
Toxic Effects of Arsenic on Four Freshwater Aquatic Species and Its Transformation Metabolism in Crucian Carp (Carassius auratus)

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Acute Toxicity Test
2.3. Arsenic Toxicology Experiment
2.4. Arsenic Speciation Analysis
2.5. Biokinetic Model of Arsenic Bioaccumulation
2.6. Determination of Oxidative Stress Levels
2.7. Statistical Analysis
3. Results and Discussion
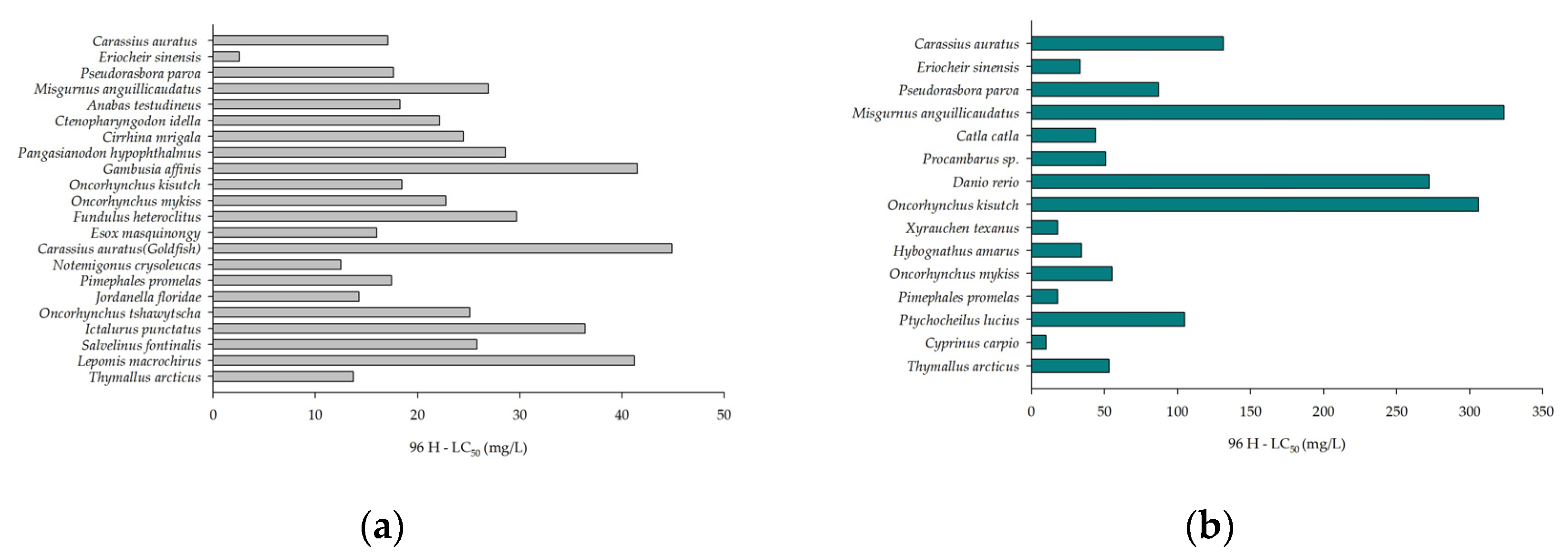
3.1. Acute Toxicity Effects
3.2. Inorganic Arsenic’s Half Lethal Concentration and Safe Concentration for Four Freshwater Aquatic Species
3.3. Biokinetic Parameters of Inorganic Arsenic Bioaccumulation
3.4. Biological Transformation and Metabolism of Arsenic Species in Crucian Carp
3.5. Enzymatic Responses and Lipoperoxidation Level
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Su, Q.; He, Y.; Pan, H.; Liu, H.Y.; Mehmood, K.; Tang, Z.X.; Hu, L.M. Toxicity of inorganic arsenic to animals and its treatment strategies. Comp. Biochem. Physiol. Part C 2023, 271, 109654. [Google Scholar] [CrossRef] [PubMed]
- Bundschuh, J.; Niazi, N.K.; Alam, M.A.; Berg, M.; Herath, I.; Tomaszewska, B.; Maity, J.P.; Ok, Y.S. Global arsenic dilemma and sustainability. J. Hazard. Mater. 2022, 436, 129197. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.Y.; Wang, L.; Li, S.; Cao, J.F.; Yang, Z.G. Species-specific bioaccumulation and correlated health risk of arsenic compounds in freshwater fish from a typical mine-impacted river. Sci. Total Environ. 2018, 625, 600–607. [Google Scholar] [CrossRef]
- Ye, Z.J.; Huang, L.P.; Zhao, Q.Y.; Zhang, W.; Zhang, L. Key genes for arsenobetaine synthesis in marine medaka (Oryzias melastigma) by transcriptomics. Aquat. Toxicol. 2022, 253, 106349. [Google Scholar] [CrossRef]
- Nasrullah; Iqbal, R.K. Phytoremediation of Arsenic in Soil & Water. Res. Rev. J. Bot. Sci. 2019, 8, 8–16. [Google Scholar]
- Fishery and Administration Authority of Agriculture and Rural Affairs of the People’s Republic of China. China Fishery Statistical Yearbook 2022; China Agriculture Press: Beijing, China, 2022. [Google Scholar]
- Wang, N.X.; Ye, Z.J.; Huang, L.P.; Zhang, C.S.; Guo, Y.X.; Zhang, W. Arsenic Occurrence and Cycling in the Aquatic Environment: A Comparison between Freshwater and Seawater. Water 2023, 15, 147. [Google Scholar] [CrossRef]
- Fan, Z.; Wang, Q.; Chen, X.; Qiu, L.P.; Yin, Y.T.; Fan, L.M.; Song, C.; Meng, S.L. Quantitative benefit and risk assessment of arsenic and nutrient levels in cooked and raw Chinese mitten crab (Eriocheir sinensis) using an in vitro digestion model. Food Chem. 2022, 368, 130826. [Google Scholar]
- Byeon, E.; Kang, H.M.; Yoon, C.; Lee, J.S. Toxicity mechanisms of arsenic compounds in aquatic organisms. Aquat. Toxicol. 2021, 237, 105901. [Google Scholar] [CrossRef]
- Cui, D.; Zhang, P.; Li, H.P.; Zhang, Z.X.; Song, Y.; Yang, Z.G. The dynamic changes of arsenic biotransformation and bioaccumulation in muscle of freshwater food fish crucian carp during chronic dietborne exposure. J. Environ. Sci. 2021, 100, 74–81. [Google Scholar] [CrossRef]
- He, J.T.; Ma, S.N.; Liu, L.; Liu, C.; Dong, S.J. A comparison study of test organism species and methodologies for combined toxicity assay of copper ions and zinc ions. Environ. Sci. Pollut. Res. 2020, 36, 45992–46002. [Google Scholar] [CrossRef]
- Bhavani, K.; Karuppasamy, R. Acute toxicity bioassay and behavioural changes on zebra fish, Danio rerio (Hamilton) under arsenic trioxide. Int. J. Mod. Res. Rev. 2014, 2, 40–46. [Google Scholar]
- Dominic, E.P.; Jorge, R.I.; Raphael, A.L.; Gretchen, L.L.; Thomas, A.J.; Jennifer, A.G.; Megan, R.; Caitlyn, D.; Mark, P.; John, M.G.; et al. Mercury, selenium and arsenic concentrations in Canadian freshwater fish and a perspective on human consumption intake and risk. J. Hazard. Mater. 2022, 6, 100060. [Google Scholar]
- Zhang, J.C.; Tan, Q.G.; Huang, L.P.; Ye, Z.J.; Wang, X.H.; Xiao, T.F.; Wu, Y.; Zhang, W.; Yan, B. Intestinal uptake and low transformation increase the bioaccumulation of inorganic arsenic in freshwater zebrafish. J. Hazard. Mater. 2022, 434, 128904. [Google Scholar] [CrossRef]
- Fernanda, G.B.; Sabina, L.V.; Barbara, M.D.C.; Marta, B.; Pamela, T.; Alejandra, V.V.; Gustavo, T. Health risk assessment and differential distribution of Arsenic and metals in organs of Urophycis brasiliensis a commercial fish from Southwestern Atlantic coast. Mar. Pollut. Bull. 2023, 187, 114499. [Google Scholar]
- Tang, S.Z.; Song, D.; Bai, S.Y.; Huang, X.L.; Chen, Z.X.; Wang, P.; Qin, D.L. Trace elements accumulation and health risk assessment of Chinese mitten crab (Eriocheir sinensis) from monoculture and rice-crab co-culture mode. J. Food Compos. Anal. 2023, 123, 105640. [Google Scholar] [CrossRef]
- Nawrocka, A.; Durkalec, M.; Michalski, M.; Posyniak, A. Simple and reliable determination of total arsenic and its species in seafood by ICP-MS and HPLC-ICP-MS. Food Chem. 2022, 379, 132045. [Google Scholar] [CrossRef]
- U.S. EPA (U.S Environmental Protection Agency). Definition and Procedure for the Determination of the Method Detection Limit-Revision 1.11. 2011. Available online: http://www.gpo.gov/fdsys/granule/CFR-2011-title40-vol23/CFR-2011-title40-vol23-part136-appB/content-detail.html (accessed on 6 October 2023).
- Chen, L.Z.; Zhang, W.; Guo, Z.Q.; Zhang, L. Effects of acclimation on arsenic bioaccumulation and biotransformation in freshwater medaka Oryzias mekongensis after chronic arsenic exposure. Environ. Pollut. 2018, 238, 17–25. [Google Scholar] [CrossRef]
- Tang, S.Z.; Chen, Z.X.; Bai, S.Y.; Wang, H.T.; Hao, Q.R.; Wu, S.; Du, N.N.; Mou, Z.B. Acute toxicity of five drugs in aquaculture on rainbow trout Oncorhynchus mykiss Juveniles. Chin. J. Fish 2015, 28, 33–37. [Google Scholar]
- Crespo, E.B.; Pereyra, P.J.; Silvestro, A.; Hidalgo, K.; Rossini, B. Acute toxicity of Cd2+, Cr6+, and Ni2+ to the Golden mussel Limnoperna fortunei (Dunker 1857). Bull. Environ. Contam. Toxicol. 2020, 104, 748–754. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.G.; Wang, W.X. Two-Compartment Toxicokinetic-Toxicodynamic Model to Predict Metal Toxicity in Daphnia magna. Environ. Sci. Technol. 2012, 46, 9709–9715. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Guptab, S.K.; Bhushan, S.; Singh, N.P. Impacts of acute toxicity of arsenic (III) alone and with high temperature on stress biomarkers, immunological status and cellular metabolism in fish. Aquat. Toxicol. 2019, 214, 105233. [Google Scholar] [CrossRef]
- U.S. Environmental Protection Agency. ECOTOX User Guide: ECOTOXicology Knowledgebase System. Version 4.0. 2017. Available online: http://www.epa.gov/ecotox/ (accessed on 6 October 2023).
- Moja, T.N.; Mishra, A.K.; Mishra, S.B. Nano Size Magnetite Particles Layered with the Blend of Conductive Polymer and Superadsorbent Hydrogel: A Core–Shell Based Nanocomposite for Trivalent Arsenide Uptake form Aqueous Solution. J. Inorg. Organomet. Polym. Mater. 2018, 28, 2131–2142. [Google Scholar] [CrossRef]
- Perry, H.; Isphording, W.; Trigg, C.; Riedel, R. Heavy metals in red crabs, Chaceon quinquedens, from the Gulf of Mexico. Mar. Pollut. Bull. 2015, 101, 845–851. [Google Scholar] [CrossRef]
- Liu, X.L.; Wang, Q.; He, J.; Zhao, M.F.; Qiu, L.P.; Fan, L.M.; Meng, S.L.; Yang, G.; Li, T.; Li, Q.; et al. Occurrence, speciation analysis and health risk assessment of arsenic in Chinese mitten crabs (Eriocheir sinensis) collected from China. J. Food Compos. Anal. 2020, 94, 103647. [Google Scholar] [CrossRef]
- Ferreira, N.S.; Oliveira, L.H.B.; Agrelli, V.; Oliveira, A.F.D.; Nogueira, A.R.A.; Oliveira, A.; Gonzalez, M.H. Bioaccumulation and Acute Toxicity of As(III) and As(V) in Nile Tilapia (Oreochromis niloticus). Chemosphere 2019, 217, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Z.; Song, D.D.; Zhang, W.; Zhang, C.C.; Zhang, L. The dynamic changes of arsenic bioaccumulation and antioxidant responses in the marine medaka Oryzias melastigma during chronic exposure. Aquat. Toxicol. 2019, 212, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Chen, L.Z.; Zhou, Y.Y.; Wu, Y.; Zhang, L. Biotransformation of inorganic arsenic in a marine herbivorous fish Siganus fuscescens after dietborne exposure. Chemosphere 2016, 147, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Kumari, B.; Kumar, V.; Sinha, A.K.; Ahsan, J.; Ghosh, A.K.; Wang, H.P.; DeBoeck, G. Toxicology of arsenic in fish and aquatic systems. Environ Chem Lett 2017, 15, 43–64. [Google Scholar] [CrossRef]
- Suhendrayatna, A.O.; Nakajima, T.; Maeda, S. Studies on the accumulation and transformation of arsenic in freshwater organisms I. Accumulation, transformation and toxicity of arsenic compounds on the Japanese Medaka, Oryzias latipes. Chemosphere 2002, 46, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Shinichi, M.; Masahito, S.; Yoshiaki, K.; Takayoshi, K.; Osamu, S.; Shoko, F.; Kevin, A.F.; Toshikazu, K. Rapid determination of arsenic species in freshwater organisms from the arsenic-rich Hayakawa River in Japan using HPLC-ICP-MS. Chemosphere 2009, 75, 1065–1073. [Google Scholar]
- María, J.C.; Alessandra, C.; Gabriela, C.; Vicenta, D.; Dinoraz, V. Dietary compounds to reduce in vivo inorganic arsenic bioavailability. J. Agric. Food Chem. 2019, 67, 9032–9038. [Google Scholar]
- Ventura-Lima, J.; Castro, M.; Acosta, D.; Fattorini, D.; Regoli, F.; Carvalho, L.M.D.; Bohrer, D.; Geracitano, L.A.; Barros, D.M.; Marins, L.F.F.; et al. Effects of arsenic (As) exposure on the antioxidant status of gills of the zebrafish Danio rerio (Cyprinidae). Comp. Biochem. Phys. C 2009, 149, 538–543. [Google Scholar] [CrossRef] [PubMed]
- Pei, J.; Zuo, J.X.; Wang, X.Y.; Yin, J.Y.; Liu, L.P.; Fan, W.H. The Bioaccumulation and Tissue Distribution of Arsenic Species in Tilapia. Int. J. Environ. Res. Public Health 2019, 16, 757. [Google Scholar] [CrossRef] [PubMed]
- Christian, S.; Annette, F.; Christoph, S. Determination of lipid content in fish samples from bioaccumulation studies: Contributions to the revision of guideline OECD 305. Environ. Sci. Eur. 2012, 24, 13. [Google Scholar]
- Song, D.D.; Zhu, S.Q.; Chen, L.Z.; Zhang, T.; Zhang, L. The strategy of arsenic metabolism in an arsenic-resistant bacterium Stenotrophomonas maltophilia SCSIOOM isolated from fish gut. Environ. Pollut. 2022, 312, 120085. [Google Scholar] [CrossRef] [PubMed]
- Juncos, R.; Arcagni, M.; Squadrone, S.; Rizzo, A.; Arribére, M.; Barriga, J.P.; Battini, M.A.; Campbell, L.M.; Brizio, P.; Abete, M.C.; et al. Interspecific differences in the bioaccumulation of arsenic of three Patagonian top predator fish: Organ distribution and arsenic speciation. Ecotoxicol. Environ. Saf. 2019, 168, 431–442. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.Y.; Liu, L.P.; Liang, D.Y.; Liu, Y.Y.; Zhao, Q.; Huang, P.; Li, X.M.; Fan, W.H. Accumulation, transformation and subcellular distribution of arsenite associated with five carbon nanomaterials in freshwater zebrafish specific-tissues. J. Hazard. Mater. 2021, 415, 125579. [Google Scholar] [CrossRef]
- Lu, D.L.; Luo, W.B.; Li, H.P.; Yang, Z.G. Biotransformation and detoxification mechanism of inorganic arsenic in a freshwater benthic fish Tachysurus fulvidraco with dietborne exposure. Ecotoxicology 2023, 32, 46–56. [Google Scholar] [CrossRef]
- Zhang, W.; Guo, Z.Q.; Song, D.D.; Du, S.; Zhang, L. Arsenic speciation in wild marine organisms and a health risk assessment in a subtropical bay of China. Sci. Total Environ. 2018, 626, 621–629. [Google Scholar] [CrossRef]
- Molin, M.; Ulven, S.M.; Meltzer, H.M.; Alexander, J. Arsenic in the human food chain, biotransformation and toxicology—Review focusing on seafood arsenic. J. Trace Elem. Med. Biol. 2015, 31, 249–259. [Google Scholar] [CrossRef]
- Perumal, E.; Eswaran, S.; Parvin, R.; Balasubramanian, S. Mitigation of arsenic induced developmental cardiotoxicity by ferulic acid in zebrafish. Comp. Biochem. Phys. C 2021, 244, 109021–109030. [Google Scholar] [CrossRef] [PubMed]
- Greani, S.; Lourkisti, R.; Berti, L.; Marchand, B.; Giannettini, J.; Santini, J.; Quilichini, Y. Effect of chronic arsenic exposure under environmental conditions on bioaccumulation, oxidative stress, and antioxidant enzymatic defenses in wild trout Salmo trutta (pisces, teleostei). Ecotoxicology 2017, 26, 930–941. [Google Scholar] [CrossRef] [PubMed]
- Altikat, S.; Uysal, K.; Kuru, H.I.; Kavasoglu, M.; Kucuk, A. The effect of arsenic on some antioxidant enzyme activities and lipid peroxidation in various tissues of mirror carp (Cyprinus carpio carpio). Environ. Sci. Pollut. Res. 2014, 22, 3212–3218. [Google Scholar] [CrossRef] [PubMed]
- Cui, D.; Zhang, P.; Li, H.; Zhang, Z.; Yang, Z. Biotransformation of dietary inorganic arsenic in a freshwater fish Carassius auratus and the unique association between arsenic dimethylation and oxidative damage. J. Hazard. Mater. 2020, 391, 122153. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Scientific Name | Time | As(III) | As(V) | ||
|---|---|---|---|---|---|
| Regression Equation | R2 | Regression Equation | R2 | ||
| M. anguillicaudatus | 24 h | y = −4.75 + 2.20x | 0.924 | y = −8.76 + 3.03x | 0.989 |
| 48 h | y = −4.25 + 2.57x | 0.984 | y = −9.27 + 3.43x | 0.953 | |
| 96 h | y = −5.55 + 3.95x | 0.997 | y = −10.61 + 4.25x | 0.987 | |
| P. parva | 24 h | y = −3.59 + 1.97x | 0.931 | y = −5.64 + 2.22x | 0.956 |
| 48 h | y = −3.29 + 2.40x | 0.912 | y = −5.2 + 2.38x | 0.981 | |
| 96 h | y = −3.23 + 2.63x | 0.979 | y = −4.26 + 2.26x | 0.980 | |
| E. sinensis | 24 h | y = −1.63 + 1.04x | 0.930 | y = −4.55 + 2.06x | 0.948 |
| 48 h | y = −1.32 + 1.76x | 0.969 | y = −3.61 + 2.06x | 0.979 | |
| 96 h | y = −0.64 + 1.74x | 0.964 | y = −3.26 + 2.26x | 0.967 | |
| C. auratus | 24 h | y = −2.53 + 1.39x | 0.973 | y = −5.6 + 2.63x | 0.952 |
| 48 h | y = −2.93 + 2.00x | 0.995 | y = −6.34 + 2.83x | 0.993 | |
| 96 h | y = −4.17 + 3.39x | 0.969 | y = −5.79 + 2.30x | 0.919 | |
| Inorganic Arsenic | Species Scientific Name | 24 h | 48 h | 96 h | SC | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| LC50 | Lower Limit | Upper Limit | LC50 | Lower Limit | Upper Limit | LC50 | Lower Limit | Upper Limit | |||
| As(III) | M. anguillicaudatus | 112.49 | 95.55 | 132.44 | 46.14 | 38.38 | 55.49 | 26.92 | 23.40 | 30.97 | 2.33 |
| P. parva | 50.71 | 42.48 | 60.53 | 25.59 | 21.08 | 31.08 | 17.62 | 14.92 | 20.81 | 1.96 | |
| E. sinensis | 21.13 | 15.13 | 29.53 | 5.58 | 4.09 | 7.62 | 2.55 | 1.89 | 3.45 | 0.12 | |
| C. auratus | 48.75 | 37.63 | 63.17 | 29.31 | 22.78 | 37.71 | 17.10 | 14.10 | 20.75 | 3.18 | |
| As(V) | M. anguillicaudatus | 690.24 | 644.56 | 739.16 | 484.17 | 449.35 | 521.70 | 323.59 | 302.73 | 345.90 | 71.47 |
| P. parva | 294.42 | 267.27 | 324.33 | 151.00 | 135.94 | 167.72 | 86.89 | 78.87 | 95.73 | 11.91 | |
| E. sinensis | 126.19 | 114.42 | 139.18 | 57.68 | 51.75 | 64.29 | 33.19 | 30.09 | 36.61 | 3.62 | |
| C. auratus | 281.17 | 254.56 | 310.57 | 169.42 | 153.22 | 187.33 | 131.51 | 119.37 | 144.89 | 18.45 | |
| Tissue | Cw (μg/L) | k1 | k2 | BCF | CAmax (μg/kg) | B1/2 (d) | R2 |
|---|---|---|---|---|---|---|---|
| Muscle | 1000 | 0.015335 | 0.03727 | 0.41145 | 411.45 | 18.59404 | 0.88712 |
| Gills | 1000 | 0.412295 | 0.726 | 0.5679 | 567.9 | 0.954545 | 0.45647 |
| Intestine | 1000 | 0.159382 | 0.1855 | 0.8592 | 859.2 | 3.735849 | 0.87028 |
| Skin | 1000 | 0.059684 | 0.0262 | 2.278 | 2278 | 26.45038 | 0.88427 |
| Liver | 1000 | 0.232551 | 0 | - | - | - | 0.89525 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, S.; Gao, L.; Qin, D.; Wang, H.; Huang, L.; Wu, S.; Bai, S.; Du, N.; Sun, Y.; Wang, P.; et al. Toxic Effects of Arsenic on Four Freshwater Aquatic Species and Its Transformation Metabolism in Crucian Carp (Carassius auratus). Toxics 2024, 12, 221. https://doi.org/10.3390/toxics12030221
Tang S, Gao L, Qin D, Wang H, Huang L, Wu S, Bai S, Du N, Sun Y, Wang P, et al. Toxic Effects of Arsenic on Four Freshwater Aquatic Species and Its Transformation Metabolism in Crucian Carp (Carassius auratus). Toxics. 2024; 12(3):221. https://doi.org/10.3390/toxics12030221
Chicago/Turabian StyleTang, Shizhan, Lei Gao, Dongli Qin, Haitao Wang, Li Huang, Song Wu, Shuyan Bai, Ningning Du, Yanchun Sun, Peng Wang, and et al. 2024. "Toxic Effects of Arsenic on Four Freshwater Aquatic Species and Its Transformation Metabolism in Crucian Carp (Carassius auratus)" Toxics 12, no. 3: 221. https://doi.org/10.3390/toxics12030221
APA StyleTang, S., Gao, L., Qin, D., Wang, H., Huang, L., Wu, S., Bai, S., Du, N., Sun, Y., Wang, P., & Chen, Z. (2024). Toxic Effects of Arsenic on Four Freshwater Aquatic Species and Its Transformation Metabolism in Crucian Carp (Carassius auratus). Toxics, 12(3), 221. https://doi.org/10.3390/toxics12030221