Regulatory Effect of Lactobacillus brevis Bmb6 on Gut Barrier Functions in Experimental Colitis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Acid and Bile Acid Tolerance Assay
2.2. Preparation of Bacterial Cell Lysate
2.3. Determination of Antioxidant Activity
2.4. Determination of β-Galactosidase Activity
2.5. 16S rDNA Sequencing and Identification
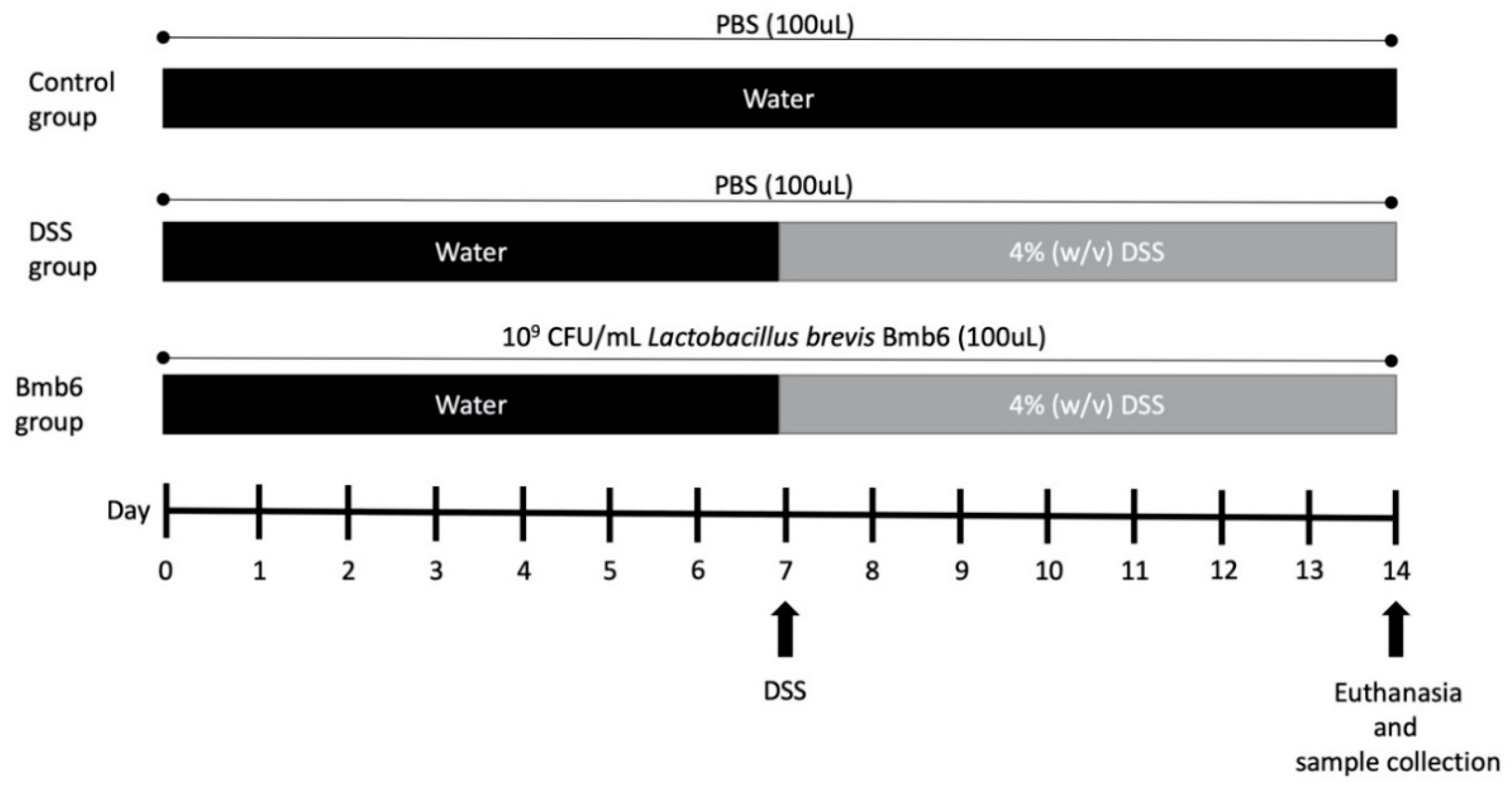
2.6. Induction and Assessment of DSS-Induced Colitis
2.7. Histological Assessment
2.8. RNA Extraction and Gene Expression Analysis
2.9. Western Blotting
2.10. Statistical Analysis
3. Results
3.1. In Vitro Characteristic of L. brevis Bmb6
3.2. Antioxidant Activity
3.3. β-Galactosidase Activity
3.4. 16S Identification
3.5. Effects of L. brevis Bmb6 on DSS-Induced Colitis Mice
3.6. Histological Analysis of Colon Sections
3.7. Effects of L. brevis Bmb6 on TJ Proteins
3.8. Effects of L. brevis on Colonic Inflammatory Cytokine Expression
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kamm, M.A. Rapid changes in epidemiology of inflammatory bowel disease. Lancet 2018, 390, 2741–2742. [Google Scholar] [CrossRef]
- Khor, B.; Gardet, A.; Xavier, R.J. Genetics and pathogenesis of inflammatory bowel disease. Nature 2011, 474, 307–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, V.; Olsen, A.; Carbonnel, F.; Tjønneland, A.; Vogel, U. Diet and risk of inflammatory bowel disease. Dig. Liver Dis. 2012, 44, 185–194. [Google Scholar] [CrossRef]
- Mukhopadhya, I.; Hansen, R.; El-Omar, E.M.; Hold, G.L. IBD-what role do proteobacteria play? Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 219. [Google Scholar] [CrossRef] [PubMed]
- Pithadia, A.B.; Jain, S. Treatment of inflammatory bowel disease (IBD). Pharmacol. Rep. 2011, 63, 629–642. [Google Scholar] [CrossRef]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.J.M.; Duckworth, C.A.; Guan, Y.; Montrose, M.H. Mechanisms of epithelial cell shedding in the mammalian intestine and maintenance of barrier function. Ann. N. Y. Acad. Sci. 2009, 1165, 135. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.A.; Uppada, S.; Achkar, I.W.; Hashem, S.; Yadav, S.K.; Shanmugakonar, M.; Al-Naemi, H.A.; Haris, M.; Uddin, S. Tight junction proteins and signaling pathways in cancer and inflammation: A functional crosstalk. Front. Physiol. 2019, 9, 1942. [Google Scholar] [CrossRef] [Green Version]
- Fujibe, M.; Chiba, H.; Kojima, T.; Soma, T.; Wada, T.; Yamashita, T.; Sawada, N. Thr203 of claudin-1, a putative phosphorylation site for MAP kinase, is required to promote the barrier function of tight junctions. Exp. Cell Res. 2004, 295, 36–47. [Google Scholar] [CrossRef]
- Umeda, K.; Matsui, T.; Nakayama, M.; Furuse, K.; Sasaki, H.; Furuse, M.; Tsukita, S. Establishment and characterization of cultured epithelial cells lacking expression of ZO-1. J. Biol. Chem. 2004, 279, 44785–44794. [Google Scholar] [CrossRef] [Green Version]
- Heller, F.; Florian, P.; Bojarski, C.; Richter, J.; Christ, M.; Hillenbrand, B.; Mankertz, J.; Gitter, A.H.; Bürgel, N.; Fromm, M. Interleukin-13 is the key effector Th2 cytokine in ulcerative colitis that affects epithelial tight junctions, apoptosis, and cell restitution. Gastroenterology 2005, 129, 550–564. [Google Scholar] [CrossRef] [PubMed]
- Runkle, E.A.; Mu, D. Tight junction proteins: From barrier to tumorigenesis. Cancer Lett. 2013, 337, 41–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruewer, M.; Luegering, A.; Kucharzik, T.; Parkos, C.A.; Madara, J.L.; Hopkins, A.M.; Nusrat, A. Proinflammatory cytokines disrupt epithelial barrier function by apoptosis-independent mechanisms. J. Immunol. 2003, 171, 6164–6172. [Google Scholar] [CrossRef] [Green Version]
- Gaón, D.; Doweck, Y.; Gómez Zavaglia, A.; Ruiz Holgado, A.; Oliver, G. Lactose digestion by milk fermented with Lactobacillus acidophilus and Lactobacillus casei of human origin. Medicina 1995, 55, 237–242. [Google Scholar]
- Tuohy, K.M.; Probert, H.M.; Smejkal, C.W.; Gibson, G.R. Using probiotics and prebiotics to improve gut health. Drug Discov. Today 2003, 8, 692–700. [Google Scholar] [CrossRef]
- Gawrońska, A.; Dziechciarz, P.; Horvath, A.; Szajewska, H. A randomized double-blind placebo-controlled trial of Lactobacillus GG for abdominal pain disorders in children. Aliment. Pharmacol. Ther. 2007, 25, 177–184. [Google Scholar] [CrossRef]
- Miele, E.; Pascarella, F.; Giannetti, E.; Quaglietta, L.; Baldassano, R.N.; Staiano, A. Effect of a probiotic preparation (VSL#3) on induction and maintenance of remission in children with ulcerative colitis. Am. J. Gastroenterol. 2009, 104, 437–443. [Google Scholar]
- Liu, Y.W.; Ong, W.K.; Su, Y.W.; Hsu, C.C.; Cheng, T.H.; Tsai, Y.C. Anti-inflammatory effects of Lactobacillus brevis K65 on RAW 264.7 cells and in mice with dextran sulphate sodium-induced ulcerative colitis. Benef. Microbes 2016, 7, 387–396. [Google Scholar] [CrossRef]
- Park, J.S.; Joe, I.; Rhee, P.D.; Jeong, C.S.; Jeong, G. A lactic acid bacterium isolated from kimchi ameliorates intestinal inflammation in DSS-induced colitis. J. Microbiol. 2017, 55, 304–310. [Google Scholar] [CrossRef]
- Lee, B.; Yong, C.-C.; Yi, H.-C.; Kim, S.; Oh, S. A non-yeast kefir-like fermented milk development with Lactobacillus acidophilus KCNU and Lactobacillus brevis Bmb6. Food Sci. Anim. Resour. 2020. [Google Scholar] [CrossRef]
- Facino, R.M.; Carini, M.; Aldini, G.; Berti, F.; Rossoni, G. Panax ginseng administration in the rat prevents myocardial ischemia-reperfusion damage induced by hyperbaric oxygen: Evidence for an antioxidant intervention. Planta Med. 1999, 65, 614–619. [Google Scholar] [CrossRef]
- Dhindsa, R.S.; Plumb-dhindsa, P.; Thorpe, T.A. Leaf senescence: Correlated with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J. Exp. Bot. 1981, 32, 93–101. [Google Scholar] [CrossRef]
- Miller, J. Assay for β-galactosidase. In Experiments in Molecular Genetics; Miller, J.M., Ed.; Cold Spring Harbor Laboratory: Cold Spring Harbor, NY, USA, 1972; pp. 352–355. [Google Scholar]
- Murthy, S.N.S.; Cooper, H.S.; Shim, H.; Shah, R.S.; Ibrahim, S.A.; Sedergran, D.J. Treatment of dextran sulfate sodium-induced murine colitis by intracolonic cyclosporin. Dig. Dis. Sci. 1993, 38, 1722–1734. [Google Scholar] [CrossRef]
- Song, S.; Lee, S.-J.; Park, D.-J.; Oh, S.; Lim, K.-T. The anti-allergic activity of Lactobacillus plantarum L67 and its application to yogurt. J. Dairy Sci. 2016, 99, 9372–9382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.H.; Cho, S.S.; Simkhada, J.R.; Rahman, M.S.; Choi, Y.S.; Kim, C.S.; Yoo, J.C. A novel multifunctional peptide oligomer of bacitracin with possible bioindustrial and therapeutic applications from a Korean food-source Bacillus strain. PLoS ONE 2017, 12, e0176971. [Google Scholar] [CrossRef]
- Kim, Y.; Koh, J.H.; Ahn, Y.J.; Oh, S.; Kim, S.H. The synergic anti-inflammatory impact of Gleditsia sinensis Lam. and Lactobacillus brevis KY21 on intestinal epithelial cells in a DSS-induced colitis model. Korean J. food Sci. Anim. Resour. 2015, 35, 604–610. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, S.; Watanabe, N.; Imaoka, A.; Okabe, Y. Preventive effects of Bifidobacterium- and Lactobacillus-fermented milk on the development of inflammatory bowel disease in senescence-accelerated mouse P1/Yit strain mice. Digestion 2001, 64, 92–99. [Google Scholar] [CrossRef]
- Yoon, J.-W.; Ahn, S.-I.; Jhoo, J.-W.; Kim, G.-Y. Antioxidant activity of yogurt fermented at low temperature and its anti-inflammatory effect on DSS-induced colitis in mice. Food Sci. Anim. Resour. 2019, 39, 162–176. [Google Scholar] [CrossRef]
- Zagato, E.; Mileti, E.; Massimiliano, L.; Fasano, F.; Budelli, A.; Penna, G.; Rescigno, M. Lactobacillus paracasei CBA L74 metabolic products and fermented milk for infant formula have anti-inflammatory activity on dendritic cells in vitro and protective effects against colitis and an enteric pathogen in vivo. PLoS ONE 2014, 9, e87615. [Google Scholar] [CrossRef]
- Yoda, K.; Miyazawa, K.; Hosoda, M.; Hiramatsu, M.; Yan, F.; He, F. Lactobacillus GG-fermented milk prevents DSS-induced colitis and regulates intestinal epithelial homeostasis through activation of epidermal growth factor receptor. Eur. J. Nutr. 2014, 53, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, M.; Nemoto, M.; Nakata, T.; Kondo, S.; Takahashi, H.; Kimura, B.; Kuda, T. Anti-inflammatory properties of fermented soy milk with Lactococcus lactis subsp. lactis S-SU2 in murine macrophage RAW264.7 cells and DSS-induced IBD model mice. Int. Immunopharmacol. 2015, 26, 295–303. [Google Scholar] [CrossRef] [PubMed]
- Lim, J.; Seo, B.J.; Kim, J.E.; Chae, C.S.; Im, S.H.; Hahn, Y.S.; Park, Y.H. Characteristics of immunomodulation by a Lactobacillus sakei proBio65 isolated from Kimchi. Korean J. Microbiol. Biotechnol. 2011, 39, 313–316. [Google Scholar]
- Granier, A.; Goulet, O.; Hoarau, C. Fermentation products: Immunological effects on human and animal models. Pediatr. Res. 2013, 74, 238–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balamurugan, R.; Chandragunasekaran, A.S.; Chellappan, G.; Rajaram, K.; Ramamoorthi, G.; Ramakrishna, B.S. Probiotic potential of lactic acid bacteria present in home made curd in southern India. Indian J. Med. Res. 2014, 140, 345. [Google Scholar]
- Asakura, H.; Kitahora, T. Antioxidants and Polyphenols in Inflammatory Bowel Disease: Ulcerative Colitis and Crohn Disease. In Polyphenols: Prevention and Treatment of Human Disease, 2nd ed.; Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Acadamic Press: Cambridge, MA, USA, 2018; Volume 2, pp. 279–292. [Google Scholar]
- Chen, L.; You, Q.; Hu, L.; Gao, J.; Meng, Q.; Liu, W.; Wu, X.; Xu, Q. The antioxidant procyanidin reduces reactive oxygen species signaling in macrophages and ameliorates experimental colitis in mice. Front. Immunol. 2018, 8, 1910. [Google Scholar] [CrossRef] [Green Version]
- Ruan, W.; Engevik, M.; Chang-Graham, A.; Hyser, J.; Versalovic, J. 7 Lactobacillus reuteri suppresses pro-inflammatory driven reactive oxygen species in vitro in human intestinal epithelial cells and in vivo in a TNBS colitis mouse model. Inflamm. Bowel Dis. 2020, 26, S41. [Google Scholar] [CrossRef]
- Seguí, J.; Gil, F.; Gironella, M.; Alvarez, M.; Gimeno, M.; Coronel, P.; Closa, D.; Piqué, J.M.; Panés, J. Down-regulation of endothelial adhesion molecules and leukocyte adhesion by treatment with superoxide dismutase is beneficial in chronic immune experimental colitis. Inflamm. Bowel Dis. 2005, 11, 872–882. [Google Scholar] [CrossRef] [Green Version]
- Carroll, I.M.; Andrus, J.M.; Bruno-Bárcena, J.M.; Klaenhammer, T.R.; Hassan, H.M.; Threadgill, D.S. Anti-inflammatory properties of Lactobacillus gasseri expressing manganese superoxide dismutase using the interleukin 10-deficient mouse model of colitis. Am. J. Physiol. Liver Physiol. 2007, 293, G729–G738. [Google Scholar]
- Ishihara, T.; Tanaka, K.-I.; Tasaka, Y.; Namba, T.; Suzuki, J.; Ishihara, T.; Okamoto, S.; Hibi, T.; Takenaga, M.; Igarashi, R.; et al. Therapeutic effect of lecithinized superoxide dismutase against colitis. J. Pharmacol. Exp. Ther. 2009, 328, 152–164. [Google Scholar] [CrossRef] [Green Version]
- Watterlot, L.; Rochat, T.; Sokol, H.; Cherbuy, C.; Bouloufa, I.; Lefèvre, F.; Gratadoux, J.-J.; Honvo-Hueto, E.; Chilmonczyk, S.; Blugeon, S. Intragastric administration of a superoxide dismutase-producing recombinant Lactobacillus casei BL23 strain attenuates DSS colitis in mice. Int. J. Food Microbiol. 2010, 144, 35–41. [Google Scholar] [CrossRef]
- Baumgart, D.C.; Sandborn, W.J. Inflammatory bowel disease: Clinical aspects and established and evolving therapies. Lancet 2007, 369, 1641–1657. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, X.; Wang, H.; Yang, Z.; Li, J.; Suo, H. Prevent effects of Lactobacillus Fermentum HY01 on dextran sulfate sodium-induced colitis in mice. Nutrients 2017, 9, 545. [Google Scholar] [CrossRef] [PubMed]
- Song, J.-L.; Choi, J.-H.; Seo, J.-H.; Lim, Y.-I.; Park, K.-Y. Anti-colitic effects of kanjangs (fermented soy sauce and sesame sauce) in dextran sulfate sodium-induced colitis in mice. J. Med. Food 2014, 17, 1027–1035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duary, R.K.; Bhausaheb, M.A.; Batish, V.K.; Grover, S. Anti-inflammatory and immunomodulatory efficacy of indigenous probiotic Lactobacillus plantarum Lp91 in colitis mouse model. Mol. Biol. Rep. 2012, 39, 4765–4775. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Ho, S.B. Intestinal goblet cells and mucins in health and disease: Recent insights and progress. Curr. Gastroenterol. Rep. 2010, 12, 319–330. [Google Scholar] [CrossRef] [Green Version]
- Wong, E.T.; Tergaonkar, V. Roles of NF-κB in health and disease: Mechanisms and therapeutic potential. Clin. Sci. 2009, 116, 451–465. [Google Scholar] [CrossRef]
- Suzuki, T. Regulation of intestinal epithelial permeability by tight junctions. Cell. Mol. Life Sci. 2013, 70, 631–659. [Google Scholar] [CrossRef]
- Lee, S.H. Intestinal permeability regulation by tight junction: Implication on inflammatory bowel diseases. Intest. Res. 2015, 13, 11. [Google Scholar] [CrossRef] [Green Version]
- Fanning, A.S.; Ma, T.Y.; Anderson, J.M. Isolation and functional characterization of the actin-binding region in the tight junction protein ZO-1. FASEB J. 2002, 16, 1–23. [Google Scholar] [CrossRef]
- Tsukita, S.; Furuse, M. Pores in the wall: Claudins constitute tight junction strands containing aqueous pores. J. Cell Biol. 2000, 149, 13–16. [Google Scholar] [CrossRef] [Green Version]
- Tamura, A.; Hayashi, H.; Imasato, M.; Yamazaki, Y.; Hagiwara, A.; Wada, M.; Noda, T.; Watanabe, M.; Suzuki, Y.; Tsukita, S. Loss of claudin-15, but not claudin-2, causes Na+ deficiency and glucose malabsorption in mouse small intestine. Gastroenterology 2011, 140, 913–923. [Google Scholar] [CrossRef] [PubMed]
- Van Itallie, C.M.; Anderson, J.M. Claudins and epithelial paracellular transport. Annu. Rev. Physiol. 2006, 68, 403–429. [Google Scholar] [CrossRef]
- Furuse, M.; Hata, M.; Furuse, K.; Yoshida, Y.; Haratake, A.; Sugitani, Y.; Noda, T.; Kubo, A.; Tsukita, S. Claudin-based tight junctions are crucial for the mammalian epidermal barrier: A lesson from claudin-1-deficient mice. J. Cell Biol. 2002, 156, 1099–1111. [Google Scholar] [CrossRef]
- Weber, C.R.; Nalle, S.C.; Tretiakova, M.; Rubin, D.T.; Turner, J.R. Claudin-1 and claudin-2 expression is elevated in inflammatory bowel disease and may contribute to early neoplastic transformation. Lab. Investig. 2008, 88, 1110–1120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poritz, L.S.; Harris, L.R.; Kelly, A.A.; Koltun, W.A. Increase in the tight junction protein claudin-1 in intestinal inflammation. Dig. Dis. Sci. 2011, 56, 2802. [Google Scholar] [CrossRef] [Green Version]
- Kinugasa, T.; Akagi, Y.; Yoshida, T.; Ryu, Y.; Shiratuchi, I.; Ishibashi, N.; Shirouzu, K. Increased claudin-1 protein expression contributes to tumorigenesis in ulcerative colitis-associated colorectal cancer. Anticancer Res. 2010, 30, 3181–3186. [Google Scholar]
- Castro-Ochoa, K.F.; Vargas-Robles, H.; Chánez-Paredes, S.; Felipe-López, A.; Cabrera-Silva, R.I.; Shibayama, M.; Betanzos, A.; Nava, P.; Galinski, E.A.; Schnoor, M. Homoectoine protects against colitis by preventing a claudin switch in epithelial tight junctions. Dig. Dis. Sci. 2019, 64, 409–420. [Google Scholar] [CrossRef]
- Xu, Q.; Li, X.; Wang, E.; He, Y.; Yin, B.; Fang, D.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. A cellular model for screening of lactobacilli that can enhance tight junctions. RSC Adv. 2016, 6, 111812–111821. [Google Scholar] [CrossRef]
- Xiaolin, W.U.; Zhao, C.; Jiang, H.; Fan, C.; Wang, L. Changes of expressions of claudin-1,-2,-4 in experimental colitis rats. Chin. J. Gastroenterol. 2017, 22, 599–603. [Google Scholar]
- Ma, T.Y.; Boivin, M.A.; Ye, D.; Pedram, A.; Said, H.M. Mechanism of TNF-α modulation of Caco-2 intestinal epithelial tight junction barrier: Role of myosin light-chain kinase protein expression. Am. J. Physiol. Liver Physiol. 2005, 288, G422–G430. [Google Scholar] [CrossRef] [Green Version]
- Schulzke, J.; Bojarski, C.; Zeissig, S.; Heller, F.; Gitter, A.H.; Fromm, M. Disrupted barrier function through epithelial cell apoptosis. Ann. N. Y. Acad. Sci. 2006, 1072, 288–299. [Google Scholar] [CrossRef]
- Bouma, G.; Strober, W. The immunological and genetic basis of inflammatory bowel disease. Nat. Rev. Immunol. 2003, 3, 521–533. [Google Scholar] [CrossRef]
- Al-Sadi, R.; Ye, D.; Dokladny, K.; Ma, T.Y. Mechanism of IL-1β-induced increase in intestinal epithelial tight junction permeability. J. Immunol. 2008, 180, 5653–5661. [Google Scholar] [CrossRef] [Green Version]
- Stallmach, A.; Giese, T.; Schmidt, C.; Ludwig, B.; Mueller-Molaian, I.; Meuer, S.C. Cytokine/chemokine transcript profiles reflect mucosal inflammation in Crohn’s disease. Int. J. Colorectal Dis. 2004, 19, 308–315. [Google Scholar] [CrossRef]
- Bruewer, M.; Utech, M.; Ivanov, A.I.; Hopkins, A.M.; Parkos, C.A.; Nusrat, A. Interferon-γ induces internalization of epithelial tight junction proteins via a macropinocytosis-like process. FASEB J. 2005, 19, 923–933. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score | Weight Loss (%) | Stool Consistency 2 | Gross Bleeding |
|---|---|---|---|
| 0 | 0 | Normal | Negative |
| 1 | 1–5 | Loose | Negative |
| 2 | 6–10 | Loose | Hemoccult positive |
| 3 | 11–15 | Diarrhea | Hemoccult positive |
| 4 | >15 | Diarrhea | Bleeding |
| Genes | Oligonucleotide Sequences | References |
|---|---|---|
| IL-1β | Forward: TTGACGGACCCCAAAAGATG | [25] |
| Reverse:AGAAGGTGCTCATGTCCTCA | ||
| TNF-α | Forward: TCTCATCAGTTCTATGGCCC | [26] |
| Reverse: GGGAGTAGACAAGGTACAAC | ||
| IFN-γ | Forward: CTGAGACAATGAACGCTACACACTGC | [27] |
| Reverse: AACAGCTGGTGGACCACTCGGAT | ||
| GAPDH | Forward: CAAAAGGGTCATCATCTCTG | [25] |
| Reverse: CCTGCTTCACCACCTTCTTG |
| Lactobacillus Strains | Acid Tolerance 1 (log10 CFU/mL) | Bile Acid Tolerance 2 (log10 CFU/mL) | ||||
|---|---|---|---|---|---|---|
| 0 h | 1 h | 2 h | 0 h | 24 h | 48 h | |
| 1 | 5.71 ± 0.00 | 5.66 ± 0.00 | 0.17 ± 0.29 c | 5.61 ± 0.01 | 6.79 ± 0.00 | 6.82 ± 0.00 b,c |
| 2 | 5.72 ± 0.00 | 5.72 ± 0.04 | 4.67 ± 0.00 a | 5.76 ± 0.00 | 6.87 ± 0.00 | 5.72 ± 0.00 f |
| 3 | 5.83 ± 0.02 | 4.63 ± 0.00 | 0.13 ± 0.23 c | 4.84 ± 0.00 | 6.88 ± 0.00 | 5.54 ± 0.05 e |
| 4 | 5.74 ± 0.01 | 5.78 ± 0.01 | 4.67 ± 0.00 a | 4.80 ± 0.00 | 5.66 ± 0.00 | 5.52 ± 0.00 e |
| 5 | 6.17 ± 0.01 | 5.63 ± 0.05 | 0.14 ± 0.25 d | 5.73 ± 0.00 | 6.83 ± 0.00 | 6.72 ± 0.00 d |
| 8 | 6.76 ± 0.05 | 6.73 ± 0.00 | 4.58 ± 0.01 b | 5.70 ± 0.00 | 6.87 ± 0.00 | 6.78 ± 0.03 b |
| 9 | 6.84 ± 0.05 | 5.68 ± 0.00 | 4.53 ± 0.00 b | 5.84 ± 0.00 | 6.92 ± 0.00 | 6.92 ± 0.00 b |
| R10 | 6.81 ± 0.00 | 6.74 ± 0.00 | 5.52 ± 0.01 a | 5.84 ± 0.00 | 6.92 ± 0.04 | 6.93 ± 0.00 b |
| P11 | 6.81 ± 0.01 | 6.82 ± 0.00 | 5.75 ± 0.01 a | 5.85 ± 0.00 | 6.85 ± 0.04 | 6.85 ± 0.01 b |
| Bmb6 | 6.86 ± 0.00 | 5.71 ± 0.00 | 5.76 ± 0.01 a | 5.83 ± 0.00 | 6.90 ± 0.00 | 7.94 ± 0.00 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, M.-Y.; Yong, C.-C.; Oh, S. Regulatory Effect of Lactobacillus brevis Bmb6 on Gut Barrier Functions in Experimental Colitis. Foods 2020, 9, 864. https://doi.org/10.3390/foods9070864
Shin M-Y, Yong C-C, Oh S. Regulatory Effect of Lactobacillus brevis Bmb6 on Gut Barrier Functions in Experimental Colitis. Foods. 2020; 9(7):864. https://doi.org/10.3390/foods9070864
Chicago/Turabian StyleShin, Mi-Young, Cheng-Chung Yong, and Sejong Oh. 2020. "Regulatory Effect of Lactobacillus brevis Bmb6 on Gut Barrier Functions in Experimental Colitis" Foods 9, no. 7: 864. https://doi.org/10.3390/foods9070864