

Developmental Toxicity of PEDOT:PSS in Zebrafish: Effects on Morphology, Cardiac Function, and Intestinal Health


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Husbandry and Zebrafish Egg Production
2.3. Chemicals
2.4. Measurement of Zebrafish Developmental Toxicity and LC50 of PEDOT:PSS
2.5. Measurement of Heart Rate and Other Basic Parameters
2.6. Behavioral Experiments
2.7. Expression Level Analysis of Oxidative Stress-Related Genes
2.8. In Situ Hybridization Experiment
2.9. Histocytological Analysis of Zebrafish Larvae
2.10. Transcriptomic Analysis
2.11. Statistical Analysis
3. Results
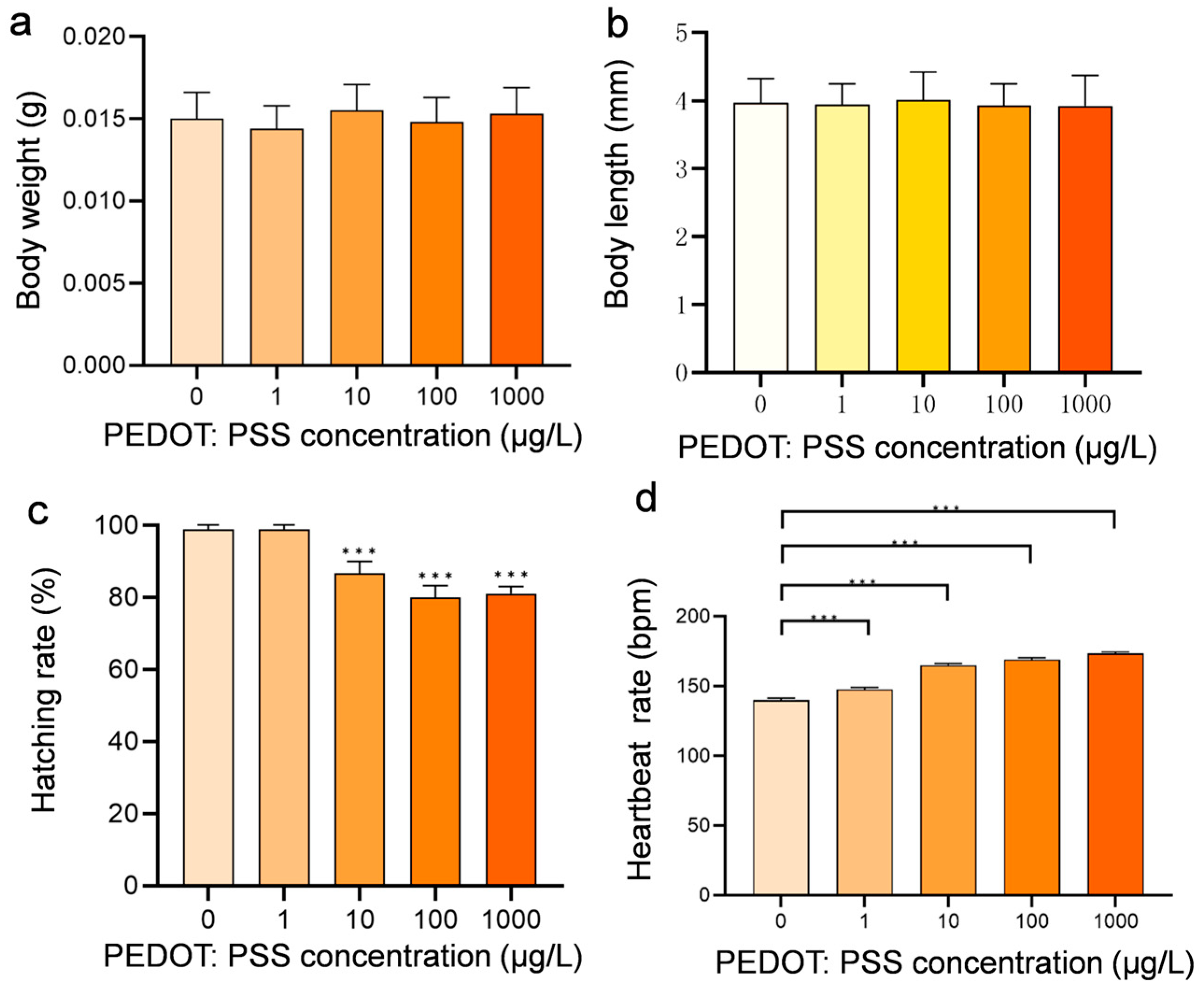
3.1. Developmental Toxicity Effects after Exposure to PEDOT:PSS
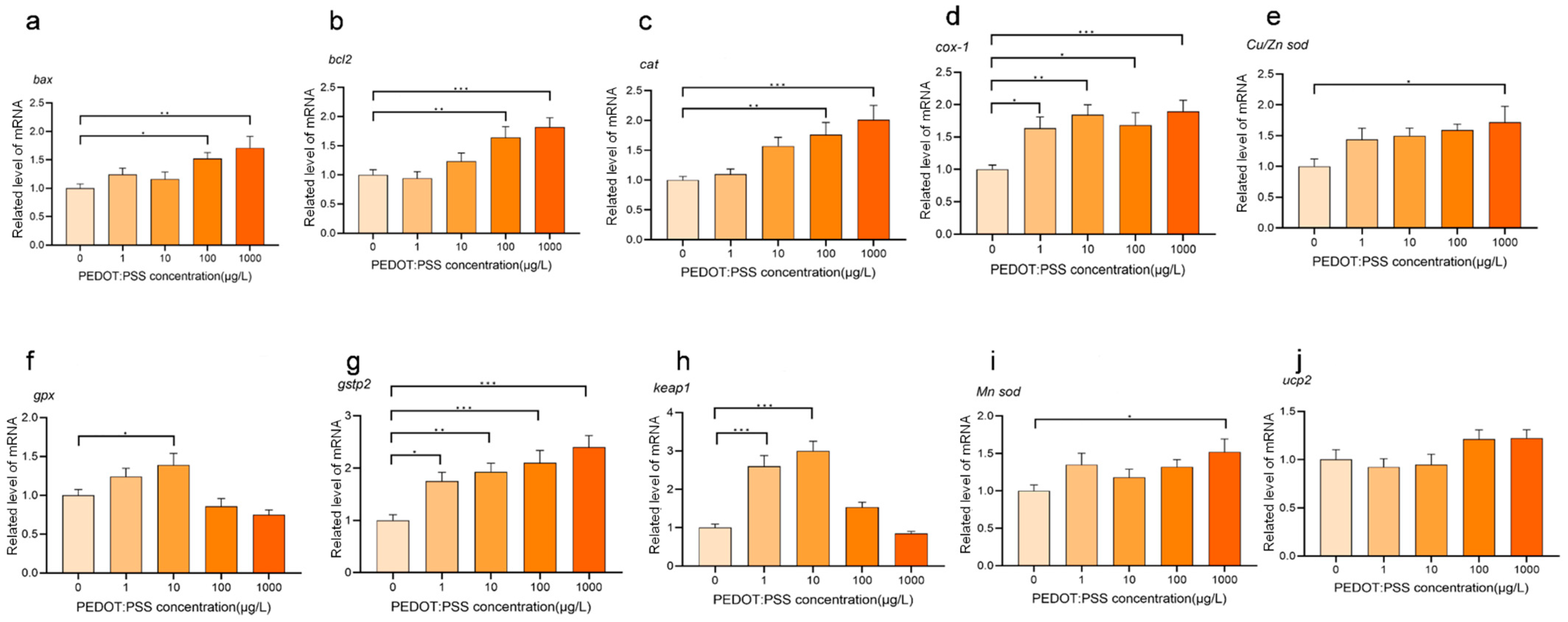
3.2. Effects of PEDOT:PSS on Oxidative Stress-Related Gene Expression
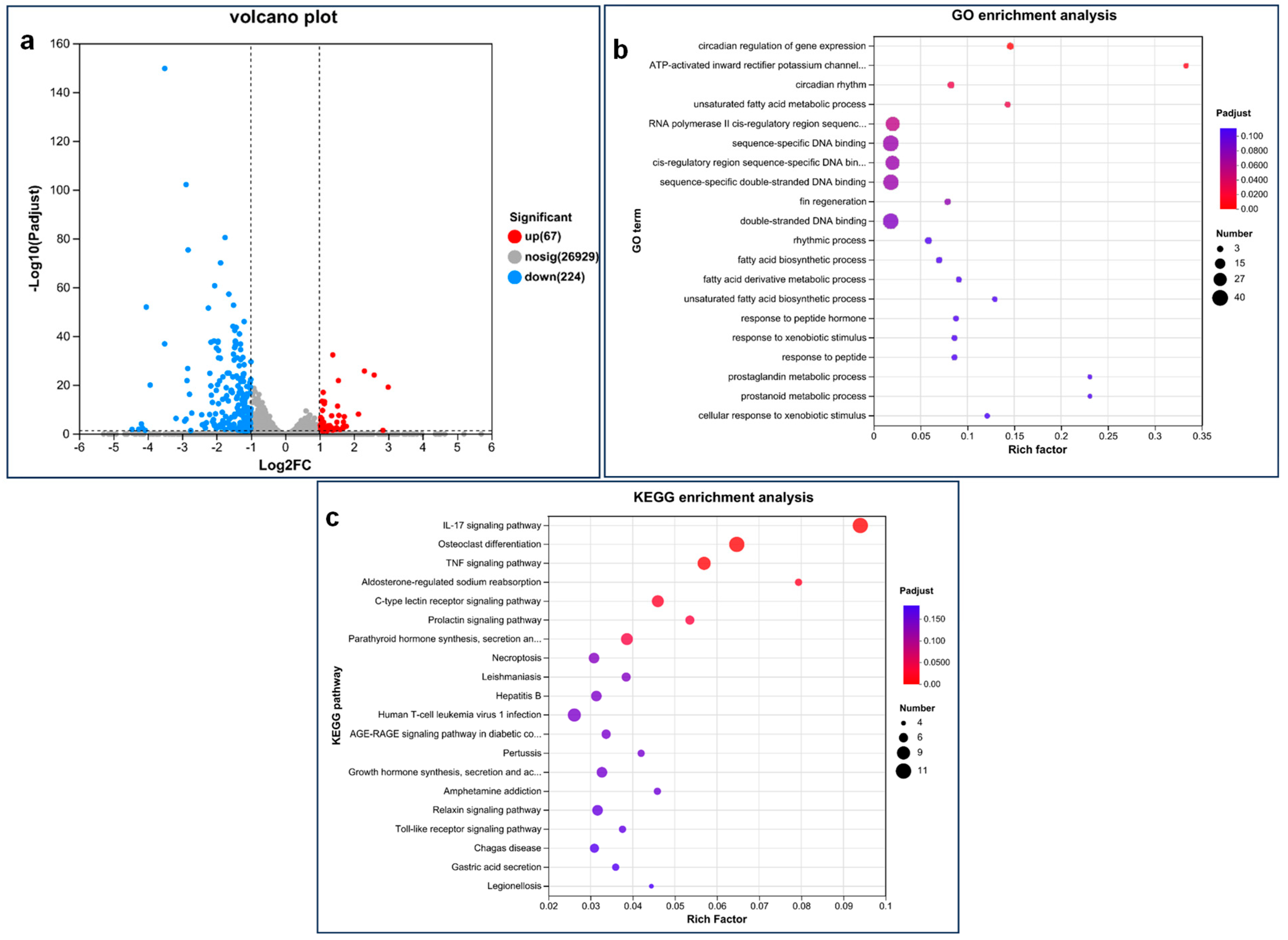
3.3. Transcriptomic Analysis of Zebrafish Larvae after Exposure to PEDOT:PSS
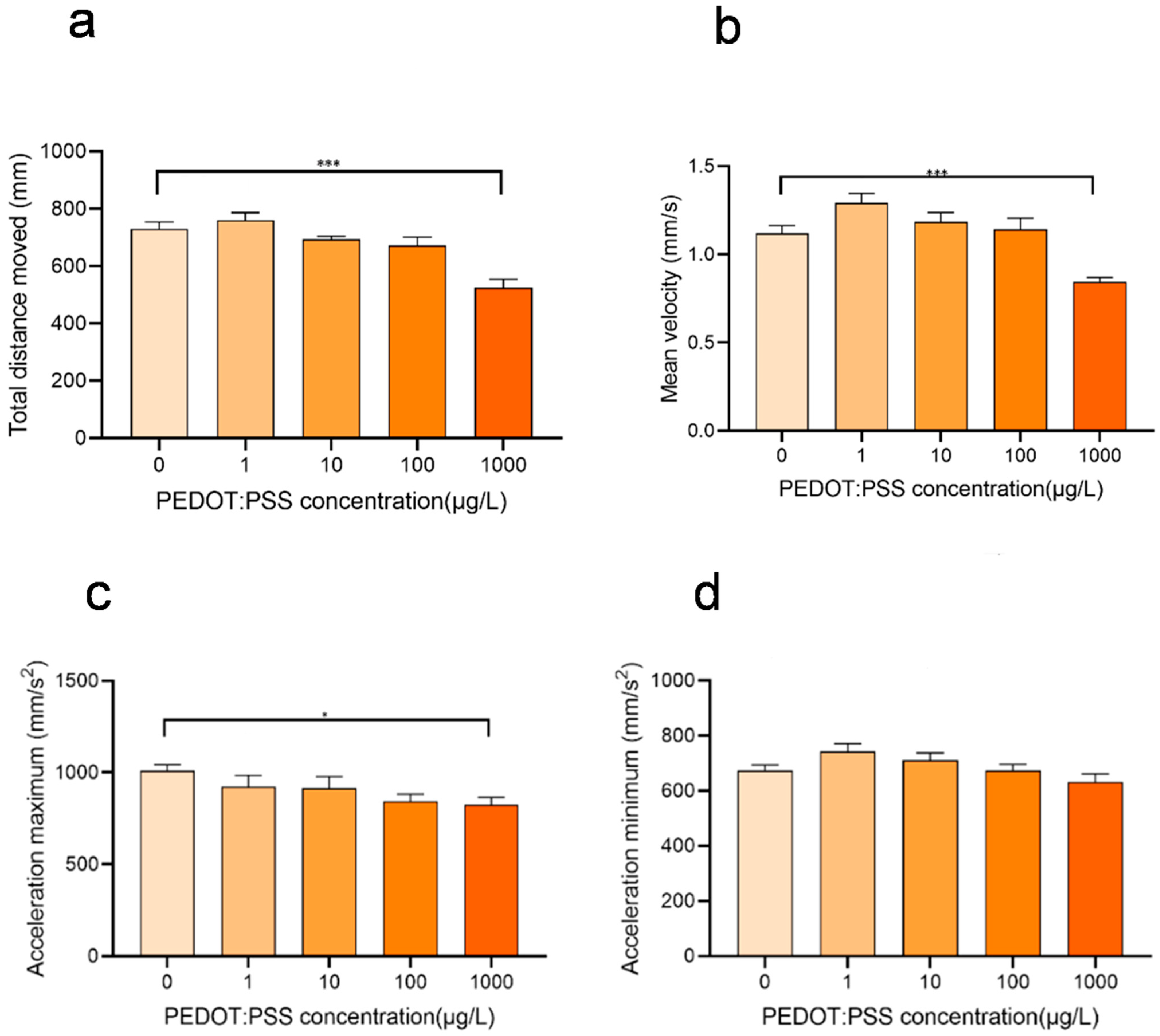
3.4. Concentration-Dependent Effects of PEDOT:PSS on Zebrafish Behavior
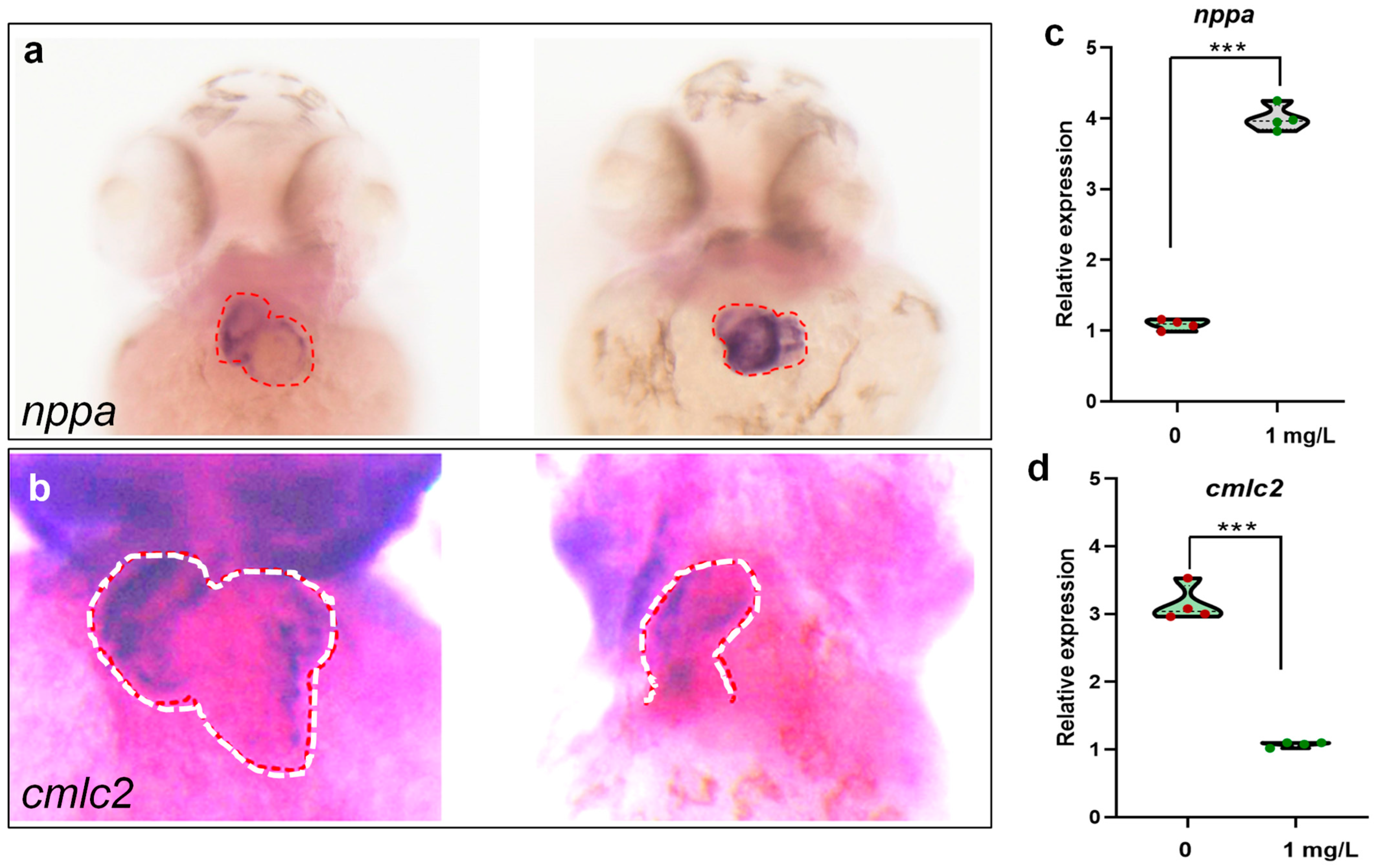
3.5. PEDOT:PSS Exposure Impairs Cardiac Development
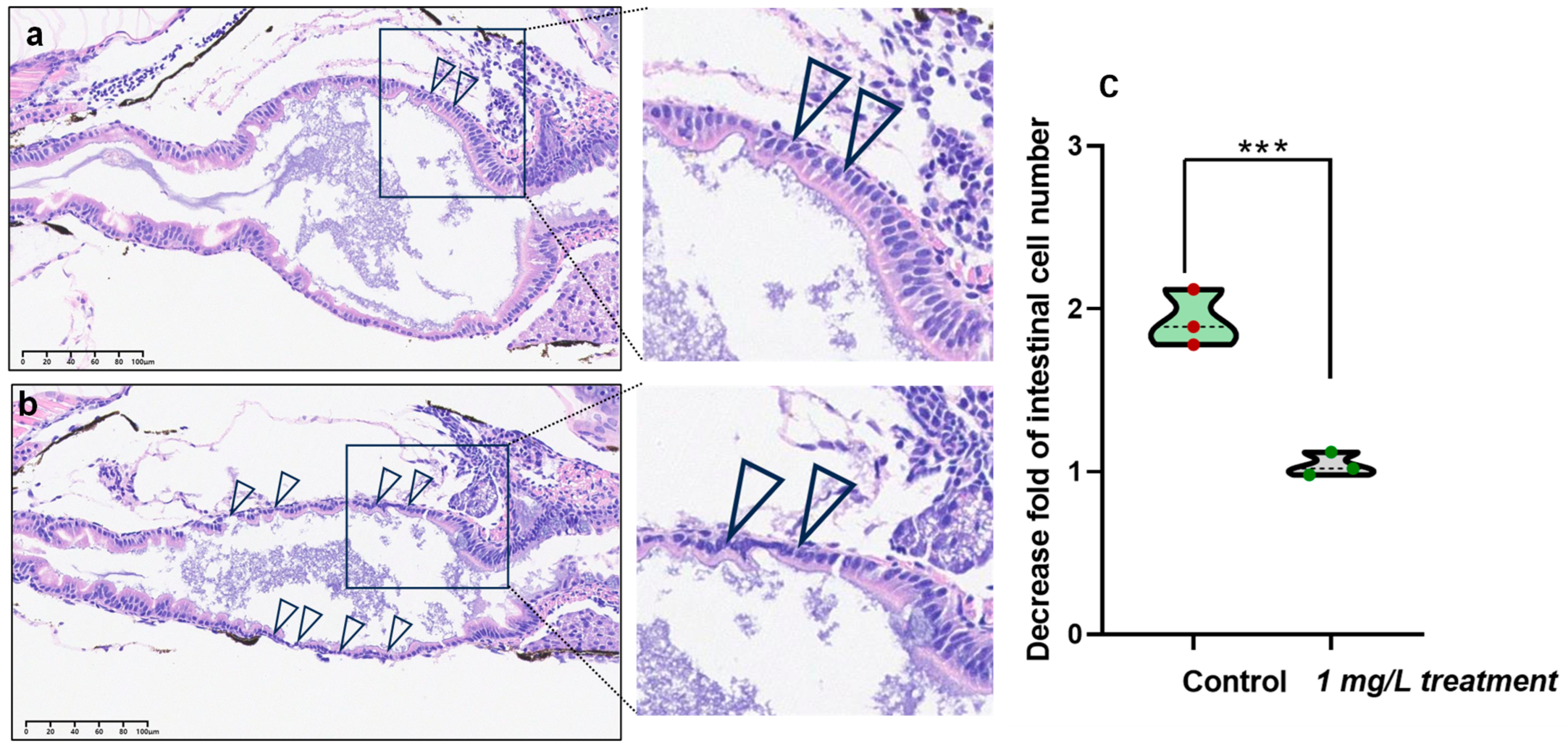
3.6. Detrimental Effects of PEDOT:PSS on Intestinal Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Conti, J.; Holtberg, P.; Diefenderfer, J.; Larose, A.; Turnure, J.T.; Westfall, L. International Energy Outlook 2016 with Projections to 2040; USDOE Energy Information Administration (EIA): Washington, DC, USA, 2016.
- Panigrahy, S.; Kandasubramanian, B. Polymeric thermoelectric PEDOT: PSS & composites: Synthesis, progress, and applications. Eur. Polym. J. 2020, 132, 109726. [Google Scholar]
- Abas, A.; Sheng, H.; Ma, Y.; Zhang, X.; Wei, Y.; Su, Q.; Lan, W.; Xie, E. PEDOT:PSS coated CuO nanowire arrays grown on Cu foam for high-performance supercapacitor electrodes. J. Mater. Sci. Mater. Electron. 2019, 30, 10953–10960. [Google Scholar] [CrossRef]
- Gong, C.; Cheng, S.; Meng, X.; Hu, X.; Chen, Y. Recent Advances of PEDOT in Flexible Energy Conversion and Storage Devices. Acta Chim. Sin. 2021, 79, 853–868. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, W.; Zhang, H.; Xie, M.; Duan, X. PEDOT:PSS: From conductive polymers to sensors. Nanotechnol. Precis. Eng. 2021, 4, 045004. [Google Scholar] [CrossRef]
- Gupta, S.; Datt, R.; Mishra, A.; Tsoi, W.C.; Patra, A.; Bober, P. Poly(3,4-ethylenedioxythiophene):Poly(styrene sulfonate) in antibacterial, tissue engineering and biosensors applications: Progress, challenges and perspectives. J. Appl. Polym. Sci. 2022, 139, e52663. [Google Scholar] [CrossRef]
- Liang, Y.; Offenhausser, A.; Ingebrandt, S.; Mayer, D. PEDOT:PSS-Based Bioelectronic Devices for Recording and Modulation of Electrophysiological and Biochemical Cell Signals. Adv. Healthc. Mater. 2021, 10, 2100061. [Google Scholar] [CrossRef]
- Zhang, W.; Su, Z.; Zhang, X.; Wang, W.; Li, Z. Recent progress on PEDOT-based wearable bioelectronics. View 2022, 3, 20220030. [Google Scholar] [CrossRef]
- Fan, Z.; Ouyang, J. Thermoelectric Properties of PEDOT:PSS. Adv. Electron. Mater. 2019, 5, 1800769. [Google Scholar] [CrossRef]
- Hung, J.-N.; Vo, D.N.K.; Ho, H.P.T.; Tsai, M.-H. PEDOT:PSS in Solution Form Exhibits Strong Potential in Inhibiting SARS-CoV-2 Infection of the Host Cells by Targeting Viruses and Also the Host Cells. Biomacromolecules 2022, 23, 3535–3548. [Google Scholar] [CrossRef]
- Zhu, J.; Omura, T.; Wakisaka, M. Biological response of protists Haematococcus lacustris and Euglena gracilis to conductive polymer poly (3,4-ethylenedioxythiophene) polystyrene sulfonate. Lett. Appl. Microbiol. 2021, 72, 619–625. [Google Scholar] [CrossRef]
- Wixon, J. Danio rerio, the zebrafish. Yeast 2000, 17, 225–231. [Google Scholar] [CrossRef]
- Cassar, S.; Adatto, I.; Freeman, J.L.; Gamse, J.T.; Iturria, I.; Lawrence, C.; Muriana, A.; Peterson, R.T.; Van Cruchten, S.; Zon, L.I. Use of Zebrafish in Drug Discovery Toxicology. Chem. Res. Toxicol. 2020, 33, 95–118. [Google Scholar] [CrossRef]
- Choi, T.-Y.; Choi, T.-I.; Lee, Y.-R.; Choe, S.-K.; Kim, C.-H. Zebrafish as an animal model for biomedical research. Exp. Mol. Med. 2021, 53, 310–317. [Google Scholar] [CrossRef]
- Adhish, M.; Manjubala, I. Effectiveness of zebrafish models in understanding human diseases—A review of models. Heliyon 2023, 9, e14557. [Google Scholar] [CrossRef]
- Bowley, G.; Kugler, E.; Wilkinson, R.; Lawrie, A.; van Eeden, F.; Chico, T.J.A.; Evans, P.C.; Noël, E.S.; Serbanovic-Canic, J. Zebrafish as a tractable model of human cardiovascular disease. Br. J. Pharmacol. 2022, 179, 900–917. [Google Scholar] [CrossRef]
- Kolvenbach, C.M.; Dworschak, G.C.; Rieke, J.M.; Woolf, A.S.; Reutter, H.; Odermatt, B.; Hilger, A.C. Modelling human lower urinary tract malformations in zebrafish. Mol. Cell. Pediatr. 2023, 10, 2. [Google Scholar] [CrossRef]
- MacArthur Clark, J.A.; Sun, D. Guidelines for the ethical review of laboratory animal welfare People’s Republic of China National Standard GB/T 35892-2018 [Issued 6 February 2018 Effective from 1 September 2018]. Anim. Models Exp. Med. 2020, 3, 103–113. [Google Scholar] [CrossRef]
- Yang, H.; Liang, X.; Zhao, Y.; Gu, X.; Mao, Z.; Zeng, Q.; Chen, H.; Martyniuk, C.J. Molecular and behavioral responses of zebrafish embryos/larvae after sertraline exposure. Ecotoxicol. Environ. Saf. 2020, 208, 111700. [Google Scholar] [CrossRef]
- Jia, P.-P.; Chandrajith, R.; Junaid, M.; Li, T.-Y.; Li, Y.-Z.; Wei, X.-Y.; Liu, L.; Pei, D.-S. Elucidating environmental factors and their combined effects on CKDu in Sri Lanka using zebrafish. Environ. Pollut. 2023, 332, 121967. [Google Scholar] [CrossRef]
- Katayama, S. Oxidative stress marker. Nihon rinsho. Jpn. J. Clin. Med. 2006, 64 (Suppl. S6), 142–146. [Google Scholar]
- Rudnicka, E.; Duszewska, A.M.; Kucharski, M.; Tyczyński, P.; Smolarczyk, R. Oxidative stress and reproductive function: Oxidative stress in polycystic ovary syndrome. Reproduction 2022, 164, F145. [Google Scholar] [CrossRef]
- Scavuzzi, B.M.; Holoshitz, J. Endoplasmic Reticulum Stress, Oxidative Stress, and Rheumatic Diseases. Antioxidants 2022, 11, 1306. [Google Scholar] [CrossRef]
- Pei, D.-S.; Sun, Y.-H.; Chen, C.-H.; Chen, S.-P.; Wang, Y.-P.; Hu, W.; Zhu, Z.-Y. Identification and characterization of a novel gene differentially expressed in zebrafish cross-subfamily cloned embryos. BMC Dev. Biol. 2008, 8, 29. [Google Scholar] [CrossRef]
- Garbinato, C.; Schneider, S.E.; Sachett, A.; Decui, L.; Conterato, G.M.; Müller, L.G.; Siebel, A.M. Exposure to ractopamine hydrochloride induces changes in heart rate and behavior in zebrafish embryos and larvae. Environ. Sci. Pollut. Res. Int. 2020, 27, 21468–21475. [Google Scholar] [CrossRef]
- Wiprich, M.T.; Zanandrea, R.; Altenhofen, S.; Bonan, C.D. Influence of 3-nitropropionic acid on physiological and behavioral responses in zebrafish larvae and adults. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2020, 234, 108772. [Google Scholar] [CrossRef]
- Patel, M.Y.; Stovall, K.; Franklin, J.L. The intrinsic apoptotic pathway lies upstream of oxidative stress in multiple organs. Free Radic. Biol. Med. 2020, 158, 13–19. [Google Scholar] [CrossRef]
- Carlsson, G.; Blomberg, M.; Pohl, J.; Örn, S. Swimming activity in zebrafish larvae exposed to veterinary antiparasitic pharmaceuticals. Environ. Toxicol. Pharmacol. 2018, 63, 74–77. [Google Scholar] [CrossRef]
- Burnett, J.C., Jr. Atrial Natriuretic Peptide, Heart Failure and the Heart as an Endocrine Organ. Clin. Chem. 2019, 65, 1602–1603. [Google Scholar] [CrossRef]
- Man, J.; Barnett, P.; Christoffels, V.M. Structure and function of the Nppa–Nppb cluster locus during heart development and disease. Cell. Mol. Life Sci. 2018, 75, 1435–1444. [Google Scholar] [CrossRef]
- Park, J.J.; Lee, H.; Kim, K.H.; Yoo, B.; Kang, S.; Baek, S.H.; Jeon, E.; Kim, J.; Cho, M.; Chae, S.C.; et al. Heart failure and atrial fibrillation: Tachycardia-mediated acute decompensation. ESC Hear. Fail. 2021, 8, 2816–2825. [Google Scholar] [CrossRef]
- Hong, T.; Park, H.; An, G.; Park, J.; Song, G.; Lim, W. Fluchloralin induces developmental toxicity in heart, liver, and nervous system during early zebrafish embryogenesis. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2023, 271, 109679. [Google Scholar] [CrossRef]
- Huang, Y.; Chen, Z.; Meng, Y.; Wei, Y.; Xu, Z.; Ma, J.; Zhong, K.; Cao, Z.; Liao, X.; Lu, H. Famoxadone-cymoxanil induced cardiotoxicity in zebrafish embryos. Ecotoxicol. Environ. Saf. 2020, 205, 111339. [Google Scholar] [CrossRef]
- Park, H.; Song, G.; Hong, T.; An, G.; Park, S.; Lim, W. Exposure to the herbicide fluridone induces cardiovascular toxicity in early developmental stages of zebrafish. Sci. Total Environ. 2023, 867, 161535. [Google Scholar] [CrossRef]
- Zhang, Q.; He, X.; Yao, S.; Lin, T.; Zhang, L.; Chen, D.; Chen, C.; Yang, Q.; Li, F.; Zhu, Y.-M.; et al. Ablation of Mto1 in zebrafish exhibited hypertrophic cardiomyopathy manifested by mitochondrion RNA maturation deficiency. Nucleic Acids Res. 2021, 49, 4689–4704. [Google Scholar] [CrossRef]
- Imperatore, R.; Orso, G.; Facchiano, S.; Scarano, P.; Hoseinifar, S.H.; Ashouri, G.; Guarino, C.; Paolucci, M. Anti-inflammatory and immunostimulant effect of different timing-related administration of dietary polyphenols on intestinal inflammation in zebrafish, Danio rerio. Aquaculture 2023, 563, 738878. [Google Scholar] [CrossRef]
- Shi, Y.; Chen, C.; Han, Z.; Chen, K.; Wu, X.; Qiu, X. Combined exposure to microplastics and amitriptyline caused intestinal damage, oxidative stress and gut microbiota dysbiosis in zebrafish (Danio rerio). Aquat. Toxicol. 2023, 260, 106589. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Y.; Liang, H.; Ma, C.; Cui, N.; Cao, H.; Wei, W.; Liu, Y. Single and combined effects of polyethylene microplastics and acetochlor on accumulation and intestinal toxicity of zebrafish (Danio rerio). Environ. Pollut. 2023, 333, 122089. [Google Scholar] [CrossRef]
- McGeachy, M.J.; Cua, D.J.; Gaffen, S.L. The IL-17 Family of Cytokines in Health and Disease. Immunity 2019, 50, 892–906. [Google Scholar] [CrossRef]
- Ruiz de Morales, J.M.G.; Puig, L.; Daudén, E.; Cañete, J.D.; Pablos, J.L.; Martín, A.O.; Juanatey, C.G.; Adán, A.; Montalbán, X.; Borruel, N.; et al. Critical role of interleukin (IL)-17 in inflammatory and immune disorders: An updated review of the evidence focusing in controversies. Autoimmun. Rev. 2020, 19, 102429. [Google Scholar] [CrossRef]
- Varfolomeev, E.; Vucic, D. Intracellular regulation of TNF activity in health and disease. Cytokine 2018, 101, 26–32. [Google Scholar] [CrossRef]
- Yao, Z.; Getting, S.J.; Locke, I.C. Regulation of TNF-Induced Osteoclast Differentiation. Cells 2021, 11, 132. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, G.; Gou, D.; Bu, L.-K.; Wei, X.-Y.; Hu, H.; Huo, W.-B.; Sultan, M.; Pei, D.-S. Developmental Toxicity of PEDOT:PSS in Zebrafish: Effects on Morphology, Cardiac Function, and Intestinal Health. Toxics 2024, 12, 150. https://doi.org/10.3390/toxics12020150
Yang G, Gou D, Bu L-K, Wei X-Y, Hu H, Huo W-B, Sultan M, Pei D-S. Developmental Toxicity of PEDOT:PSS in Zebrafish: Effects on Morphology, Cardiac Function, and Intestinal Health. Toxics. 2024; 12(2):150. https://doi.org/10.3390/toxics12020150
Chicago/Turabian StyleYang, Guan, Dongzhi Gou, Ling-Kang Bu, Xing-Yi Wei, Huan Hu, Wen-Bo Huo, Marriya Sultan, and De-Sheng Pei. 2024. "Developmental Toxicity of PEDOT:PSS in Zebrafish: Effects on Morphology, Cardiac Function, and Intestinal Health" Toxics 12, no. 2: 150. https://doi.org/10.3390/toxics12020150
APA StyleYang, G., Gou, D., Bu, L.-K., Wei, X.-Y., Hu, H., Huo, W.-B., Sultan, M., & Pei, D.-S. (2024). Developmental Toxicity of PEDOT:PSS in Zebrafish: Effects on Morphology, Cardiac Function, and Intestinal Health. Toxics, 12(2), 150. https://doi.org/10.3390/toxics12020150