
Lead and Other Trace Element Levels in Brains of Croatian Large Terrestrial Carnivores: Influence of Biological and Ecological Factors
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Sampling
2.2. Element Analyses
2.3. Statistical Procedure
3. Results
3.1. Between-Species Differences in Trace Elements
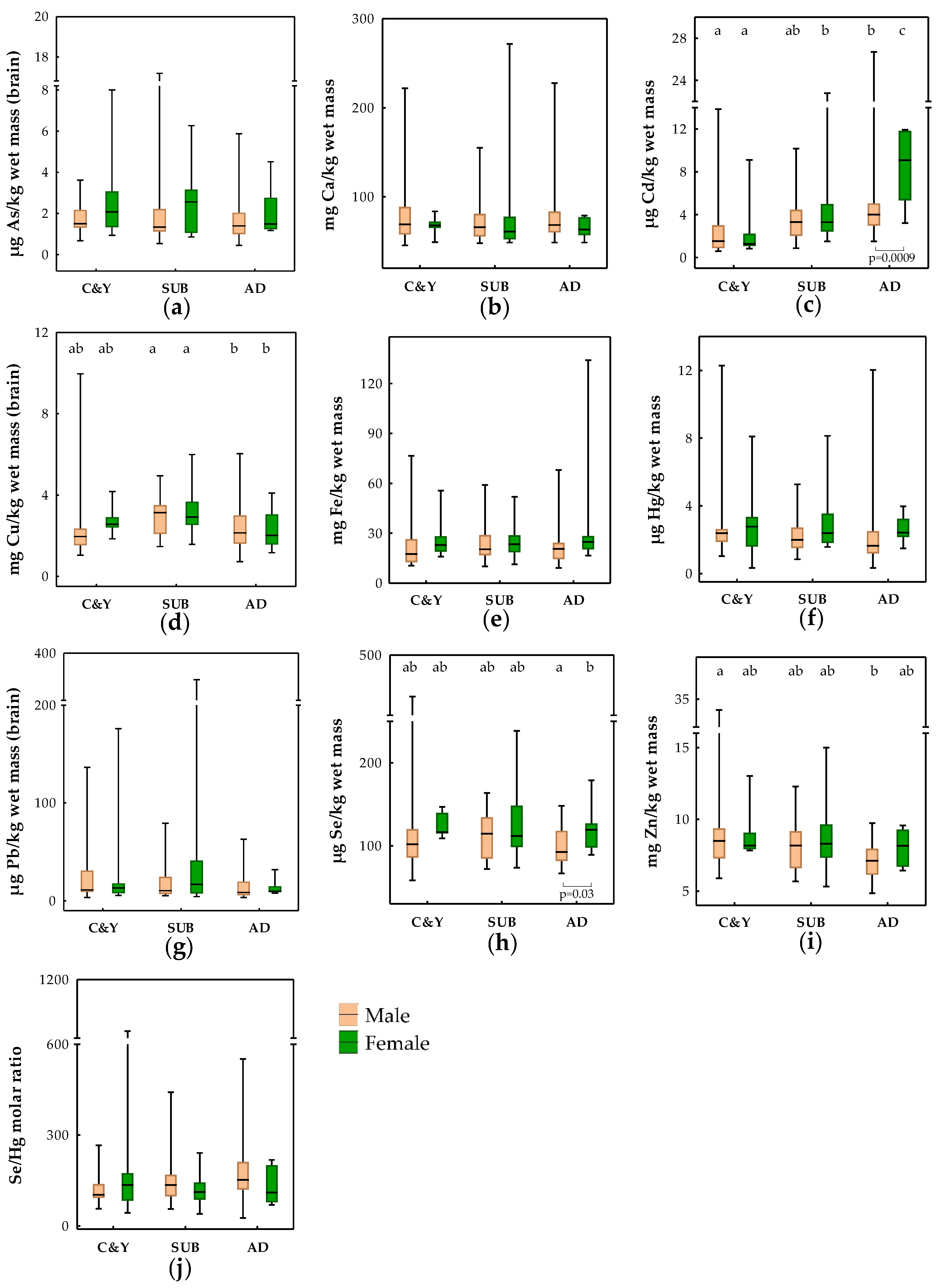
3.2. Influence of Age, Sex, BCI, and Season on Trace Elements in the Brains of Brown Bears
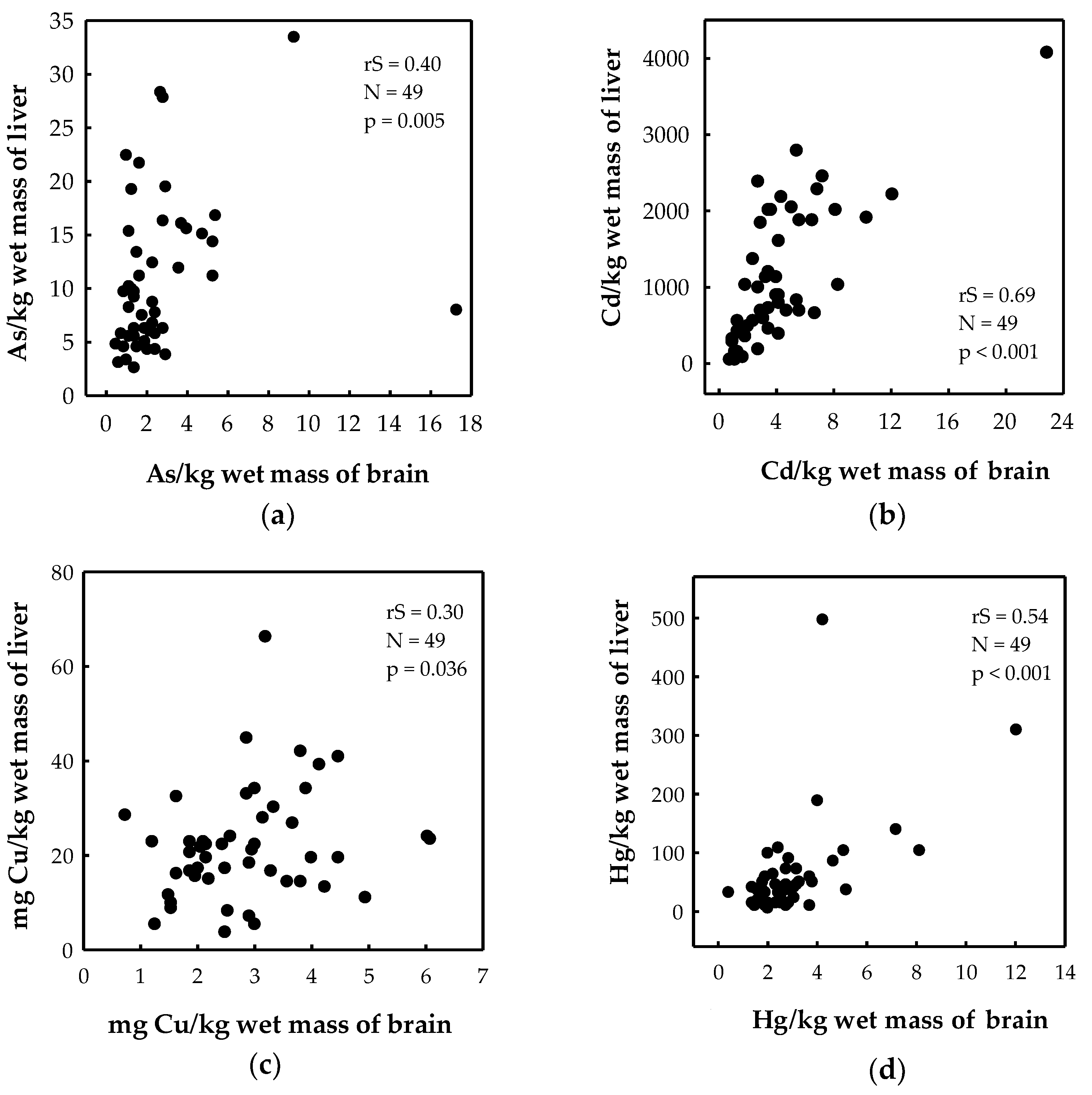
3.3. Association of Trace Elements between the Brain and Liver
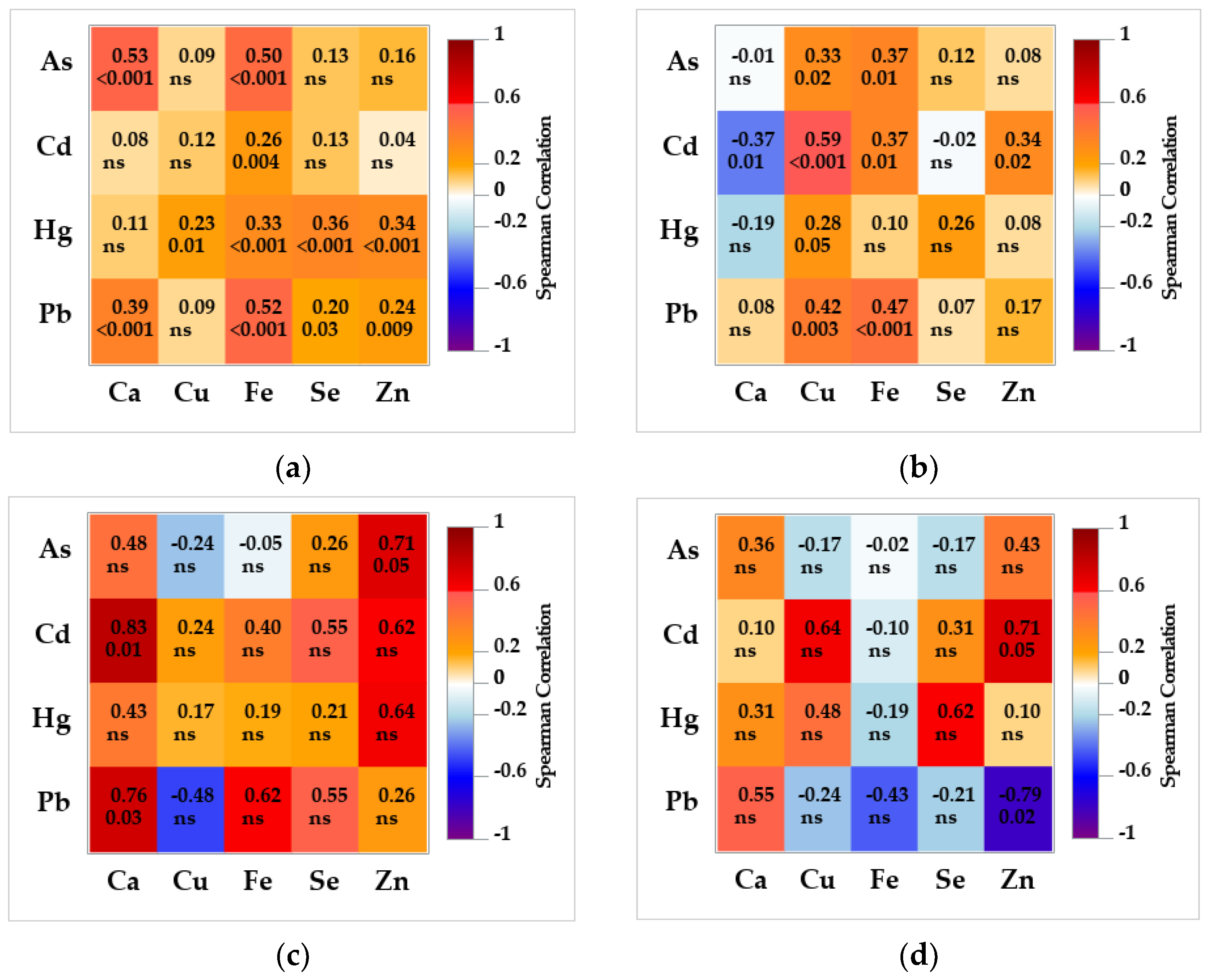
3.4. Association between Non-Essential and Essential Trace Elements in the Brain and Liver
4. Discussion
4.1. Between-Species Differences in Trace Elements
4.2. Influence of Age, Sex, BCI, and Season on Trace Elements in Brains of Brown Bears
4.3. Association of Trace Elements between the Brain and Liver
4.4. Associations between Non-Essential and Essential Trace Elements in the Brain and Liver
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- López-Berenguer, G.; Peñalver, J.; Martínez-López, E. A critical review about neurotoxic effects in marine mammals of mercury and other trace elements. Chemosphere 2020, 246, 125688. [Google Scholar] [CrossRef] [PubMed]
- Desforges, J.P.; Mikkelsen, B.; Dam, M.; Rigét, F.; Sveegaard, S.; Sonne, C.; Dietz, R.; Basu, N. Mercury and neurochemical biomarkers in multiple brain regions of five Arctic marine mammals. Neurotoxicology 2021, 84, 136–145. [Google Scholar] [CrossRef] [PubMed]
- Caito, S.; Aschner, M. Chapter Eleven—Neurotoxicity of metals. In Occupational Neurology; Lotti, M., Bleecker, M.L., Eds.; Handbook of Clinical Neurology; Elsevier: Amsterdam, The Netherlands, 2015; Volume 131, pp. 169–189. [Google Scholar]
- Wu, X.; Cobbina, S.J.; Mao, G.; Xu, H.; Zhang, Z.; Yang, L. A review of toxicity and mechanisms of individual and mixtures of heavy metals in the environment. Environ. Sci. Pollut. Res. 2016, 23, 8244–8259. [Google Scholar] [CrossRef] [PubMed]
- Ruczaj, A.; Brzóska, M.M. Environmental exposure of the general population to cadmium as a risk factor of the damage to the nervous system: A critical review of current data. J. Appl. Toxicol. 2022, 43, 66–88. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.S.; Nogara, P.A.; Ardisson-Araújo, D.M.P.; Aschner, M.; Rocha, J.B.T.; Dórea, J.G. Chapter Two—Neurodevelopmental effects of mercury. In Advances in Neurotoxicology; Aschner, M., Costa, L.G., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2018; Volume 2, pp. 27–86. [Google Scholar]
- Andrade, V.M.; Aschner, M.; Marreilha dos Santos, A.P. Neurotoxicity of metal mixtures. In Advances in Neurobiology; Aschner, M., Costa, L., Eds.; Springer: Cham, Switzerland, 2017; Volume 18, pp. 227–265. [Google Scholar]
- Masjosthusmann, S.; Tigges, J.; Fritsche, E.; Koch, K. Chapter Two—Arsenic-mediated developmental neurotoxicity: Recent advances in understanding the adverse outcomes and underlying mechanisms. In Neurotoxicity of Metals: Old Issues and New Developments. Advances in Neurotoxicology; Aschner, M., Costa, L.G., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2021; Volume 5, pp. 35–80. [Google Scholar]
- Thakur, M.; Rachamalla, M.; Niyogi, S.; Datusalia, A.K.; Flora, S.J.S. Molecular mechanism of arsenic-induced neurotoxicity including neuronal dysfunctions. Int. J. Mol. Sci. 2021, 22, 10077. [Google Scholar] [CrossRef]
- Gonçalves, J.F.; Dressler, V.L.; Assmann, C.E.; Morsch, V.M.M.; Schetinger, M.R.C. Chapter Three—Cadmium neurotoxicity: From its analytical aspects to neuronal impairment. In Neurotoxicity of Metals: Old Issues and New Developments; Aschner, M., Costa, L.G., Eds.; Advances in Neurotoxicology; Elsevier Inc.: Amsterdam, The Netherlands, 2021; Volume 5, pp. 81–113. [Google Scholar]
- Bjørklund, G.; Dadar, M.; Mutter, J.; Aaseth, J. The toxicology of mercury: Current research and emerging trends. Environ. Res. 2017, 159, 545–554. [Google Scholar] [CrossRef]
- Branco, V.; Aschner, M.; Carvalho, C. Chapter Seven—Neurotoxicity of mercury: An old issue with contemporary significance. In Neurotoxicity of Metals: Old Issues and New Developments. Advances in Neurotoxicology; Aschner, M., Costa, L.G., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2021; Volume 5, pp. 239–262. [Google Scholar]
- Cory-Slechta, D.A. Relationships between lead-induced learning impairments and changes in dopaminergic, cholinergic, and glutamatergic neurotransmitter system functions. Annu. Rev. Pharmacol. Toxicol. 1995, 35, 391–415. [Google Scholar] [CrossRef]
- Agency for Toxic Substances and Disease Registry (ATSDR). Toxicological Profile for Lead. Department of Health and Human Services, Public Health Service, Centers for Disease Control Atlanta, ATSDR, Atlanta, GA, USA, 2020. Available online: https://www.atsdr.cdc.gov/toxprofiles/tp13.pdf (accessed on 25 May 2022).
- Virgolini, M.B.; Aschner, M. Molecular mechanisms of lead neurotoxicity. In Neurotoxicity of Metals: Old Issues and New Developments. Advances in Neurotoxicology; Aschner, M., Costa, L.G., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2021; Volume 5, pp. 159–213. [Google Scholar]
- European Food Safety Authority (EFSA). Scientific opinion on lead in food. EFSA J. 2010, 8, 1570. [Google Scholar]
- Grandjean, P.; Herz, K.T. Trace elements as paradigms of developmental neurotoxicants: Lead, methylmercury and arsenic. J. Trace Elem. Med. Biol. 2015, 31, 130–134. [Google Scholar] [CrossRef] [Green Version]
- Bertram, M.G.; Martin, J.M.; McCallum, E.S.; Alton, L.A.; Brand, J.A.; Brooks, B.W.; Cerveny, D.; Fick, J.; Ford, A.T.; Hellström, G.; et al. Frontiers in quantifying wildlife behavioural responses to chemical pollution. Biol. Rev. 2022, 97, 1346–1364. [Google Scholar] [CrossRef]
- National Research Council (U.S.). Pesticides in the Diets of Infants and Children. Committee on Pesticides in the Diets of Infants and Children; National Academy Press: Washington, DC, USA, 1993; Available online: https://nap.nationalacademies.org/catalog/2126/pesticides-in-the-diets-of-infants-and-children (accessed on 25 May 2022).
- Landrigan, P.J. Children as a vulnerable population. Int. J. Occup. Med. Environ. Health 2005, 17, 175–177. [Google Scholar] [CrossRef]
- Dietz, R.; Fort, J.; Sonne, C.; Albert, C.; Bustnes, J.O.; Christensen, T.K.; Ciesielski, T.M.; Danielsen, J.; Dastnai, S.; Eens, M.; et al. A risk assessment of the effects of mercury on Baltic Sea, Greater North Sea and North Atlantic wildlife, fish and bivalves. Environ. Int. 2021, 146, 106178. [Google Scholar] [CrossRef] [PubMed]
- Chiverton, L.; Cromie, R.; Kock, R. European mammal exposure to lead from ammunition and fishing weight sources. Heliyon 2022, 8, e10014. [Google Scholar] [CrossRef] [PubMed]
- Nam, D.H.; Rutkiewicz, J.; Basu, N. Multiple metals exposure and neurotoxic risk in bald eagles (Haliaeetus leucocephalus) from two Great Lakes states. Environ. Toxicol. Chem. 2012, 31, 623–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rutkiewicz, J.; Nam, D.H.; Cooley, T.; Neumann, K.; Padilla, I.B.; Route, W.; Strom, S.; Basu, N. Mercury exposure and neurochemical impacts in bald eagles across several Great Lakes states. Ecotoxicology 2011, 20, 1669–1676. [Google Scholar] [CrossRef] [PubMed]
- Grunst, A.S.; Grunst, M.L.; Daem, N.; Pinxten, R.; Bervoets, L.; Eens, M. An important personality trait varies with blood and plumage metal concentrations in a free-living songbird. Environ. Sci. Technol. 2019, 53, 10487–10496. [Google Scholar] [CrossRef]
- Franson, J.C.; Pain, D.J. Lead in birds. In Environmental Contaminants in Biota: Interpreting Tissue Concentrations, 2nd ed.; Beyer, W.N., Meador, J.P., Eds.; USGS Staff—Published Research; CRC Press: Boca Raton, FL, USA, 2011; pp. 563–594. [Google Scholar]
- Ecke, F.; Singh, N.J.; Arnemo, J.M.; Bignert, A.; Helander, B.; Berglund, Å.M.M.; Borg, H.; Bröjer, C.; Holm, K.; Lanzone, M.; et al. Sublethal lead exposure alters movement behavior in free-ranging golden eagles. Environ. Sci. Technol. 2017, 51, 5729–5736. [Google Scholar] [CrossRef]
- McClelland, S.C.; Durães Ribeiro, R.; Mielke, H.W.; Finkelstein, M.E.; Gonzales, C.R.; Jones, J.A.; Komdeur, J.; Derryberry, E.; Saltzberg, E.B.; Karubian, J. Sub-lethal exposure to lead is associated with heightened aggression in an urban songbird. Sci. Total Environ. 2019, 654, 593–603. [Google Scholar] [CrossRef]
- Basu, N.; Scheuhammer, A.M.; Sonne, C.; Letcher, R.J.; Born, E.W.; Dietz, R. Is dietary mercury of neurotoxicological concern to wild polar bears (Ursus maritimus)? Environ. Toxicol. Chem. 2009, 28, 133–140. [Google Scholar] [CrossRef]
- Haines, K.J.R.; Evans, R.D.; O’Brien, M.; Evans, H.E. Accumulation of mercury and selenium in the brain of river otters (Lontra canadensis) and wild mink (Mustela vison) from Nova Scotia, Canada. Sci. Total Environ. 2010, 408, 537–542. [Google Scholar] [CrossRef]
- Krey, A.; Kwan, M.; Chan, H.M. Mercury speciation in brain tissue of polar bears (Ursus maritimus) from the Canadian Arctic. Environ. Res. 2012, 114, 24–30. [Google Scholar] [CrossRef] [PubMed]
- Krey, A.; Kwan, M.; Chan, H.M. In vivo and in vitro changes in neurochemical parameters related to mercury concentrations from specific brain regions of polar bears (Ursus maritimus). Environ. Toxicol. Chem. 2014, 33, 2463–2471. [Google Scholar] [CrossRef] [PubMed]
- Krey, A.; Ostertag, S.K.; Chan, H.M. Assessment of neurotoxic effects of mercury in beluga whales (Delphinapterus leucas), ringed seals (Pusa hispida), and polar bears (Ursus maritimus) from the Canadian Arctic. Sci. Total Environ. 2015, 509–510, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Scheuhammer, A.; Braune, B.; Chan, H.M.; Frouin, H.; Krey, A.; Letcher, R.; Loseto, L.; Noël, M.; Ostertag, S.; Ross, P.; et al. Recent progress on our understanding of the biological effects of mercury in fish and wildlife in the Canadian Arctic. Sci. Total Environ. 2015, 509–510, 91–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, S.; Coelho, J.P.; Bandeira, V.; Barros, T.; Duarte, A.C.; Fonseca, C.; Pereira, M.E. Mercury bioaccumulation in the egyptian mongoose (Herpestes ichneumon): Geographical, tissue, gender and age differences. Water Air Soil Pollut. 2014, 225, 2005. [Google Scholar] [CrossRef]
- Komov, V.T.; Ivanova, E.S.; Gremyachikh, V.A.; Poddubnaya, N.Y. Mercury content in organs and tissues of indigenous (Vulpes vulpes L.) and invasive (Nyctereutes procyonoides Gray.) species of canids from areas near Cherepovets (North-Western Industrial Region, Russia). Bull. Environ. Contam. Toxicol. 2016, 97, 480–485. [Google Scholar] [CrossRef]
- Lanocha, N.; Kalisinska, E.; Kosik-Bogacka, D.I.; Budis, H.; Podlasinska, J.; Jedrzejewska, E. Mercury levels in raccoons (Procyon lotor) from the Warta Mouth National Park, Northwestern Poland. Biol. Trace Elem. Res. 2014, 159, 152–160. [Google Scholar] [CrossRef] [Green Version]
- Kalisinska, E.; Lanocha-Arendarczyk, N.; Kosik-Bogacka, D.; Budis, H.; Podlasinska, J.; Popiolek, M.; Pirog, A.; Jedrzejewska, E. Brains of native and alien mesocarnivores in biomonitoring of toxic metals in Europe. PLoS ONE 2016, 11, e0159935. [Google Scholar] [CrossRef]
- Rodríguez-Jorquera, I.A.; Vitale, N.; Garner, L.; Perez-Venegas, D.J.; Galbán-Malagón, C.J.; Duque-Wilckens, N.; Toor, G.S. Contamination of the upper class: Occurrence and effects of chemical pollutants in terrestrial top predators. Curr. Pollut. Rep. 2017, 3, 206–219. [Google Scholar] [CrossRef]
- Lazarus, M.; Sekovanić, A.; Orct, T.; Reljić, S.; Kusak, J.; Jurasović, J.; Huber, Đ. Apex predatory mammals as bioindicator species in environmental monitoring of elements in Dinaric Alps (Croatia). Environ. Sci. Pollut. Res. 2017, 24, 23977–23991. [Google Scholar] [CrossRef]
- Lazarus, M.; Orct, T.; Sergiel, A.; Vranković, L.; Marijić, V.F.; Rašić, D.; Reljić, S.; Aladrović, J.; Zwijacz-Kozica, T.; Zięba, F.; et al. Metal(loid) exposure assessment and biomarker responses in captive and free-ranging European brown bear (Ursus arctos). Environ. Res. 2020, 183, 109166. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, H.; Taki, Y.; Nouchi, R.; Yokoyama, R.; Kotozaki, Y.; Nakagawa, S.; Sekiguchi, A.; Iizuka, K.; Hanawa, S.; Araki, T.; et al. Lead exposure is associated with functional and microstructural changes in the healthy human brain. Commun. Biol. 2021, 4, 912. [Google Scholar] [CrossRef] [PubMed]
- Lazarus, M.; Sekovanić, A.; Orct, T.; Reljić, S.; Jurasović, J.; Huber, Đ. Sexual maturity and life stage influences toxic metal accumulation in Croatian brown bears. Arch. Environ. Contam. Toxicol. 2018, 74, 339–348. [Google Scholar] [CrossRef]
- Bechshoft, T.; Derocher, A.E.; Viengkone, M.; Routti, H.; Aars, J.; Letcher, R.J.; Dietz, R.; Sonne, C.; Jenssen, B.M.; Richardson, E.; et al. On the integration of ecological and physiological variables in polar bear toxicology research: A systematic review. Environ. Rev. 2018, 26, 1–12. [Google Scholar] [CrossRef]
- Malvandi, H.; Ghasempouri, S.M.; Esmaili-Sari, A.; Bahramifar, N. Evaluation of the suitability of application of golden jackal (Canis aureus) hair as a noninvasive technique for determination of body burden mercury. Ecotoxicology 2010, 19, 997–1002. [Google Scholar] [CrossRef] [PubMed]
- Cattet, M.R.L.; Caulkett, N.A.; Obbard, M.E.; Stenhouse, G.B. A body-condition index for Ursids. Can. J. Zool. 2002, 80, 1156–1161. [Google Scholar] [CrossRef] [Green Version]
- Lazarus, M.; Sekovanić, A.; Reljić, S.; Kusak, J.; Kovačić, J.; Orct, T.; Jurasović, J.; Huber, D. Selenium in brown bears (Ursus arctos) from Croatia: Relation to cadmium and mercury. J. Environ. Sci. Health Part A Toxic/Hazardous Subst. Environ. Eng. 2014, 49, 1392–1401. [Google Scholar] [CrossRef] [PubMed]
- Romanić, S.H.; Klinčić, D.; Kljakovic-Gašpić, Z.; Kusak, J.; Reljić, S.; Huber, D. Organochlorine pesticides and polychlorinated biphenyl congeners in wild terrestrial mammals from Croatia: Interspecies comparison of residue levels and compositions. Chemosphere 2015, 137, 52–58. [Google Scholar] [CrossRef]
- Rodríguez-Estival, J.; Mateo, R. Exposure to anthropogenic chemicals in wild carnivores: A silent conservation threat demanding long-term surveillance. Curr. Opin. Environ. Sci. Health 2019, 11, 21–25. [Google Scholar] [CrossRef]
- Chapron, G.; Kaczensky, P.; Linnell, J.D.C.; von Arx, M.; Huber, D.; Andrén, H.; López-Bao, J.V.; Adamec, M.; Álvares, F.; Anders, O.; et al. Recovery of large carnivores in Europe’s modern human-dominated landscapes. Science 2014, 346, 1517–1519. [Google Scholar] [CrossRef] [Green Version]
- Trouwborst, A.; Krofel, M.; Linnell, J.D.C. Legal implications of range expansions in a terrestrial carnivore: The case of the golden jackal (Canis aureus) in Europe. Biodivers. Conserv. 2015, 24, 2593–2610. [Google Scholar] [CrossRef]
- Sienkiewicz, T.; Sergiel, A.; Huber, D.; Maślak, R.; Wrzosek, M.; Podgórski, P.; Reljić, S.; Paśko, Ł. The brain anatomy of the brown bear (Carnivora, Ursus arctos L., 1758) Compared to that of other carnivorans: A cross-sectional study using MRI. Front. Neuroanat. 2019, 13, 79. [Google Scholar] [CrossRef] [Green Version]
- Huber, D.; Kusak, J.; Majić-Skrbinšek, A.; Majnarić, D.; Sindičić, M. A multidimensional approach to managing the European brown bear in Croatia. Ursus 2008, 19, 22–32. [Google Scholar] [CrossRef]
- Stoneberg, R.P.; Jonkel, C.J. Age determination of black bears by cementum layers. J. Wildl. Manag. 1966, 30, 411–414. [Google Scholar] [CrossRef]
- Gipson, P.S.; Ballard, W.B.; Nowak, R.M.; Mech, L.D. Accuracy and precision of estimating age of gray wolves by tooth wear. J. Wildl. Manag. 2000, 64, 752. [Google Scholar] [CrossRef]
- Marti, I.; Ryser-Degiorgis, M.P. A tooth wear scoring scheme for age estimation of the Eurasian lynx (Lynx lynx) under field conditions. Eur. J. Wildl. Res. 2018, 64, 37. [Google Scholar] [CrossRef] [Green Version]
- Knott, E.J.; Bunnefeld, N.; Huber, D.; Reljić, S.; Kereži, V.; Milner-Gulland, E.J. The potential impacts of changes in bear hunting policy for hunting organisations in Croatia. Eur. J. Wildl. Res. 2014, 60, 85–97. [Google Scholar] [CrossRef] [Green Version]
- Wikenros, C.; Gicquel, M.; Zimmermann, B.; Flagstad, Ø.; Åkesson, M. Age at first reproduction in wolves: Different patterns of density dependence for females and males. Proc. R. Soc. B Biol. Sci. 2021, 288, 20210207. [Google Scholar] [CrossRef]
- Mech, L. The Wolf: The Ecology and Behavior of an Endangered Species; The Natural History Press: Garden City, New York, NY, USA, 1970. [Google Scholar]
- Lazarus, M.V.; Sekovanić, A.; Kljaković-Gašpic, Z.; Orct, T.; Jurasović, J.; Kusak, J.; Reljić, S.; Huber, D. Cadmium and lead in grey wolf liver samples: Optimisation of a microwaveassisted digestion method. Arh. Hig. Rada Toksikol. 2013, 64, 395–403. [Google Scholar] [CrossRef] [Green Version]
- Krebs, N.; Langkammer, C.; Goessler, W.; Ropele, S.; Fazekas, F.; Yen, K.; Scheurer, E. Assessment of trace elements in human brain using inductively coupled plasma mass spectrometry. J. Trace Elem. Med. Biol. 2014, 28, 1–7. [Google Scholar] [CrossRef]
- Huber, Đ.; Manen, F.T. Bear Morphology. In Encyclopedia of Animal Cognition and Behavior; Vonk, J., Shackelford, T., Eds.; Springer: Berlin/Heidelberg, Germany, 2019. [Google Scholar]
- Puls, R. Mineral Levels in Animal Health. Diagnostic Data; Sherpa International: Clearbrook, BC, Canada, 1994. [Google Scholar]
- Monclús, L.; Shore, R.F.; Krone, O. Lead contamination in raptors in Europe: A systematic review and meta-analysis. Sci. Total Environ. 2020, 748, 141437. [Google Scholar] [CrossRef] [PubMed]
- Ma, W. Lead in mammals. In Environmental Contaminants in Biota: Interpreting Tissue Concentrations; Beyer, W.N., Meador, J.P., Eds.; CRC Press Inc: Boca Raton, FL, USA, 2011; pp. 595–608. [Google Scholar]
- Souza, M.J.; Ramsay, E.C.; Donnell, R.L. Metal accumulation and health effects in raccoons (Procyon lotor) associated with coal fly ash exposure. Arch. Environ. Contam. Toxicol. 2013, 64, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Beyer, W.N.; Connor, E.E.; Gerould, S. Estimates of soil ingestion by wildlife. J. Wildl. Manag. 1994, 58, 375. [Google Scholar] [CrossRef]
- García, M.H.D.M.; Hernández Moreno, D.; Soler Rodríguez, F.; Beceiro, A.L.; Álvarez, L.E.F.; López, M.P. Sex- and age-dependent accumulation of heavy metals (Cd, Pb and Zn) in liver, kidney and muscle of roe deer (Capreolus capreolus) from NW Spain. J. Environ. Sci. Health Part A Toxic/Hazardous Subst. Environ. Eng. 2011, 46, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Nordberg, M.; Nordberg, G.F. Metallothionein and cadmium toxicology—Historical review and commentary. Biomolecules 2022, 12, 360. [Google Scholar] [CrossRef]
- Nordberg, G.; Nogawa, K.; Nordberg, M. Cadmium. In Handbook on the Toxicology of Metals, 4th ed.; Academic Press: Amsterdam, The Netherlands, 2015; Volume 2, pp. 667–716. [Google Scholar]
- Wang, B.; Du, Y. Cadmium and its neurotoxic effects. Oxid. Med. Cell. Longev. 2013, 2013, 898034. [Google Scholar] [CrossRef] [Green Version]
- Malavolta, M.; Cipriano, C.; Costarelli, L.; Giacconi, R.; Tesei, S.; Muti, E.; Piacenza, F.; Pierpaoli, S.; Larbi, A.; Pawelec, G.; et al. Metallothionein downregulation in very old age: A phenomenon associated with cellular senescence? Rejuvenation Res. 2008, 11, 455–459. [Google Scholar] [CrossRef]
- Bridges, C.C.; Zalups, R.K. Molecular and ionic mimicry and the transport of toxic metals. Toxicol. Appl. Pharmacol. 2005, 204, 274–308. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.-T.; Zhen, J.; Leng, J.Y.; Cai, L.; Ji, H.L.; Keller, B.B. Zinc as a countermeasure for cadmium toxicity. Acta Pharmacol. Sin. 2021, 42, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Huber, D.; Kulier, I.; Poljak, A.; Devčić-Kuhar, B. Food intake and mass gain of hand-reared brown bear cubs. Zoo Biol. 1993, 12, 525–533. [Google Scholar] [CrossRef]
- Hissa, R. Physiology of the European brown bear (Ursus Arctos Arctos). Ann. Zool. Fennici 1997, 34, 267–287. [Google Scholar]
- Vahter, M.; Åkesson, A.; Lidén, C.; Ceccatelli, S.; Berglund, M. Gender differences in the disposition and toxicity of metals. Environ. Res. 2007, 104, 85–95. [Google Scholar] [CrossRef]
- Llop, S.; Lopez-Espinosa, M.J.; Rebagliato, M.; Ballester, F. Gender differences in the neurotoxicity of metals in children. Toxicology 2013, 311, 3–12. [Google Scholar] [CrossRef]
- Zaidi, Z.F. Gender differences in human brain: A review. Open Anat. J. 2010, 2, 37–55. [Google Scholar] [CrossRef] [Green Version]
- Tyler, C.R.; Allan, A.M. The effects of arsenic exposure on neurological and cognitive dysfunction in human and rodent studies: A review. Curr. Environ. Health Rep. 2014, 1, 132–147. [Google Scholar] [CrossRef] [Green Version]
- Reglero, M.M.; Taggart, M.A.; Monsalve-González, L.; Mateo, R. Heavy metal exposure in large game from a lead mining area: Effects on oxidative stress and fatty acid composition in liver. Environ. Pollut. 2009, 157, 1388–1395. [Google Scholar] [CrossRef]
- Berzas Nevado, J.J.; Rodríguez Martin-Doimeadios, R.C.; Mateo, R.; Rodríguez Fariñas, N.; Rodríguez-Estival, J.; Patiño Ropero, M.J. Mercury exposure and mechanism of response in large game using the almadén mercury mining area (Spain) as a case study. Environ. Res. 2012, 112, 58–66. [Google Scholar] [CrossRef] [Green Version]
- Vighi, M.; Borrell, A.; Aguilar, A. Bone as a surrogate tissue to monitor metals in Baleen whales. Chemosphere 2017, 171, 81–88. [Google Scholar] [CrossRef]
- Peraza, M.A.; Ayala-Fierro, F.; Barber, D.S.; Casarez, E.; Rael, L.T. Effects of micronutrients on metal toxicity. Environ. Health Perspect. 1998, 106, 203–216. [Google Scholar]
- Hudson, L.G.; Cooper, K.L.; Atlas, S.R.; King, B.S.; Jian, L.K. Arsenic interaction with zinc finger motifs. In Arsenic: Exposure Sources, Health Risks, and Mechanisms of Toxicity; States, C., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; pp. 291–314. [Google Scholar]
- Porcella, D.B.; Zillioux, E.J.; Grieb, T.M.; Newman, J.R.; West, G.B. Retrospective study of mercury in raccoons (Procyon lotor) in South Florida. Ecotoxicology 2004, 13, 207–221. [Google Scholar] [CrossRef]
- Woshner, V.M.; O’Hara, T.M.; Bratton, G.R.; Suydam, R.S.; Beasley, V.R. Concentrations and interactions of selected essential and non-essential elements in bowhead and Beluga whales of Arctic Alaska. J. Wildl. Dis. 2001, 37, 693–710. [Google Scholar] [CrossRef] [Green Version]
- Hoekstra, P.F.; Braune, B.M.; Elkin, B.; Armstrong, F.A.J.; Muir, D.C.G. Concentrations of selected essential and non-essential elements in Arctic fox (Alopex lagopus) and wolverines (Gulo gulo) from the Canadian Arctic. Sci. Total Environ. 2003, 309, 81–92. [Google Scholar] [CrossRef]
- Rush, S.A.; Borgå, K.; Dietz, R.; Born, E.W.; Sonne, C.; Evans, T.; Muir, D.C.G.; Letcher, R.J.; Norstrom, R.J.; Fisk, A.T. Geographic distribution of selected elements in the livers of polar bears from Greenland, Canada and the United States. Environ. Pollut. 2008, 153, 618–626. [Google Scholar] [CrossRef]
- Dietz, R.; Riget, F.; Born, E.W. An assessment of selenium to mercury in Greenland marine animals. Sci. Total Environ. 2000, 245, 15–24. [Google Scholar] [CrossRef]
- Khan, M.A.K.; Wang, F. Mercury-selenium compounds and their toxicological significance: Toward a molecular understanding of the mercury-selenium antagonism. Environ. Toxicol. Chem. 2009, 28, 1567–1577. [Google Scholar] [CrossRef]
- Toscano, C.D.; Guilarte, T.R. Lead neurotoxicity: From exposure to molecular effects. Brain Res. Rev. 2005, 49, 529–554. [Google Scholar] [CrossRef]
- Słota, M.; Wąsik, M.; Stołtny, T.; Machoń-Grecka, A.; Kasperczyk, S. Effects of environmental and occupational lead toxicity and its association with iron metabolism. Toxicol. Appl. Pharmacol. 2022, 434, 115794. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Brown Bear (n = 114) | Grey Wolf (n = 8) | |||||
|---|---|---|---|---|---|---|
| Male | Female | Male | Female | |||
| Age (y) | C&Y | 1.0 (0.5, 1.0); 14 A | 0.9 (1.0, 1.0); 9 A | SUB | 1; 1 | 0.9 (0.6, 1.3); 4 |
| SUB | 2.0 (2.0, 3.0); 30 B | 3.0 (2.0, 3.0); 21 B | AD | 2.5 (2.4, 2.5); 2 | 5.5; 1 | |
| AD | 5.0 (4.0, 10.0); 30 C | 6.0 (5.0, 9.0); 10 C | ||||
| Body mass (kg) | C&Y | 54.8 (30.1, 75.0); 14 A | 67.4 (40.7, 81.0); 8 A | SUB | 20; 1 | 24.2 (20.6, 24.6); 4 |
| SUB | 135 (82.0, 190); 29 aB | 92.0 (71.0, 100); 21 bA | AD | 36.2 (35.0, 37.4); 2 | 28.3; 1 | |
| AD | 210 (179, 248); 30 aC | 130 (110, 185); 10 bB | ||||
| Body length (cm) | C&Y | 125 (117, 133); 14 A | 136 (114, 149); 8 A | SUB | 114; 1 | 107 (97.5, 117); 4 |
| SUB | 175 (162, 185); 30 aB | 149 (143, 158); 20 bB | AD | 123 (122, 124); 2 | 120; 1 | |
| AD | 187 (180, 200); 29 aC | 160 (154, 170); 9 bB | ||||
| BCI | C&Y | −0.09 (−0.49, 1.13); 14 | 0.61 (−0.99, 0.78); 8 | |||
| SUB | 0.32 (−0.77, 0.92); 30 | 0.55 (−0.73, 1.27); 20 | ||||
| AD | 0.68 (0.09, 1.19); 29 | 1.46 (0.76, 1.57); 9 | ||||
| Element | MDL | Tissue | Brown Bear | Grey Wolf | p |
|---|---|---|---|---|---|
| As (μg/kg) | 0.412 | brain | 1.52 (1.12, 2.56) 0.452–17.2 | 1.27 (0.88, 1.96) 0.556–2.63 | NS |
| liver | 8.88 (5.55, 15.3) 2.60–33.7 | 3.49 (2.42, 5.18) 1.53–17.2 | 0.003 | ||
| Cd (μg/kg) | 0.155 | brain | 3.40 (2.22, 5.19) 0.582–26.7 | 0.535 (0.289, 0.890) 0.167–3.36 | <0.001 |
| liver | 926 (495, 1918) 81.9–4093 | 38.2 (22.1, 71.6) 16.5–137 | <0.001 | ||
| Ca (mg/kg) | 0.667 | brain | 66.3 (56.5, 80.3) 45.6–271 | 124 (111, 154) 108–324 | <0.001 |
| liver | 40.7 (36.9, 52.2) 27.2–104 | 65 (51.1, 119) 28.0–302 | 0.02 | ||
| Cu (mg/kg) | 0.100 | brain | 2.59 (1.89, 3.31) 0.723–9.97 | 1.96 (1.22, 2.37) 1.09–2.81 | 0.02 |
| liver | 21.3 (15.1, 27.4) 3.93–66.4 | 8.13 (4.09, 14.7) 3.14–33.0 | 0.004 | ||
| Fe (mg/kg) | 0.068 | brain | 21.2 (16.7, 28.3) 9.20–134 | 33.8 (26.3, 41.9) 23.3–92.9 | 0.003 |
| liver | 131 (87.8, 190) 38.5–549 | 148 (110, 200) 99.8–222 | NS | ||
| Hg (μg/kg) | 0.668 | brain | 2.14 (1.57, 2.97) < MDL–12.3 | 2.04 (1.54, 2.89) 1.16–6.25 | NS |
| liver | 39.5 (18.2, 64.7) 8.64–499 | 7.95 (6.25, 18.1) 4.84–26.1 | <0.001 | ||
| Pb (μg/kg) | 0.454 | brain | 11.1 (7.13, 24.1) 3.45–372 | 7.68 (3.55, 11.8) 1.10–16.6 | 0.04 |
| liver | 590 (310, 831) 143–4046 | 120 (72.3, 167) 51.3–258 | <0.001 | ||
| Se (μg/kg) | 0.770 | brain | 112 (86.8, 130) 58.3–448 | 133 (117, 147) 107–150 | 0.03 |
| liver | 406 (328, 449) 249–689 | 489 (424, 564) 370–630 | 0.01 | ||
| Zn (mg/kg) | 0.042 | brain | 7.89 (6.71, 9.04) 4.85–34.2 | 7.54 (6.95, 8.36) 5.82–11.1 | NS |
| liver | 35.8 (32.4, 45.3) 21.1–93.3 | 26.1 (22.3, 29.4) 14.6–50.1 | 0.003 | ||
| Se/Hg | brain | 128 (94.9, 174) 26.2–1024 | 161 (119, 192) 58.9–328 | NS | |
| liver | 25.8 (13.1, 53.2) 2.96–123 | 144 (94.6, 197) 47.8–230 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lazarus, M.; Sekovanić, A.; Reljić, S.; Kusak, J.; Ferenčaković, M.; Sindičić, M.; Gomerčić, T.; Huber, Đ. Lead and Other Trace Element Levels in Brains of Croatian Large Terrestrial Carnivores: Influence of Biological and Ecological Factors. Toxics 2023, 11, 4. https://doi.org/10.3390/toxics11010004
Lazarus M, Sekovanić A, Reljić S, Kusak J, Ferenčaković M, Sindičić M, Gomerčić T, Huber Đ. Lead and Other Trace Element Levels in Brains of Croatian Large Terrestrial Carnivores: Influence of Biological and Ecological Factors. Toxics. 2023; 11(1):4. https://doi.org/10.3390/toxics11010004
Chicago/Turabian StyleLazarus, Maja, Ankica Sekovanić, Slaven Reljić, Josip Kusak, Maja Ferenčaković, Magda Sindičić, Tomislav Gomerčić, and Đuro Huber. 2023. "Lead and Other Trace Element Levels in Brains of Croatian Large Terrestrial Carnivores: Influence of Biological and Ecological Factors" Toxics 11, no. 1: 4. https://doi.org/10.3390/toxics11010004