The Role of Enterococcus faecium as a Key Producer and Fermentation Condition as an Influencing Factor in Tyramine Accumulation in Cheonggukjang

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Cheonggukjang Products
2.2. Physicochemical Analyses
2.3. Microbial Analyses
2.4. Isolation and Identification of Enterococcus Strains from Retail Cheonggukjang Products
2.5. Preparation of Cheonggukjang
2.6. BA Analyses in Cheonggukjang Samples and Bacterial Cultures
2.6.1. BA Extraction from Cheonggukjang Samples and Bacterial Cultures
2.6.2. Preparation of Standard Solutions for High Performance Liquid Chromatography (HPLC) Analysis
2.6.3. Derivatization of Extracts and Standards
2.6.4. Chromatographic Separations
2.7. Gene Expression Analyses in Bacterial Cultures and Cheonggukjang
2.7.1. RNA Extraction and Reverse Transcription
2.7.2. Quantitative PCR Analysis
2.8. Statistical Analyses
3. Results and Discussion
3.1. Physicochemical Properties of Retail Cheonggukjang Products
3.2. Microbial Properties of Retail Cheonggukjang Products
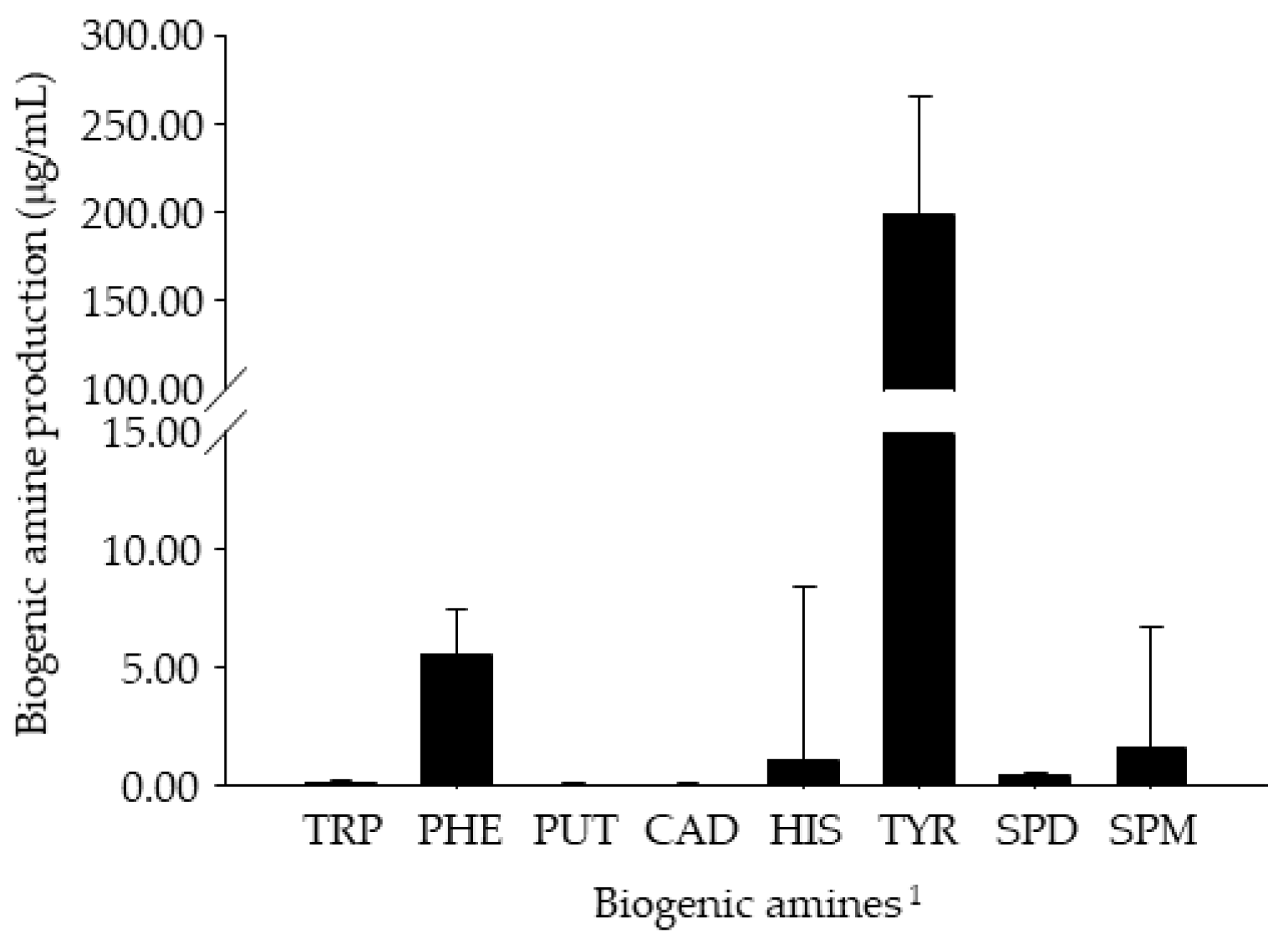
3.3. BA Content of Retail Cheonggukjang Products
3.4. In Vitro BA Production by Enterococcus Strains Isolated from Retail Cheonggukjang Products
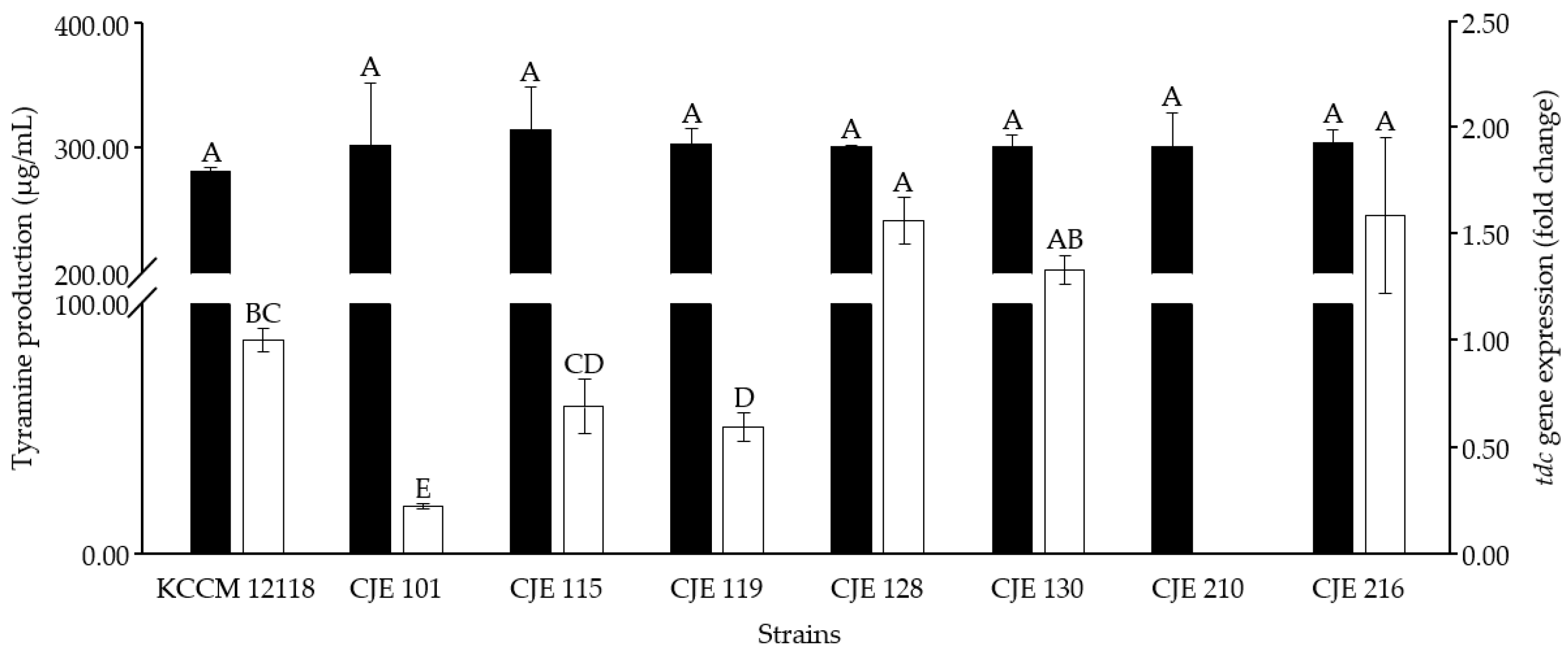
3.5. Selection of Tyramine-Producing E. faecium Strain for Cheonggukjang Fermentation Based on Tyrosine Decarboxylase Gene Expression In Vitro
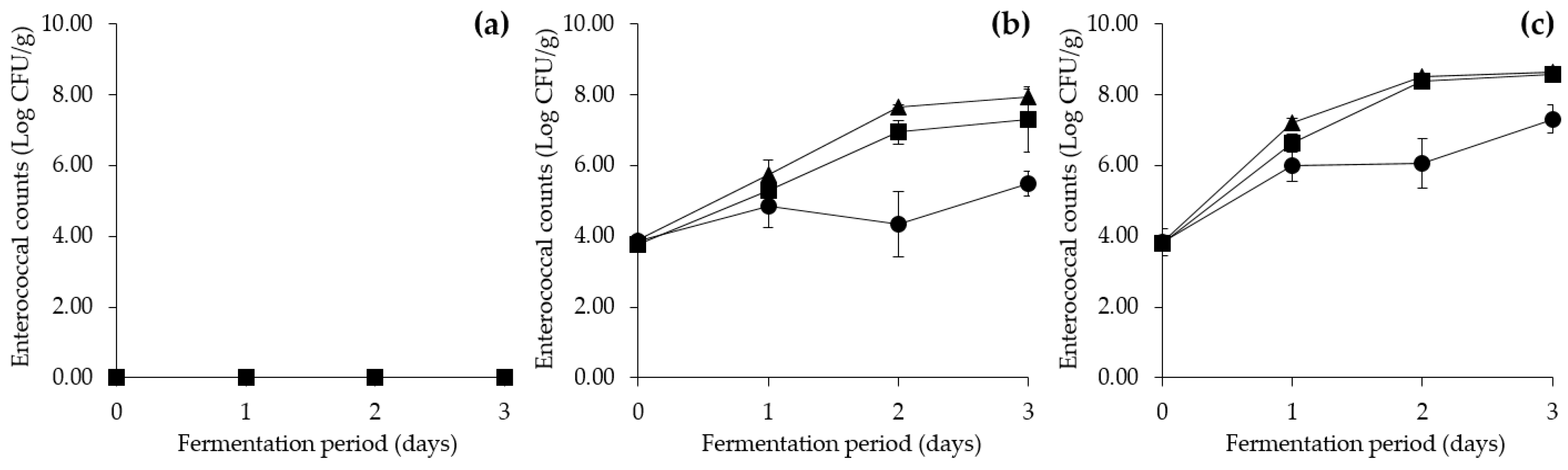
3.6. Tyramine Production by E. faecium during Cheonggukjang Fermentation at Various Temperatures
3.6.1. Changes in Physicochemical and Microbial Properties during Cheonggukjang Fermentation at Various Temperatures
3.6.2. Effect of Fermentation Temperature on Tyramine Content in Cheonggukjang
3.6.3. Effect of Fermentation Temperature on tdc Gene Expression by E. faecium Strains in Cheonggukjang
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jang, C.H.; Lim, J.K.; Kim, J.H.; Park, C.S.; Kwon, D.Y.; Kim, Y.-S.; Shin, D.H.; Kim, J.-S. Change of isoflavone content during manufacturing of cheonggukjang, a traditional Korean fermented soyfood. Food Sci. Biotechnol. 2006, 15, 643–646. [Google Scholar]
- Kim, K.-J.; Ryu, M.-K.; Kim, S.-S. Chungkook-jang Koji fermentation with rice straw. Korean J. Food Sci. Technol. 1982, 14, 301–308. [Google Scholar]
- Hong, S.W.; Kim, J.Y.; Lee, B.K.; Chung, K.S. The bacterial biological response modifier enriched Chungkookjang fermentation. Korean J. Food Sci. Technol. 2006, 38, 548–553. [Google Scholar]
- Lee, J.-O.; Ha, S.-D.; Kim, A.-J.; Yuh, C.-S.; Bang, I.-S.; Park, S.-H. Industrial application and physiological functions of Chongkukjang. Food Sci. Ind. 2005, 38, 69–78. [Google Scholar]
- Hwang, J.-S.; Kim, S.-J.; Kim, H.-B. Antioxidant and blood-pressure reduction effects of fermented soybean, Chungkookjang. Korean J. Microbiol. 2009, 45, 54–57. [Google Scholar]
- Askar, A.; Treptow, H. Biogene Amine in Lebensmitteln: Vorkommen, Bedeutung und Bestimmung, 1st ed.; Eugen Ulmer: Stuttgart, Germany, 1986; pp. 21–74. [Google Scholar]
- Silla Santos, M.H. Biogenic amines: Their importance in foods. Int. J. Food Microbiol. 1996, 29, 213–231. [Google Scholar] [CrossRef]
- Shalaby, A.R. Significance of biogenic amines to food safety and human health. Food Res. Int. 1996, 29, 675–690. [Google Scholar] [CrossRef]
- Ladero, V.; Calles-Enriquez, M.; Fernández, M.; Alvarez, M.A. Toxicological effects of dietary biogenic amines. Curr. Nutr. Food Sci. 2010, 6, 145–156. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards (BIOHAZ). Scientific opinion on risk based control of biogenic amine formation in fermented foods. EFSA J. 2011, 9, 2393. [Google Scholar] [CrossRef]
- Taylor, S.L.; Eitnmiller, R.R. Histamine food poisoning: Toxicology and clinical aspects. Crit. Rev. Toxicol. 1986, 17, 91–128. [Google Scholar] [CrossRef]
- Kovacova-Hanuskova, E.; Buday, T.; Gavliakova, S.; Plevkova, J. Histamine, histamine intoxication and intolerance. Allergol. Immunopathol. 2015, 43, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Maintz, L.; Novak, N. Histamine and histamine intolerance. Am. J. Clin. Nutr. 2007, 85, 1185–1196. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.A. Amines in food. Food Chem. 1981, 6, 169–200. [Google Scholar] [CrossRef]
- Stratton, J.E.; Hutkins, R.W.; Taylor, S.L. Biogenic amines in cheese and other fermented foods: A review. J. Food Prot. 1991, 54, 460–470. [Google Scholar] [CrossRef]
- Linares, D.M.; Martín, M.; Ladero, V.; Alvarez, M.A.; Fernández, M. Biogenic amines in dairy products. Crit. Rev. Food Sci. Nutr. 2011, 51, 691–703. [Google Scholar] [CrossRef]
- Schirone, M.; Tofalo, T.; Fasoli, G.; Perpetuini, G.; Corsetti, A.; Manetta, A.C.; Ciarrocchi, A.; Suzzi, G. High content of biogenic amines in Pecorino cheeses. Food Microbiol. 2013, 34, 137–144. [Google Scholar] [CrossRef]
- Ko, Y.-J.; Son, Y.-H.; Kim, E.-J.; Seol, H.-G.; Lee, G.-R.; Kim, D.-H.; Ryu, C.-H. Quality properties of commercial Chungkukjang in Korea. J. Agric. Life Sci. 2012, 46, 177–187. [Google Scholar]
- Jeon, A.R.; Lee, J.H.; Mah, J.-H. Biogenic amine formation and bacterial contribution in Cheonggukjang, a Korean traditional fermented soybean food. LWT Food Sci. Technol. 2018, 92, 282–289. [Google Scholar] [CrossRef]
- Seo, M.-J.; Lee, C.-D.; Lee, J.-N.; Yang, H.-J.; Jeong, D.-Y.; Lee, G.-H. Analysis of biogenic amines and inorganic elements in Cheonggukjang. Korean J. Food Preserv. 2019, 26, 101–108. [Google Scholar] [CrossRef]
- Ten Brink, B.; Damink, C.; Joosten, H.M.L.J.; Huis in ‘t Veld, J.H.J. Occurrence and formation of biologically active amines in foods. Int. J. Food Microbiol. 1990, 11, 73–84. [Google Scholar] [CrossRef]
- Ibe, A.; Nishima, T.; Kasai, N. Bacteriological properties of and amine-production conditions for tyramine-and histamine-producing bacterial strains isolated from soybean paste (miso) starting materials. Jpn. J. Toxicol. Environ. Health 1992, 38, 403–409. [Google Scholar] [CrossRef]
- Torriani, S.; Gatto, V.; Sembeni, S.; Tofalo, R.; Suzzi, G.; Belletti, N.; Gardini, F.; Bover-Cid, S. Rapid detection and quantification of tyrosine decarboxylase gene (tdc) and its expression in gram-positive bacteria associated with fermented foods using PCR-based methods. J. Food Prot. 2008, 71, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Ladero, V.; Fernández, M.; Cuesta, I.; Alvarez, M.A. Quantitative detection and identification of tyramine-producing enterococci and lactobacilli in cheese by multiplex qPCR. Food Microbiol. 2010, 27, 933–939. [Google Scholar] [CrossRef] [PubMed]
- Kang, T.-M.; Park, J.-H. Isolation and antibiotic susceptibility of Enterococcus spp. from fermented soy paste. J. Korean Soc. Food Sci. Nutr. 2012, 41, 714–720. [Google Scholar] [CrossRef][Green Version]
- Kang, H.-R.; Lee, Y.-L.; Hwang, H.-J. Potential for application as a starter culture of tyramine-reducing strain. J. Korean Soc. Food Sci. Nutr. 2017, 46, 1561–1567. [Google Scholar] [CrossRef]
- Svec, P.; Devriese, L.A. Enterococcus. In Bergey’s Manual of Systematics of Archaea and Bacteria, 2nd ed.; De Vos, P., Garrity, G.M., Jones, D., Krieg, N.R., Ludwig, W., Rainey, F.A., Schleifer, K.-H., Whitman, W.B., Eds.; Springer: New York, NY, USA, 2015; Volume 3, pp. 594–607. [Google Scholar]
- Food Information Statistics System. Available online: http://www.atfis.or.kr/article/M001050000/view.do?articleId=2452&page=5&searchKey=&searchString=&searchCategory= (accessed on 29 May 2020).
- Kim, I.-J.; Kim, H.-K.; Chung, J.-H.; Jeong, Y.-K.; Ryu, C.-H. Study of functional Chungkukjang contain fibrinolytic enzyme. Korean J. Life Sci. 2002, 12, 357–362. [Google Scholar]
- Lee, N.-R.; Go, T.-H.; Lee, S.-M.; Hong, C.-O.; Park, K.-M.; Park, G.-T.; Hwang, D.-Y.; Son, H.-J. Characteristics of Chungkookjang prepared by Bacillus amyloliquefaciens with different soybeans and fermentation temperatures. Korean J. Microbiol. 2013, 49, 71–77. [Google Scholar] [CrossRef]
- Bhardwaj, A.; Gupta, H.; Iyer, R.; Kumar, N.; Malik, R.K. Tyramine-producing enterococci are equally detected on tyramine production medium, by quantification of tyramine by HPLC, or by tdc gene-targeted PCR. Dairy Sci. Technol. 2009, 89, 601–611. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of AOAC International, 18th ed.; AOAC International: Gaithersburg, MD, USA, 2005. [Google Scholar]
- ISO 7218:2007. Microbiology of Food and Animal Feeding Stuffs—General Requirements and Guidance for Microbiological Examinations; ISO: Geneva, Switzerland, 2007. [Google Scholar]
- Mareková, M.; Lauková, A.; DeVuyst, L.; Skaugen, M.; Nes, I.F. Partial characterization of bacteriocins produced by environmental strain Enterococcus faecium EK13. J. Appl. Microbiol. 2003, 94, 523–530. [Google Scholar] [CrossRef]
- Ryu, M.S.; Yang, H.-J.; Kim, J.W.; Jeong, S.-J.; Jeong, S.-Y.; Eom, J.-S.; Jeong, D.-Y. Potential probiotics activity of Bacillus spp. from traditional soybean pastes and fermentation characteristics of Cheonggukjang. Korean J. Food Preserv. 2017, 24, 1168–1179. [Google Scholar] [CrossRef]
- Lee, J.S.; Lee, M.H.; Kim, J.M. Changes in quality characteristics of cheonggukjang added with quinoa during fermentation period. Korean J. Food Nutr. 2018, 31, 24–32. [Google Scholar]
- National Institute of Agricultural Sciences. Available online: https://koreanfood.rda.go.kr:2360/eng/fctFoodSrchEng/engMain (accessed on 7 May 2020).
- Ben-Gigirey, B.; Vieites Baptista De Sousa, J.M.; Villa, T.G.; Barros-Velazquez, J. Changes in biogenic amines and microbiological analysis in albacore (Thunnus alalunga) muscle during frozen storage. J. Food Prot. 1998, 61, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Eerola, S.; Hinkkanen, R.; Lindfors, E.; Hirvi, T. Liquid chromatographic determination of biogenic amines in dry sausages. J. AOAC Int. 1993, 76, 575–577. [Google Scholar] [CrossRef]
- Ben-Gigirey, B.; Vieites Baptista De Sousa, J.M.; Villa, T.G.; Barros-Velazquez, J. Histamine and cadaverine production by bacteria isolated from fresh and frozen albacore (Thunnus alalunga). J. Food Prot. 1999, 62, 933–939. [Google Scholar] [CrossRef] [PubMed]
- Marcobal, Á.; Martín-Álvarez, P.J.; Moreno-Arribas, M.V.; Muñoz, R. A multifactorial design for studying factors influencing growth and tyramine production of the lactic acid bacteria Lactobacillus brevis CECT 4669 and Enterococcus faecium BIFI-58. Res. Microbiol. 2006, 157, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Yoon, H.; Park, J.H.; Choi, A.; Hwang, H.-J.; Mah, J.-H. Validation of an HPLC analytical method for determination of biogenic amines in agricultural products and monitoring of biogenic amines in Korean fermented agricultural products. Toxicol. Res. 2015, 31, 299–305. [Google Scholar] [CrossRef]
- Kang, H.-R.; Kim, H.-S.; Mah, J.-H.; Kim, Y.-W.; Hwang, H.-J. Tyramine reduction by tyrosine decarboxylase inhibitor in Enterococcus faecium for tyramine controlled cheonggukjang. Food Sci. Biotechnol. 2018, 27, 87–93. [Google Scholar] [CrossRef]
- Top, J.; Paganelli, F.L.; Zhang, X.; van Schaik, W.; Leavis, H.L.; Van Luit-Asbroek, M.; van der Poll, T.; Leendertse, M.; Bonten, M.J.M.; Willems, R.J.L. The Enterococcus faecium enterococcal biofilm regulator, EbrB, regulates the esp operon and is implicated in biofilm formation and intestinal colonization. PLoS ONE 2013, 8, e65224. [Google Scholar] [CrossRef]
- Lee, E.S.; Kim, Y.S.; Ryu, M.S.; Jeong, D.Y.; Uhm, T.B.; Cho, S.H. Characterization of Bacillus licheniformis SCK A08 with antagonistic property against Bacillus cereus and degrading capacity of biogenic amines. J. Food Hyg. Saf. 2014, 29, 40–46. [Google Scholar] [CrossRef][Green Version]
- Yoo, S.-M.; Choe, J.-S.; Park, H.-J.; Hong, S.-P.; Chang, C.-M.; Kim, J.-S. Physicochemical properties of traditional Chonggugjang produced in different regions. Appl. Biol. Chem. 1998, 41, 377–383. [Google Scholar]
- Kim, J.-W.; Kim, Y.-S.; Jeong, P.-H.; Kim, H.-E.; Shin, D.-H. Physicochemical characteristics of traditional fermented soybean products manufactured in folk villages of Sunchang region. J. Food Hyg. Saf. 2006, 21, 223–230. [Google Scholar]
- Jeong, W.J.; Lee, A.R.; Chun, J.; Cha, J.; Song, Y.-S.; Kim, J.H. Properties of cheonggukjang fermented with Bacillus strains with high fibrinolytic activities. J. Food Sci. Nutr. 2009, 14, 252–259. [Google Scholar] [CrossRef]
- Kang, S.J.; Kim, S.S.; Chung, H.Y. Comparison of physicochemical characteristics and consumer perception of Cheongkukjang. J. Korean Soc. Food Sci. Nutr. 2014, 43, 1104–1111. [Google Scholar] [CrossRef]
- Oh, S.-J.; Mah, J.-H.; Kim, J.-H.; Kim, Y.-W.; Hwang, H.-J. Reduction of tyramine by addition of Schizandra chinensis Baillon in Cheonggukjang. J. Med. Food 2012, 15, 1109–1115. [Google Scholar] [CrossRef] [PubMed]
- International Commission on Microbiological Specifications for Foods International Association of Microbiological Societies. Reduced water activity. In Microbial Ecology of Foods, 1st ed.; Silliker, J.H., Elliot, R.P., Baird-Parker, A.C., Bryan, F.L., Christian, J.H.B., Clark, D.S., Olson, J.C., Roberts, T.A., Eds.; Academic Press: New York, NY, USA, 1980; Volume 1, pp. 70–91. [Google Scholar]
- Li, L.; Ruan, L.; Ji, A.; Wen, Z.; Chen, S.; Wang, L.; Wei, X. Biogenic amines analysis and microbial contribution in traditional fermented food of Douchi. Sci. Rep. 2018, 8, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Takebe, Y.; Takizaki, M.; Tanaka, H.; Ohta, H.; Niidome, T.; Morimura, S. Evaluation of the biogenic amine-production ability of lactic acid bacteria isolated from tofu-misozuke. Food Sci. Technol. Res. 2016, 22, 673–678. [Google Scholar] [CrossRef]
- Giraffa, G. Enterococci from foods. FEMS Microbiol. Rev. 2002, 26, 163–171. [Google Scholar] [CrossRef]
- Giraffa, G.; Carminati, D.; Neviani, E. Enterococci isolated from dairy products: A review of risks and potential technological use. J. Food Prot. 1997, 60, 732–738. [Google Scholar] [CrossRef]
- Bandara, N.; Chung, S.-J.; Jeong, D.-Y.; Kim, K.-P. The use of the pathogen-specific bacteriophage BCP8-2 to develop a rice straw-derived Bacillus cereus-free starter culture. Korean J. Food Sci. Technol. 2014, 46, 115–120. [Google Scholar] [CrossRef]
- Heu, J.-S.; Lee, I.-J.; Yoon, M.-H.; Choi, W.-Y. Adhesive microbial populations of rice straws and their effects on Chungkukjang fermentation. Korean J. Agric. Sci. 1999, 26, 77–83. [Google Scholar]
- Seok, Y.-R.; Kim, Y.-H.; Kim, S.; Woo, H.-S.; Kim, T.-W.; Lee, S.-H.; Choi, C. Change of protein and amino acid composition during Chungkook-Jang fermentation using Bacillus licheniformis CN-115. Korean J. Agic. Sci. 1994, 37, 65–71. [Google Scholar]
- Cho, T.-Y.; Han, G.-H.; Bahn, K.-N.; Son, Y.-W.; Jang, M.-R.; Lee, C.-H.; Kim, S.-H.; Kim, D.-B.; Kim, S.-B. Evaluation of biogenic amines in Korean commercial fermented foods. Korean J. Food Sci. Technol. 2006, 38, 730–737. [Google Scholar]
- Han, G.-H.; Cho, T.-Y.; Yoo, M.-S.; Kim, C.-S.; Kim, J.-M.; Kim, H.-A.; Kim, M.-O.; Kim, S.-C.; Lee, S.-A.; Ko, Y.-S.; et al. Biogenic amines formation and content in fermented soybean paste (cheonggukjang). Korean J. Food Sci. Technol. 2007, 39, 541–545. [Google Scholar]
- Rice, S.L.; Eitenmiller, R.R.; Koehler, P.E. Biologically active amines in food: A review. J. Milk Food Technol. 1976, 39, 353–358. [Google Scholar] [CrossRef]
- Rodriguez-Jerez, J.J.; Giaccone, V.; Colavita, G.; Parisi, E. Bacillus macerans—A new potent histamine producing micro-organism isolated from Italian cheese. Food Microbiol. 1994, 11, 409–415. [Google Scholar] [CrossRef]
- Novella-Rodríguez, S.; Veciana-Nogues, M.T.; Roig-Sagues, A.X.; Trujillo-Mesa, A.J.; Vidal-Carou, M.C. Evaluation of biogenic amines and microbial counts throughout the ripening of goat cheeses from pasteurized and raw milk. J. Dairy Res. 2004, 71, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Marcobal, A.; de las Rivas, B.; Moreno-Arribas, M.V.; Munoz, R. Evidence for horizontal gene transfer as origin of putrescine production in Oenococcus oeni RM83. Appl. Environ. Microbiol. 2006, 72, 7954–7958. [Google Scholar] [CrossRef] [PubMed]
- Condori, J.; Nopo-Olazabal, C.; Medrano, G.; Medina-Bolivar, F. Selection of reference genes for qPCR in hairy root cultures of peanut. BMC Res. Notes 2011, 4, 392. [Google Scholar] [CrossRef] [PubMed]
- Loizzo, M.R.; Menichini, F.; Picci, N.; Puoci, F.; Spizzirri, U.G.; Restuccia, D. Technological aspects and analytical determination of biogenic amines in cheese. Trends Food Sci. Technol. 2013, 30, 38–55. [Google Scholar] [CrossRef]
- Cosansu, S. Determination of biogenic amines in a fermented beverage, boza. J. Food Agric. Environ. 2009, 7, 54–58. [Google Scholar]
- Mann, S.-Y.; Kim, E.-A.; Lee, G.-Y.; Kim, R.-U.; Hwang, D.-Y.; Son, H.-J.; Kim, D.-S. Isolation and identification of GABA-producing microorganism from Chungkookjang. J. Life Sci. 2013, 23, 102–109. [Google Scholar] [CrossRef][Green Version]
- Morandi, S.; Brasca, M.; Alfieri, P.; Lodi, R.; Tamburini, A. Influence of pH and temperature on the growth of Enterococcus faecium and Enterococcus faecalis. Le Lait 2005, 85, 181–192. [Google Scholar] [CrossRef]
- Van den Berghe, E.; De Winter, T.; De Vuyst, L. Enterocin A production by Enterococcus faecium FAIR-E 406 is characterised by a temperature-and pH-dependent switch-off mechanism when growth is limited due to nutrient depletion. Int. J. Food Microbiol. 2006, 107, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Kalhotka, L.; Manga, I.; Přichystalová, J.; Hůlová, M.; Vyletělová, M.; Šustová, K. Decarboxylase activity test of the genus Enterococcus isolated from goat milk and cheese. Acta Vet. BRNO 2012, 81, 145–151. [Google Scholar] [CrossRef]
- Glanemann, C.; Loos, A.; Gorret, N.; Willis, L.B.; O’brien, X.M.; Lessard, P.A.; Sinskey, A.J. Disparity between changes in mRNA abundance and enzyme activity in Corynebacterium glutamicum: Implications for DNA microarray analysis. Appl. Microbiol. Biotechnol. 2003, 61, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Ladero, V.; Linares, D.M.; Fernández, M.; Alvarez, M.A. Real time quantitative PCR detection of histamine-producing lactic acid bacteria in cheese: Relation with histamine content. Food Res. Int. 2008, 41, 1015–1019. [Google Scholar] [CrossRef]
- Spano, G.; Russo, P.; Lonvaud-Funel, A.; Lucas, P.; Alexandre, H.; Grandvalet, C.; Coton, E.; Coton, M.; Barnavon, L.; Bach, B.; et al. Biogenic amines in fermented foods. Eur. J. Clin. Nutr. 2010, 64, S95–S100. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Products 1 | pH | Salinity (%) | Water Activity |
|---|---|---|---|
| CJ1 | 6.91 ± 0.03 2 | 5.54 ± 0.09 | 0.948 ± 0.002 |
| CJ2 | 6.84 ± 0.02 | 7.25 ± 0.06 | 0.919 ± 0.001 |
| CJ3 | 6.87 ± 0.02 | 3.16 ± 0.06 | 0.968 ± 0.002 |
| CJ4 | 6.99 ± 0.03 | 1.95 ± 0.09 | 0.973 ± 0.002 |
| CJ5 | 7.05 ± 0.05 | 9.36 ± 0.59 | 0.944 ± 0.003 |
| CJ6 | 6.39 ± 0.03 | 3.71 ± 0.34 | 0.954 ± 0.003 |
| Average | 6.84 ± 0.23 | 5.16 ± 2.78 | 0.951 ± 0.019 |
| Products 1 | Total Mesophilic Viable Bacteria (Log CFU/g) | Enterococcus spp. (Log CFU/g) | Lactic Acid Bacteria (Log CFU/g) |
|---|---|---|---|
| CJ1 | 9.45 ± 0.06 2 | 7.32 ± 0.03 | 7.45 ± 0.10 |
| CJ2 | 9.04 ± 0.09 | 6.78 ± 0.14 | 6.66 ± 0.10 |
| CJ3 | 9.81 ± 0.25 | 6.64 ± 0.01 | 6.70 ± 0.15 |
| CJ4 | 9.57 ± 0.15 | 7.27 ± 0.03 | 6.96 ± 0.09 |
| CJ5 | 8.54 ± 0.48 | 7.00 ± 0.08 | 6.67 ± 0.18 |
| CJ6 | 9.20 ± 0.44 | 7.99 ± 0.04 | 8.12 ± 0.01 |
| Average | 9.27 ± 0.45 | 7.17 ± 0.49 | 7.09 ± 0.58 |
| Products 1 | BA Content (mg/kg) 2 | |||||||
|---|---|---|---|---|---|---|---|---|
| TRP | PHE | PUT | CAD | HIS | TYR | SPD | SPM | |
| CJ1 | 115.06 ± 19.72 A,3 | 31.66 ± 3.82 B | 8.54 ± 3.45 BC | 22.66 ± 0.82 C | 8.60 ± 0.88 B | 222.25 ± 15.1 C | 137.88 ± 1.76 A | 292.99 ± 27.86 A |
| CJ2 | 86.02 ± 3.12 B | 16.06 ± 2.24 B | 12.18 ± 0.73 B | 29.95 ± 0.82 A | 12.69 ± 0.28 A | 57.14 ± 8.06 D | 99.99 ± 4.25 C | 206.32 ± 23.82 B |
| CJ3 | 118.27 ± 5.97 A | 27.22 ± 2.00 B | 18.33 ± 4.93 A | 26.58 ± 1.65 B | 6.50 ± 1.00 C | 80.83 ± 3.91 D | 96.82 ± 4.09 C | 201.63 ± 5.77 B |
| CJ4 | 54.87 ± 3.18 C | 22.20 ± 7.06 B | 11.36 ± 2.28 B | 8.28 ± 0.54 F | 5.86 ± 0.37 C | 898.41 ± 79.43 B | 125.61 ± 4.64 B | 91.09 ± 24.03 C |
| CJ5 | 47.85 ± 4.04 C | 24.48 ± 1.97 B | 11.61 ± 4.21 B | 13.57 ± 0.21 D | 8.65 ± 0.51 B | 61.98 ± 6.30 D | 125.98 ± 7.60 B | 305.05 ± 17.35 A |
| CJ6 | 1.70 ± 2.94 D | 95.58 ± 46.97 A | 2.81 ± 1.77 C | 10.19 ± 0.50 E | 7.84 ± 0.33 B | 1424.04 ± 62.43 A | 145.18 ± 7.64 A | 26.20 ± 8.51 D |
| Average | 70.63 ± 44.74 | 36.22 ± 29.55 | 10.80 ± 5.07 | 18.57 ± 9.08 | 8.37 ± 2.40 | 457.42 ± 573.15 | 121.92 ± 19.69 | 187.20 ± 110.27 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, Y.K.; Jin, Y.H.; Lee, J.-H.; Byun, B.Y.; Lee, J.; Jeong, K.C.; Mah, J.-H. The Role of Enterococcus faecium as a Key Producer and Fermentation Condition as an Influencing Factor in Tyramine Accumulation in Cheonggukjang. Foods 2020, 9, 915. https://doi.org/10.3390/foods9070915
Park YK, Jin YH, Lee J-H, Byun BY, Lee J, Jeong KC, Mah J-H. The Role of Enterococcus faecium as a Key Producer and Fermentation Condition as an Influencing Factor in Tyramine Accumulation in Cheonggukjang. Foods. 2020; 9(7):915. https://doi.org/10.3390/foods9070915
Chicago/Turabian StylePark, Young Kyoung, Young Hun Jin, Jun-Hee Lee, Bo Young Byun, Junsu Lee, KwangCheol Casey Jeong, and Jae-Hyung Mah. 2020. "The Role of Enterococcus faecium as a Key Producer and Fermentation Condition as an Influencing Factor in Tyramine Accumulation in Cheonggukjang" Foods 9, no. 7: 915. https://doi.org/10.3390/foods9070915
APA StylePark, Y. K., Jin, Y. H., Lee, J.-H., Byun, B. Y., Lee, J., Jeong, K. C., & Mah, J.-H. (2020). The Role of Enterococcus faecium as a Key Producer and Fermentation Condition as an Influencing Factor in Tyramine Accumulation in Cheonggukjang. Foods, 9(7), 915. https://doi.org/10.3390/foods9070915