

Selenomethionine and Allicin Synergistically Mitigate Intestinal Oxidative Injury by Activating the Nrf2 Pathway


Abstract

1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Allicin Preparation
2.3. Cell Culture
2.4. H2O2-Induced IPEC-J2 Cells: Oxidative Damage and SeMet + Allicin Treatment
2.4.1. Establishment of Oxidative Stress Model and Screening of Optimal Drug Combination
2.4.2. Determination of Combination Index (CI) Values
2.4.3. Cell Groups and Treatment
2.4.4. Measurement of ROS in IPEC-J2 Cells
2.4.5. Measurement of oxidative stress markers
2.4.6. Q-PCR
2.4.7. Western Blot
2.4.8. Wound Closure Assay
2.4.9. Transepithelial Electrical Resistance (TEER) and Permeability Assay
2.5. Diquat-Induced Intestinal Barrier Injury and SeMet + Allicin Treatment In Vivo
2.5.1. Establishment of Oxidative Stress Animal Model
2.5.2. Mice Groups and Treatment
2.5.3. Assessment of the Intestinal Permeability of FD4
2.5.4. SOD and MDA assay
2.5.5. Histopathological Examination
2.5.6. ROS Determination in Jejunum
2.5.7. Western Blot Analysis of Related Pathway Proteins in Jejunum Tissue
2.6. Statistical Analysis
3. Results
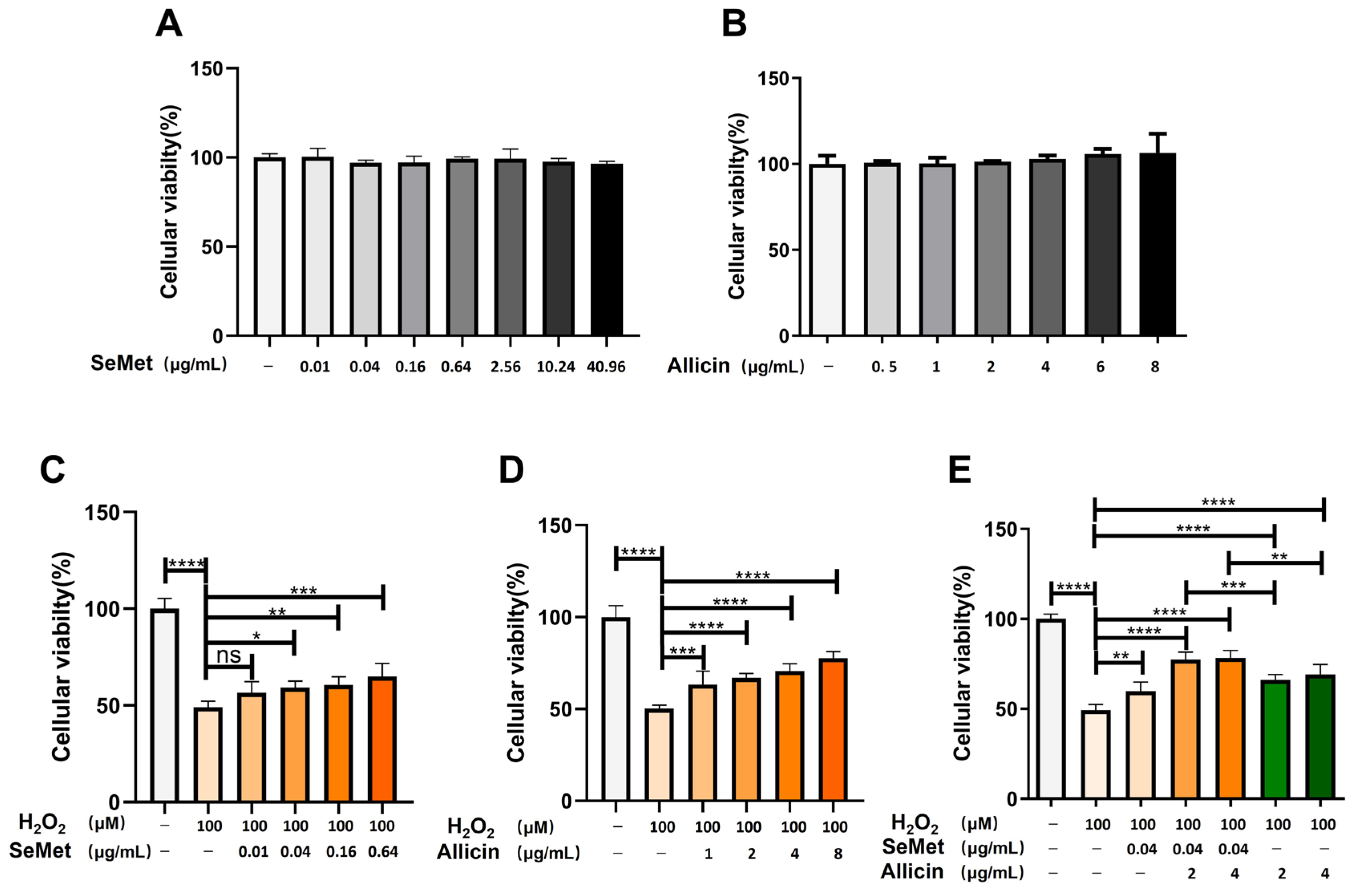
3.1. Candidate H2O2, SeMet or Allicin Dosage under Selection
3.2. SeMet, Allicin, or Both Alleviates H2O2-Induced IPEC-J2 Cell Damage
3.3. Synergistic Effect of SeMet and Allicin Combination
3.4. SeMet, Allicin, or Both Regulated the Generation of ROS and Redox Balance
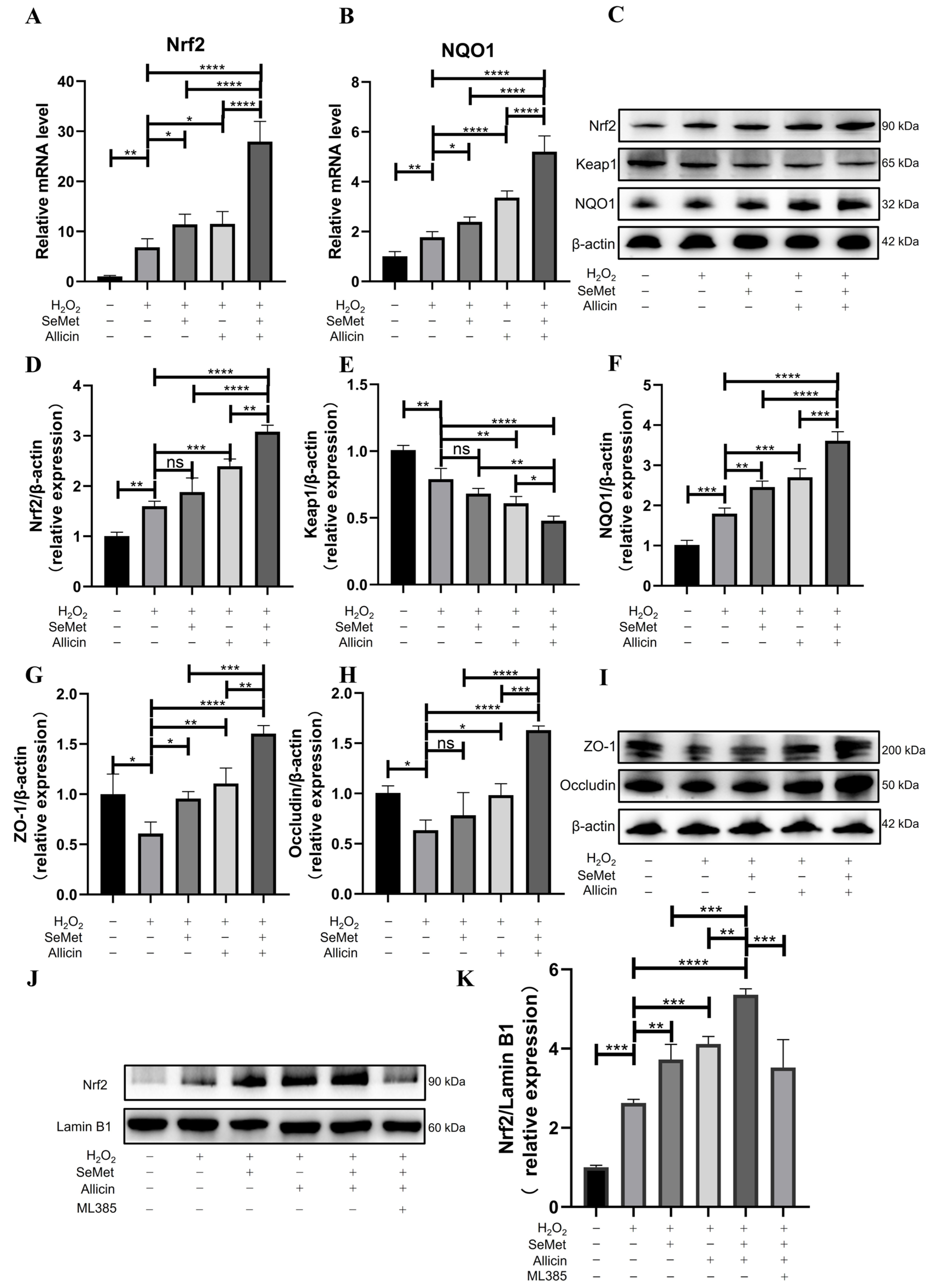
3.5. SeMet, Allicin, or Both Activate the Nrf2 Signaling Pathway
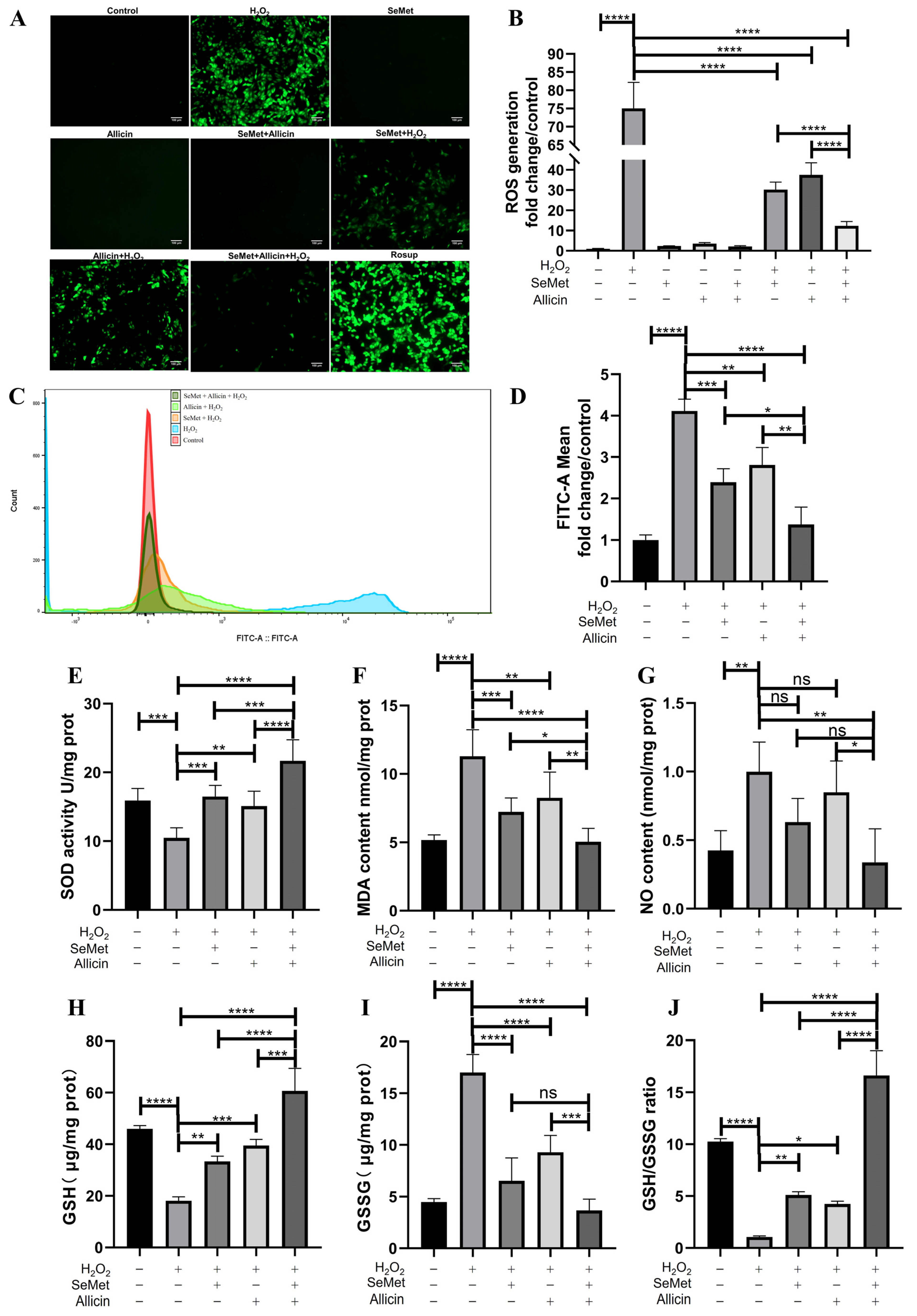
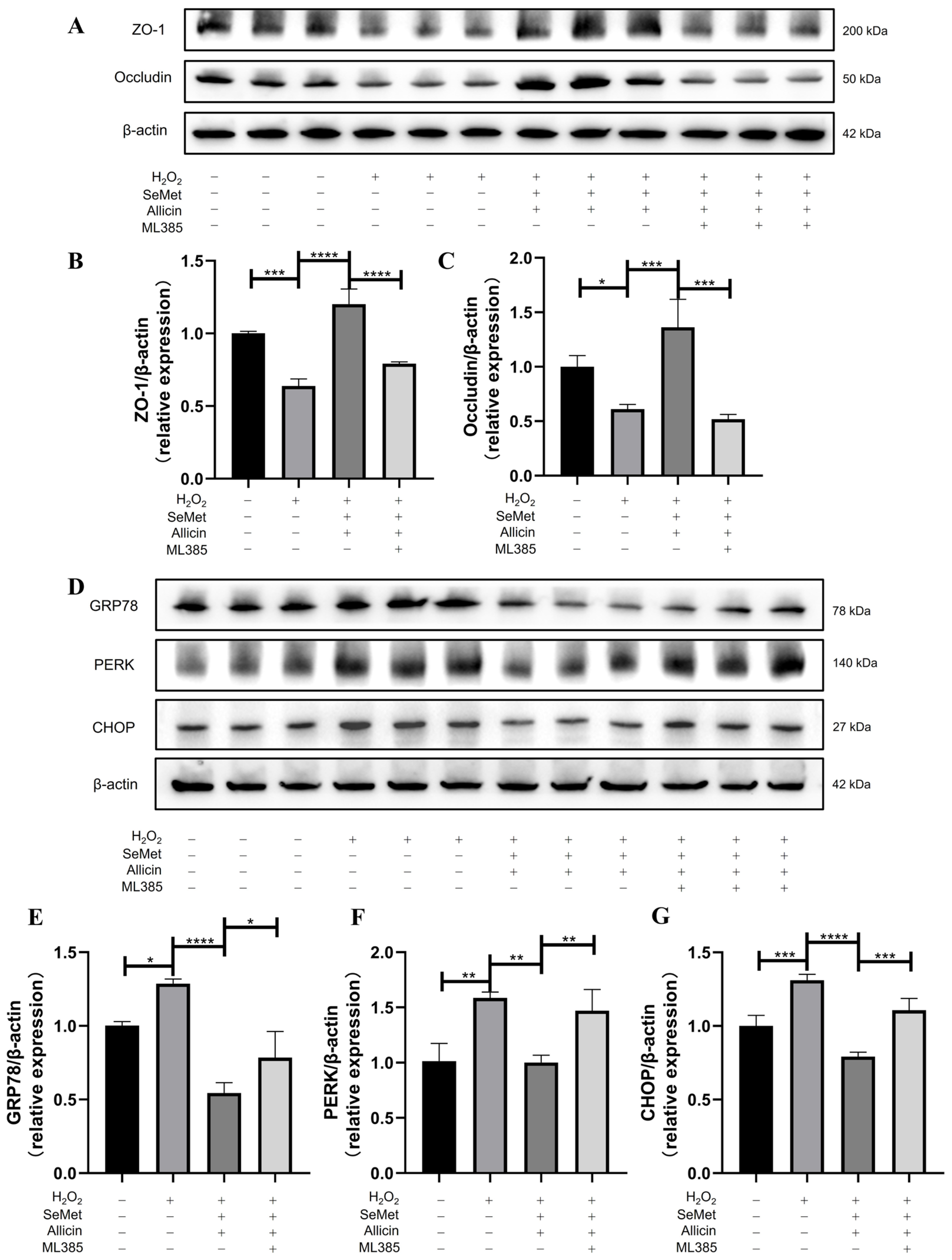
3.6. SeMet, Allicin, or Both Increase TJ Protein Expression
3.7. SeMet, Allicin, or Both Attenuate ERS
3.8. SeMet and Allicin Synergistically Activate Nrf2 Signaling Pathway
3.9. SeMet and Allicin Improve Cell Migration and Maintain Barrier Function Depending on Nrf2 Activation


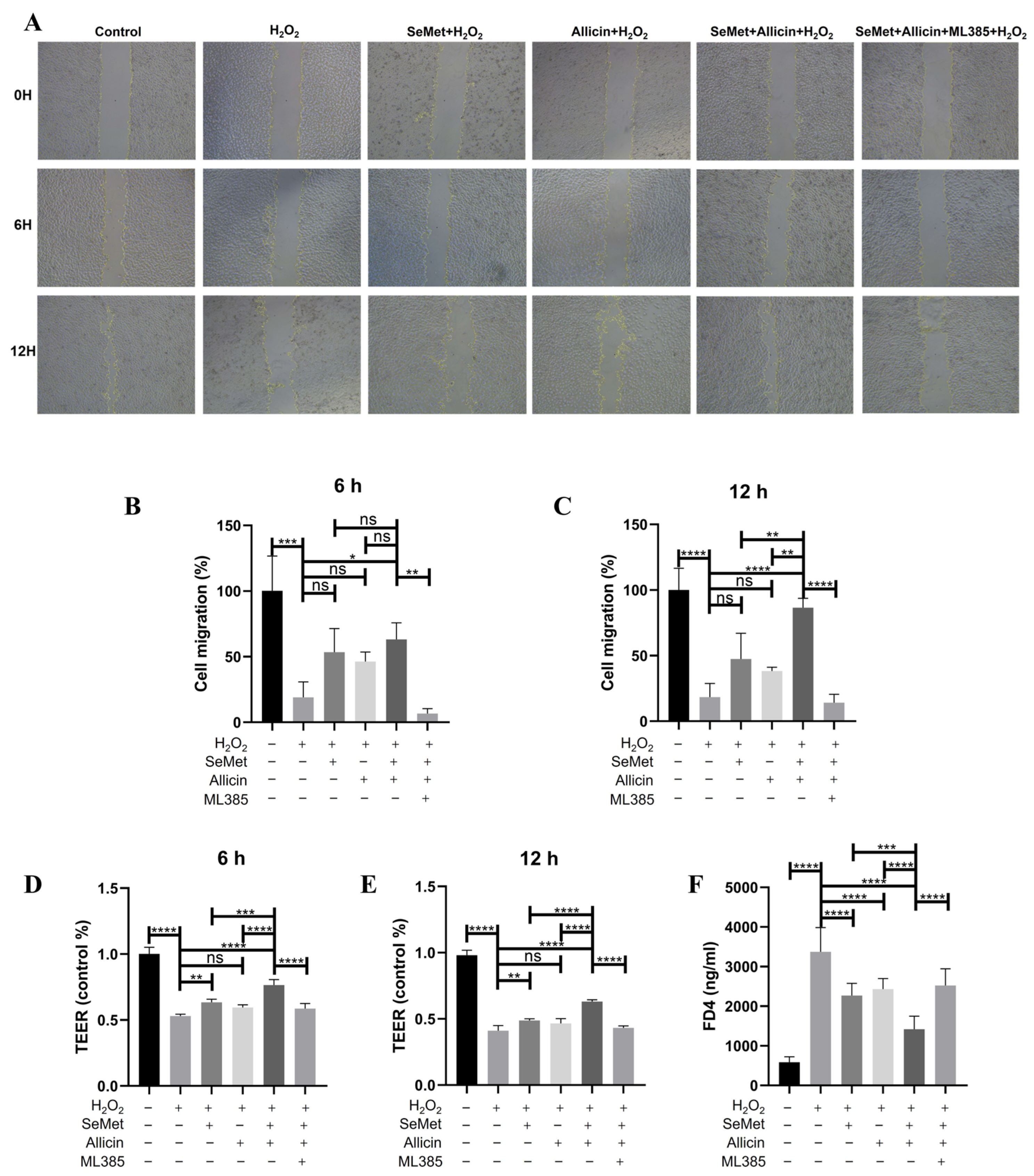
3.10. SeMet and Allicin Improve Tight Junction and Inhibit ER Stress Depends on Nrf2 Activation
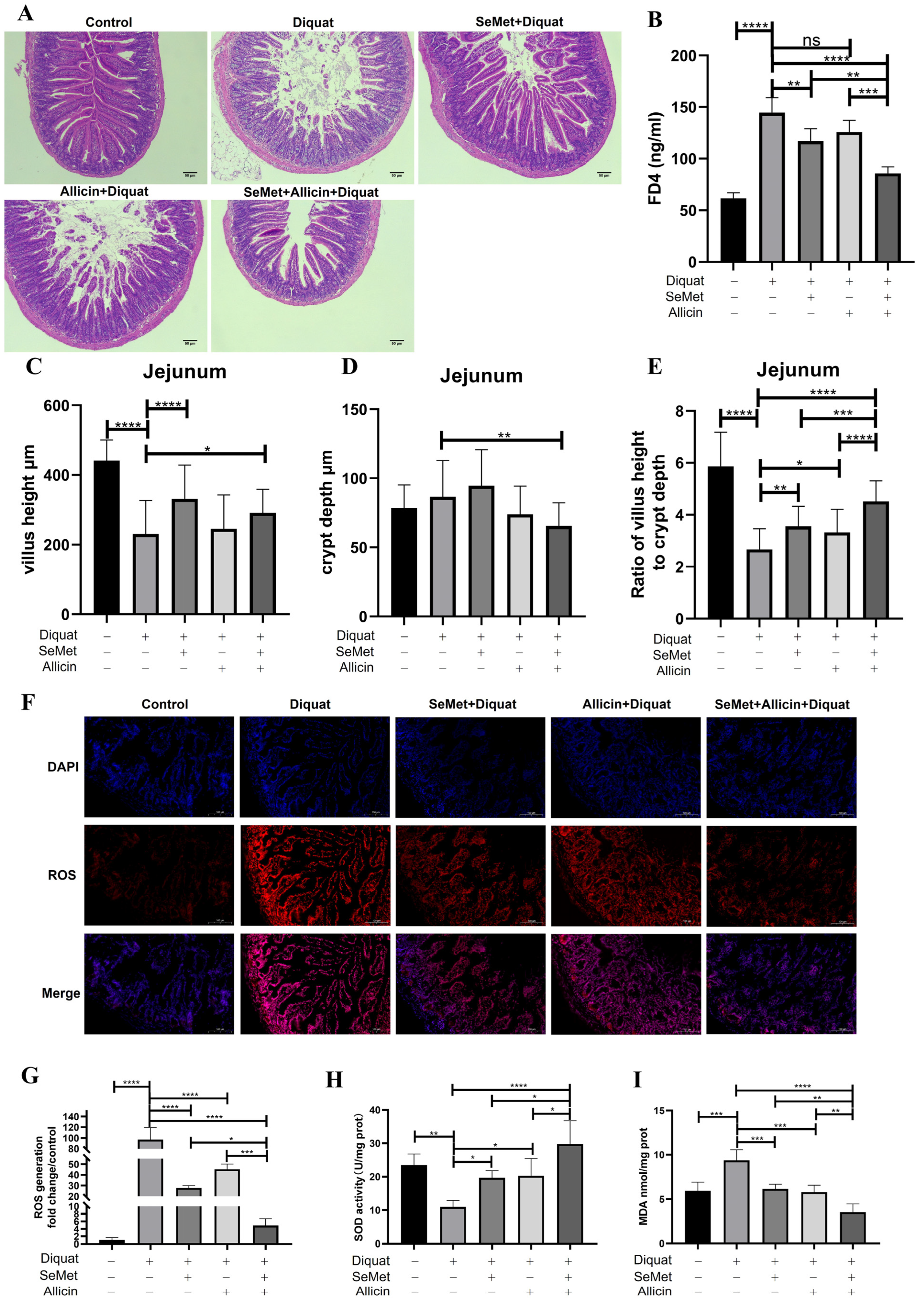
3.11. SeMet, Allicin, or Both Increased the Intestinal Function and Alleviated Intestinal Injury in Mice
3.12. SeMet, Allicin, or Both Improve Antioxidative Capacity in Jejunum of Mice
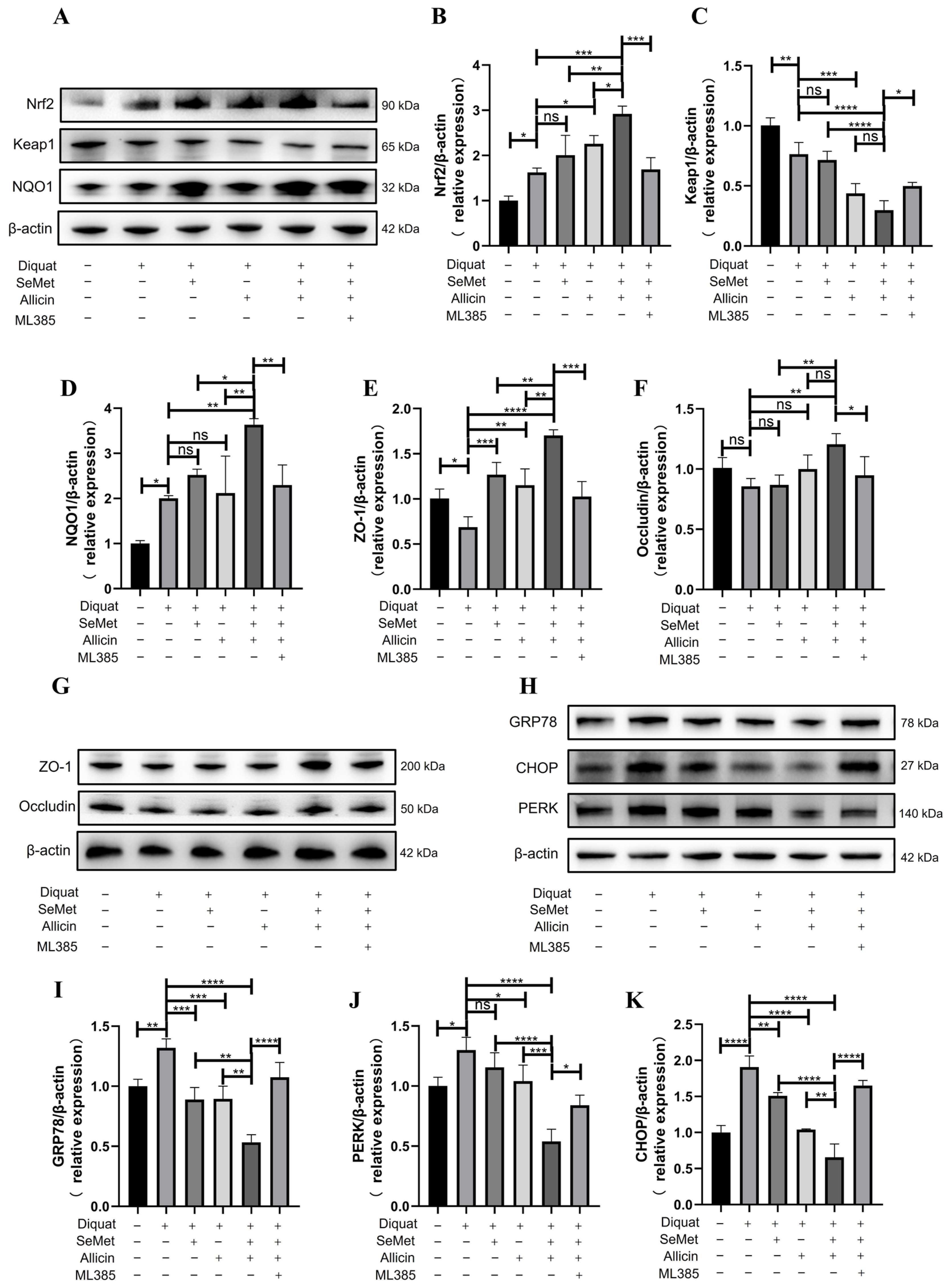
3.13. SeMet and Allicin Synergistically Activate the Nrf2 Pathway to Improve Barrier Function and Inhibit ER Stress in the Jejunum of Mice
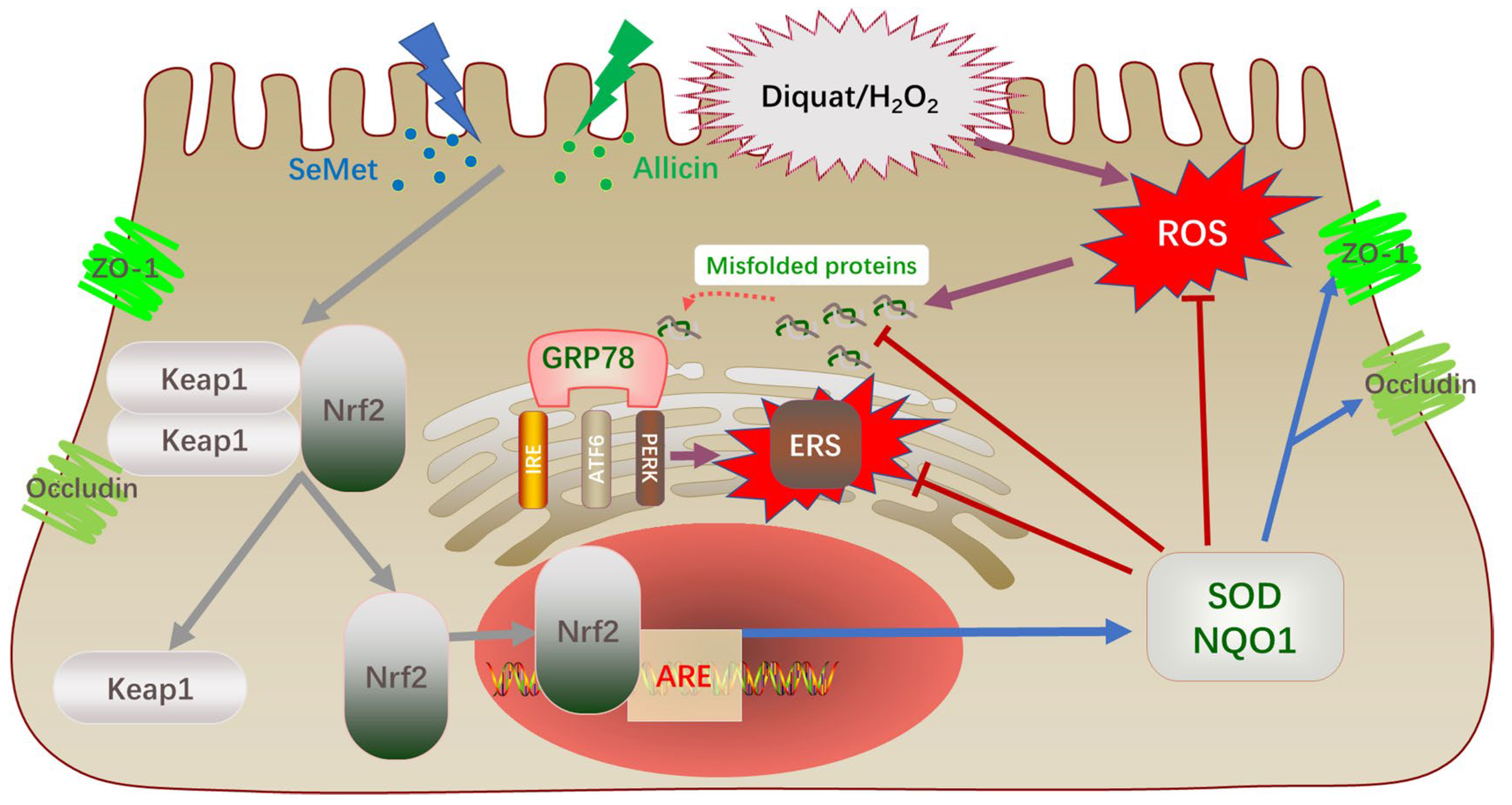
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Mou, D.; Ding, D.; Yang, M.; Jiang, X.; Zhao, L.; Che, L.; Fang, Z.; Xu, S.; Lin, Y.; Zhuo, Y.; et al. Maternal organic selenium supplementation during gestation improves the antioxidant capacity and reduces the inflammation level in the intestine of offspring through the NF-κB and ERK/Beclin-1 pathways. Food Funct. 2021, 12, 315–327. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, A.; Chanez-Paredes, S.D.; Haest, X.; Turner, J.R. Paracellular permeability and tight junction regulation in gut health and disease. Nat. Rev. Gastroenterol. Hepatol. 2023, 20, 417–432. [Google Scholar] [CrossRef] [PubMed]
- Oda, Y.; Takahashi, C.; Harada, S.; Nakamura, S.; Sun, D.; Kiso, K.; Urata, Y.; Miyachi, H.; Fujiyoshi, Y.; Honigmann, A.; et al. Discovery of anti-inflammatory physiological peptides that promote tissue repair by reinforcing epithelial barrier formation. Sci. Adv. 2021, 7, eabj6895. [Google Scholar] [CrossRef]
- Jin, Y.; Zhai, Z.; Jia, H.; Lai, J.; Si, X.; Wu, Z. Kaempferol attenuates diquat-induced oxidative damage and apoptosis in intestinal porcine epithelial cells. Food Funct. 2021, 12, 6889–6899. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Mo, S.; Jin, Y.; Zheng, Z.; Wang, H.; Wu, S.; Ren, Z.; Wu, J. Ammonia-induced excess ROS causes impairment and apoptosis in porcine IPEC-J2 intestinal epithelial cells. Ecotoxicol. Environ. Saf. 2022, 243, 114006. [Google Scholar] [CrossRef] [PubMed]
- Hartwick Bjorkman, S.; Oliveira Pereira, R. The Interplay Between Mitochondrial Reactive Oxygen Species, Endoplasmic Reticulum Stress, and Nrf2 Signaling in Cardiometabolic Health. Antioxid. Redox Signal. 2021, 35, 252–269. [Google Scholar] [CrossRef] [PubMed]
- Jaud, M.; Philippe, C.; Di Bella, D.; Tang, W.; Pyronnet, S.; Laurell, H.; Mazzolini, L.; Rouault-Pierre, K.; Touriol, C. Translational Regulations in Response to Endoplasmic Reticulum Stress in Cancers. Cells 2020, 9, 540. [Google Scholar] [CrossRef]
- Yang, Y.; Lu, D.; Wang, M.; Liu, G.; Feng, Y.; Ren, Y.; Sun, X.; Chen, Z.; Wang, Z. Endoplasmic reticulum stress and the unfolded protein response: Emerging regulators in progression of traumatic brain injury. Cell Death Dis. 2024, 15, 156. [Google Scholar] [CrossRef]
- Zhang, R.; Jiang, M.; Zhang, J.; Qiu, Y.; Li, D.; Li, S.; Liu, J.; Liu, C.; Fang, Z.; Cao, F. Regulation of the cerebrovascular smooth muscle cell phenotype by mitochondrial oxidative injury and endoplasmic reticulum stress in simulated microgravity rats via the PERK-eIF2α-ATF4-CHOP pathway. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165799. [Google Scholar] [CrossRef]
- Sun, R.; Deng, Z.; Han, X.; Zhang, Y.; Zhou, Y.; Shan, Y.; Fang, W.; Li, X. Porcine Circovirus 2 Manipulates the PERK-ERO1α Axis of the Endoplasmic Reticulum To Favor Its Replication by Derepressing Viral DNA from HMGB1 Sequestration within Nuclei. J. Virol. 2021, 95, e0100921. [Google Scholar] [CrossRef]
- Zhang, G.; Yang, W.; Jiang, F.; Zou, P.; Zeng, Y.; Ling, X.; Zhou, Z.; Cao, J.; Ao, L. PERK regulates Nrf2/ARE antioxidant pathway against dibutyl phthalate-induced mitochondrial damage and apoptosis dependent of reactive oxygen species in mouse spermatocyte-derived cells. Toxicol. Lett. 2019, 308, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Xing, S.; Chen, Y.; Liao, Q.; Li, Q.; Liu, Y.; He, S.; Feng, F.; Chen, Y.; Zhang, J.; et al. Reasonably activating Nrf2: A long-term, effective and controllable strategy for neurodegenerative diseases. Eur. J. Med. Chem. 2020, 185, 111862. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.; Dong, Y.; Liu, H.; Ren, H.; Cui, Z. Hesperetin protects against HO-triggered oxidative damage via upregulation of the Keap1-Nrf2/HO-1 signal pathway in ARPE-19 cells. Biomed. Pharmacother. 2017, 88, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.; Li, S.; Guo, W.; Yang, Y.; Zhang, W.; Zhang, Q.; He, Y.; Yi, X.; Cui, T.; An, Y.; et al. Simvastatin Protects Human Melanocytes from H2O2-Induced Oxidative Stress by Activating Nrf2. J. Investig. Dermatol. 2017, 137, 1286–1296. [Google Scholar] [CrossRef] [PubMed]
- Fão, L.; Mota, S.I.; Rego, A.C. Shaping the Nrf2-ARE-related pathways in Alzheimer’s and Parkinson’s diseases. Ageing Res. Rev. 2019, 54, 100942. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Cheng, Y.; Li, X.; Wang, F.; Lu, Z.; Xiao, X.; Wang, Y. Biogenic Nanoselenium Particles Effectively Attenuate Oxidative Stress-Induced Intestinal Epithelial Barrier Injury by Activating the Nrf2 Antioxidant Pathway. ACS Appl. Mater. Interfaces 2017, 9, 14724–14740. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P. The use of high-selenium yeast to raise selenium status: How does it measure up? Br. J. Nutr. 2004, 92, 557–573. [Google Scholar] [CrossRef]
- Gruhlke, M.C.H.; Antelmann, H.; Bernhardt, J.; Kloubert, V.; Rink, L.; Slusarenko, A.J. The human allicin-proteome: S-thioallylation of proteins by the garlic defence substance allicin and its biological effects. Free Radic. Biol. Med. 2019, 131, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Daim, M.M.; Abushouk, A.I.; Donia, T.; Alarifi, S.; Alkahtani, S.; Aleya, L.; Bungau, S.G. The nephroprotective effects of allicin and ascorbic acid against cisplatin-induced toxicity in rats. Environ. Sci. Pollut. Res. Int. 2019, 26, 13502–13509. [Google Scholar] [CrossRef]
- Borlinghaus, J.; Albrecht, F.; Gruhlke, M.C.H.; Nwachukwu, I.D.; Slusarenko, A.J. Allicin: Chemistry and biological properties. Molecules 2014, 19, 12591–12618. [Google Scholar] [CrossRef] [PubMed]
- Pandurangan, A.K.; Ismail, S.; Saadatdoust, Z.; Esa, N.M. Allicin Alleviates Dextran Sodium Sulfate- (DSS-) Induced Ulcerative Colitis in BALB/c Mice. Oxid. Med. Cell. Longev. 2015, 2015, 605208. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Sun, C.; Qiu, G.; Yao, L.; Yu, J.; Al Sberi, H.; Fouda, M.S.; Othman, M.S.; Lokman, M.S.; Kassab, R.B.; et al. Allicin mitigates hepatic injury following cyclophosphamide administration via activation of Nrf2/ARE pathways and through inhibition of inflammatory and apoptotic machinery. Environ. Sci. Pollut. Res. Int. 2021, 28, 39625–39636. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.; Dalton, C.; Hall, A.; Woodroofe, M.N.; Gardiner, P.H.E. Recent developments in selenium research. Br. J. Biomed. Sci. 2009, 66, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Nan, B.; Yang, C.; Li, L.; Ye, H.; Yan, H.; Wang, M.; Yuan, Y. Allicin alleviated acrylamide-induced NLRP3 inflammasome activation via oxidative stress and endoplasmic reticulum stress in Kupffer cells and SD rats liver. Food Chem. Toxicol. 2021, 148, 111937. [Google Scholar] [CrossRef]
- Alim, I.; Caulfield, J.T.; Chen, Y.; Swarup, V.; Geschwind, D.H.; Ivanova, E.; Seravalli, J.; Ai, Y.; Sansing, L.H.; Ste Marie, E.J.; et al. Selenium Drives a Transcriptional Adaptive Program to Block Ferroptosis and Treat Stroke. Cell 2019, 177, 1262–1279.e25. [Google Scholar] [CrossRef]
- Zhang, C.; Ge, J.; Lv, M.; Zhang, Q.; Talukder, M.; Li, J.-L. Selenium prevent cadmium-induced hepatotoxicity through modulation of endoplasmic reticulum-resident selenoproteins and attenuation of endoplasmic reticulum stress. Environ. Pollut. 2020, 260, 113873. [Google Scholar] [CrossRef]
- Ding, D.; Mou, D.; Zhu, H.; Jiang, X.; Che, L.; Fang, Z.; Xu, S.; Lin, Y.; Zhuo, Y.; Li, J.; et al. Maternal Organic Selenium Supplementation Relieves Intestinal Endoplasmic Reticulum Stress in Piglets by Enhancing the Expression of Glutathione Peroxidase 4 and Selenoprotein S. Front. Nutr. 2022, 9, 900421. [Google Scholar] [CrossRef]
- Ferro, C.; Florindo, H.F.; Santos, H.A. Selenium Nanoparticles for Biomedical Applications: From Development and Characterization to Therapeutics. Adv. Healthc. Mater. 2021, 10, e2100598. [Google Scholar] [CrossRef] [PubMed]
- Trevisani, M.; Siemens, J.; Materazzi, S.; Bautista, D.M.; Nassini, R.; Campi, B.; Imamachi, N.; Andrè, E.; Patacchini, R.; Cottrell, G.S.; et al. 4-Hydroxynonenal, an endogenous aldehyde, causes pain and neurogenic inflammation through activation of the irritant receptor TRPA1. Proc. Natl. Acad. Sci. USA 2007, 104, 13519–13524. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Pan, J.; Liu, H.; Jiao, Z. Characterization of the Synergistic Antioxidant Activity of Epigallocatechin Gallate (EGCG) and Kaempferol. Molecules 2023, 28, 5265. [Google Scholar] [CrossRef] [PubMed]
- Nisar, R.; Hanson, P.S.; He, L.; Taylor, R.W.; Blain, P.G.; Morris, C.M. Diquat causes caspase-independent cell death in SH-SY5Y cells by production of ROS independently of mitochondria. Arch. Toxicol. 2015, 89, 1811–1825. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Zhang, J.-J.; Zhang, Q.-B.; Wang, Z.-X.; Yin, Z.-N.; Li, X.-X.; Chen, J.; Ye, L.-M. Release test of alliin/alliinase double-layer tablet by HPLC-Allicin determination. J. Pharm. Anal. 2013, 3, 187–192. [Google Scholar] [CrossRef]
- Gong, Q.; Wang, X.; Liu, Y.; Yuan, H.; Ge, Z.; Li, Y.; Huang, J.; Liu, Y.; Chen, M.; Xiao, W.; et al. Potential Hepatoprotective Effects of Allicin on Carbon Tetrachloride-Induced Acute Liver Injury in Mice by Inhibiting Oxidative Stress, Inflammation, and Apoptosis. Toxics 2024, 12, 328. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, Z.; Ma, J.; Xv, Q.; Gao, H.; Yin, H.; Yan, G.; Jiang, X.; Yu, W. Lycopene attenuates the inflammation and apoptosis in aristolochic acid nephropathy by targeting the Nrf2 antioxidant system. Redox Biol. 2022, 57, 102494. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, V.; Sharda, N.; Huse, J.; Singh, D.; Sokolov, D.; Czinn, S.J.; Blanchard, T.G.; Banerjee, A. Synergistic potential of dual andrographolide and melatonin targeting of metastatic colon cancer cells: Using the Chou-Talalay combination index method. Eur. J. Pharmacol. 2021, 897, 173919. [Google Scholar] [CrossRef]
- Del Rio, B.; Redruello, B.; Linares, D.M.; Ladero, V.; Fernandez, M.; Martin, M.C.; Ruas-Madiedo, P.; Alvarez, M.A. The dietary biogenic amines tyramine and histamine show synergistic toxicity towards intestinal cells in culture. Food Chem. 2017, 218, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Li, Y.; Fan, L.; Sun, Y.; Gu, Z.; Xu, Q.-Q.; Wang, Y.; Xiong, L.; Xiao, K.; Chen, Z.-S.; et al. Bioactive Ceria Nanoenzymes Target Mitochondria in Reperfusion Injury to Treat Ischemic Stroke. ACS Nano 2024, 18, 5510–5529. [Google Scholar] [CrossRef]
- Xu, J.; Xu, J.; Shi, T.; Zhang, Y.; Chen, F.; Yang, C.; Guo, X.; Liu, G.; Shao, D.; Leong, K.W.; et al. Probiotic-Inspired Nanomedicine Restores Intestinal Homeostasis in Colitis by Regulating Redox Balance, Immune Responses, and the Gut Microbiome. Adv. Mater. 2023, 35, e2207890. [Google Scholar] [CrossRef]
- Song, H.; Qiu, J.; Yu, C.; Xiong, M.; Ou, C.; Ren, B.; Zhong, M.; Zeng, M.; Peng, Q. Traditional Chinese Medicine prescription Huang-Qi-Jian-Zhong-Tang ameliorates indomethacin-induced duodenal ulcers in rats by affecting NF-κB and STAT signaling pathways. Biomed. Pharmacother. 2022, 156, 113866. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zeng, L.; Wu, N.; Liang, Y.; Jin, J.; Fan, M.; Lai, X.; Chen, Z.-S.; Pan, Y.; Zeng, F.; et al. Inhibition of phosphoglycerate dehydrogenase induces ferroptosis and overcomes enzalutamide resistance in castration-resistant prostate cancer cells. Drug Resist. Updat. 2023, 70, 100985. [Google Scholar] [CrossRef]
- Li, K.; Xu, K.; He, Y.; Yang, Y.; Tan, M.; Mao, Y.; Zou, Y.; Feng, Q.; Luo, Z.; Cai, K. Oxygen Self-Generating Nanoreactor Mediated Ferroptosis Activation and Immunotherapy in Triple-Negative Breast Cancer. ACS Nano 2023, 17, 4667–4687. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.-W.; Zhang, Z.-S.; Zhou, Y.-F.; Zhou, T.-Y.; Sun, P.-L.; Zheng, B. Protective effect of seleno-amino-oligosaccharide on oxidative damage of IPEC-1 cells by activating Keap1/Nrf2 signaling pathway. Int. J. Biol. Macromol. 2020, 155, 972–978. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Gong, Z.; Liu, K.; Kou, J.; Liu, B.; Liu, K. Baicalin combats glutamate excitotoxicity via protecting glutamine synthetase from ROS-induced 20S proteasomal degradation. Redox Biol. 2020, 34, 101559. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Zhou, Y.; Cheng, X.; Chen, J.; Cao, H.; Guo, X.; Zhang, C.; Zhuang, Y.; Hu, G. Baicalin Attenuates H2O2-Induced Oxidative Stress by Regulating the AMPK/Nrf2 Signaling Pathway in IPEC-J2 Cells. Int. J. Mol. Sci. 2023, 24, 9435. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Song, G.; Shen, H.; Wu, Y.; Zhao, C.; Zhang, Z.; Jiang, Q.; Li, X.; Ma, X.; Tan, B.; et al. Allicin Improves Intestinal Epithelial Barrier Function and Prevents LPS-Induced Barrier Damages of Intestinal Epithelial Cell Monolayers. Front. Immunol. 2022, 13, 847861. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Zhong, H.; Li, T.; Wang, Z.; Chen, X.; Zou, T.; You, J.; Chen, J. Selenomethionine Alleviates Deoxynivalenol-Induced Oxidative Injury in Porcine Intestinal Epithelial Cells Independent of MAPK Pathway Regulation. Antioxidants 2024, 13, 356. [Google Scholar] [CrossRef] [PubMed]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef]
- Masuda, J.; Umemura, C.; Yokozawa, M.; Yamauchi, K.; Seko, T.; Yamashita, M.; Yamashita, Y. Dietary Supplementation of Selenoneine-Containing Tuna Dark Muscle Extract Effectively Reduces Pathology of Experimental Colorectal Cancers in Mice. Nutrients 2018, 10, 356. [Google Scholar] [CrossRef]
- Gao, Z.; Chen, J.; Qiu, S.; Li, Y.; Wang, D.; Liu, C.; Li, X.; Hou, R.; Yue, C.; Liu, J.; et al. Optimization of selenylation modification for garlic polysaccharide based on immune-enhancing activity. Carbohydr. Polym. 2016, 136, 560–569. [Google Scholar] [CrossRef]
- Campo-Sabariz, J.; Moral-Anter, D.; Brufau, M.T.; Briens, M.; Pinloche, E.; Ferrer, R.; Martín-Venegas, R. 2-Hydroxy-(4-methylseleno)butanoic Acid Is Used by Intestinal Caco-2 Cells as a Source of Selenium and Protects against Oxidative Stress. J. Nutr. 2019, 149, 2191–2198. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Nan, B.; Wu, X.; Yan, H.; Yuan, Y. Allicin alleviates acrylamide-induced oxidative stress in BRL-3A cells. Life Sci. 2019, 231, 116550. [Google Scholar] [CrossRef]
- Xiang, Q.; Cheng, Z.; Wang, J.; Feng, X.; Hua, W.; Luo, R.; Wang, B.; Liao, Z.; Ma, L.; Li, G.; et al. Allicin Attenuated Advanced Oxidation Protein Product-Induced Oxidative Stress and Mitochondrial Apoptosis in Human Nucleus Pulposus Cells. Oxid. Med. Cell. Longev. 2020, 2020, 6685043. [Google Scholar] [CrossRef] [PubMed]
- Krug, P.; Mielczarek, L.; Wiktorska, K.; Kaczyńska, K.; Wojciechowski, P.; Andrzejewski, K.; Ofiara, K.; Szterk, A.; Mazur, M. Sulforaphane-conjugated selenium nanoparticles: Towards a synergistic anticancer effect. Nanotechnology 2019, 30, 065101. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wang, W.; Shan, Y.; Barrera, L.N.; Howie, A.F.; Beckett, G.J.; Wu, K.; Bao, Y. Synergy between sulforaphane and selenium in the up-regulation of thioredoxin reductase and protection against hydrogen peroxide-induced cell death in human hepatocytes. Food Chem. 2012, 133, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Dacosta, C.; Wang, W.; Zhou, Z.; Liu, M.; Bao, Y. Synergy between sulforaphane and selenium in protection against oxidative damage in colonic CCD841 cells. Nutr. Res. 2015, 35, 610–617. [Google Scholar] [CrossRef] [PubMed]
- Telorack, M.; Meyer, M.; Ingold, I.; Conrad, M.; Bloch, W.; Werner, S. A Glutathione-Nrf2-Thioredoxin Cross-Talk Ensures Keratinocyte Survival and Efficient Wound Repair. PLoS Genet. 2016, 12, e1005800. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-L.; Liu, X.-H.; Qiao, Y.; Ning, L.-N.; Li, W.-J.; Sun, Y.-S.; Liu, D.-S.; Gao, W.; Ma, C.-M. Allicin alleviates inflammation of diabetic macroangiopathy via the Nrf2 and NF-kB pathway. Eur. J. Pharmacol. 2020, 876, 173052. [Google Scholar] [CrossRef]
- Li, K.; Cao, Z.; Guo, Y.; Tong, C.; Yang, S.; Long, M.; Li, P.; He, J. Selenium Yeast Alleviates Ochratoxin A-Induced Apoptosis and Oxidative Stress via Modulation of the PI3K/AKT and Nrf2/Keap1 Signaling Pathways in the Kidneys of Chickens. Oxid. Med. Cell. Longev. 2020, 2020, 4048706. [Google Scholar] [CrossRef]
- Bouvier, E.; Brouillard, F.; Molet, J.; Claverie, D.; Cabungcal, J.H.; Cresto, N.; Doligez, N.; Rivat, C.; Do, K.Q.; Bernard, C.; et al. Nrf2-dependent persistent oxidative stress results in stress-induced vulnerability to depression. Mol. Psychiatry 2017, 22, 1701–1713. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.-X.; Yang, C.; Liu, Z.-J.; Su, H.-Y.; Zhang, W.-H.; Pan, Q.; Liu, H.-F. Protection of procyanidin B2 on mitochondrial dynamics in sepsis associated acute kidney injury via promoting Nrf2 nuclear translocation. Aging 2020, 12, 15638–15655. [Google Scholar] [CrossRef]
- Singh, A.; Venkannagari, S.; Oh, K.H.; Zhang, Y.-Q.; Rohde, J.M.; Liu, L.; Nimmagadda, S.; Sudini, K.; Brimacombe, K.R.; Gajghate, S.; et al. Small Molecule Inhibitor of NRF2 Selectively Intervenes Therapeutic Resistance in KEAP1-Deficient NSCLC Tumors. ACS Chem. Biol. 2016, 11, 3214–3225. [Google Scholar] [CrossRef]
- Liu, X.; Zhu, Q.; Zhang, M.; Yin, T.; Xu, R.; Xiao, W.; Wu, J.; Deng, B.; Gao, X.; Gong, W.; et al. Isoliquiritigenin Ameliorates Acute Pancreatitis in Mice via Inhibition of Oxidative Stress and Modulation of the Nrf2/HO-1 Pathway. Oxid. Med. Cell. Longev. 2018, 2018, 7161592. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.-S.; Xue, R.; Du, M.; Tang, Z.; Xiang, X.-W.; Zheng, B.; Qu, Y.-L. Hemp seed polysaccharides protect intestinal epithelial cells from hydrogen peroxide-induced oxidative stress. Int. J. Biol. Macromol. 2019, 135, 203–211. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene Accession Number | Primer Sequence 5′-3′ |
|---|---|---|
| Nrf2 | XM_021075133.1 | F: CACCACCTCAGGGTAATA |
| R: GACAAACATTCAAGCCGC | ||
| NQO1 | XM_047791619.1 | F: CCAGCAGCCCGGCCAATCTG |
| R: AGGTCCGACACGGCGACCTC | ||
| GRP78 | X92446.1 | F: AATGGCCGTGTGGAGATCA |
| R: GAGCTGGTTCTTGGCTGCAT | ||
| IRE1 | XM_047757844.1 | F: CTGCACTCCCTCAACATCGT |
| R: GTAGGTGGGGTTCTCCTTGC | ||
| PERK | XM_003124925.4 | F: ACTACAAGCGGGAAAGGAGC |
| R: CACCAGTGCAAAAGGAGCAC | ||
| ATF6 | XM_021089516.1 | F: GCTCCTCCGTTCCTCCTTACCTC |
| R: CTGACAACATGGGCTGCCTCTG | ||
| XBP1 | XM_012934292.1 | F: TGAAGAGGAGGCAGAGACCAAGG |
| R: GGGAGATGTTCTGGAGGGGTGAC | ||
| ATF4 | XM_021090887.1 | F: GCCCCCGCAGATAGTGAA |
| R: TGGGGAAAGGGGAAGAGTTTG | ||
| CHOP | XM_008978498.4 | F: TCTGGCTTGGCTGACTGAGGAG |
| R: CCGTTTCCTGGGTCTTCTTTGGTC | ||
| β-actin | XM_003124280 | F: GATGAGATTGGCATGGCTTT |
| R: CACCTTCACCGTTCCAGTTT |
| Compound | Dose SeMet (μg/mL) | Dose Allicin (μg/mL) | CI |
|---|---|---|---|
| SeMet + allicin | 0.04 | 1 | 0.672 ± 0.495 |
| 2 | 0.263 ± 0.232 | ||
| 4 | 0.402 ± 0.307 | ||
| 8 | 0.584 ± 0.560 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Lv, X.; Yuan, H.; Wang, X.; Huang, J.; Wang, L. Selenomethionine and Allicin Synergistically Mitigate Intestinal Oxidative Injury by Activating the Nrf2 Pathway. Toxics 2024, 12, 719. https://doi.org/10.3390/toxics12100719
Liu Y, Lv X, Yuan H, Wang X, Huang J, Wang L. Selenomethionine and Allicin Synergistically Mitigate Intestinal Oxidative Injury by Activating the Nrf2 Pathway. Toxics. 2024; 12(10):719. https://doi.org/10.3390/toxics12100719
Chicago/Turabian StyleLiu, Yongshi, Xi Lv, Heling Yuan, Xiaoming Wang, Jinhu Huang, and Liping Wang. 2024. "Selenomethionine and Allicin Synergistically Mitigate Intestinal Oxidative Injury by Activating the Nrf2 Pathway" Toxics 12, no. 10: 719. https://doi.org/10.3390/toxics12100719
APA StyleLiu, Y., Lv, X., Yuan, H., Wang, X., Huang, J., & Wang, L. (2024). Selenomethionine and Allicin Synergistically Mitigate Intestinal Oxidative Injury by Activating the Nrf2 Pathway. Toxics, 12(10), 719. https://doi.org/10.3390/toxics12100719