 and 665 nm), blue (447 nm) and far red (731 nm) light | UV-A LEDs: 6.2 and 12.4, Blue LEDs: 21 Red LEDs: 122 Deep red LEDs: 155 Far red LEDs: 2.2 | 16 h light/8 h dark | Microgreens of basil (Ocimum basilicum L., var. Sweet Genovese), beet (Beta vulgaris L., var. Bulls Blood) and red pak choi (var. chinensis, Rubi) | Basil exposed to 6.2 µmol m–2 s–1 UV-A (366 nm): decreased by around 150 mg kg–1; Basil exposed to 12.4 µmol m–2 s–1 UV-A (366, 390, and 402 nm): increased by 50–100 mg kg–1; Beet exposed to 6.2 µmol m–2 s–1 UV-A (366, 390, and 402 nm): increased by 50–100 mg kg–1; Pal choi exposed to 6.2 µmol m–2 s–1 UV-A (366, 390, and 402 nm): increased by around 50 mg kg–1 | [61] | |
| Blue LED light (400–492 nm) | Blue light (450 nm), + white (400–700 nm) light | Blue LEDs: 30 White LEDs: 170 | 16 h light/8 h dark | Lettuce (var. crispa ‘Green Oak Leaf’) | Nitrate in leaves decreased by about 200 mg kg–1 | [62] |
| Green LED light (492–577 nm) | Green light (505 or 530 nm) + high pressure sodium lamp (HPL) light | Green LEDs: 30 HPL: 170 | 16 h light/8 h dark | Baby leaf lettuce: red leaf (var. Multired 4), green leaf (var. Multigreen 3), light green leaf (var. Multiblond 2) | Nitrate in red leaf and light green leaf lettuce decreased by 5666 and 4705 mg kg–1 under 505 nm LED, 5452 and 9785 mg kg–1 under 530 nm LED light; nitrate increased by 3568 and 249 mg kg–1 under 505 and 530 nm LED | [63] |
| Green light (522 nm) + white (400–700 nm) light | Green LEDs: 30 White LED: 103 | 16 h light/8 h dark | Lettuce (var. crispa ‘Green Oak Leaf’) | Nitrate decreased by around 200 mg kg–1 | [62] | |
| Green light (530 nm) + combined red (660 nm) and blue (450 nm) light | Green LEDs: 33.33 Blue LEDs: 33.33 Red LEDs: 133.34 | Illumination for 24 h before harvest | Lettuce (var. Butterhead) | Upregulated NR related gene expression; increased NR activity; decreased nitrate by 80–111 mg kg–1 | [6,14] | |
| Green light (494–565 nm) + combined red (599–644 nm) and Blue (435–489 nm) light | Green LEDs: 44.7 Blue LEDs: 108.3 Red LEDs: 147 | 18 h light/6 h dark | Lettuce (var. crispa) | Nitrate decreased by 400 mg kg–1 | [60] | |
| White LED (400–700 nm) | White light (400–700 nm) + combination of red (660 nm) and blue (454 nm) light | Total light intensity: 210 | 16 h light/8 h dark | Boston lettuce (var. capitate) | Nitrate decreased by 20 mg kg–1 (dry weight) | [16] |
| Red LED (600–700 nm) | Red light (638 nm) + HLP and nature light | Red LEDs + HPL (90) + Nature light intensity = 300 | Illumination for 3 days before harvest | Spinach, parsley (Petroselinum crispum Mill.), dill (Anethumgraveolens) | Reduced nitrate in spinach, parsley, and dill decreased by 206, 566, and 1811 mg kg–1, respectively | [64] |
| Yellow light (577–597 nm) | Yellow light (596 nm) + white light (400–700 nm) light | Yellow LEDs: 30 White LEDs: 103 | 16 h light/8 h dark | Lettuce (var. crispa ‘Green Oak Leaf) | Nitrate in leaves increased by around 200 mg kg–1 | [62] |
| Far-red (735nm and infrared light (850 nm) | Far-red light (735 nm) + cool fluorescent lamp | Far-red LEDs: 86 Cool fluorescent lamp: 187 | Illumination for 5 days before harvest | Spinach (var ‘Whitney’). | Nitrate excessive accumulation (0.12 mmoles g–1) by 10 times compared with cool fluorescent lamp only (0.01 mmoles g–1) | [65] |
| Infrared light (850 nm) + White light (400–700 nm) | Infar-red LEDs: 30 White LEDs: 103 | 16 h light/8 h dark | Lettuce (var.crispa ‘Green Oak Leaf) | Nitrate in leaves increased by about 160 mg kg–1 | [62] | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bian, Z.; Wang, Y.; Zhang, X.; Li, T.; Grundy, S.; Yang, Q.; Cheng, R. A Review of Environment Effects on Nitrate Accumulation in Leafy Vegetables Grown in Controlled Environments. Foods 2020, 9, 732. https://doi.org/10.3390/foods9060732
Bian Z, Wang Y, Zhang X, Li T, Grundy S, Yang Q, Cheng R. A Review of Environment Effects on Nitrate Accumulation in Leafy Vegetables Grown in Controlled Environments. Foods. 2020; 9(6):732. https://doi.org/10.3390/foods9060732
Chicago/Turabian StyleBian, Zhonghua, Yu Wang, Xiaoyan Zhang, Tao Li, Steven Grundy, Qichang Yang, and Ruifeng Cheng. 2020. "A Review of Environment Effects on Nitrate Accumulation in Leafy Vegetables Grown in Controlled Environments" Foods 9, no. 6: 732. https://doi.org/10.3390/foods9060732