
Isolation of DiNP-Degrading Microbes from the Mouse Colon and the Influence DiNP Exposure Has on the Microbiota, Intestinal Integrity, and Immune Status of the Colon

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials & Methods
2.1. Chemicals
2.2. Experimental Animals and Design
2.3. Tissue Collection
2.4. Immunohistology and Analysis
2.5. Anaerobic Diluent and Colon Content Collection
2.6. Isolation and Characterization of DiNP-Degrading Bacteria
2.7. Bacteroides Defined Minimal Media
2.8. Colonic Sample Collection and DNA Extraction for 16S rRNA Sequencing
2.9. 16S Microbiome Analysis
2.10. Identification of DiNP-Degrading Bacteria
2.11. Statistical Analysis
3. Results
3.1. Alpha and Beta Diversity
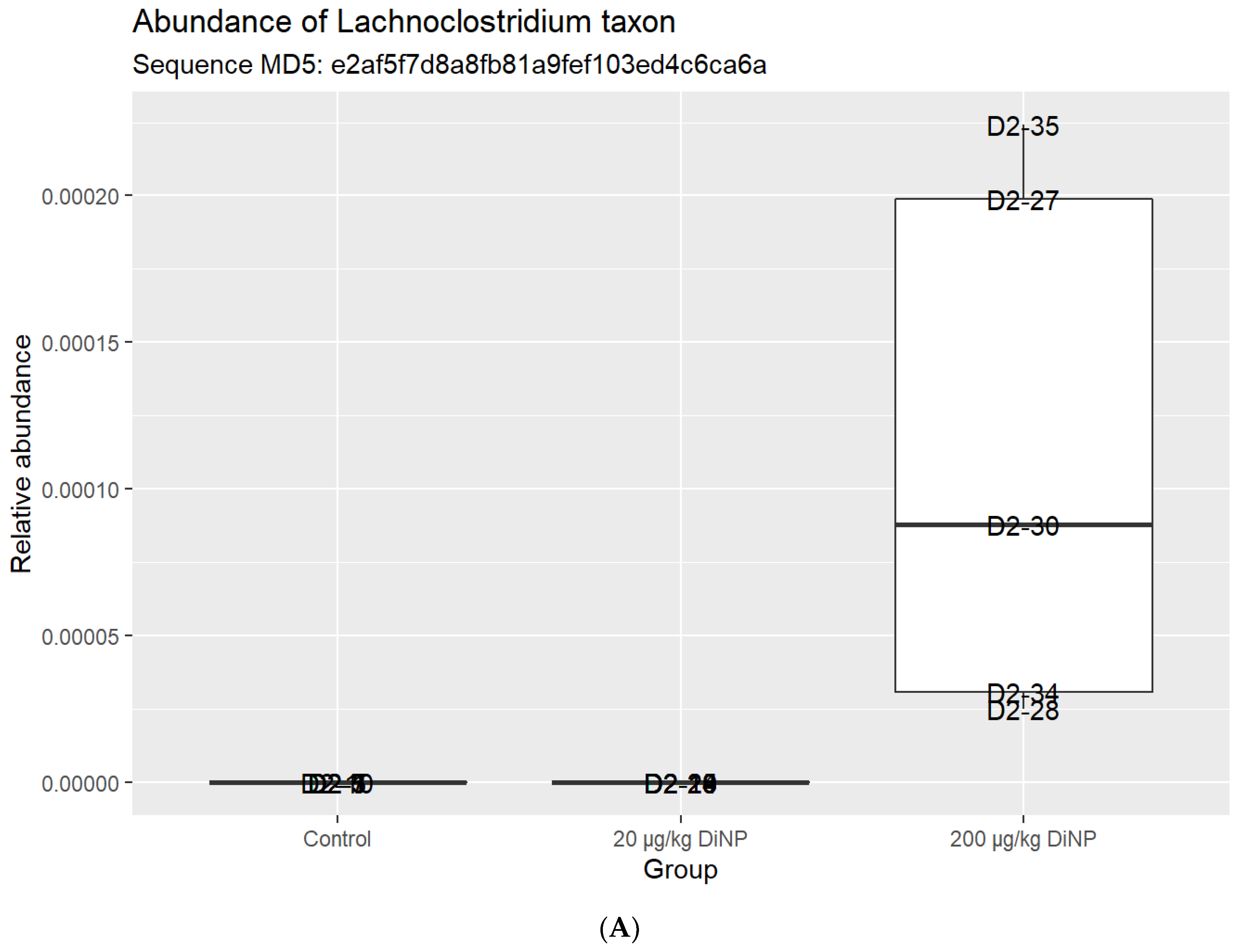
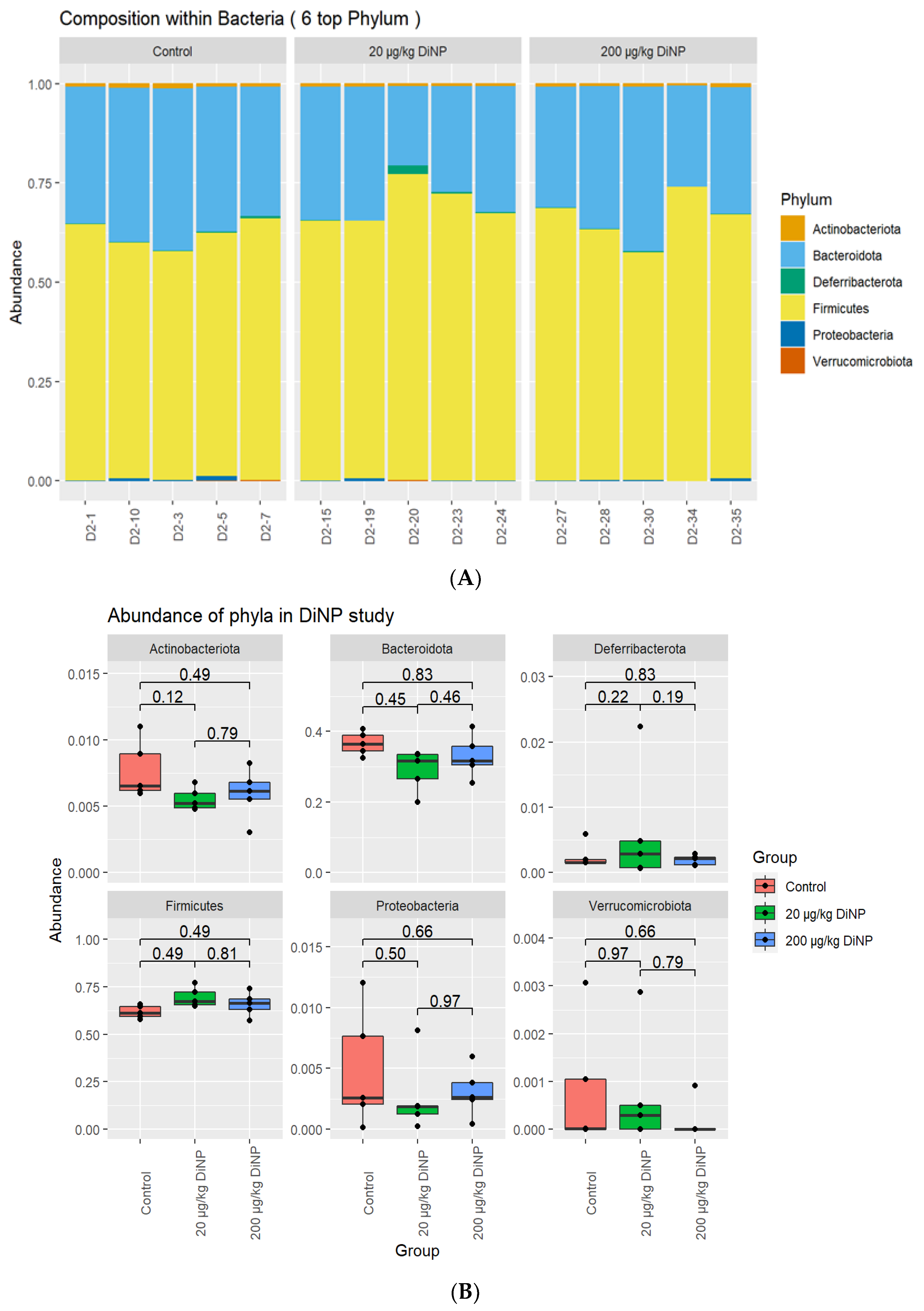
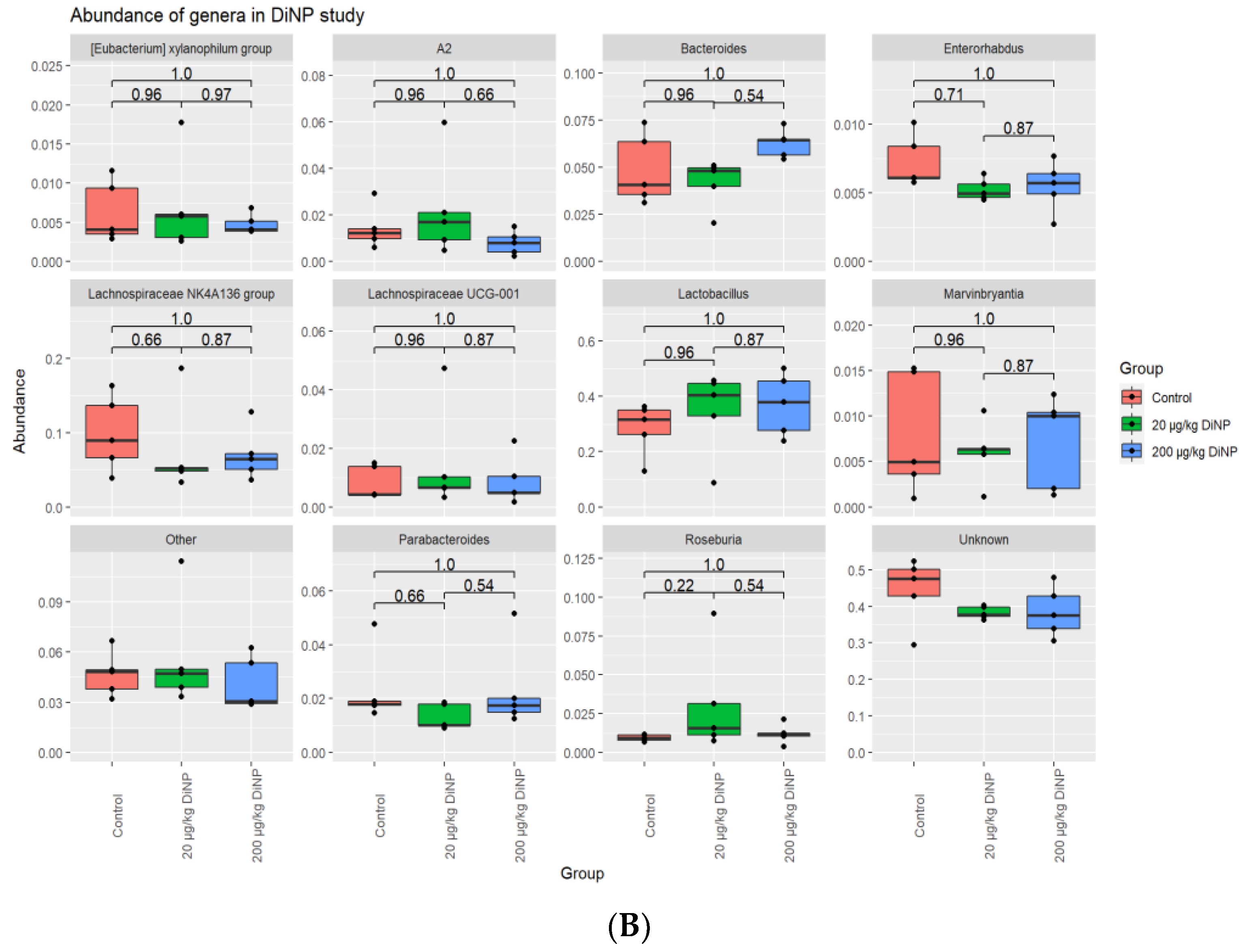
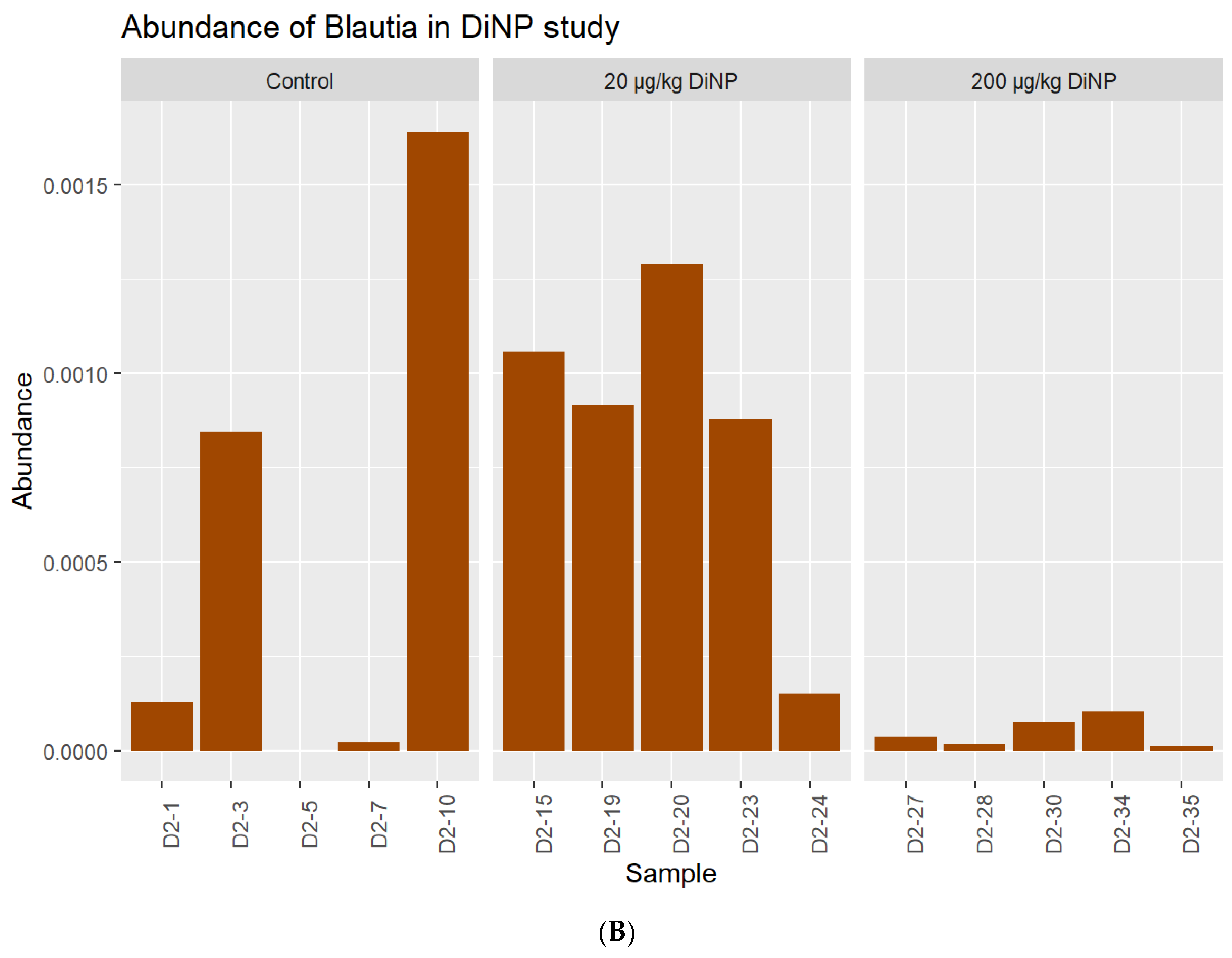
3.2. Taxonomic Results
3.3. DiNP-Degrading Microbes Residing in the Colon
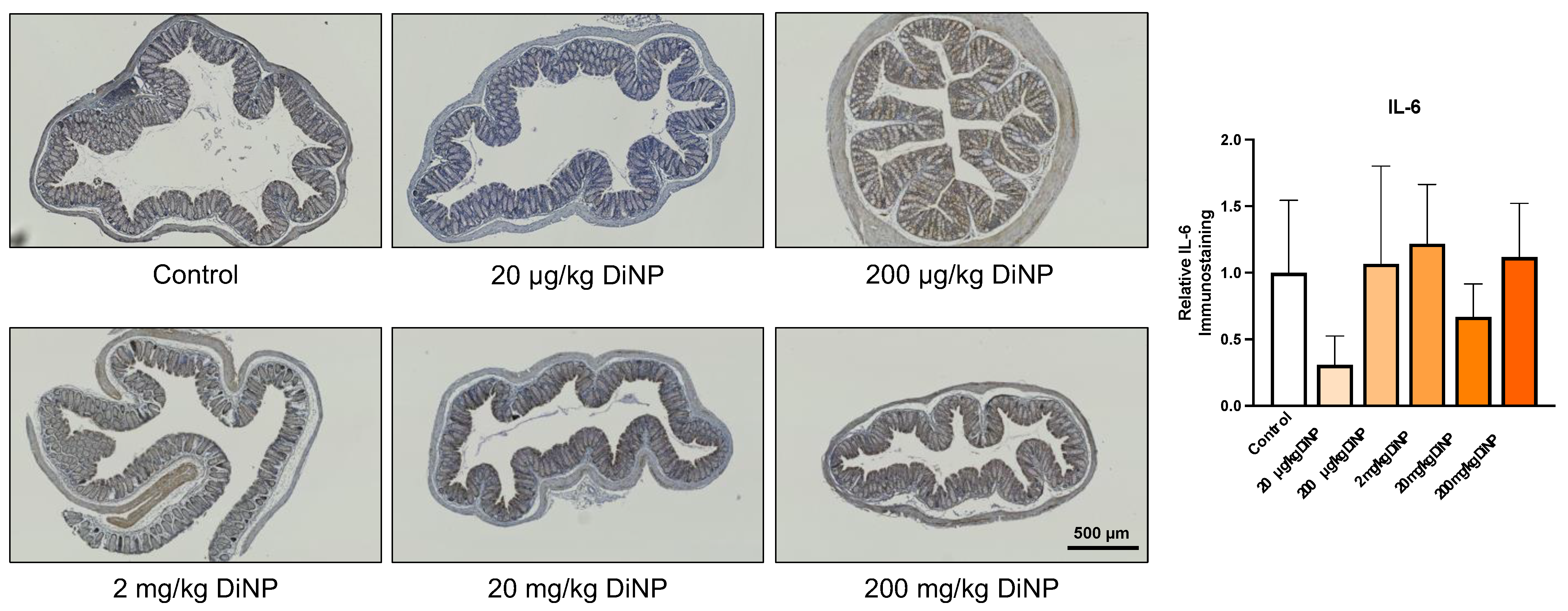
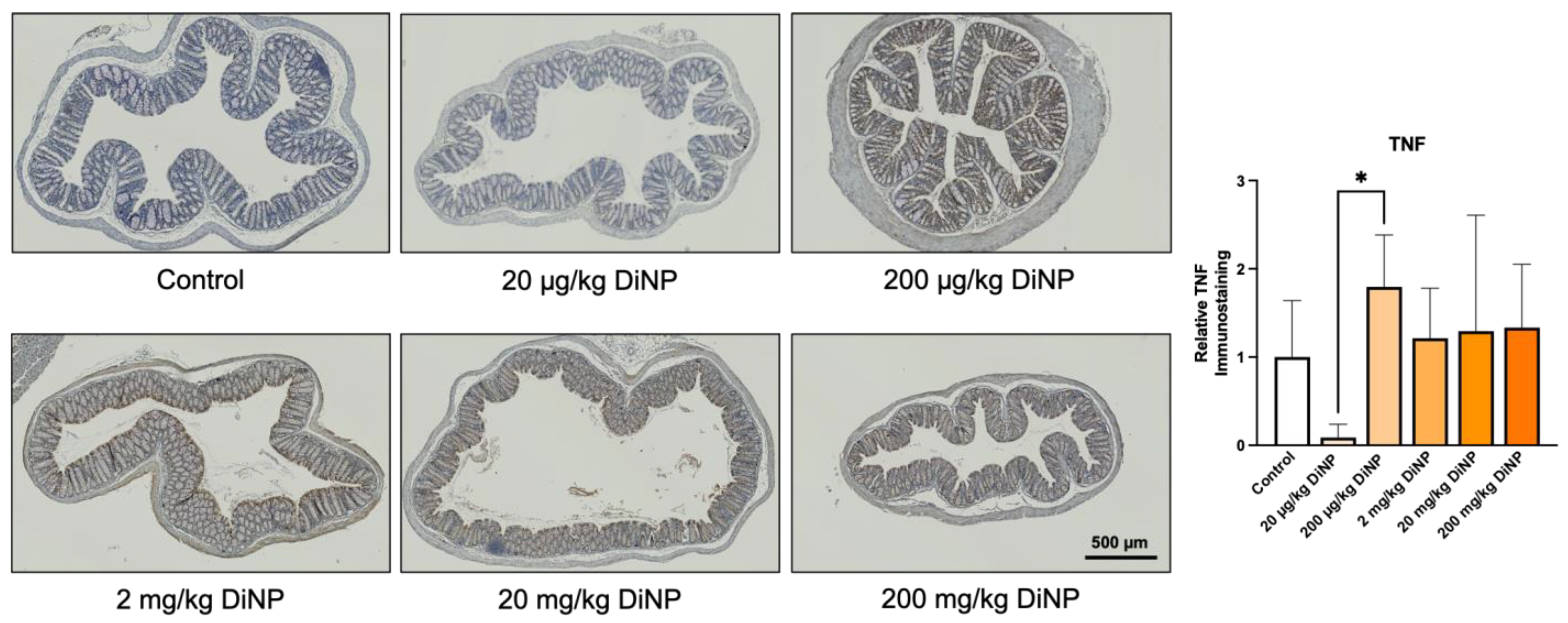
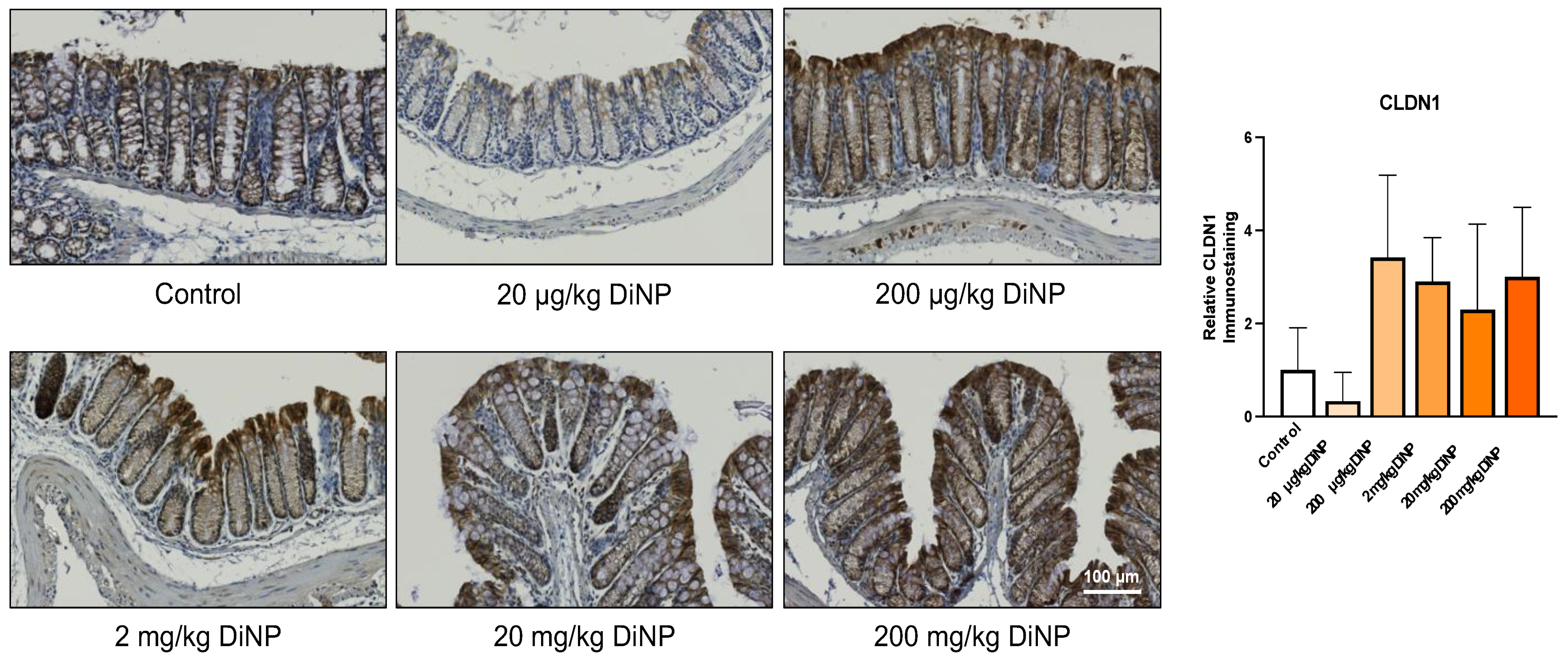
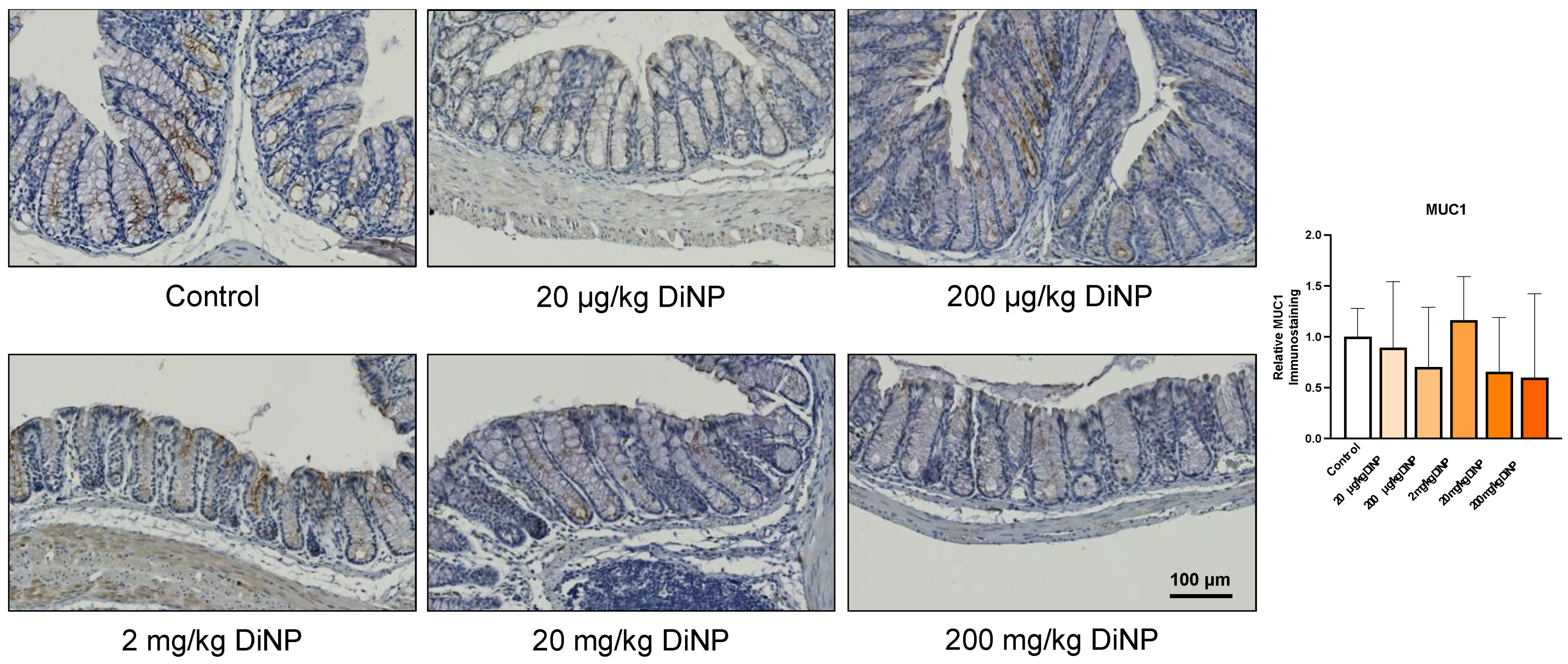
3.4. Immunohistochemistry of Cytokines and Other Factors Playing a Role in the Intestinal Immunology
4. Discussion
5. Conclusions and Future Directions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Neuman, H.; Koren, O. The Gut Microbiome. In Encyclopedia of Cell Biology; Bradshaw, R.A., Stahl, P.D., Eds.; Academic Press: Waltham, MA, USA, 2016; pp. 799–808. [Google Scholar]
- Chiu, K.; Warner, G.; Nowak, R.A.; Flaws, J.A.; Mei, W. The Impact of Environmental Chemicals on the Gut Microbiome. Toxicol. Sci. 2020, 176, 253–284. [Google Scholar] [CrossRef]
- Chiu, K.; Bashir, S.T.; Nowak, R.A.; Mei, W.; Flaws, J.A. Subacute exposure to di-isononyl phthalate alters the morphology, endocrine function, and immune system in the colon of adult female mice. Sci. Rep. 2020, 10, 18788. [Google Scholar] [CrossRef]
- Chiu, K.; Bashir, S.T.; Gao, L.; Gutierrez, J.; de Godoy, M.R.C.; Drnevich, J.; Fields, C.J.; Cann, I.; Flaws, J.A.; Nowak, R.A. Subacute Exposure to an Environmentally Relevant Dose of Di-(2-ethylhexyl) Phthalate during Gestation Alters the Cecal Microbiome, but Not Pregnancy Outcomes in Mice. Toxics 2021, 9, 215. [Google Scholar] [CrossRef] [PubMed]
- Kolb, S.A.; EJO’Loughlin, E.J.; Gsell, T.C. Data on the characterization of phthalate-degrading bacteria from Asian carp microbiomes and riverine sediments. Data Brief 2019, 25, 104375. [Google Scholar] [CrossRef] [PubMed]
- Kolb, S.A.; EJO’Loughlin, E.J.; Gsell, T.C. Characterization of phthalate-degrading bacteria from Asian carp microbiomes and riverine sediments. Int. Biodeterior. Biodegrad. 2019, 143, 104727. [Google Scholar] [CrossRef]
- Lei, M.; Menon, R.; Manteiga, S.; Alden, N.; Hunt, C.; Alaniz, R.C.; Lee, K.; Jayaraman, A. Environmental Chemical Diethylhexyl Phthalate Alters Intestinal Microbiota Community Structure and Metabolite Profile in Mice. mSystems 2019, 4, e00724-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Yue, S.; Hao, Z.; Ren, G.; Lu, D.; Zhang, Q.; Zhao, M. Pubertal exposure to the endocrine disruptor mono-2-ethylhexyl ester at body burden level caused cholesterol imbalance in mice. Environ. Pollut. 2019, 244, 657–666. [Google Scholar] [CrossRef]
- Yang, Y.N.; Yang, Y.S.H.; Lin, I.H.; Chen, Y.Y.; Lin, H.Y.; Wu, C.Y.; Su, Y.T.; Yang, Y.J.; Yang, S.N.; Suen, J.L. Phthalate exposure alters gut microbiota composition and IgM vaccine response in human newborns. Food Chem. Toxicol. 2019, 132, 110700. [Google Scholar] [CrossRef]
- National Research Council (US) Committee on the Health Risks of Phthalates. Phthalates and Cumulative Risk Assessment: The Tasks Ahead; National Academies Press: Washington, DC, USA, 2008.
- Chiang, C.; Lewis, L.R.; Borkowski, G.; Flaws, J.A. Exposure to di(2-ethylhexyl) phthalate and diisononyl phthalate during adulthood disrupts hormones and ovarian folliculogenesis throughout the prime reproductive life of the mouse. Toxicol. Appl. Pharmacol. 2020, 393, 114952. [Google Scholar] [CrossRef]
- Ahlman, H.; Nilsson, O. The Gut as the Largest Endocrine Organ in the Body. Ann. Oncol. 2001, 12 (Suppl. 2), S63–S68. [Google Scholar] [CrossRef]
- Wright, R.J.; Bosch, R.; Gibson, M.I.; Christie-Oleza, J.A. Plasticizer Degradation by Marine Bacterial Isolates: A Proteogenomic and Metabolomic Characterization. Environ. Sci. Technol. 2020, 54, 2244–2256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, S.; Hiraga, K.; Takehana, T.; Taniguchi, I.; Yamaji, H.; Maeda, Y.; Toyohara, K.; Miyamoto, K.; Kimura, Y.; Oda, K. A bacterium that degrades and assimilates poly(ethylene terephthalate). Science 2016, 351, 1196–1199. [Google Scholar] [CrossRef]
- Boll, M.; Geiger, R.; Junghare, M.; Schink, B. Microbial degradation of phthalates: Biochemistry and environmental implications. Environ. Microbiol. Rep. 2020, 12, 3–15. [Google Scholar] [CrossRef]
- Hines, C.J.; Hopf, N.B.; Deddens, J.A.; Silva, M.J.; Calafat, A.M. Occupational Exposure to Diisononyl Phthalate (DiNP) in Polyvinyl Chloride Processing Operations. Int. Arch. Occup. Environ. Health 2012, 85, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Koch, H.M.; Haller, A.; Weiss, T.; Käfferlein, H.U.; Stork, J.; Brüning, T. Phthalate exposure during cold plastisol application—A human biomonitoring study. Toxicol. Lett. 2012, 213, 100–106. [Google Scholar] [CrossRef] [PubMed]
- Porras, S.P.; Hartonen, M.; Koponen, J.; Ylinen, K.; Louhelainen, K.; Tornaeus, J.; Kiviranta, H.; Santonen, T. Occupational Exposure of Plastics Workers to Diisononyl Phthalate (DiNP) and Di(2-propylheptyl) Phthalate (DPHP) in Finland. Int. J. Environ. Res. Public Health 2020, 17, 2035. [Google Scholar] [CrossRef] [Green Version]
- Bogen, K.T.; Boekelheide, K.; Cunningham, M.L.; Jackson, B.A.; Peters, J.M.; Reddy, J.K.; Zeise, L. Report to the U.S. consumer product safety commission by the chronic hazard advisory panel on diisononyl phthalate (dinp). In Chronic Hazard Advisory Panel on Diisononyl Phthalate; U.S.C.P.S. Commission: Bethesda, MD, USA, 2001; p. 20814. [Google Scholar]
- Waterman, S.J.; Ambroso, J.L.; Keller, L.H.; Trimmer, G.W.; Nikiforov, A.I.; Harris, S.B. Developmental Toxicity of Di-Isodecyl and Di-Isononyl Phthalates in Rats. Reprod. Toxicol. 1999, 13, 131–136. [Google Scholar] [CrossRef]
- Gray, L.E., Jr.; Ostby, J.; Furr, J.; Price, M.; Veeramachaneni, D.N.R.; Parks, L. Perinatal Exposure to the Phthalates DEHP, BBP, and DINP, but Not DEP, DMP, or DOTP, Alters Sexual Differentiation of the Male Rat. Toxicol. Sci. 2000, 58, 350–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiang, C.; Flaws, J.A. Subchronic Exposure to Di(2-ethylhexyl) Phthalate and Diisononyl Phthalate during Adulthood Has Immediate and Long-Term Reproductive Consequences in Female Mice. Toxicol. Sci. 2019, 168, 620–631. [Google Scholar] [CrossRef]
- Chiu, K.; Bashir, S.T.; Chiu, J.; Nowak, R.A.; Flaws, J.A. The impact of di-isononyl phthalate exposure on specialized epithelial cells in the colon. Toxicol. Sci. 2021, 184, 142–153. [Google Scholar] [CrossRef] [PubMed]
- McSweeny, C.S.; Denman, S.E.; Mackie, R.I. Rumen bacteria. In Methods in Gut Microbial Ecology for Ruminants; Makkar, H.P.S., McSweeny, C., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 23–38. [Google Scholar]
- Chiang, C.; Pacyga, D.C.; Strakovsky, R.S.; Smith, R.L.; James-Todd, T.; Williams, P.L.; Hauser, R.; Meling, D.D.; Li, Z.; Flaws, J.A. Urinary phthalate metabolite concentrations and serum hormone levels in pre- and perimenopausal women from the Midlife Women’s Health Study. Environ. Int. 2021, 156, 106633. [Google Scholar] [CrossRef] [PubMed]
- Warner, G.R.; Pacyga, D.C.; Strakovsky, R.S.; Smith, R.; James-Todd, T.; Williams, P.L.; Hauser, R.; Meling, D.D.; Li, Z.; Flaws, J.A. Urinary phthalate metabolite concentrations and hot flashes in women from an urban convenience sample of midlife women. Environ. Res. 2021, 197, 110891. [Google Scholar] [CrossRef]
- Varel, V.H.; Bryant, M.P. Nutritional Features of Bacteroides fragilis subsp. fragilis. Appl. Microbiol. 1974, 28, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Bacic, M.K.; Smith, C.J. Laboratory maintenance and cultivation of Bacteroides species. Curr. Protoc. Microbiol. 2008, 9, 13C.1.1–13C.1.21. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Lan, Y.; Wang, Q.; Cole, J.R.; Rosen, G.L. Using the RDP Classifier to Predict Taxonomic Novelty and Reduce the Search Space for Finding Novel Organisms. PLoS ONE 2012, 7, e32491. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2–approximately maximum-likelihood trees for large alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef] [PubMed]
- McMurdie, P.J.; Holmes, S. phyloseq: An R package for reproducible interactive analysis and graphics of microbiome census data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Raikhel, V.; Gopalakrishnan, K.; Fernandez-Hernandez, H.; Lambertini, L.; Manservisi, F.; Falcioni, L.; Bua, L.; Belpoggi, F.; Teitelbaum, S.L.; et al. Effect of postnatal low-dose exposure to environmental chemicals on the gut microbiome in a rodent model. Microbiome 2016, 4, 26. [Google Scholar] [CrossRef] [Green Version]
- Witjes, V.M.; Boleij, A.; Halffman, W. Reducing versus Embracing Variation as Strategies for Reproducibility: The Microbiome of Laboratory Mice. Animals 2020, 10, 2415. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.Q.; Li, T.; Nakatsu, G.; Chen, Y.X.; Yau, T.O.; Chu, E.; Wong, S.; Szeto, C.H.; Ng, S.C.; Chan, F.K.L.; et al. A novel faecal Lachnoclostridium marker for the non-invasive diagnosis of colorectal adenoma and cancer. Gut 2020, 69, 1248–1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yutin, N.; Galperin, M.Y. A genomic update on clostridial phylogeny: Gram-negative spore formers and other misplaced clostridia. Environ. Microbiol. 2013, 15, 2631–2641. [Google Scholar] [CrossRef] [PubMed]
- Dandachi, I.; Anani, H.; Hadjadj, L.; Brahimi, S.; Lagier, J.C.; Daoud, Z.; Rolain, J.M. Genome analysis of Lachnoclostridium phocaeense isolated from a patient after kidney transplantation in Marseille. New Microbes New Infect. 2021, 41, 100863. [Google Scholar] [CrossRef]
- Momose, Y.; Maruyama, A.; Iwasaki, T.; Miyamoto, Y.; Itoh, K. 16S rRNA gene sequence-based analysis of clostridia related to conversion of germfree mice to the normal state. J. Appl. Microbiol. 2009, 107, 2088–2097. [Google Scholar] [CrossRef] [PubMed]
- Atarashi, K.; Tanoue, T.; Oshima, K.; Suda, W.; Nagano, Y.; Nishikawa, H.; Fukuda, S.; Saito, T.; Narushima, S.; Hase, K.; et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 2013, 500, 232–236. [Google Scholar] [CrossRef]
- Li, J.; Adams, V.; Bannam, T.L.; Miyamoto, K.; Garcia, J.P.; Uzal, F.A.; Rood, J.I.; McClane, B.A. Toxin plasmids of Clostridium perfringens. Microbiol. Mol. Biol. Rev. 2013, 77, 208–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Songer, J.G. Clostridia as agents of zoonotic disease. Vet. Microbiol. 2010, 140, 399–404. [Google Scholar] [CrossRef]
- Ozato, N.; Saito, S.; Yamaguchi, T.; Katashima, M.; Tokuda, I.; Sawada, K.; Katsuragi, Y.; Kakuta, M.; Imoto, S.; Ihara, K.; et al. Blautia genus associated with visceral fat accumulation in adults 20–76 years of age. NPJ Biofilms Microbiomes 2019, 5, 28. [Google Scholar] [CrossRef] [Green Version]
- Gurung, M.; Li, Z.; You, H.; Rodrigues, R.; Jump, D.B.; Morgun, A.; Shulzhenko, N. Role of gut microbiota in type 2 diabetes pathophysiology. EbioMedicine 2020, 51, 102590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenq, R.R.; Taur, Y.; Devlin, S.M.; Ponce, D.M.; Goldberg, J.D.; Ahr, K.F.; Littmann, E.R.; Ling, L.; Gobourne, A.C.; Miller, L.C.; et al. Intestinal Blautia Is Associated with Reduced Death from Graft-versus-Host Disease. Biol. Blood Marrow Transplant. 2015, 21, 1373–1383. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Anaerobic Diluent (Modified from C.S. McSweeney et al., 2005) | |
|---|---|
| 8% Na2CO3 solution | 5 mL |
| Solution No. 1 (See below) | 3.8 mL |
| Solution No. 2 (See below) | 3.8 mL |
| 0.1% Resazurin | 0.1 mL |
| ddH2O | 87.3 mL |
| Total Volume | 100 mL |
| Solution No. 1—g/L in dH2O | |
| KH2PO4 | 6 g |
| Solution No. 2—g/L in dH2O | |
| CaCl2–2H2O | 1.6 g |
| KH2PO4 | 6 g |
| NaCl | 12 g |
| (NH4)2SO4 | 6 g |
| MgSO4–7H2O | 2.5 g |
| Bacteroides Defined Minimal Media (Modified from Bacic and Smith, 2013) | |
|---|---|
| Mineral 3B solution | 50 mL/L |
| Cysteine hydrochloride | 1 g/L |
| Hemin solution | 10 mL/L |
| 0.01% vitamin B12 | 1 mL/L |
| 10% DiNP solution | 10 mL/L |
| Iron (II) solute (FeSO4) solution | 1.5 mL/L |
| 7% NaHCO3 | 14.4 mL/L |
| 0.1% resazurin | 1 mL/L |
| Distilled water | Add to make 1 L |
| Mineral 3B solution—g/L in dH2O | |
| KH2PO4 | 18 g/L |
| NaCl | 18 g/L |
| MgCl2•6H2O | 0.4 g/L |
| CaCl2•2H2O | 0.52 g/L |
| CoCl2•6H2O | 0.02 g/L |
| MnCl2•4H2O | 0.20 g/L |
| NH4Cl | 10 g/L |
| Na2SO4 | 5 g/L |
| Study | Variable | Bray | UniFrac | Weighted UniFrac |
|---|---|---|---|---|
| DiNP | Group | 0.787 | 0.599 | 0.588 |
| DiNP | Library Size | 0.328 | 0.569 | 0.712 |
| DiNP20 | Group | 0.695 | 0.752 | 0.338 |
| DiNP20 | Library Size | 0.340 | 0.753 | 0.605 |
| DiNP200 | Group | 0.520 | 0.781 | 0.386 |
| DiNP200 | Library Size | 0.612 | 0.206 | 0.727 |
| Genus | Adjusted p-Value | ||
|---|---|---|---|
| Control vs. 20 µg/kg DiNP | Control vs. 200 µg/kg DiNP | 20 µg/kg DiNP vs. 200 µg/kg DiNP | |
| GCA-900066575 | 0.9579 | 0.9965 | 0.873 |
| Lachnospiraceae UCG-006 | 0.9253 | 0.9965 | 0.5428 |
| Lachnoclostridium | 0.9279 | 0.9965 | 0.6572 |
| [Eubacterium] ventriosum group | 0.9579 | 0.9655 | 0.6572 |
| Lachnospiraceae FCS020 group | 0.9579 | 0.9965 | 0.873 |
| Roseburia | 0.2213 | 0 9965 | 0.5428 |
| Blautia | 0.9579 | 0.1565 | 0.0514 |
| Acetatifactor | 0.9579 | 0.9965 | 0.873 |
| Lachnospiraceae NK4A136 group | 0.6583 | 0.9965 | 0.873 |
| Marvinbrvantia | 0.9579 | 0.9965 | 0.873 |
| Lachnospiraceae UCG-001 | 0.9579 | 0.9965 | 0.873 |
| A2 | 0.9579 | 0.9965 | 0.6572 |
| [Eubacterium] xylanophilum group | 0.9579 | 0.9965 | 9713 |
| DefluviitaleaceaeUCG-011 | 0.9579 | 0.9965 | 0.873 |
| Tyzzerella | 0.9579 | 0.9965 | 0.6572 |
| Tuzzerella | 0.9579 | 0.9983 | 0.873 |
| ASF356 | 0.9579 | 0.9965 | 0.873 |
| Lachnospiraceae UCG-010 | 0.9579 | 0.9965 | 0.9103 |
| Christensenella | 0.9579 | 0.9965 | 0.873 |
| Christensenellaceae R-7 group | 0 9579 | 0 9965 | 0.873 |
| Monoglobus | 0.6583 | 0.9965 | 0.5428 |
| Oscillibacter | 0.9579 | 0.9965 | 0.873 |
| Colidextribacter | 0.7136 | 0 9965 | 0.873 |
| Intestinimonas | 0.7084 | 0.9965 | 0.8842 |
| NK4A214 group | 0.9579 | 0.9965 | 0.873 |
| Papillibacter | 0.9579 | 0.9965 | 0.873 |
| UCG-005 | 0.6583 | 0.9965 | 0.6522 |
| UCG-009 | 0.9579 | 0.9965 | 0.873 |
| Butyricicoccus | 0.7136 | 0.9965 | 0.873 |
| Incertae Sedis | 0 9579 | 0.9965 | 0.6572 |
| Ruminococcus | 0.9579 | 0.3723 | 0.5428 |
| [Eubacterium] siraeum group | 0.9579 | 0 9965 | 0.9713 |
| Anaerotruncus | 0.6583 | 0.9965 | 0.6047 |
| Negativibacillus | 0.9579 | 0.9965 | 0.873 |
| Harryflintia | 0.9579 | 0.9965 | 0.873 |
| Paludicola | 9579 | 0.9965 | 0.98 |
| Candidatus Arthromitus | 0.9579 | 0.9965 | 0.873 |
| Anaerofustis | 0.9579 | 0.9965 | 0.9713 |
| Anaerovorax | 0.981 | 0.9965 | 0.9277 |
| [Eubacterium] nodatum group | 0.9579 | 0.9965 | 0.873 |
| Family XIlI AD3011 group | 0.9579 | 0.9965 | 0.873 |
| [Eubacterium] brachy group | 0.6583 | 0.9965 | 0.6572 |
| Family XIII UCG-001 | 0.9579 | 0.9965 | 0 8730 |
| Lactobacillus | 0.9579 | 0.9965 | 0.873 |
| Erysipelatoclostridium | 0.7136 | 0.3782 | 0.873 |
| Enterorhabdus | 7136 | 0.9965 | 0.873 |
| Parvibacter | 0.7169 | 0.9965 | 0.5428 |
| Parasutterella | 0.7367 | 0.9965 | 0.873 |
| Mucispirillum | 0.7136 | 0 9965 | 0.6047 |
| Akkermansia | 0.9579 | 0.9965 | 0.873 |
| Alistipes | 0.9579 | 0.9965 | 0.873 |
| Parabacteroides | 0.6583 | 0.9965 | 0.5428 |
| Bacteroides | 0.9579 | 0.9965 | 0.5428 |
| Control vs. 20 µg/kg DiNP | Control vs. 200 µg/kg DiNP | 20 vs. 200 µg/kg DiNP | |
|---|---|---|---|
| Up | 0 | 1 | 3 |
| Non-significant | 430 | 429 | 425 |
| Down | 2 | 2 | 4 |
| DiNP-Degrading Bacteria Isolated from the Colon | |||
|---|---|---|---|
| Species | Coverage (%) | Identity (%), 27F Primer | Identity (%), 1492R Primer |
| Proteus mirabilis strain ATCC 29,906 | 96 | 95.32 | 96.80 |
| Desulfitobacterium hafniense DCB-2 | 89 | 92.41 | 99.07 |
| Paenibacillus barengoltzii strain NBRC 101,215 | 98 | 95.97 | 97.94 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiu, K.K.; Bashir, S.T.; Abdel-Hamid, A.M.; Clark, L.V.; Laws, M.J.; Cann, I.; Nowak, R.A.; Flaws, J.A. Isolation of DiNP-Degrading Microbes from the Mouse Colon and the Influence DiNP Exposure Has on the Microbiota, Intestinal Integrity, and Immune Status of the Colon. Toxics 2022, 10, 75. https://doi.org/10.3390/toxics10020075
Chiu KK, Bashir ST, Abdel-Hamid AM, Clark LV, Laws MJ, Cann I, Nowak RA, Flaws JA. Isolation of DiNP-Degrading Microbes from the Mouse Colon and the Influence DiNP Exposure Has on the Microbiota, Intestinal Integrity, and Immune Status of the Colon. Toxics. 2022; 10(2):75. https://doi.org/10.3390/toxics10020075
Chicago/Turabian StyleChiu, Karen K., Shah Tauseef Bashir, Ahmed M. Abdel-Hamid, Lindsay V. Clark, Mary J. Laws, Isaac Cann, Romana A. Nowak, and Jodi A. Flaws. 2022. "Isolation of DiNP-Degrading Microbes from the Mouse Colon and the Influence DiNP Exposure Has on the Microbiota, Intestinal Integrity, and Immune Status of the Colon" Toxics 10, no. 2: 75. https://doi.org/10.3390/toxics10020075