
Concordance between In Vitro and In Vivo Relative Toxic Potencies of Diesel Exhaust Particles from Different Biodiesel Blends

 , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Engine
2.2. Fuels
2.3. Engine Operation for Generation of Test Particles
2.4. Emission Characterization
2.5. Extraction of Particles
2.6. Preparation of Particle Suspensions
2.7. In Vitro Exposure
2.7.1. Cell Culture
2.7.2. DEP Exposure
2.7.3. Cytotoxicity Analyses
2.7.4. Cytokine Secretion
2.8. In Vivo Exposure
2.8.1. Animals
2.8.2. Intratracheal Instillation
2.8.3. Biological Samples
2.8.4. Cytology
2.8.5. Cytokines
2.8.6. Gene Expression Analyses
2.8.7. Potency Calculations
2.9. Statistical Analyses
3. Results
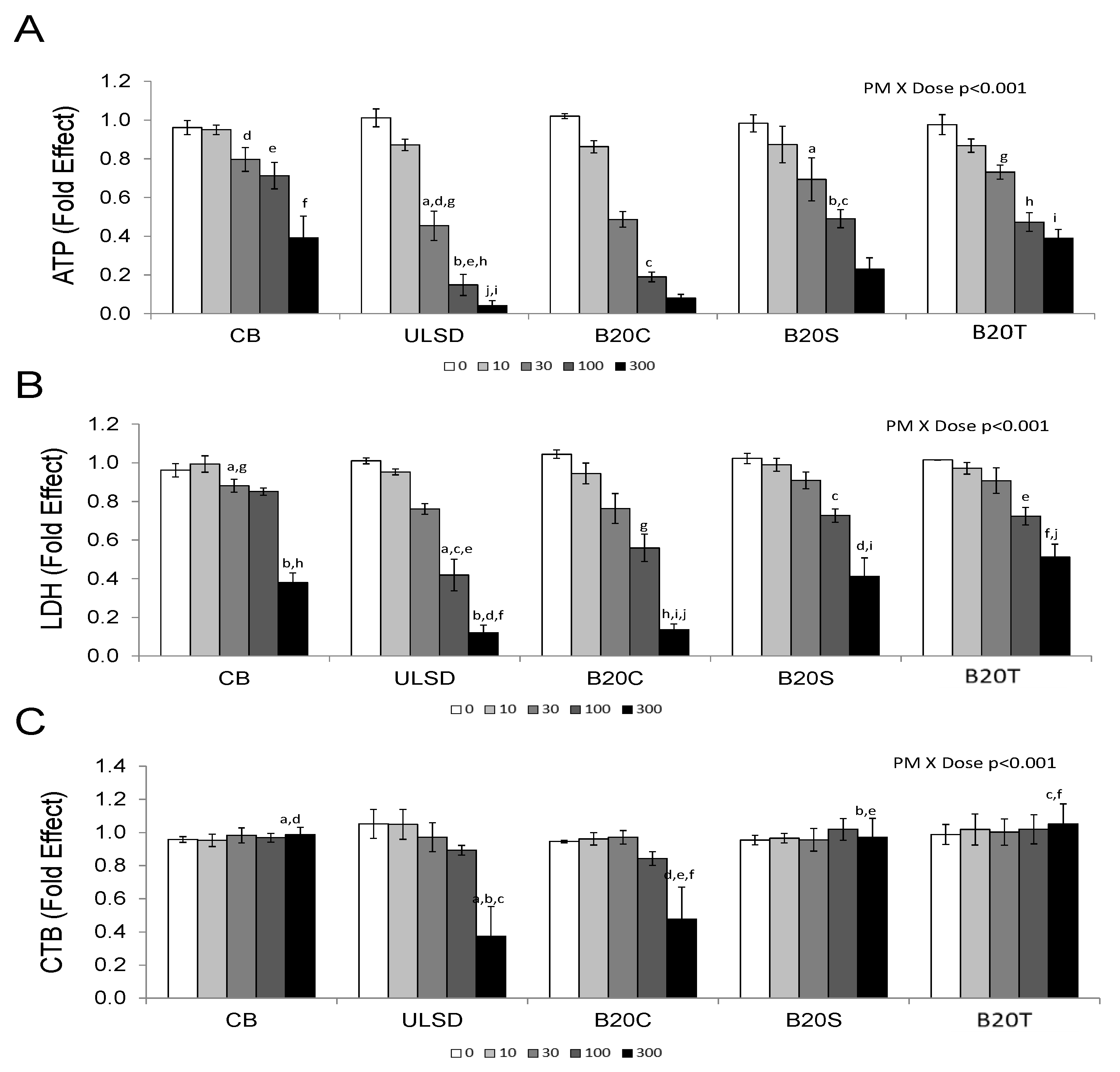
3.1. In Vitro Effects
3.1.1. Cytotoxicity
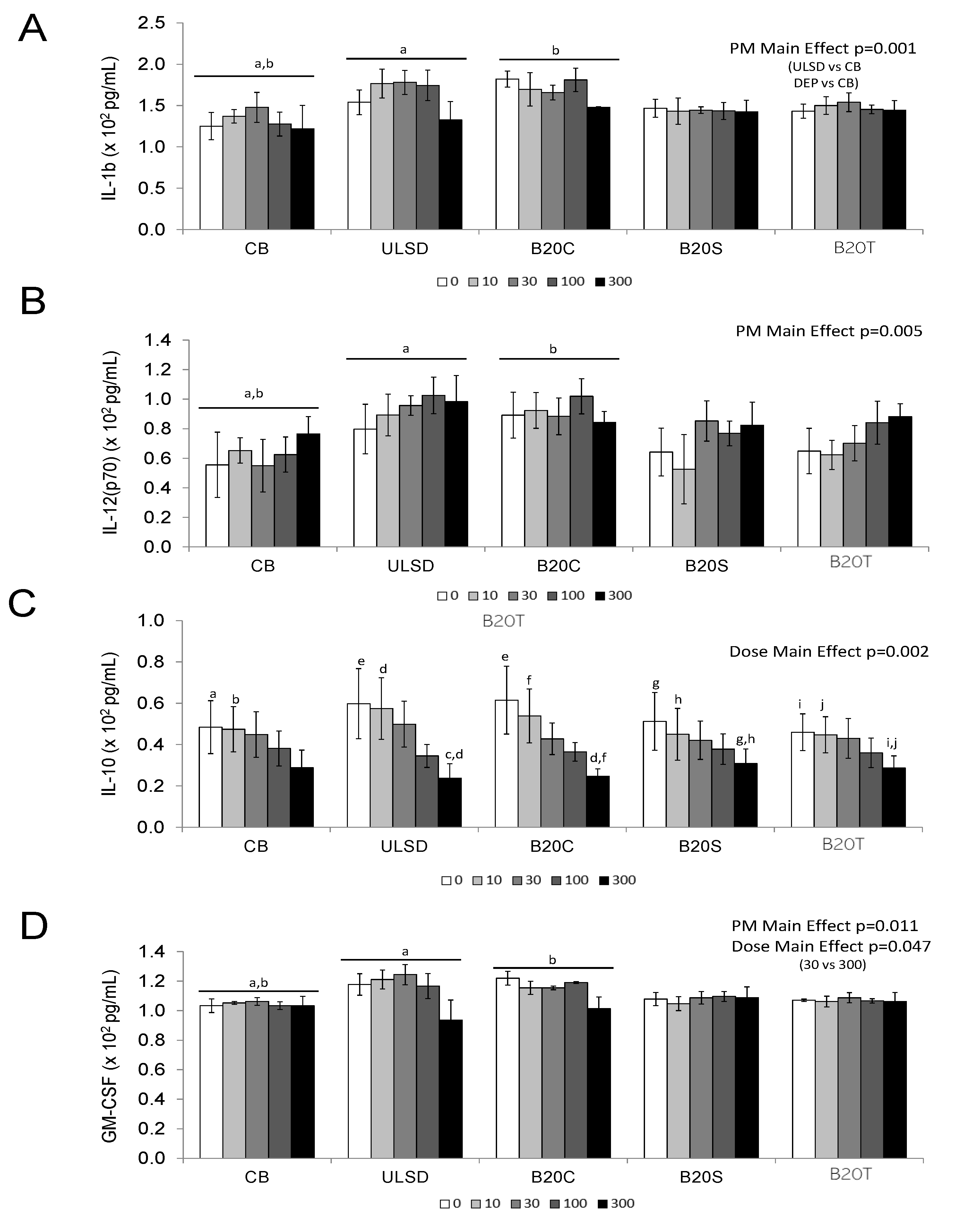
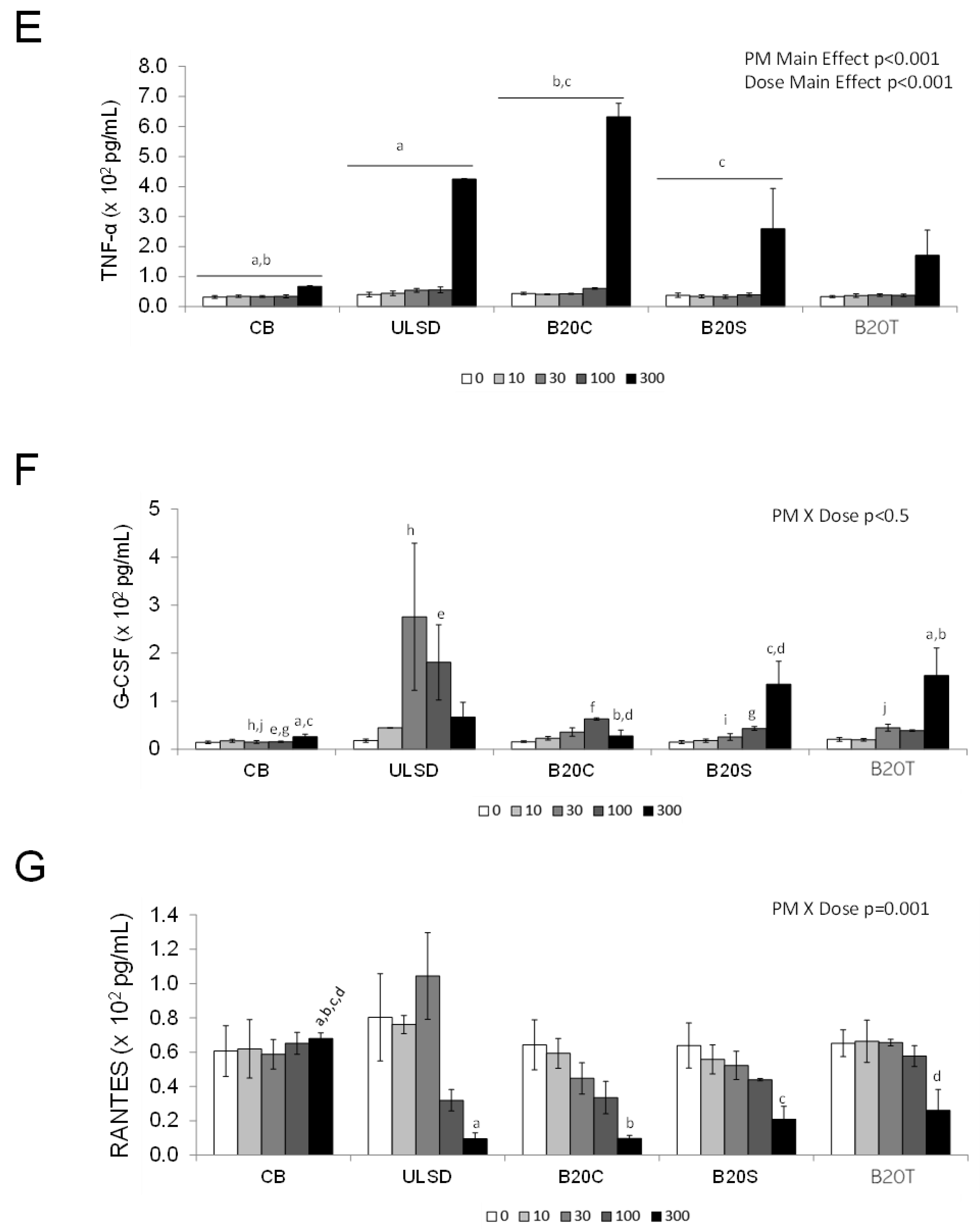
3.1.2. In Vitro Cytokine Secretion
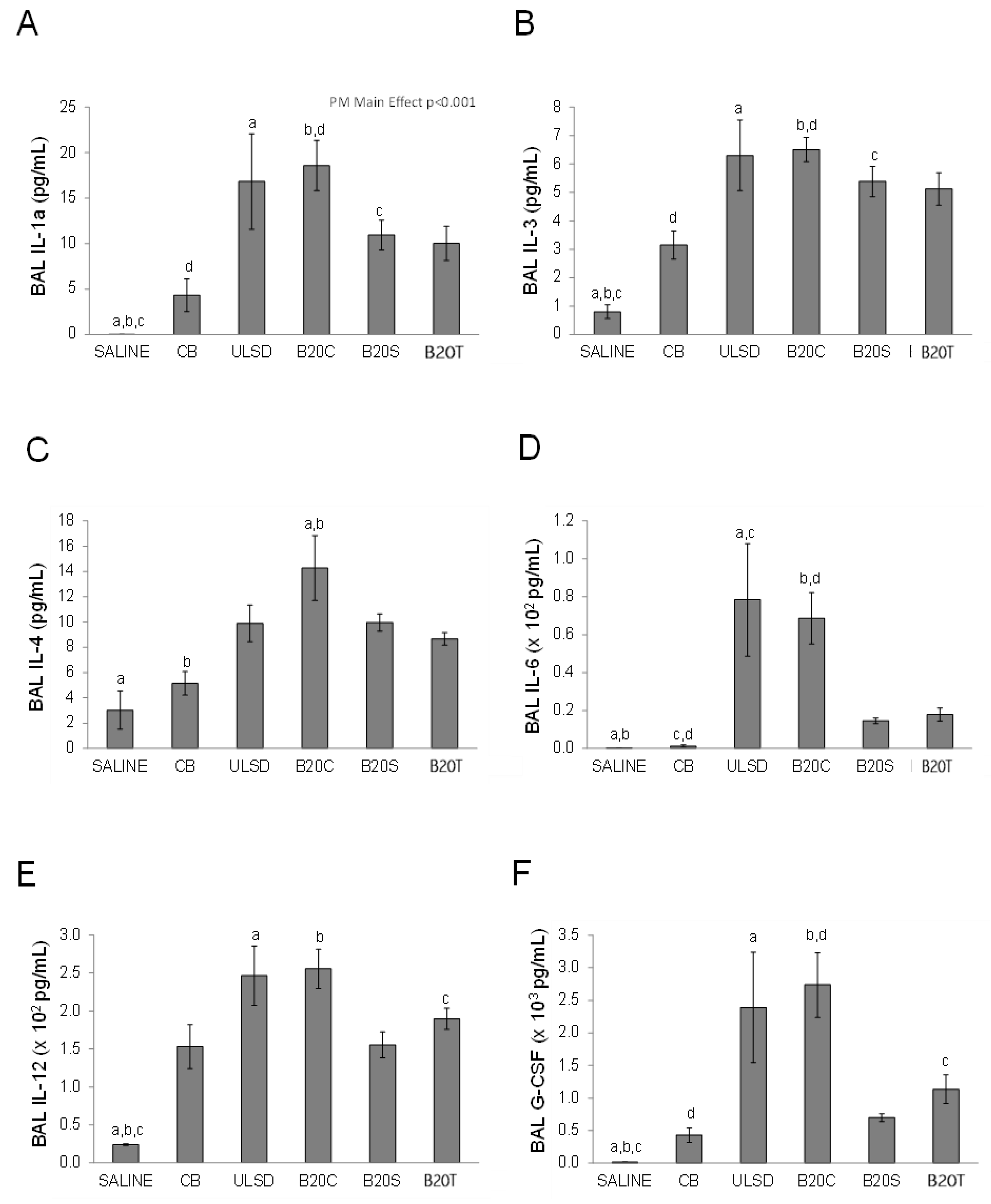
3.2. In Vivo Effects
3.2.1. BAL Neutrophil Counts
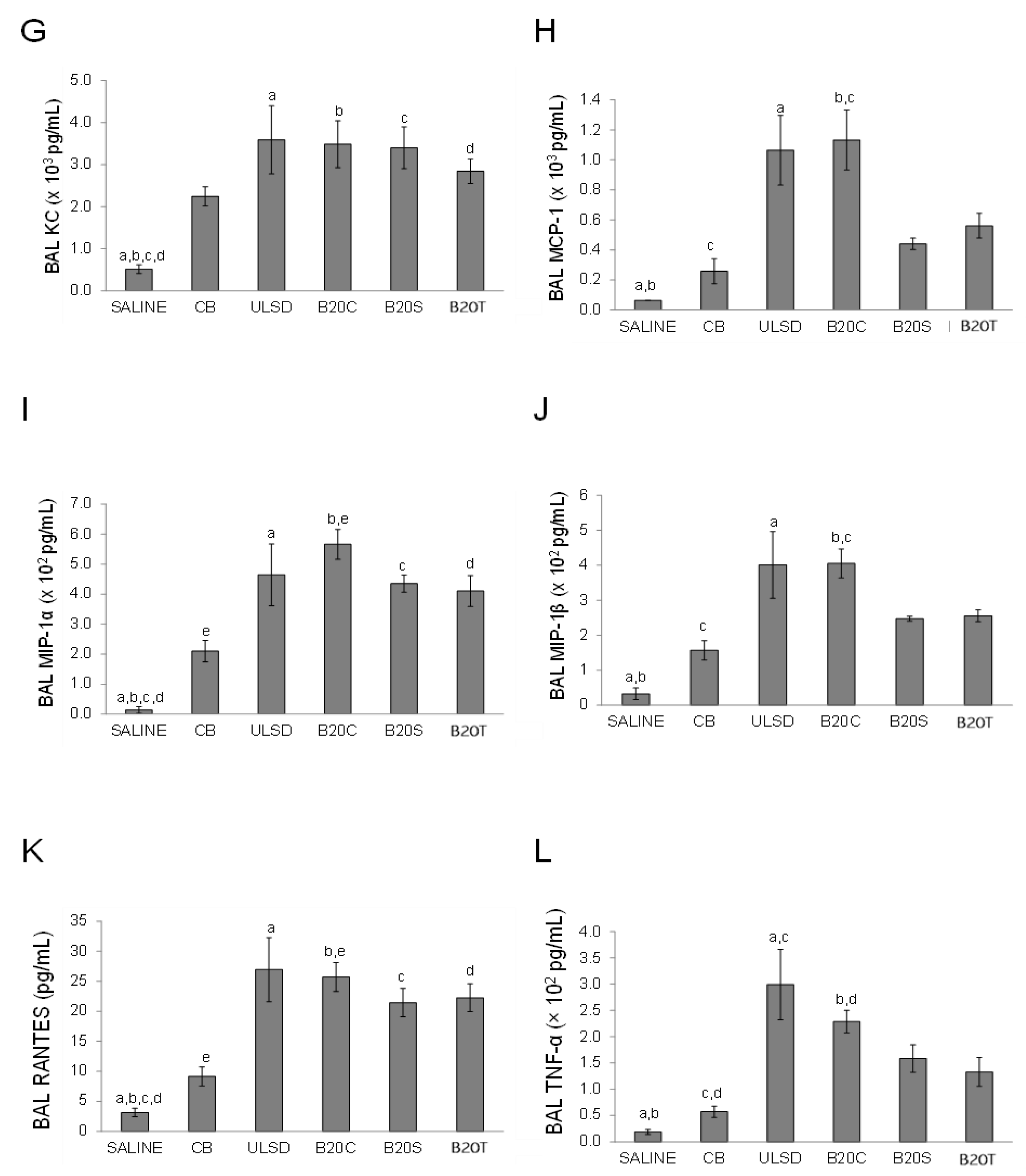
3.2.2. BAL Cytokines
3.2.3. Lung Gene Expression
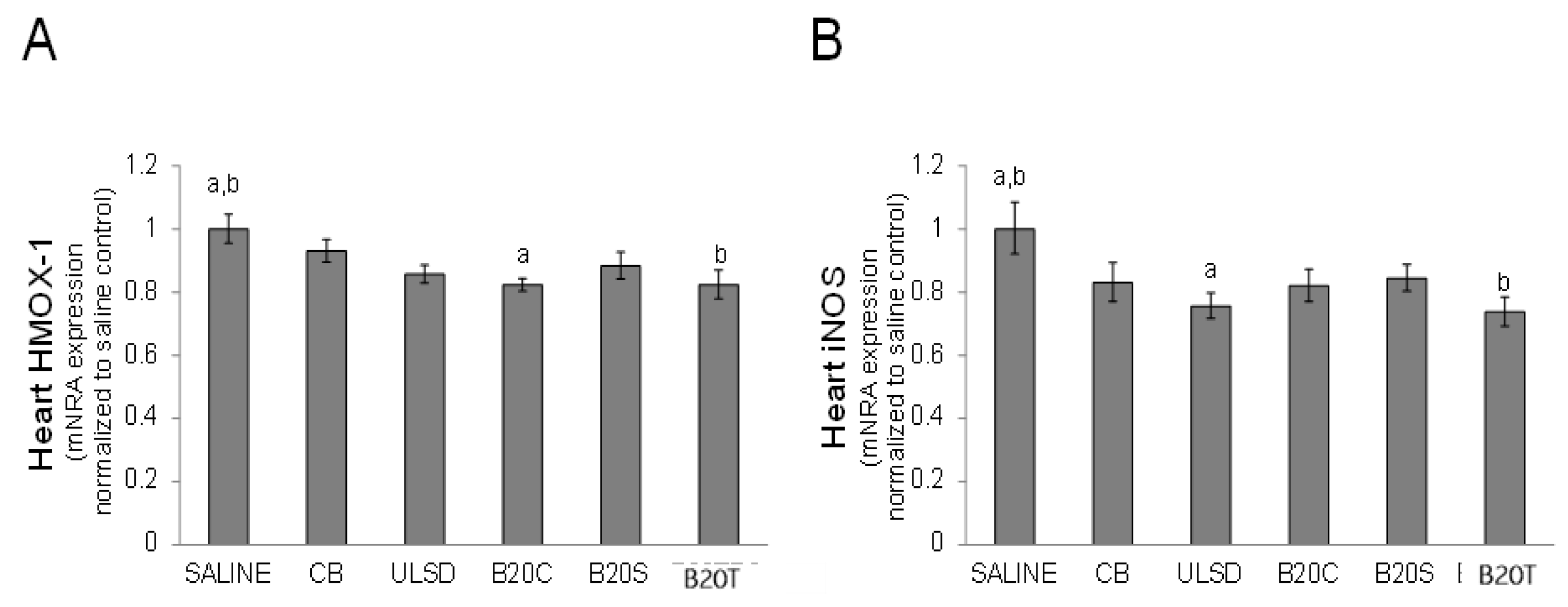
3.2.4. Heart Gene Expression
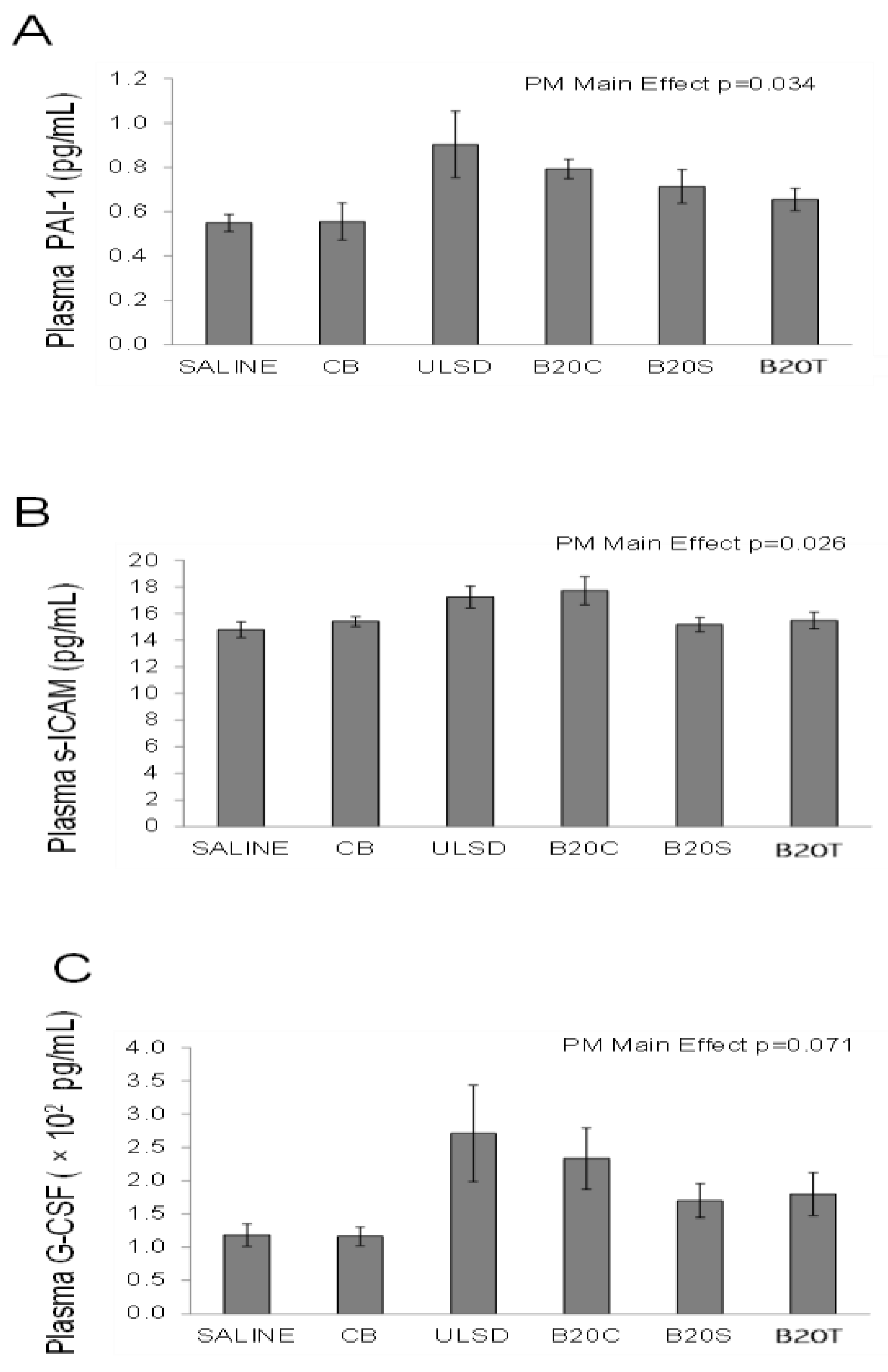
3.2.5. Plasma Cytokines
3.3. Correlations between In Vivo vs. In Vitro Toxicity Endpoints
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hoffmann, B.; Moebus, S.; Möhlenkamp, S.; Stang, A.; Lehmann, N.; Dragano, N.; Schmermund, A.; Memmesheimer, M.; Mann, K.; Erbel, R.; et al. Residential exposure to traffic is associated with coronary atherosclerosis. Circulation 2007, 116, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Tonne, C.; Melly, S.; Mittleman, M.; Coull, B.; Goldberg, R.; Schwartz, J. A case-control analysis of exposure to traffic and acute myocardial infarction. Environ. Health Perspect. 2007, 115, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Auchincloss, A.H.; Diez Roux, A.V.; Dvonch, J.T.; Brown, P.L.; Barr, R.G.; Daviglus, M.L.; Goff, D.C.; Kaufman, J.D.; O’Neill, M.S. Associations between recent exposure to ambient fine particulate matter and blood pressure in the Multi-ethnic Study of Atherosclerosis (MESA). Environ. Health Perspect. 2008, 116, 486–491. [Google Scholar] [CrossRef] [PubMed]
- Lall, R.; Ito, K.; Thurston, G.D. Distributed lag analyses of daily hospital admissions and source-apportioned fine particle air pollution. Environ. Health Perspect. 2011, 119, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Raaschou-Nielsen, O.; Andersen, Z.J.; Jensen, S.S.; Ketzel, M.; Sørensen, M.; Hansen, J.; Loft, S.; Tjønneland, A.; Overvad, K. Traffic air pollution and mortality from cardiovascular disease and all causes: A Danish cohort study. Environ. Health 2012, 11, 60. [Google Scholar] [CrossRef]
- Kirwa, K.; Eliot, M.N.; Wang, Y.; Adams, M.A.; Morgan, C.G.; Kerr, J.; Norman, G.J.; Eaton, C.B.; Allison, M.A.; Wellenius, G.A. Residential proximity to major roadways and prevalent hypertension among postmenopausal women: Results from the Women’s Health Initiative San Diego Cohort. J. Am. Heart Assoc. 2014, 3, e000727. [Google Scholar] [CrossRef]
- Wichmann, J.; Sjöberg, K.; Tang, L.; Haeger-Eugensson, M.; Rosengren, A.; Andersson, E.M.; Barregard, L.; Sallsten, G. The effect of secondary inorganic aerosols, soot and the geographical origin of air mass on acute myocardial infarction hospitalisations in Gothenburg, Sweden during 1985–2010: A case-crossover study. Environ. Health 2014, 13, 61. [Google Scholar] [CrossRef] [PubMed]
- Vincent, R.; Kumarathasan, P.; Goegan, P.; Bjarnason, S.G.; Guénette, J.; Karthikeyan, S.; Thomson, E.M.; Adamson, I.Y.; Watkinson, W.P.; Battistini, B.; et al. Acute cardiovascular effects of inhaled ambient particulate matter: Chemical composition-related oxidative stress, endothelin-1, blood pressure, and ST-segment changes in Wistar rats. Chemosphere 2022, 296, 133933. [Google Scholar] [CrossRef]
- Hazenkamp-von Arx, M.E.; Schindler, C.; Ragettli, M.S.; Künzli, N.; Braun-Fahrländer, C.; Liu, L.-J.S. Impacts of highway traffic exhaust in alpine valleys on the respiratory health in adults: A cross-sectional study. Environ. Health 2011, 10, 13. [Google Scholar] [CrossRef]
- Delfino, R.J.; Wu, J.; Tjoa, T.; Gullesserian, S.K.; Nickerson, B.; Gillen, D.L. Asthma morbidity and ambient air pollution: Effect modification by residential traffic-related air pollution. Epidemiology 2014, 25, 48–57. [Google Scholar] [CrossRef]
- Wilhelm, M.; Ritz, B. Residential proximity to traffic and adverse birth outcomes in Los Angeles county, California, 1994–1996. Environ. Health Perspect. 2003, 111, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Yorifuji, T.; Naruse, H.; Kashima, S.; Ohki, S.; Murakoshi, T.; Takao, S.; Tsuda, T.; Doi, H. Residential proximity to major roads and preterm births. Epidemiology 2011, 22, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Mitku, A.A.; Zewotir, T.; North, D.; Jeena, P.; Asharam, K.; Muttoo, S.; Tularam, H.; Naidoo, R.N. Impact of ambient air pollution exposure during pregnancy on adverse birth outcomes: Generalized structural equation modeling approach. BMC Public Health 2023, 23, 45. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Garcidueñas, L.; Ayala, A. Air Pollution, Ultrafine Particles, and Your Brain: Are Combustion Nanoparticle Emissions and Engineered Nanoparticles Causing Preventable Fatal Neurodegenerative Diseases and Common Neuropsychiatric Outcomes? Environ. Sci. Technol. 2022, 56, 6847–6856. [Google Scholar] [CrossRef] [PubMed]
- Raaschou-Nielsen, O.; Andersen, Z.J.; Hvidberg, M.; Jensen, S.S.; Ketzel, M.; Sørensen, M.; Loft, S.; Overvad, K.; Tjønneland, A. Lung cancer incidence and long-term exposure to air pollution from traffic. Environ. Health Perspect. 2011, 119, 860–865. [Google Scholar] [CrossRef] [PubMed]
- Hung, L.-J.; Chan, T.-F.; Wu, C.-H.; Chiu, H.-F.; Yang, C.-Y. Traffic air pollution and risk of death from ovarian cancer in Taiwan: Fine particulate matter (PM2.5) as a proxy marker. J. Toxicol. Environ. Health A 2012, 75, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Dummer, T.J.B.; Yu, X.; Cui, Y.; Nauta, L.; Saint-Jacques, N.; Sweeney Magee, M.; Rainham, D.G.C. Traffic-Related Air Pollution and Risk of Lung, Breast, and Urinary Tract Cancer in Halifax, Nova Scotia. J. Occup. Environ. Med. 2023, 65, e485–e490. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Robles, L.A.; Fu, J.S.; Reed, G.D. Modeling and source apportionment of diesel particulate matter. Environ. Int. 2008, 34, 1–11. [Google Scholar] [CrossRef][Green Version]
- Brandt, E.B.; Kovacic, M.B.; Lee, G.B.; Gibson, A.M.; Acciani, T.H.; Le Cras, T.D.; Ryan, P.H.; Budelsky, A.L.; Khurana Hershey, G.K. Diesel exhaust particle induction of IL-17A contributes to severe asthma. J. Allergy Clin. Immunol. 2013, 132, 1194–1204. [Google Scholar] [CrossRef]
- Gordon, C.J.; Schladweiler, M.C.; Krantz, T.; King, C.; Kodavanti, U.P. Cardiovascular and thermoregulatory responses of unrestrained rats exposed to filtered or unfiltered diesel exhaust. Inhal. Toxicol. 2012, 24, 296–309. [Google Scholar] [CrossRef]
- Labranche, N.; El Khattabi, C.; Dewachter, L.; Dreyfuss, C.; Fontaine, J.; van de Borne, P.; Berkenboom, G.; Pochet, S. Vascular oxidative stress induced by diesel exhaust microparticles: Synergism with hypertension. J. Cardiovasc. Pharmacol. 2012, 60, 530–537. [Google Scholar] [CrossRef]
- Karthikeyan, S.; Thomson, E.M.; Kumarathasan, P.; Guénette, J.; Rosenblatt, D.; Chan, T.; Rideout, G.; Vincent, R. Nitrogen dioxide and ultrafine particles dominate the biological effects of inhaled diesel exhaust treated by a catalyzed diesel particulate filter. Toxicol. Sci. 2013, 135, 437–450. [Google Scholar] [CrossRef]
- Pöss, J.; Lorenz, D.; Werner, C.; Pavlikova, V.; Gensch, C.; Speer, T.; Alessandrini, F.; Berezowski, V.; Kuntz, M.; Mempel, M.; et al. Diesel exhaust particles impair endothelial progenitor cells, compromise endothelial integrity, reduce neoangiogenesis, and increase atherogenesis in mice. Cardiovasc. Toxicol. 2013, 13, 290–300. [Google Scholar] [CrossRef]
- Kim, J.Y.; Kim, J.-H.; Kim, Y.-D.; Seo, J.H. Ultrafine Diesel Exhaust Particles Induce Apoptosis of Oligodendrocytes by Increasing Intracellular Reactive Oxygen Species through NADPH Oxidase Activation. Antioxidants 2022, 11, 1031. [Google Scholar] [CrossRef]
- McCormick, R.L. The impact of biodiesel on pollutant emissions and public health. Inhal. Toxicol. 2007, 19, 1033–1039. [Google Scholar] [CrossRef]
- Fontaras, G.; Karavalakis, G.; Kousoulidou, M.; Ntziachristos, L.; Bakeas, E.; Stournas, S.; Samaras, Z. Effects of low concentration biodiesel blends application on modern passenger cars. Part 2: Impact on carbonyl compound emissions. Environ. Pollut. 2010, 158, 2496–2503. [Google Scholar] [CrossRef]
- Cahill, T.M.; Okamoto, R.A. Emissions of acrolein and other aldehydes from biodiesel-fueled heavy-duty vehicles. Environ. Sci. Technol. 2012, 46, 8382–8388. [Google Scholar] [CrossRef]
- George, I.J.; Hays, M.D.; Snow, R.; Faircloth, J.; George, B.J.; Long, T.; Baldauf, R.W. Cold temperature and biodiesel fuel effects on speciated emissions of volatile organic compounds from diesel trucks. Environ. Sci. Technol. 2014, 48, 14782–14789. [Google Scholar] [CrossRef]
- Fontaras, G.; Kousoulidou, M.; Karavalakis, G.; Tzamkiozis, T.; Pistikopoulos, P.; Ntziachristos, L.; Bakeas, E.; Stournas, S.; Samaras, Z. Effects of low concentration biodiesel blend application on modern passenger cars. Part 1: Feedstock impact on regulated pollutants, fuel consumption and particle emissions. Environ. Pollut. 2010, 158, 1451–1460. [Google Scholar] [CrossRef]
- Surawski, N.C.; Miljevic, B.; Ayoko, G.A.; Elbagir, S.; Stevanovic, S.; Fairfull-Smith, K.E.; Bottle, S.E.; Ristovski, Z.D. Physicochemical characterization of particulate emissions from a compression ignition engine: The influence of biodiesel feedstock. Environ. Sci. Technol. 2011, 45, 10337–10343. [Google Scholar] [CrossRef]
- Hajbabaei, M.; Johnson, K.C.; Okamoto, R.A.; Mitchell, A.; Pullman, M.; Durbin, T.D. Evaluation of the impacts of biodiesel and second generation biofuels on NO(x) emissions for CARB diesel fuels. Environ. Sci. Technol. 2012, 46, 9163–9173. [Google Scholar] [CrossRef]
- Karavalakis, G.; Gysel, N.; Schmitz, D.A.; Cho, A.K.; Sioutas, C.; Schauer, J.J.; Cocker, D.R.; Durbin, T.D. Impact of biodiesel on regulated and unregulated emissions, and redox and proinflammatory properties of PM emitted from heavy-duty vehicles. Sci. Total Environ. 2017, 584–585, 1230–1238. [Google Scholar] [CrossRef]
- Jalava, P.I.; Tapanainen, M.; Kuuspalo, K.; Markkanen, A.; Hakulinen, P.; Happo, M.S.; Pennanen, A.S.; Ihalainen, M.; Yli-Pirilä, P.; Makkonen, U.; et al. Toxicological effects of emission particles from fossil- and biodiesel-fueled diesel engine with and without DOC/POC catalytic converter. Inhal. Toxicol. 2010, 22 (Suppl. 2), 48–58. [Google Scholar] [CrossRef]
- Hawley, B.; L’Orange, C.; Olsen, D.B.; Marchese, A.J.; Volckens, J. Oxidative stress and aromatic hydrocarbon response of human bronchial epithelial cells exposed to petro- or biodiesel exhaust treated with a diesel particulate filter. Toxicol. Sci. 2014, 141, 505–514. [Google Scholar] [CrossRef]
- Rouleau, M.; Egyed, M.; Taylor, B.; Chen, J.; Samaali, M.; Davignon, D.; Morneau, G. Human health impacts of biodiesel use in on-road heavy duty diesel vehicles in Canada. Environ. Sci. Technol. 2013, 47, 13113–13121. [Google Scholar] [CrossRef]
- Gerlofs-Nijland, M.E.; Totlandsdal, A.I.; Tzamkiozis, T.; Leseman, D.L.A.C.; Samaras, Z.; Låg, M.; Schwarze, P.; Ntziachristos, L.; Cassee, F.R. Cell toxicity and oxidative potential of engine exhaust particles: Impact of using particulate filter or biodiesel fuel blend. Environ. Sci. Technol. 2013, 47, 5931–5938. [Google Scholar] [CrossRef]
- Mullins, B.J.; Kicic, A.; Ling, K.-M.; Mead-Hunter, R.; Larcombe, A.N. Biodiesel exhaust-induced cytotoxicity and proinflammatory mediator production in human airway epithelial cells. Environ. Toxicol. 2016, 31, 44–57. [Google Scholar] [CrossRef]
- Agarwal, A.K.; Singh, A.P.; Gupta, T.; Agarwal, R.A.; Sharma, N.; Rajput, P.; Pandey, S.K.; Ateeq, B. Mutagenicity and Cytotoxicity of Particulate Matter Emitted from Biodiesel-Fueled Engines. Environ. Sci. Technol. 2018, 52, 14496–14507. [Google Scholar] [CrossRef]
- Yanamala, N.; Hatfield, M.K.; Farcas, M.T.; Schwegler-Berry, D.; Hummer, J.A.; Shurin, M.R.; Birch, M.E.; Gutkin, D.W.; Kisin, E.; Kagan, V.E.; et al. Biodiesel versus diesel exposure: Enhanced pulmonary inflammation, oxidative stress, and differential morphological changes in the mouse lung. Toxicol. Appl. Pharmacol. 2013, 272, 373–383. [Google Scholar] [CrossRef]
- Kisin, E.R.; Yanamala, N.; Farcas, M.T.; Gutkin, D.W.; Shurin, M.R.; Kagan, V.E.; Bugarski, A.D.; Shvedova, A.A. Abnormalities in the male reproductive system after exposure to diesel and biodiesel blend. Environ. Mol. Mutagen. 2015, 56, 265–276. [Google Scholar] [CrossRef]
- Fukagawa, N.K.; Li, M.; Poynter, M.E.; Palmer, B.C.; Parker, E.; Kasumba, J.; Holmén, B.A. Soy biodiesel and petrodiesel emissions differ in size, chemical composition and stimulation of inflammatory responses in cells and animals. Environ. Sci. Technol. 2013, 47, 12496–12504. [Google Scholar] [CrossRef]
- Douki, T.; Corbière, C.; Preterre, D.; Martin, P.J.; Lecureur, V.; André, V.; Landkocz, Y.; Pottier, I.; Keravec, V.; Fardel, O.; et al. Comparative study of diesel and biodiesel exhausts on lung oxidative stress and genotoxicity in rats. Environ. Pollut. 2018, 235, 514–524. [Google Scholar] [CrossRef]
- Gavett, S.H.; Wood, C.E.; Williams, M.A.; Cyphert, J.M.; Boykin, E.H.; Daniels, M.J.; Copeland, L.B.; King, C.; Krantz, T.Q.; Richards, J.H.; et al. Soy biodiesel emissions have reduced inflammatory effects compared to diesel emissions in healthy and allergic mice. Inhal. Toxicol. 2015, 27, 533–544. [Google Scholar] [CrossRef] [PubMed]
- Madden, M.C. A paler shade of green? The toxicology of biodiesel emissions: Recent findings from studies with this alternative fuel. Biochim. Biophys. Acta 2016, 1860, 2856–2862. [Google Scholar] [CrossRef]
- Nadeau, D.; Vincent, R.; Kumarathasan, P.; Brook, J.; Dufresne, A. Cytotoxicity of ambient air particles to rat lung macrophages: Comparison of cellular and functional assays. Toxicol. In Vitro 1996, 10, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Kumarathasan, P.; Breznan, D.; Das, D.; Salam, M.A.; Siddiqui, Y.; MacKinnon-Roy, C.; Guan, J.; de Silva, N.; Simard, B.; Vincent, R. Cytotoxicity of carbon nanotube variants: A comparative in vitro exposure study with A549 epithelial and J774 macrophage cells. Nanotoxicology 2015, 9, 148–161. [Google Scholar] [CrossRef]
- Vincent, R.; Vu, D.; Hatch, G.; Poon, R.; Dreher, K.; Guénette, J.; Bjarnason, S.; Potvin, M.; Norwood, J.; McMullen, E. Sensitivity of lungs of aging Fischer 344 rats to ozone: Assessment by bronchoalveolar lavage. Am. J. Physiol. 1996, 271, L555–L565. [Google Scholar] [CrossRef]
- Poon, R.; Nakai, J.; Yagminas, A.; Benoit, F.; Moir, D.; Chu, I.; Valli, V.E. Subchronic toxicity of chloral hydrate on rats: A drinking water study. J. Appl. Toxicol. 2002, 22, 227–236. [Google Scholar] [CrossRef]
- Thomson, E.M.; Vladisavljevic, D.; Mohottalage, S.; Kumarathasan, P.; Vincent, R. Mapping acute systemic effects of inhaled particulate matter and ozone: Multiorgan gene expression and glucocorticoid activity. Toxicol. Sci. 2013, 135, 169–181. [Google Scholar] [CrossRef]
- Vincent, R.; Goegan, P.; Johnson, G.; Brook, J.R.; Kumarathasan, P.; Bouthillier, L.; Burnett, R.T. Regulation of promoter-CAT stress genes in HepG2 cells by suspensions of particles from ambient air. Fundam. Appl. Toxicol. 1997, 39, 18–32. [Google Scholar] [CrossRef]
- Vincent, R.; Kumarathasan, P.; Goegan, P.; Bjarnason, S.G.; Guénette, J.; Bérubé, D.; Adamson, I.Y.; Desjardins, S.; Burnett, R.T.; Miller, F.J.; et al. Inhalation Toxicology of Urban Ambient Particulate Matter: Acute Cardiovascular Effects in Rats. Res. Rep. Health Eff Inst. 2001, 104, 5–54; discussion 55–62. [Google Scholar]
- DeMarini, D.M.; Mutlu, E.; Warren, S.H.; King, C.; Gilmour, M.I.; Linak, W.P. Mutagenicity emission factors of canola oil and waste vegetable oil biodiesel: Comparison to soy biodiesel. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2019, 846, 403057. [Google Scholar] [CrossRef]
- Liu, Y.-Y.; Lin, T.-C.; Wang, Y.-J.; Ho, W.-L. Biological toxicities of emissions from an unmodified engine fueled with diesel and biodiesel blend. J. Environ. Sci. Health Part A Tox. Hazard. Subst. Environ. Eng. 2008, 43, 1735–1743. [Google Scholar] [CrossRef] [PubMed]
- Bünger, J.; Krahl, J.; Baum, K.; Schröder, O.; Müller, M.; Westphal, G.; Ruhnau, P.; Schulz, T.G.; Hallier, E. Cytotoxic and mutagenic effects, particle size and concentration analysis of diesel engine emissions using biodiesel and petrol diesel as fuel. Arch. Toxicol. 2000, 74, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.K.; Gupta, T.; Dixit, N.; Shukla, P.C. Assessment of toxic potential of primary and secondary particulates/aerosols from biodiesel vis-à-vis mineral diesel fuelled engine. Inhal. Toxicol. 2013, 25, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Schwarze, P.E.; Totlandsdal, A.I.; Låg, M.; Refsnes, M.; Holme, J.A.; Øvrevik, J. Inflammation-related effects of diesel engine exhaust particles: Studies on lung cells in vitro. BioMed Res. Int. 2013, 2013, 685142. [Google Scholar] [CrossRef] [PubMed]
- Inoue, K.; Takano, H. Metallothionein as a negative regulator of pulmonary inflammation. Curr. Pharm. Biotechnol. 2013, 14, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Wright, K.; Morgan, E.T. Transcriptional and post-transcriptional suppression of P450IIC11 and P450IIC12 by inflammation. FEBS Lett. 1990, 271, 59–61. [Google Scholar] [CrossRef][Green Version]
- Witkowska, A.M.; Borawska, M.H. Soluble intercellular adhesion molecule-1 (sICAM-1): An overview. Eur. Cytokine Netw. 2004, 15, 91–98. [Google Scholar]
- Ghosh, A.K.; Vaughan, D.E. PAI-1 in tissue fibrosis. J. Cell Physiol. 2012, 227, 493–507. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Engine Manufacturer | Caterpillar |
|---|---|
| Model | C11 |
| Year | 2004 |
| Serial number | KCA018109 |
| Engine family | 4CPXH0680EBK |
| Air handling system | Series Turbo-Charged |
| Control | Electronic ACERT |
| Bore (mm) | 130 |
| Stroke (mm) | 140 |
| Cycles | 4 |
| Number of cylinders | 6 |
| Displacement (liters) | 11.1 |
| Curb idle speed (rpm) | 700 |
| Rated test speed (rpm) | 2100 |
| Maximum torque (lb-ft) | 1150 @ 1200 rpm |
| Maximum power (bhp) | 305 @ 2100 rpm |
| Compression ratio | 17.1 |
| NOx (g/bhp-hr.) * | 2.3 |
| CO (g/bhp-hr.) * | 1.6 |
| PM (g/bhp-hr.) * | 0.09 |
| Fuel Identification | Method | ULSD | Canola B20 | Soy B20 | Animal Tallow B20 |
|---|---|---|---|---|---|
| Biodiesel blend Volume % * | ASTM D7371 | n/a | 19.6 | 19.3 | 20.2 |
| Density, kg/m3 @ 15 °C | ASTM D4052 | 833.0 | 843.1 | 843.3 | 841.6 |
| Cetane No. D613 | ASTM D613 | 49.6 | 52.8 | 55.6 | 53.8 |
| Carbon, %m | ASTM D5291 | 86.20 | 84.32 | 84.17 | 84.10 |
| Hydrogen, %m | ASTM D5291 | 12.79 | 13.02 | 13.38 | 13.66 |
| Sulphur, mg/kg | ASTM D5453 | 4.6 | 3.8 | 4.6 | 7.6 |
| Fuel | Engine Load | CO (g/bhp-hr.) | CO2 (g/bhp-hr.) | NOX (g/bhp-hr.) | THC (g/bhp-hr.) | PM (g/bhp-hr.) | FC (g/bhp-hr.) | Torque (lb.-ft) | Power (bhp) |
|---|---|---|---|---|---|---|---|---|---|
| ULSD | 25% | 0.07 (0.01) | 559.8 (5.10) | 1.73 (0.03) | 0.023 (0.00) | 0.107 (0.01) | 177.3 (1.61) | 561 (4.58) | 129 (1.03) |
| 50% | 0.04 (0.01) | 492.6 (2.22) | 1.63 (0.02) | 0.004 (0.00) | 0.059 (0.00) | 156.0 (0.71) | 844 (3.94) | 194 (0.91) | |
| B20 canola | 25% | 0.08 (0.02) | 553.7 (0.96) | 1.65 (0.02) | 0.036 (0.00) | 0.115 (0.00) | 179.4 (0.31) | 529 (1.29) | 121 (0.31) |
| 50% | 0.05 (0.01) | 497.9 (6.21) | 1.72 (0.02) | 0.000 (0.00) | 0.059 (0.00) | 161.3 (2.02) | 810 (8.63) | 186 (1.98) | |
| B20 Soy | 25% | 0.08 (0.02) | 555.9 (1.23) | 1.57 (0.01) | 0.029 (0.00) | 0.112 (0.00) | 179.7 (0.40) | 545 (0.60) | 125 (0.13) |
| 50% | 0.05 (0.01) | 499.7 (2.07) | 1.62 (0.01) | 0.001 (0.00) | 0.057 (0.00) | 161.5 (0.67) | 802 (2.98) | 184 (0.64) | |
| B20 Tallow | 25% | 0.06 (0.01) | 566.2 (1.45) | 1.78 (0.03) | 0.028 (0.00) | 0.115 (0.00) | 183.0 (0.46) | 534 (1.89) | 122 (0.48) |
| 50% | 0.04 (0.01) | 504.0 (1.06) | 1.71 (0.00) | 0.007 (0.00) | 0.058 (0.00) | 162.9 (0.35) | 799 (0.59) | 183 (0.20) |
| In Vivo Endpoints | In Vitro Cytotoxicity | In Vitro TNF-α Secretion | |||
|---|---|---|---|---|---|
| Compartment | Endpoint | ATP | CTB | LDH | |
| BAL cells | PMN Cell Number | 0.792 (0.110) | 0.688 (0.199) | 0.739 (0.153) | 0.774 (0.124) |
| BAL cytokines | G-CSF | 0.945 (0.015) | 0.919 (0.027) | 0.923 (0.025) | 0.909 (0.032) |
| IL-1a | 0.964 (0.008) | 0.850 (0.068) | 0.871 (0.054) | 0.993 (0.001) | |
| IL-3 | 0.901 (0.037) | 0.708 (0.181) | 0.756 (0.139) | 0.974 (0.005) | |
| IL-4 | 0.781 (0.119) | 0.656 (0.230) | 0.634 (0.251) | 0.947 (0.014) | |
| IL-6 | 0.986 (0.002) | 0.953 (0.012) | 0.980 (0.003) | 0.889 (0.044) | |
| IL-10 | 0.842 (0.074) | 0.920 (0.027) | 0.888 (0.044) | 0.884 (0.046) | |
| IL-12(p40) | 0.921 (0.026) | 0.901 (0.037) | 0.913 (0.030) | 0.926 (0.024) | |
| MCP-1 | 0.967 (0.007) | 0.915 (0.029) | 0.931 (0.021) | 0.931 (0.021) | |
| MIP-1α | 0.821 (0.088) | 0.631 (0.254) | 0.652 (0.233) | 0.967 (0.007) | |
| MIP-1β | 0.983 (0.003) | 0.892 (0.042) | 0.921 (0.026) | 0.961 (0.009) | |
| KC | 0.867 (0.057) | 0.668 (0.217) | 0.722 (0.169) | 0.920 (0.027) | |
| TNF-a | 0.972 (0.006) | 0.850 (0.068) | 0.915 (0.029) | 0.890 (0.043) | |
| Lung gene expression | CYP1A1 | −0.867 (0.057) | −0.966 (0.008) | −0.952 (0.012) | −0.682 (0.204) |
| MTII | 0.913 (0.031) | 0.748 (0.146) | 0.840 (0.075) | 0.838 (0.076) | |
| IL-1β | 0.949 (0.014) | 0.793 (0.109) | 0.869 (0.056) | 0.897 (0.039) | |
| IL-6 | 0.944 (0.016) | 0.795 (0.108) | 0.873 (0.053) | 0.879 (0.050) | |
| Heart gene expression | IL-1β | 0.906 (0.034) | 0.922 (0.026) | 0.880 (0.049) | 0.913 (0.030) |
| Plasma cytokines | MMP-9 | 0.817 (0.091) | 0.913 (0.030) | 0.925 (0.025) | 0.576 (0.310) |
| G-CSF | 0.966 (0.008) | 0.843 (0.073) | 0.909 (0.033) | 0.895 (0.040) | |
| PAI-1 | 0.962 (0.009) | 0.836 (0.078) | 0.904 (0.035) | 0.879 (0.050) | |
| s-ICAM | 0.911 (0.031) | 0.965 (0.008) | 0.943 (0.016) | 0.831 (0.081) | |
| s-VCAM | 0.899 (0.038) | 0.895 (0.040) | 0.863 (0.059) | 0.923 (0.025) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Karthikeyan, S.; Breznan, D.; Thomson, E.M.; Blais, E.; Vincent, R.; Kumarathasan, P. Concordance between In Vitro and In Vivo Relative Toxic Potencies of Diesel Exhaust Particles from Different Biodiesel Blends. Toxics 2024, 12, 290. https://doi.org/10.3390/toxics12040290
Karthikeyan S, Breznan D, Thomson EM, Blais E, Vincent R, Kumarathasan P. Concordance between In Vitro and In Vivo Relative Toxic Potencies of Diesel Exhaust Particles from Different Biodiesel Blends. Toxics. 2024; 12(4):290. https://doi.org/10.3390/toxics12040290
Chicago/Turabian StyleKarthikeyan, Subramanian, Dalibor Breznan, Errol M. Thomson, Erica Blais, Renaud Vincent, and Premkumari Kumarathasan. 2024. "Concordance between In Vitro and In Vivo Relative Toxic Potencies of Diesel Exhaust Particles from Different Biodiesel Blends" Toxics 12, no. 4: 290. https://doi.org/10.3390/toxics12040290
APA StyleKarthikeyan, S., Breznan, D., Thomson, E. M., Blais, E., Vincent, R., & Kumarathasan, P. (2024). Concordance between In Vitro and In Vivo Relative Toxic Potencies of Diesel Exhaust Particles from Different Biodiesel Blends. Toxics, 12(4), 290. https://doi.org/10.3390/toxics12040290