
Biochemical and Metabolomic Responses of Antarctic Bacterium Planococcus sp. O5 Induced by Copper Ion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture
2.2. MIC Determination of Heavy Metals
2.3. Measurements of Electrical Conductivity and Biomass
2.4. Measurements of Antioxidant System
2.5. GC-MS analysis of Metabolites
2.5.1. Sample Preparation
2.5.2. GC-MS Analysis of Metabolites
2.5.3. Data Processing
2.5.4. Statistical Analysis
3. Results
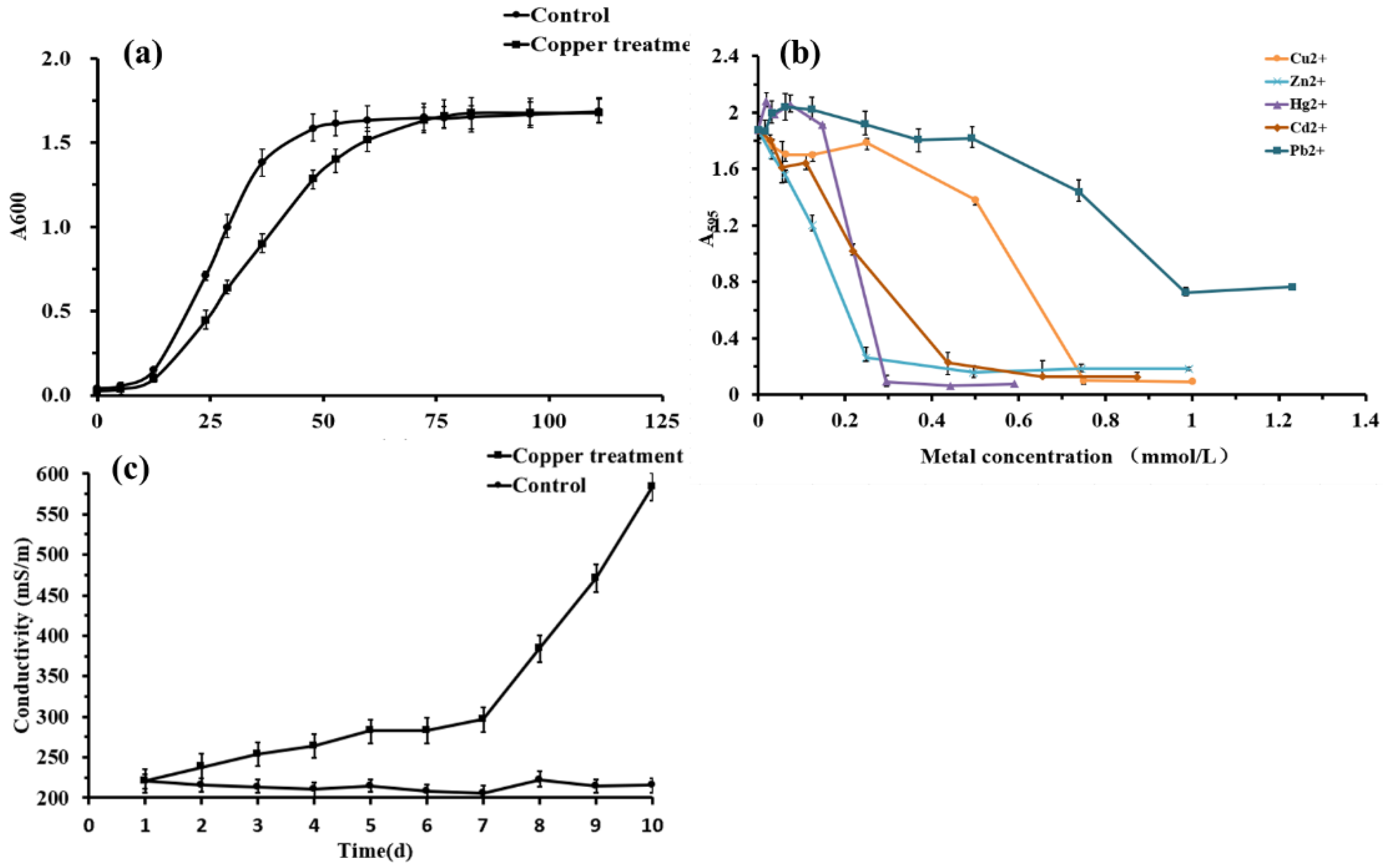
3.1. Heavy Metals Resistance Analysis
3.2. Change in the Membrane Permeability
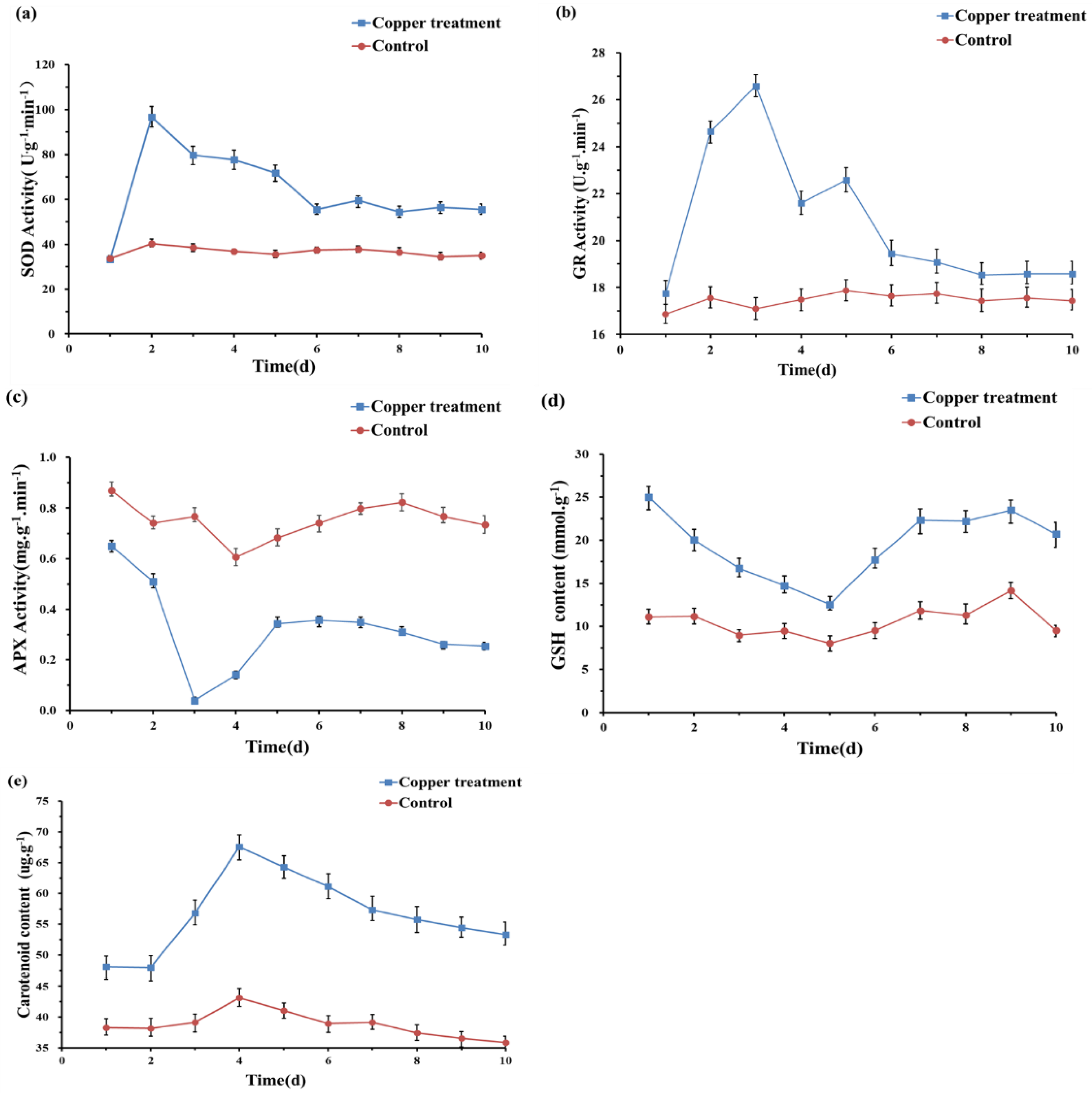
3.3. Response of the Antioxidant System
3.4. Metabolic Response of Strain O5 to Cu Induction
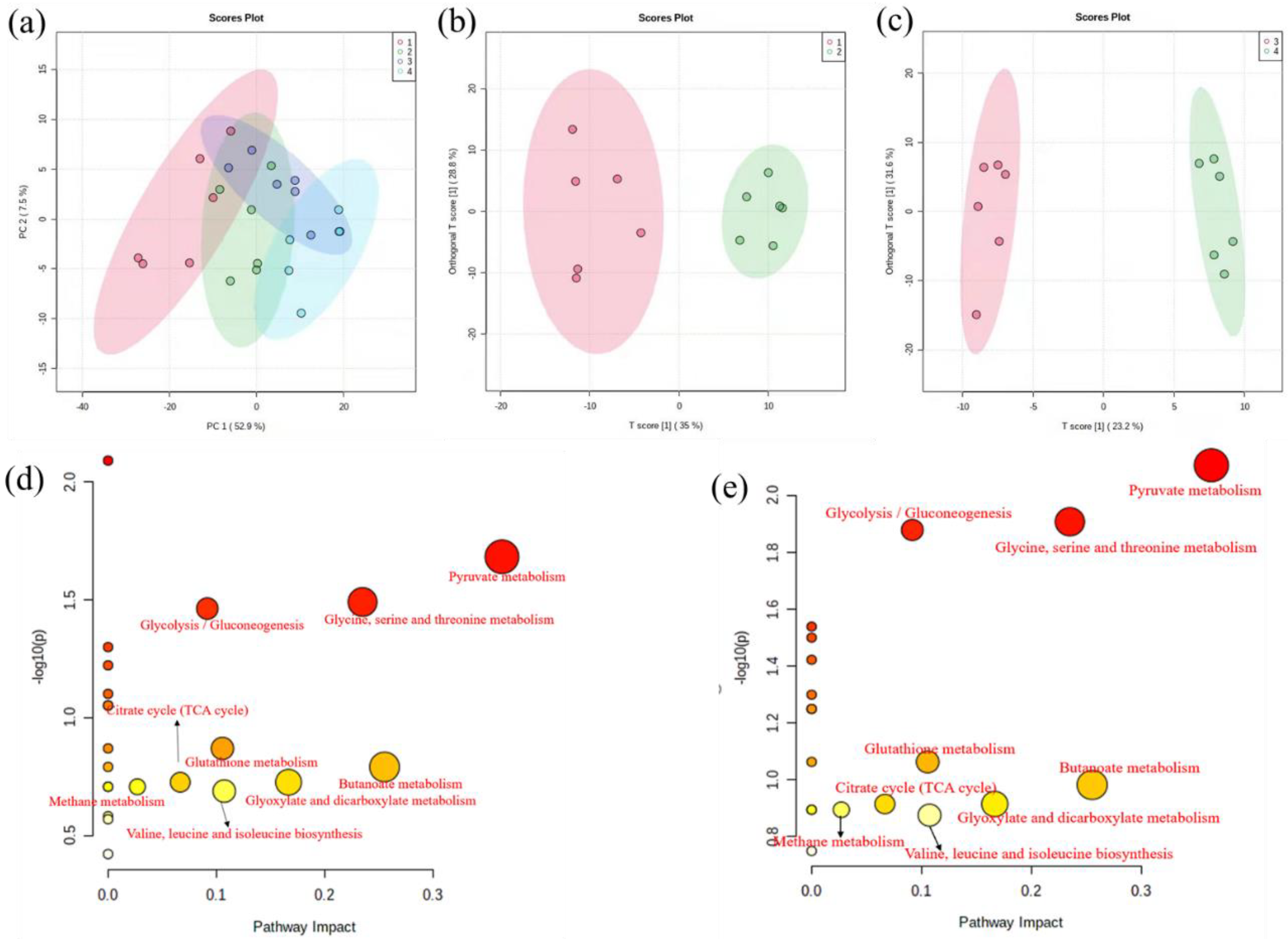
3.4.1. Metabolic Profile Analysis
3.4.2. Identification and Analysis of Differential Metabolites
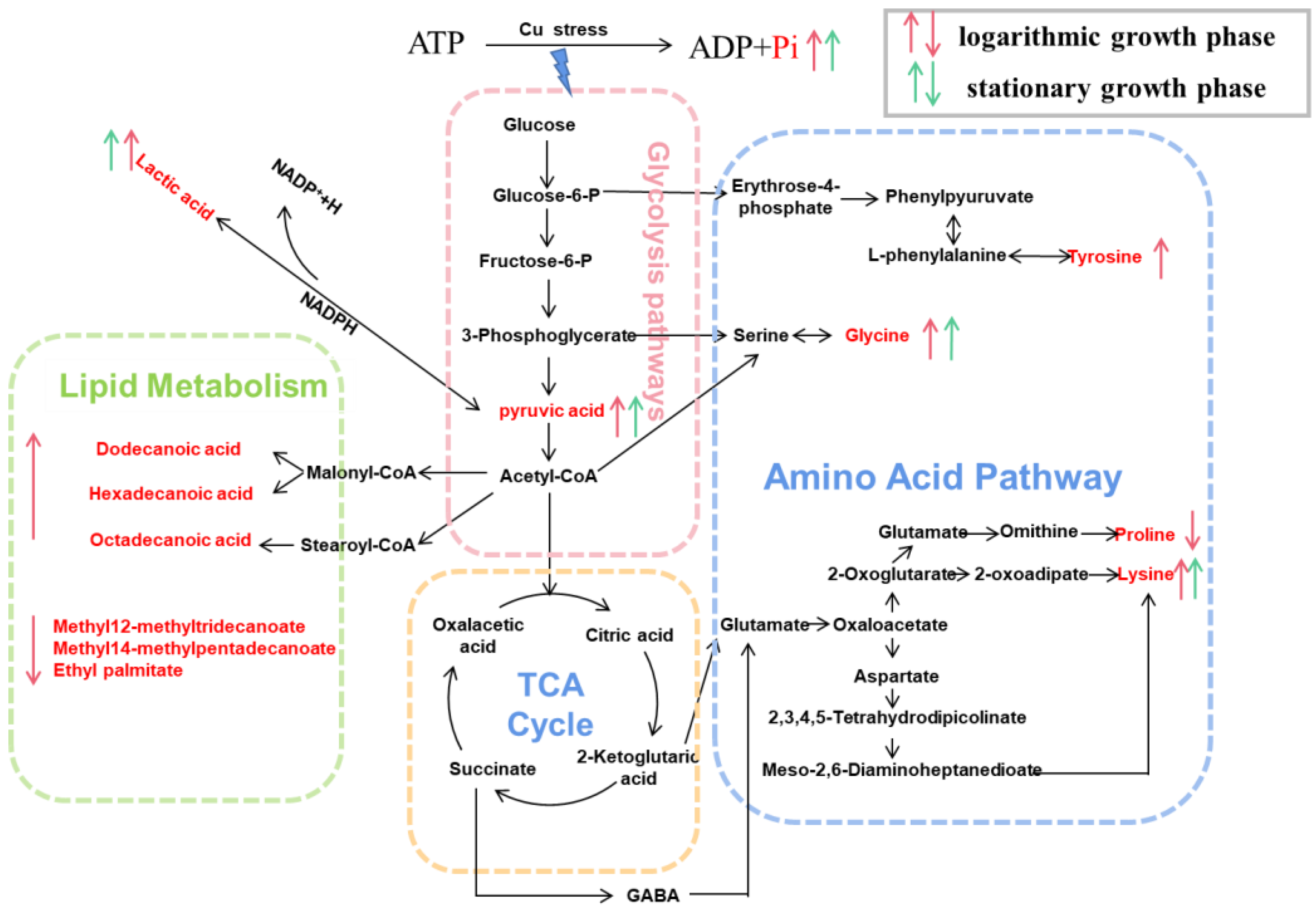
3.4.3. Perturbed Biological Pathway Responded to Copper Stress
4. Discussion
4.1. Heavy Metals Resistance
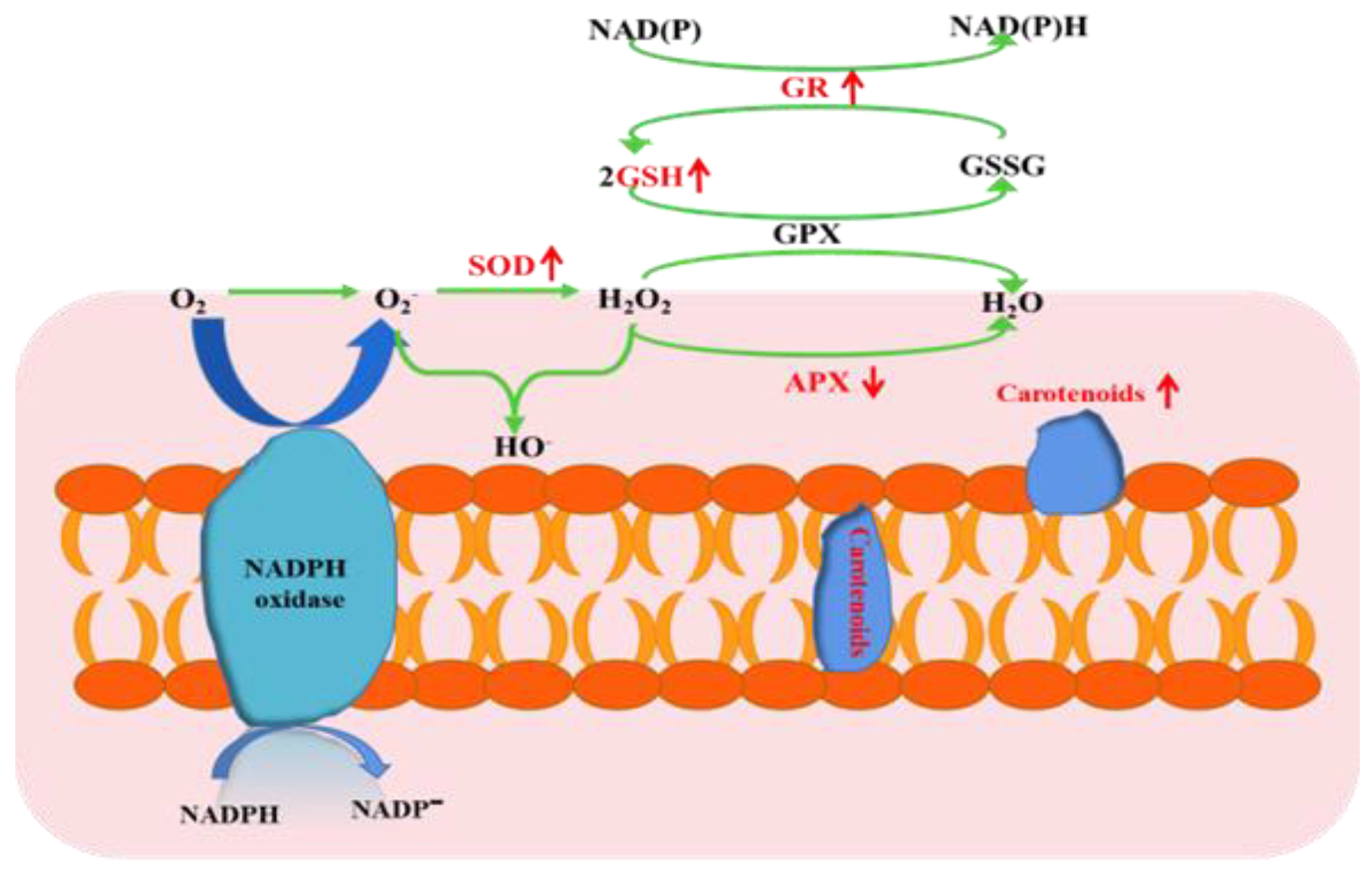
4.2. Effect on Redox Status
4.3. Metabolic Reprogramming
4.3.1. Energy Metabolism
4.3.2. Amino Acid Metabolism
4.3.3. Lipid Metabolism
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaur, N.; Flora, G.; Yadav, M.; Tiwari, A. A review with recent advancements on bioremediation-based abolition of heavy metals. Environ. Sci. Process. Impacts 2014, 16, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Malidareh, H.B.; Mahvi, A.; Yunesian, M.; Alimohammadi, M.; Nazmara, S. Effect of fertilizer application on paddy soil heavy metals concentration and groundwater in North of Iran. Middle-East J. Sci. Res. 2014, 20, 1721–1727. [Google Scholar]
- Zeraatkar, A.K.; Ahmadzadeh, H.; Talebi, A.F.; Moheimani, N.R.; McHenry, M.P. Potential use of algae for heavy metal bioremediation, a critical review. J. Environ. Manag. 2016, 181, 817–831. [Google Scholar] [CrossRef] [PubMed]
- Lemire, J.A.; Harrison, J.J.; Turner, R.J. Antimicrobial activity of metals: Mechanisms, molecular targets and applications. Nat. Rev. Microbiol. 2013, 11, 371–384. [Google Scholar] [CrossRef]
- Pal, A.; Bhattacharjee, S.; Saha, J.; Sarkar, M.; Mandal, P. Bacterial survival strategies and responses under heavy metal stress: A comprehensive overview. Crit. Rev. Microbiol. 2022, 48, 327–355. [Google Scholar] [CrossRef]
- Sogin, E.M.; Leisch, N.; Dubilier, N. Chemosynthetic symbioses. Curr. Biol. 2020, 30, R1137–R1142. [Google Scholar] [CrossRef] [PubMed]
- Hao, X.; Zhu, J.; Rensing, C.; Liu, Y.; Gao, S.; Chen, W.; Huang, Q.; Liu, Y.R. Recent advances in exploring the heavy metal(loid) resistant microbiome. Comput. Struct. Biotechnol. J. 2021, 19, 94–109. [Google Scholar] [CrossRef]
- Nanda, M.; Kumar, V.; Sharma, D.K. Multimetal tolerance mechanisms in bacteria: The resistance strategies acquired by bacteria that can be exploited to ‘clean-up’ heavy metal contaminants from water. Aquat. Toxicol. 2019, 212, 1–10. [Google Scholar] [CrossRef]
- MacLean, A.; Bley, A.M.; Appanna, V.P.; Appanna, V.D. Metabolic manipulation by Pseudomonas fluorescens: A powerful stratagem against oxidative and mental stress. J. Med. Microbiol. 2020, 69, 339–346. [Google Scholar] [CrossRef]
- Jurkiewicz, P.; Olzynska, A.; Cwiklik, L.; Conte, E.; Jungwirth, P.; Megli, F.M.; Hof, M. Biophysics of lipid bilayers containing oxidatively modified phospholipids: Insights from fluorescence and EPR experiments and from MD simulations. Biochim. Biophys. Acta 2012, 1818, 2388–2402. [Google Scholar] [CrossRef] [Green Version]
- Alhasawi, A.; Costanzi, J.; Auger, C.; Appanna, N.D.; Appanna, V.D. Metabolic reconfigurations aimed. at the detoxification of a multi-mental stress in Pseudomonas fluorescens: Implications for the bioremediation of metal pollutants. J. Biotechnol. 2015, 200, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Bouskill, N.J.; Barker-Finkel, J.; Galloway, T.S.; Handy, R.D.; Ford, T.E. Temporal bacterial diversity associated with metal-contaminated river sediments. Ecotoxicology 2010, 19, 317–328. [Google Scholar] [CrossRef]
- Fakhar, A.; Gul, B.; Gurmani, A.R.; Khan, S.M.; Ali, S.; Sultan, T.; Chaudhary, H.J.; Rafique, M.; Rizwan, M. Heavy metal remediation and resistance mechanism of Aeromonas, Bacillus, and Pseudomonas: A review. Crit. Rev. Environ. Sci. Technol. 2022, 52, 1868–1914. [Google Scholar] [CrossRef]
- Murinova, S.; Dercova, K. Response mechanisms of bacterial degraders to environmental contaminants on the level of cell walls and cytoplasmic membrane. Int. J. Microbiol. 2014, 2014, 873081. [Google Scholar] [CrossRef] [Green Version]
- Dave, D.; Sarma, S.; Parmar, P.; Shukla, A.; Goswami, D.; Shukla, A.; Saraf, M. Microbes as a boon for the bane of heavy metals. Environ. Sustain. 2020, 3, 233–255. [Google Scholar] [CrossRef]
- Alvarez, S.; Jerez, C.A. Copper ions stimulate polyphosphate degradation and phosphate efflux in Acidithiobacillus ferrooxidans. Appl. Environ. Microbiol. 2004, 70, 5177–5182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulakovskaya, T. Inorganic polyphosphates and heavy metal resistance in microorganisms. World J. Microbiol. Biotechnol. 2018, 34, 139. [Google Scholar] [CrossRef] [PubMed]
- Merroun, M.L.; Raff, J.; Rossberg, A.; Hennig, C.; Reich, T.; Selenska-Pobell, S. Complexation of uranium by cells and S-layer sheets of Bacillus sphaericus JG-A12. Appl. Environ. Microbiol. 2005, 71, 5532–5543. [Google Scholar] [CrossRef] [Green Version]
- Grillo-Puertas, M.; Schurig-Briccio, L.A.; Rodriguez-Montelongo, L.; Rintoul, M.R.; Rapisarda, V.A. Copper tolerance mediated by polyphosphate degradation and low-affinity inorganic phosphate transport system in Escherichia coli. BMC Microbiol. 2014, 14, 72. [Google Scholar] [CrossRef] [Green Version]
- Mailloux, R.J.; Lemire, J.; Kalyuzhnyi, S.; Appanna, V. A novel metabolic network leads to enhanced citrate biogenesis in Pseudomonas fluorescens exposed to aluminum toxicity. Extremophiles 2008, 12, 451–459. [Google Scholar] [CrossRef]
- Yin, K.; Wang, Q.; Lv, M.; Chen, L. Microorganism remediation strategies towards heavy metals. Chem. Eng. J. 2019, 360, 1553–1563. [Google Scholar] [CrossRef]
- Ferreira, P.A.A.; Bomfeti, C.A.; Soares, C.R.F.d.S.; Soares, B.L.; Moreira, F.M.d.S. Cupriavidus necator strains: Zinc and cadmium tolerance and bioaccumulation. Sci. Agric. 2018, 75, 452–460. [Google Scholar] [CrossRef]
- Ghavidel, A.; Naji Rad, S.; Alikhani, H.A.; Sharari, M.; Ghanbari, A. Bioleaching of heavy metals from sewage sludge, direct action of Acidithiobacillus ferrooxidans or only the impact of pH? J. Mater. Cycles Waste Manag. 2017, 20, 1179–1187. [Google Scholar] [CrossRef]
- Eggers, S.; Safdar, N.; Sethi, A.K.; Suen, G.; Peppard, P.E.; Kates, A.E.; Skarlupka, J.H.; Kanarek, M.; Malecki, K.M.C. Urinary lead concentration and composition of the adult gut microbiota in a cross-sectional population-based sample. Environ. Int. 2019, 133 Pt A, 105122. [Google Scholar] [CrossRef] [PubMed]
- Mangal, V.; Nguyen, T.Q.; Fiering, Q.; Gueguen, C. An untargeted metabolomics approach for the putative characterization of metabolites from Scenedesmus obliquus in response to cadmium stress. Environ. Pollut. 2020, 266 Pt 2, 115123. [Google Scholar] [CrossRef] [PubMed]
- Booth, S.C.; Weljie, A.M.; Turner, R.J. Metabolomics reveals differences of metal toxicity in cultures of Pseudomonas pseudoalcaligenes KF707 grown on different carbon sources. Front. Microbiol. 2015, 6, 827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bargagli, R.; Agnorelli, C.; Borghini, F.; Monaci, F. Enhanced deposition and bioaccumulation of mercury in Antarctic terrestrial ecosystems facing a coastal polynya. Environ. Sci. Technol. 2005, 39, 8150–8185. [Google Scholar] [CrossRef]
- Lo Giudice, A.; Casella, P.; Bruni, V.; Michaud, L. Response of bacterial isolates from Antarctic shallow sediments towards heavy metals, antibiotics and polychlorinated biphenyls. Ecotoxicology 2013, 22, 240–250. [Google Scholar] [CrossRef]
- Bargagli, R. Trace metals in Antarctica related to climate change and increasing human impact. Rev. Environ. Contam. Toxicol. 2000, 166, 129–173. [Google Scholar]
- Chu, W.L.; Dang, N.L.; Kok, Y.Y.; Yap, K.S.I.; Phang, S.M.; Convey, P. Heavy metal pollution in Antarctica and its potential impacts on algae. Polar Sci. 2019, 20, 75–83. [Google Scholar] [CrossRef]
- Puasa, N.A.; Zulkharnain, A.; Verasoundarapandian, G.; Wong, C.Y.; Zahri, K.N.M.; Merican, F.; Shaharuddin, N.A.; Gomez-Fuentes, C.; Ahmad, S.A. Effects of Diesel, Heavy Metals and Plastics Pollution on Penguins in Antarctica: A Review. Animals 2021, 11, 2505. [Google Scholar] [CrossRef] [PubMed]
- Ravet, K.; Pilon, M. Copper and iron homeostasis in plants: The challenges of oxidative stress. Antioxid. Redox Signal. 2013, 19, 919–932. [Google Scholar] [CrossRef] [Green Version]
- Baker, J.; Sitthisak, S.; Sengupta, M.; Johnson, M.; Jayaswal, R.K.; Morrissey, J.A. Copper stress induces a global stress response in Staphylococcus aureus and represses sae and agr expression and biofilm formation. Appl. Environ. Microbiol. 2010, 76, 150–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barwinska-Sendra, A.; Waldron, K.J. The Role of Intermetal Competition and Mis-Metalation in Metal Toxicity. Adv. Microb. Physiol. 2017, 70, 315–379. [Google Scholar] [PubMed]
- Sfiligoj, B.J.; King, C.K.; Candy, S.G.; Mondon, J.A. Determining the sensitivity of the Antarctic amphipod Orchomenella pinguides to metals using a joint. model of survival response to exposure concentration and duration. Ecotoxicology 2015, 24, 583–594. [Google Scholar] [CrossRef] [PubMed]
- Chapman, P.M.; Riddle, M.J. Toxic effects of contaminants in polar marine environments. Environ. Sci. Technol. 2005, 39, 200A–207A. [Google Scholar] [CrossRef] [Green Version]
- Fenice, M. The Psychrotolerant Antarctic Fungus Lecanicillium muscarium CCFEE 5003: A Powerful Producer of Cold-Tolerant Chitinolytic Enzymes. Molecules 2016, 21, 447. [Google Scholar] [CrossRef] [Green Version]
- Yusof, N.A.; Hashim, N.H.F.; Bharudin, I. Cold Adaptation Strategies and the Potential of Psychrophilic Enzymes from the Antarctic Yeast, Glaciozyma antarctica PI12. J. Fungi 2021, 7, 528. [Google Scholar] [CrossRef]
- Min, W.; Guangfeng, K.; Cuijuan, S.; Qiuju, X.; Yingying, H.; Zhenhuan, L. In Heavy metal tolerance of an antarctic bacterial Strain O5 and its antioxidant enzyme activity changes induced by Cu2+. In Proceedings of the 2011 IEEE International Conference on Systems Biology (ISB), Zhuhai, China, 2–4 September 2011; pp. 303–306. [Google Scholar]
- Rajpert, L.; Sklodowska, A.; Matlakowska, R. Biotransformation of copper from Kupferschiefer black shale (Fore-Sudetic Monocline, Poland) by yeast Rhodotorula mucilaginosa LM9. Chemosphere 2013, 91, 1257–1265. [Google Scholar] [CrossRef]
- Ghaffari, S.; Sarp, A.S.K.; Lange, D.; Gulsoy, M. Potassium iodide potentiated photodynamic inactivation of Enterococcus faecalis using Toluidine Blue: Comparative analysis and post-treatment biofilm formation study. Photodiagn. Photodyn. Ther. 2018, 24, 245–249. [Google Scholar] [CrossRef]
- Zhang, H.B.; Wang, L.; Sha, T. Absorption of heavy metals by microorganisms isolated from lead-zinc tailing. J. Microbiol. 2004, 24, 34–37. [Google Scholar]
- Pinto, M.C.; Mata, A.M.; Lopez-Barea, J. Reversible inactivation of Saccharomyces cerevisiae glutathione reductase under reducing conditions. Arch. Biochem. Biophys. 1984, 228, 1–12. [Google Scholar] [CrossRef]
- Nazar, R.; Umar, S.; Khan, N.A. Exogenous salicylic acid improves photosynthesis and growth through an increase in ascorbate-glutathione metabolism and S assimilation in mustard under salt stress. Plant Signal. Behav. 2015, 10, e1003751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, Y.H.; Byun, J.R. Occurrence of glutathione sulphydryl (GSH) and antioxidant activities in probiotic Lactobacillus spp. Asian-Australas. J. Anim. Sci. 2004, 17, 1582–1585. [Google Scholar] [CrossRef]
- Freitas, C.; Nobre, B.; Gouveia, L.; Roseiro, J.; Reis, A.; da Silva, T.L. New at-line flow cytometric protocols for determining carotenoid content and cell viability during Rhodosporidium toruloides NCYC 921 batch growth. Process Biochem. 2014, 49, 554–562. [Google Scholar] [CrossRef] [Green Version]
- Ojuederie, O.B.; Babalola, O.O. Microbial and Plant-Assisted Bioremediation of Heavy Metal Polluted Environments: A Review. Int. J. Environ. Res. Public Health 2017, 14, 1504. [Google Scholar] [CrossRef] [Green Version]
- Tashirev, A.B.; Rokitko, P.V.; Levishko, A.S.; Romanovskaia, V.A.; Tashireva, A.A. Resistance of chemoorganotrophic bacteria isolated from Antarctic cliffs to toxic metals. Mikrobiol. Z. 2012, 74, 3–7. [Google Scholar]
- Kan, G.; Wang, X.; Jiang, J.; Zhang, C.; Chi, M.; Ju, Y.; Shi, C. Copper stress response in yeast Rhodotorula mucilaginosa AN5 isolated from sea ice, Antarctic. Microbiologyopen 2019, 8, e00657. [Google Scholar] [CrossRef]
- Escamilla-Rodríguez, A.; Carlos-Hernández, S.; Díaz-Jiménez, L. Evidence of Resistance of Heavy Metals from Bacteria Isolated from Natural Waters of a Mining Area in Mexico. Water 2021, 13, 2766. [Google Scholar] [CrossRef]
- Dias, M.; Gomes de Lacerda, J.T.J.; Perdigao Cota de Almeida, S.; de Andrade, L.M.; Oller do Nascimento, C.A.; Rozas, E.E.; Mendes, M.A. Response mechanism of mine-isolated fungus Aspergillus niger IOC 4687 to copper stress determined by proteomics. Metallomics 2019, 11, 1558–1566. [Google Scholar] [CrossRef]
- Elahi, A.; Rehman, A.; Zajif Hussain, S.; Zulfiqar, S.; Shakoori, A.R. Isolation and characterization of a highly effective bacterium Bacillus cereus b-525k for hexavalent chromium detoxification. Saudi J. Biol. Sci. 2022, 29, 2878–2885. [Google Scholar] [CrossRef] [PubMed]
- Jomova, K.; Valko, M. Advances in metal-induced oxidative stress and human disease. Toxicology 2011, 283, 65–87. [Google Scholar] [CrossRef] [PubMed]
- Banthia, S.; Hazra, C.; Sen, R.; Das, S.; Das, K. Electrodeposited functionally graded coating inhibits Gram-positive and Gram-negative bacteria by a lipid peroxidation mediated membrane damage mechanism. Mater. Sci. Eng. C Mater. Biol. Appl. 2019, 102, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Bruno-Barcena, J.M.; Azcarate-Peril, M.A.; Hassan, H.M. Role of antioxidant enzymes in bacterial resistance to organic acids. Appl. Environ. Microbiol. 2010, 76, 2747–2753. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Zhou, X.; Zhang, J.; Wang, J.; Xie, H.; Wu, Z. alpha-Lipoic acid protects against the cytotoxicity and oxidative stress induced by cadmium in HepG2 cells through regeneration of glutathione by glutathione reductase via Nrf2/ARE signaling pathway. Environ. Toxicol. Pharmacol. 2016, 45, 274–281. [Google Scholar] [CrossRef]
- Maria, P.; Antonella, C.; Marianna, D.; Elisa, Z.; Mario, S.; Filomena, D.; Carmen, R.; Clara, U.; Emilio, D.; Marco, L.G.; et al. Heavy-metal resistant microorganisms in sediments from submarine canyons and the adjacent continental slope in the northeastern Ligurian margin (Western Mediterranean Sea). Prog. Oceanogr. 2018, 168, 155–168. [Google Scholar]
- Yan, X.; Zhao, X.-E.; Sun, J.; Zhu, S.; Lei, C.; Li, R.; Gong, P.; Ling, B.; Wang, R.; Wang, H. Probing glutathione reductase activity with graphene quantum dots and gold nanoparticles system. Sens. Actuators B Chem. 2018, 263, 27–35. [Google Scholar] [CrossRef]
- Gonzalez-Dominguez, A.; Visiedo, F.; Dominguez-Riscart, J.; Ruiz-Mateos, B.; Saez-Benito, A.; Lechuga-Sancho, A.M.; Mateos, R.M. Blunted Reducing Power Generation in Erythrocytes Contributes to Oxidative Stress in Prepubertal Obese Children with Insulin Resistance. Antioxidants 2021, 10, 244. [Google Scholar] [CrossRef]
- Souri, Z.; Karimi, N.; Ahmad, P. The effect of NADPH oxidase inhibitor diphenyleneiodonium (DPI) and glutathione (GSH) on Isatis cappadocica, under Arsenic (As) toxicity. Int. J. Phytoremediat. 2021, 23, 945–957. [Google Scholar] [CrossRef]
- Singhvi, M.; Zendo, T.; Sonomoto, K. Free lactic acid production under acidic conditions by lactic acid bacteria strains: Challenges and future prospects. Appl. Microbiol. Biotechnol. 2018, 102, 5911–5924. [Google Scholar] [CrossRef]
- Zhai, Q.; Xiao, Y.; Narbad, A.; Chen, W. Comparative metabolomic analysis reveals global cadmium stress response of Lactobacillus plantarum strains. Metallomics 2018, 10, 1065–1077. [Google Scholar] [CrossRef] [PubMed]
- Fotiadis, D.; Kanai, Y.; Palacin, M. The SLC3 and SLC7 families of amino acid transporters. Mol. Aspects Med. 2013, 34, 139–158. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, R.; Tao, H.; Purvis, J.E.; York, S.W.; Shanmugam, K.T.; Ingram, L.O. Gene array-based identification of changes that contribute to ethanol tolerance in ethanologenic Escherichia coli: Comparison of KO11 (parent) to LY01 (resistant mutant). Biotechnol. Prog. 2003, 19, 612–623. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, H.; Hu, T.; Li, H.; Jin, G.; Zhang, Y. Metabonomic profiling in study hepatoprotective effect of polysaccharides from Flammulina velutipes on carbon tetrachloride-induced acute liver injury rats using GC-MS. Int. J. Biol. Macromol. 2018, 110, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Olin-Sandoval, V.; Yu, J.S.L.; Miller-Fleming, L.; Alam, M.T.; Kamrad, S.; Correia-Melo, C.; Haas, R.; Segal, J.; Pena Navarro, D.A.; Herrera-Dominguez, L.; et al. Lysine harvesting is an antioxidant strategy and triggers underground polyamine metabolism. Nature 2019, 572, 249–253. [Google Scholar] [CrossRef]
- Tian, B.; Qiao, Z.; Zhang, L.; Li, H.; Pei, Y. Hydrogen sulfide and proline cooperate to alleviate cadmium stress in foxtail millet seedlings. Plant Physiol. Biochem. 2016, 109, 293–299. [Google Scholar] [CrossRef]
- Srivastava, S.; Srivastava, S. Prescience of endogenous regulation in Arabidopsis thaliana by Pseudomonas putida MTCC 5279 under phosphate starved salinity stress condition. Sci. Rep. 2020, 10, 5855. [Google Scholar] [CrossRef] [Green Version]
- Raza, M.F.; Wang, Y.; Cai, Z.; Bai, S.; Yao, Z.; Awan, U.A.; Zhang, Z.; Zheng, W.; Zhang, H. Gut microbiota promotes host resistance to low-temperature stress by stimulating its arginine and proline metabolism pathway in adult Bactrocera dorsalis. PLoS Pathog. 2020, 16, e1008441. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Ni, S.; Cai, S.; Zhang, G. Comprehensive dissection of primary metabolites in response to diverse abiotic stress in barley at the seedling stage. Plant Physiol. Biochem. 2021, 161, 54–64. [Google Scholar] [CrossRef]
- Geng, P.; Qin, W.; Xu, G. Proline metabolism in cancer. Amino Acids 2021, 53, 1769–1777. [Google Scholar] [CrossRef]
- Shavalikohshori, O.; Zalaghi, R.; Sorkheh, K.; Enaytizamir, N. The expression of proline production/degradation genes under salinity and cadmium stresses in Triticum aestivum inoculated with Pseudomonas sp. Int. J. Environ. Sci. Technol. 2019, 17, 2233–2242. [Google Scholar] [CrossRef]
- Tyminski, M.; Ciacka, K.; Staszek, P.; Gniazdowska, A.; Krasuska, U. Toxicity of meta-Tyrosine. Plants 2021, 10, 2800. [Google Scholar] [CrossRef]
- Ipson, B.R.; Fisher, A.L. Roles of the tyrosine isomers meta-tyrosine and ortho-tyrosine in oxidative stress. Ageing Res. Rev. 2016, 27, 93–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lokvam, J.; Brenes-Arguedas, T.; Lee, J.S.; Coley, P.D.; Kursar, T.A. Allelochemic function for a primary metabolite: The case of l-tyrosine hyper-production in Inga umbellifera (Fabaceae). Am. J. Bot. 2006, 93, 1109–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacevic, V.; Simpson, A.J.; Simpson, M.J. Evaluation of Daphnia magna metabolic responses to organic contaminant exposure with and without dissolved organic matter using 1H nuclear magnetic resonance (NMR)-based metabolomics. Ecotoxicol. Environ. Saf. 2018, 164, 189–200. [Google Scholar] [CrossRef]
- Li, W.H.; Shi, Y.C.; Chang, C.H.; Huang, C.W.; Hsiu-Chuan Liao, V. Selenite protects Caenorhabditis elegans from oxidative stress via DAF-16 and TRXR-1. Mol. Nutr. Food Res. 2014, 58, 863–874. [Google Scholar] [CrossRef] [PubMed]
- Shi, K.; Gao, Z.; Shi, T.Q.; Song, P.; Ren, L.J.; Huang, H.; Ji, X.J. Reactive Oxygen Species-Mediated Cellular Stress Response and Lipid Accumulation in Oleaginous Microorganisms: The State of the Art and Future Perspectives. Front. Microbiol. 2017, 8, 793. [Google Scholar] [CrossRef]
- Mirosława, S.; Przemysław, B.; Sylwia, R.; Justyna, N.; Jerzy, D. Comparative study of metal induced phospholipid modifications in the heavy metal tolerant filamentous fungus Paecilomyces marquandii and implications for the fungal membrane integrity. Acta Biochim. Pol. 2013, 60, 695–700. [Google Scholar]
- Baoukina, S.; Rozmanov, D.; Tieleman, D.P. Composition Fluctuations in Lipid Bilayers. Biophys. J. 2017, 113, 2750–2761. [Google Scholar] [CrossRef] [Green Version]
- Mikami, K.; Murata, N. Membrane fluidity and the perception of environmental signals in cyanobacteria and plants. Prog. Lipid Res. 2003, 42, 527–543. [Google Scholar] [CrossRef]
- Ramos, J.L.; Duque, E.; Gallegos, M.T.; Godoy, P.; Ramos-Gonzalez, M.I.; Rojas, A.; Teran, W.; Segura, A. Mechanisms of solvent tolerance in gram-negative bacteria. Annu. Rev. Microbiol. 2002, 56, 743–768. [Google Scholar] [CrossRef]
- Antsotegi-Uskola, M.; Markina-Inarrairaegui, A.; Ugalde, U. New insights into copper homeostasis in filamentous fungi. Int. Microbiol. 2020, 23, 65–73. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Fu, L.; Hu, Z.; Li, Y.; Zheng, X.; Zhang, Z.; Jiang, C.; Zeng, B. Modulation of Fatty Acid Composition of Aspergillus oryzae in Response to Ethanol Stress. Microorganisms 2019, 7, 158. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, Z.; Shi, C.; Gao, X.; Wang, X.; Kan, G. Biochemical and Metabolomic Responses of Antarctic Bacterium Planococcus sp. O5 Induced by Copper Ion. Toxics 2022, 10, 302. https://doi.org/10.3390/toxics10060302
Cheng Z, Shi C, Gao X, Wang X, Kan G. Biochemical and Metabolomic Responses of Antarctic Bacterium Planococcus sp. O5 Induced by Copper Ion. Toxics. 2022; 10(6):302. https://doi.org/10.3390/toxics10060302
Chicago/Turabian StyleCheng, Ziyi, Cuijuan Shi, Xiujun Gao, Xiaofei Wang, and Guangfeng Kan. 2022. "Biochemical and Metabolomic Responses of Antarctic Bacterium Planococcus sp. O5 Induced by Copper Ion" Toxics 10, no. 6: 302. https://doi.org/10.3390/toxics10060302
APA StyleCheng, Z., Shi, C., Gao, X., Wang, X., & Kan, G. (2022). Biochemical and Metabolomic Responses of Antarctic Bacterium Planococcus sp. O5 Induced by Copper Ion. Toxics, 10(6), 302. https://doi.org/10.3390/toxics10060302