
SOS! Hydrogen Sulfide Enhances the Flavonoid Early Warning System in Rice Plants to Cope with Thiocyanate Pollution


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods and Materials
2.1. Plant Growth and Experimental Design
2.2. Measurement of Growth Parameters
2.3. Quantification of Flavonoids in the Rice Plants
2.4. Quantification of ROS Levels in Rice Plants
2.5. Modeling of Positive/Negative Feedback Networks of Flavonoids in Rice Plants
2.5.1. Evaluating Genetic Expression Variation Factors
2.5.2. Evaluation of Flavonoid Variation Factors
2.6. Data Analysis
3. Results
3.1. The Relative Growth Rate of Rice Plants
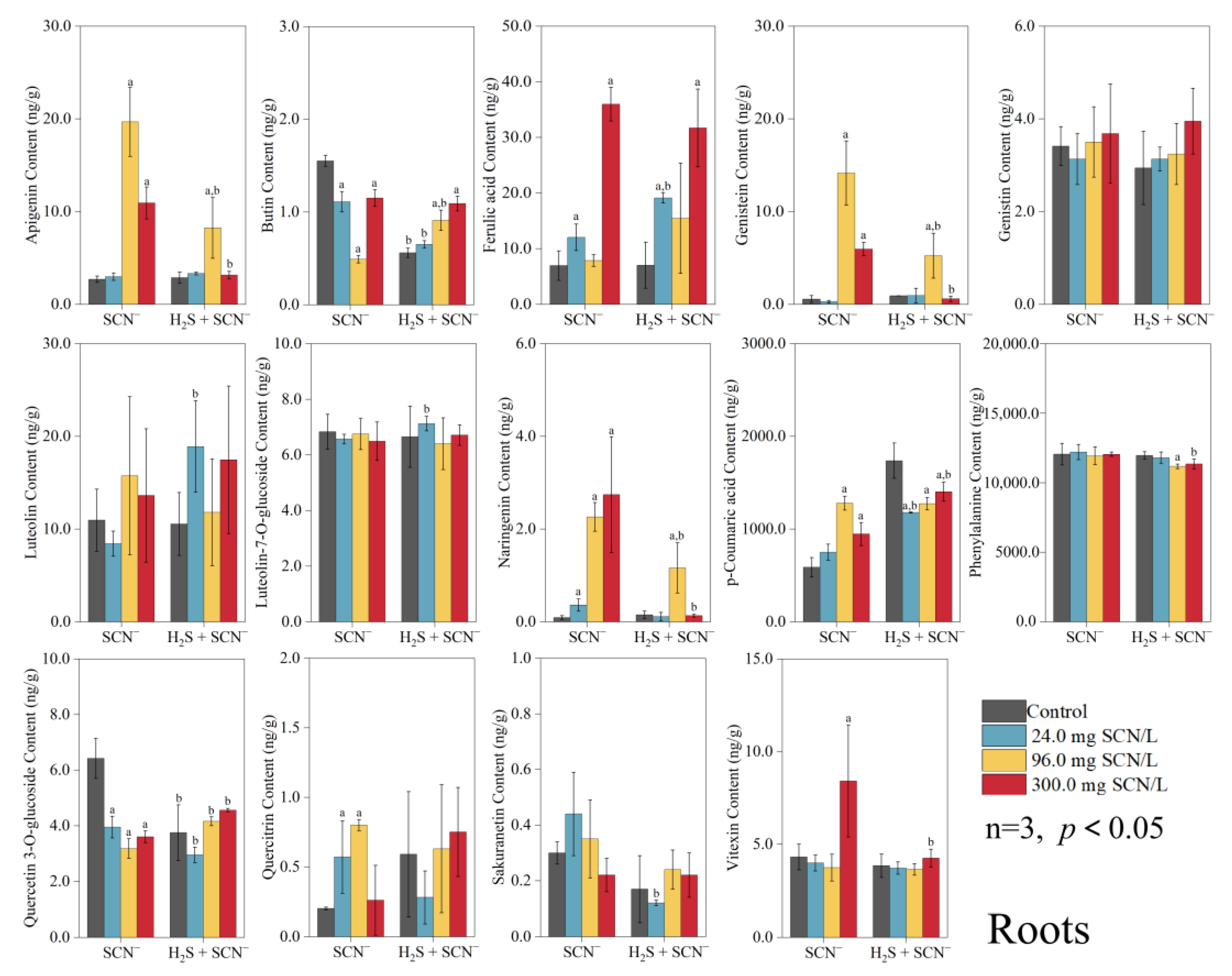
3.2. Response of Rice Plants to Flavonoids
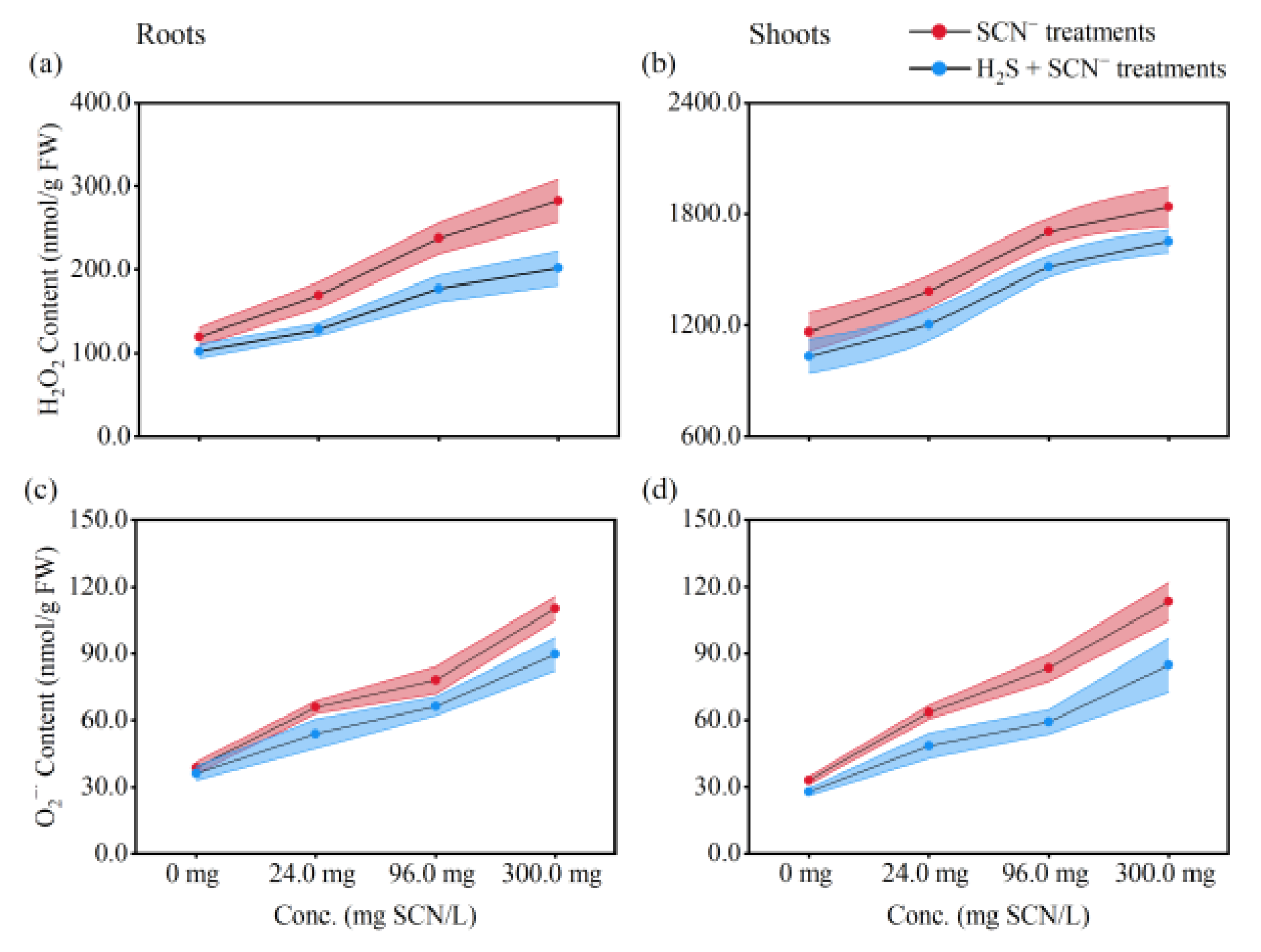
3.3. ROS Levels in Rice Plants
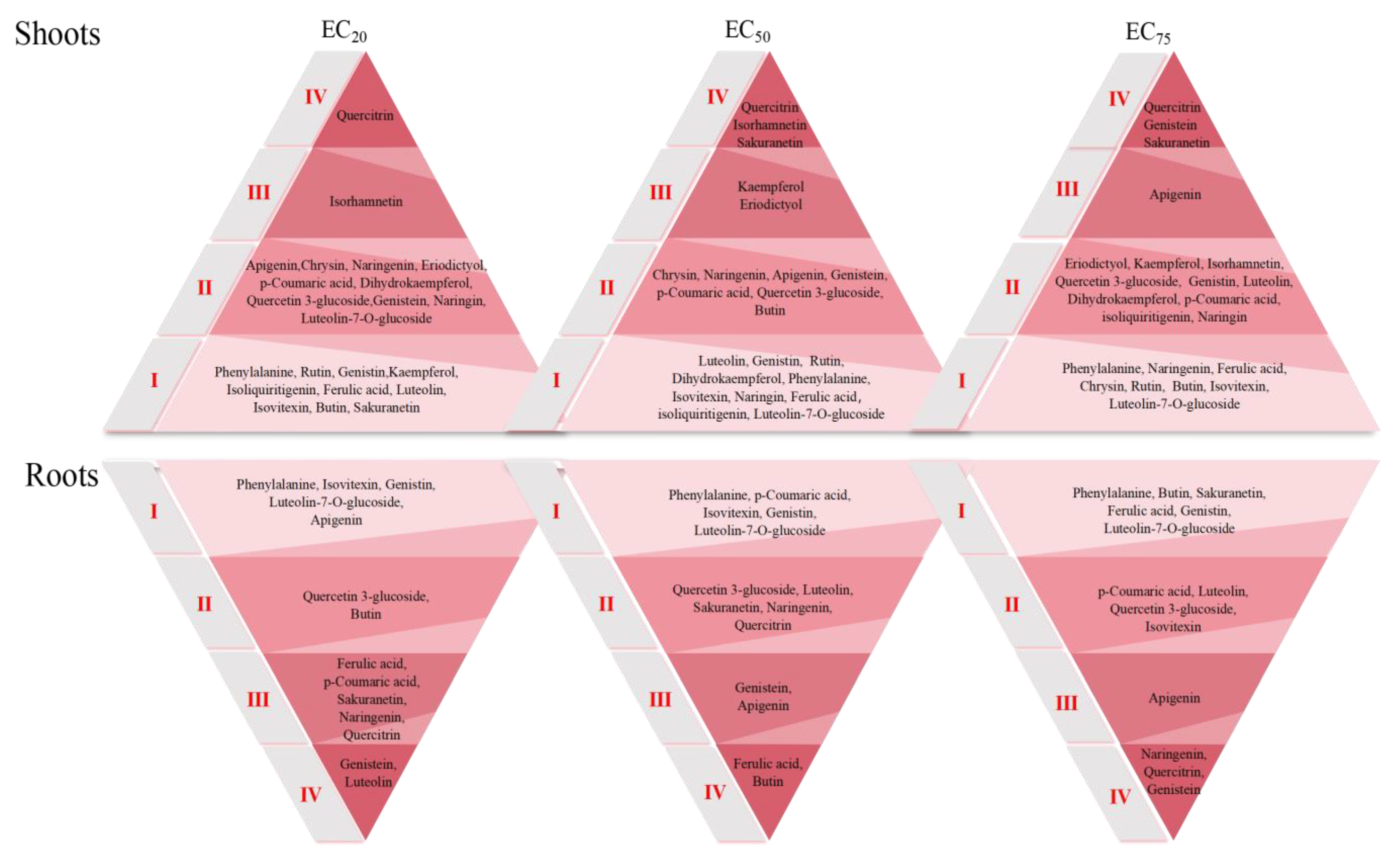
3.4. Evaluation of Flavonoid Early Warning Response Levels
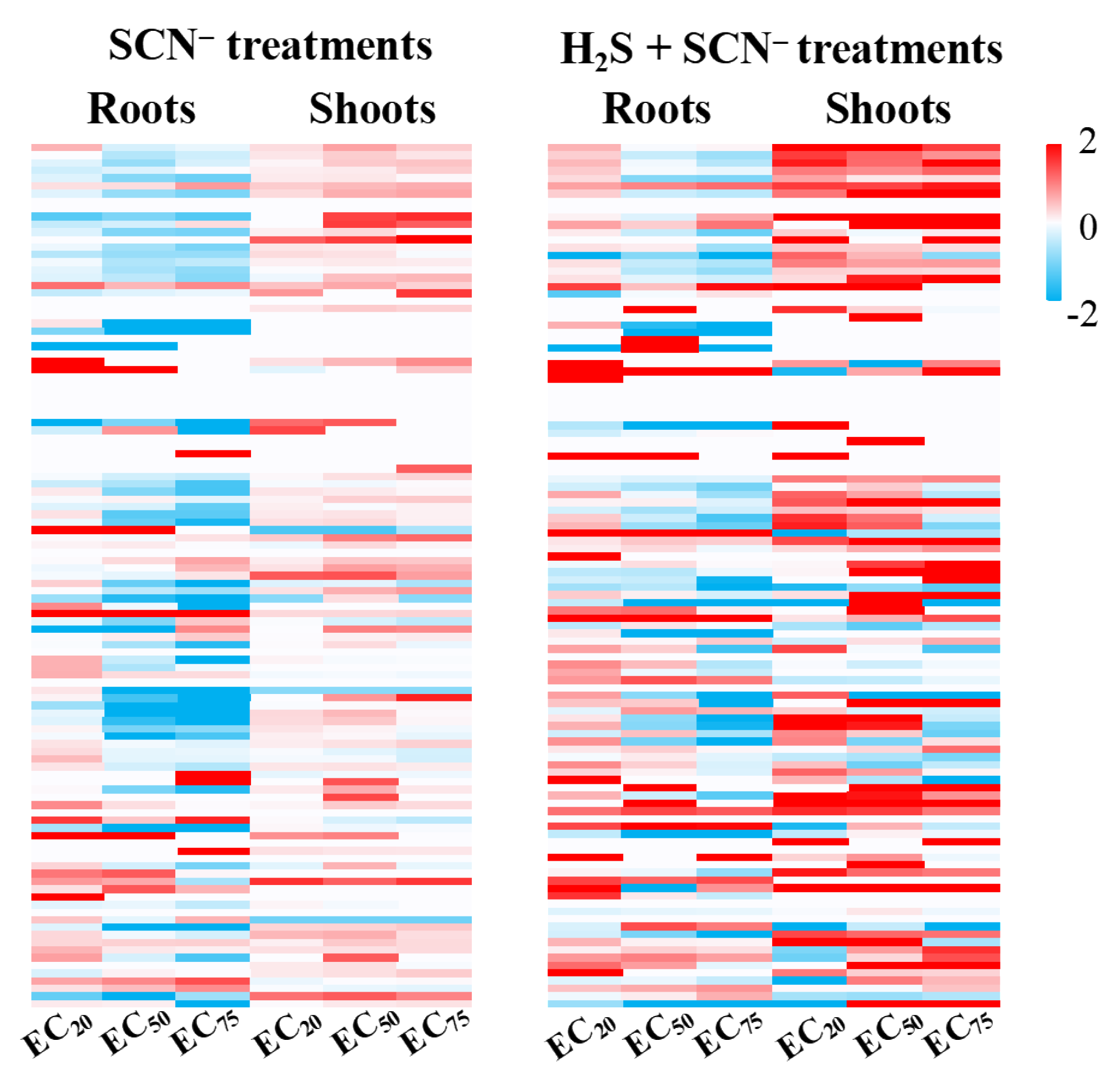
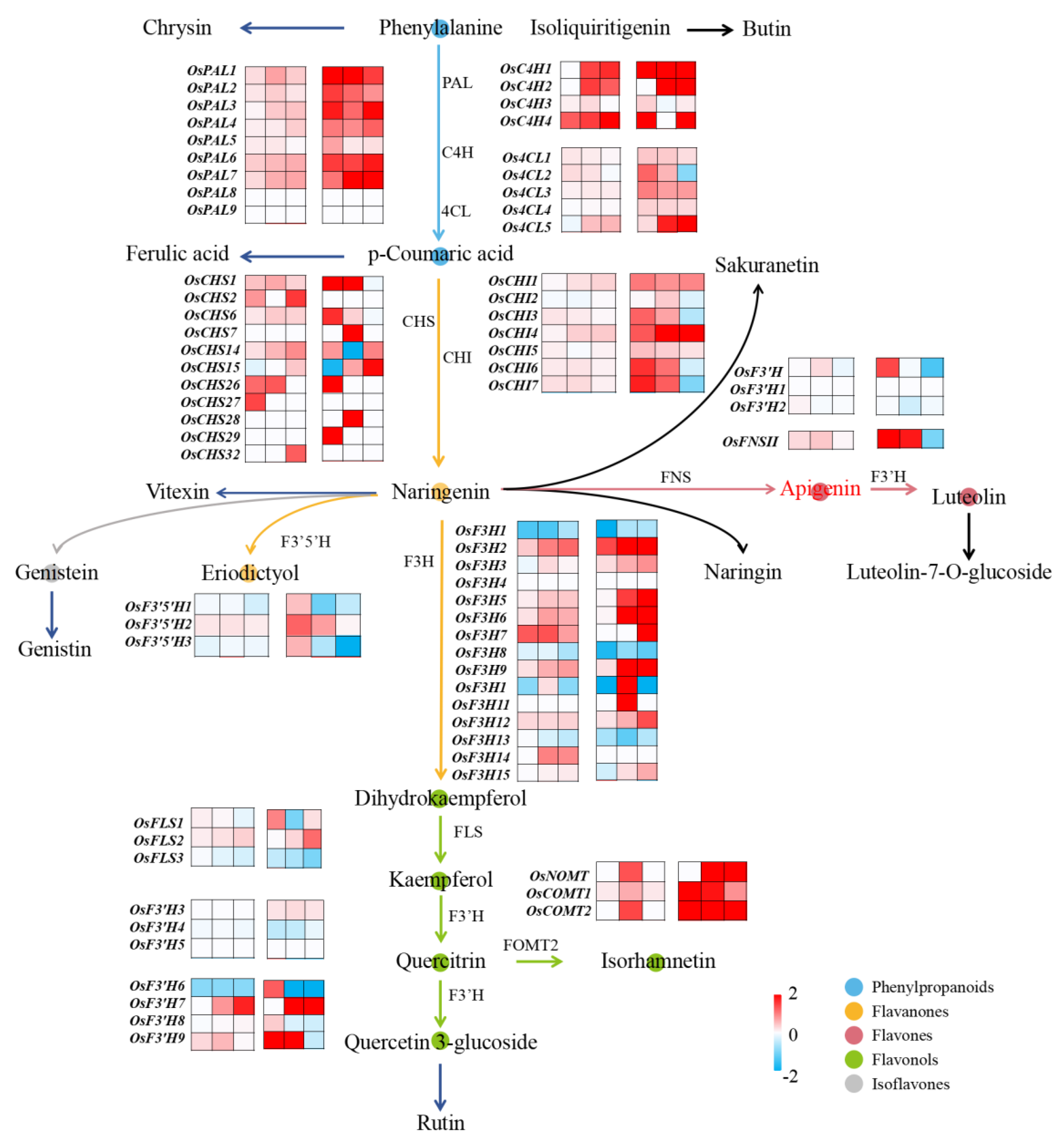
3.5. Response of Flavonoid-Biosynthesis- and Metabolism-Related Genes in Rice
3.6. Genetic Expression Variation Factors at Various EC Levels of SCN−
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, L.W.; Zhang, L.Z.; An, X.J.; Xiao, X.S.; Zhang, S.L.; Xu, Z.H.; Cai, H.X.; Zhang, Q.H. Thiocyanate-degrading microflora alleviates thiocyanate stress on tomato seedlings by improving plant and rhizosphere microenvironment. Environ. Pollut. 2023, 32, 116423. [Google Scholar] [CrossRef]
- Gupta, N.; Balomajumder, C.; Agarwal, V.K. Enzymatic mechanism and biochemistry for cyanide degradation: A review. J. Hazard. Mater. 2010, 176, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.J.; Yu, X.Z.; Li, Y.H.; Yang, L. Inhibition of the mitochondrial respiratory components (complex I and complex III) as stimuli to induce oxidative damage in Oryza sativa L. under thiocyanate exposure. Chemosphere 2020, 243, 125472. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Feng, Y.X.; Yu, X.Z.; Zhang, H.; Liang, Y.P. Effects of nitrogen fertilization on removal kinetics of thiocyanate (SCN−) in rice plants. Int. J. Environ. Sci. Technol. 2019, 17, 4291–4298. [Google Scholar] [CrossRef]
- Gao, J.J.; Wang, B.; Li, Z.J.; Xu, J.; Fu, X.Y.; Han, H.J.; Wang, L.J.; Zhang, W.H.; Deng, Y.D.; Wang, Y.; et al. Metabolic engineering of Oryza sativa for complete biodegradation of thiocyanate. Sci. Total Environ. 2022, 820, 153283. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.X.; Yang, L.; Lin, Y.J.; Song, Y.; Yu, X.Z. Merging the occurrence possibility into gene co-expression network deciphers the importance of exogenous 2-oxoglutarate in improving the growth of rice plants under thiocyanate stress. Front. Plant Sci. 2023, 14, 1086098. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.X.; Tian, P.; Lin, Y.J.; Cao, D.Y.; Li, C.Z.; Ullah, A. Gaseous signaling molecule H2S as a multitasking signal molecule in ROS metabolism of Oryza sativa under thiocyanate (SCN−) pollution. Environ. Pollut. 2024, 340, 122816. [Google Scholar] [CrossRef] [PubMed]
- King, L.; Wang, Q.; Xia, L.L.; Wang, P.; Jiang, G.H.; Li, W.Y.; Huang, Y.; Liang, X.L.; Peng, X.L.; Li, Y.G.; et al. Environmental exposure to perchlorate, nitrate and thiocyanate, and thyroid function in Chinese adults: A community-based cross-sectional study. Environ. Int. 2023, 171, 107713. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.W.; An, X.J.; Xiaoo, X.S.; Li, N.J.; Xie, D.; Lai, F.J.; Zhang, Q.H. Treatment of thiocyanate-containing wastewater: A critical review of thiocyanate destruction in industrial effluents. World J. Microb. Biot. 2023, 39, 35. [Google Scholar] [CrossRef]
- Bhunia, F.; Saha, N.C.; Kaviraj, A. Toxicity of thiocyanate to fish, plankton, worm, and aquatic ecosystem. Bull. Environ. Contam. Toxicol. 2000, 64, 197. [Google Scholar] [CrossRef]
- Yu, X.Z.; Zhang, F.Z. Effects of exogenous thiocyanate on mineral nutrients, antioxidative responses and free amino acids in rice plants. Ecotoxicology 2013, 22, 752–760. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.J.; Feng, Y.X.; Yu, X.Z. The importance of utilizing nitrate (NO3−) over ammonium (NH4+) as nitrogen source during detoxification of exogenous thiocyanate (SCN−) in Oryza sativa. Environ. Sci. Pollut. Res. 2022, 29, 5622–5633. [Google Scholar] [CrossRef] [PubMed]
- Afendi, F.M.; Okada, T.; Yamazaki, M.; Hirai-Morita, A.; Nakamura, Y.; Nakamura, K.; Ikeda, S.; Takahashi, H.; Altaf-Ul-Amin, M.; Darusman, L.K.; et al. KNApSAcK family databases: Integrated metabolite-plant species databases for multifaceted plant research. Plant Cell Physiol. 2012, 53, e1. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.X.; Li, Y.Y.; Dang, P.Q.; Zhao, S.J.; Lai, D.W.; Zhou, L.G. Rice secondary metabolites: Structures, roles, biosynthesis, and metabolic regulation. Molecules 2018, 23, 3098. [Google Scholar] [CrossRef] [PubMed]
- Yonekura-Sakakibara, K.; Higashi, Y.; Nakabayashi, R. The origin and evolution of plant flavonoid metabolism. Front. Plant Sci. 2019, 10, 468057. [Google Scholar] [CrossRef] [PubMed]
- Nagula, R.L.; Wairkar, S. Recent advances in topical delivery of flavonoids: A review. J. Control. Release 2019, 296, 190–201. [Google Scholar] [CrossRef]
- Wu, J.T.; Lv, S.D.; Zhao, L.; Gao, T.; Yu, C.; Hu, J.N.; Ma, F. Advances in the study of the function and mechanism of the action of flavonoids in plants under environmental stresses. Planta 2023, 257, 108. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.C.; Pinto, D.C.; Silva, A.M. Plant flavonoids: Chemical characteristics and biological activity. Molecules 2021, 26, 5377. [Google Scholar] [CrossRef]
- Agati, G.; Brunetti, C.; Fini, A.; Gori, A.; Guidi, L.; Landi, M.; Sebastiani, F.; Tattini, M. Are flavonoids effective antioxidants in plants? Twenty years of our investigation. Antioxidants 2020, 9, 1098. [Google Scholar] [CrossRef]
- Zhang, P.P.; Du, H.Y.; Wang, J.; Pu, Y.X.; Yang, C.Y.; Yan, R.J.; Yang, H.; Cheng, H.; Yu, D.Y. Multiplex CRISPR/Cas9-mediated metabolic engineering increases soya bean isoflavone content and resistance to soya bean mosaic virus. Plant Biotechnol. J. 2019, 18, 1384–1395. [Google Scholar] [CrossRef]
- Pourcel, L.; Routaboul, J.M.; Cheynier, V.; Lepiniec, L.; Debeaujon, I. Flavonoid oxidation in plants: From biochemical properties to physiological functions. Trends Plant Sci. 2007, 12, 29–36. [Google Scholar] [CrossRef]
- Kaya, C.; Ashraf, M.; Al-Huqail, A.A.; Alqahtani, M.A.; Ahmad, P. Silicon is dependent on hydrogen sulfide to improve boron toxicity tolerance in pepper plants by regulating the AsA-GSH cycle and glyoxalase system. Chemosphere 2020, 257, 127241. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Fahad, S.; Ali, M.A.; Hussain, S.; Tariq, M.; Ilyas, F.; Ahmed, S.; Saud, S.; Hammad, H.M.; Nasim, W.; et al. Hydrogen sulfide: A novel gaseous molecule for plant adaptation to stress. J. Plant Growth Regul. 2021, 40, 2485–2501. [Google Scholar] [CrossRef]
- Bhadwal, S.S.; Verma, S.; Hassan, S.; Kaur, S. Unraveling the potential of hydrogen sulfide as a signaling molecule for plant development and environmental stress responses: A state-of-the-art review. Plant Physiol. Biochem. 2024, 212, 108730. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Tian, M.; Han, Y. Hydrogen sulfide: A multi-tasking signal molecule in the regulation of oxidative stress responses. J. Exp. Bot. 2020, 71, 2862–2869. [Google Scholar] [PubMed]
- Lin, Y.J.; Feng, Y.X.; Li, Y.H.; Yu, G.; Yu, X.Z. Fuzzy synthetic evaluation of the impact of plant growth regulators on the root phenotype traits of rice plants under thiocyanate stress. Plant Physiol. Biochem. 2021, 158, 182–189. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Feng, Y.X.; Lin, Y.J.; Yu, X.Z. Comparative effects of sodium hydrosulfide and proline on functional repair in rice chloroplast through the D1 protein and thioredoxin system under simulated thiocyanate pollution. Chemosphere 2021, 284, 131389. [Google Scholar] [CrossRef] [PubMed]
- Tian, P.; Feng, Y.X.; Li, Y.H. Transcriptome reveals the crucial role of exogenous hydrogen sulfide in alleviation of thiocyanate (SCN−) toxicity in rice plants. Environ. Sci. Pollut. Res. 2023, 30, 26901–26913. [Google Scholar]
- Banerjee, A.; Tripathi, D.K.; Roychoudhury, A. Hydrogen sulphide trapeze: Environmental stress amelioration and phytohormone crosstalk. Plant Physiol. Biochem. 2018, 132, 46–53. [Google Scholar] [CrossRef]
- Wang, C.L.; Deng, Y.Z.; Liu, Z.S.; Liao, W.B. Hydrogen sulfide in plants: Crosstalk with other signal molecules in response to abiotic stresses. Int. J. Mol. Sci. 2021, 22, 12068. [Google Scholar] [CrossRef]
- Ge, Y.; Hu, K.D.; Wang, S.S.; Hu, L.Y.; Chen, X.Y.; Li, Y.H.; Yang, Y.; Yang, F.; Zhang, H. Hydrogen sulfide alleviates postharvest ripening and senescence of banana by antagonizing the effect of ethylene. PLoS ONE 2017, 12, e0180113. [Google Scholar] [CrossRef] [PubMed]
- Fakhari, S.; Sharifi, M.; De Michele, R.; Ghanati, F.; Safaie, N.; Sadeghnezhad, E. Hydrogen sulfide directs metabolic flux towards the lignan biosynthesis in Linum album hairy roots. Plant Physiol. Biochem. 2019, 135, 359–371. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Feng, Y.X.; Lin, Y.J.; Yu, X.Z. Indigenous proline is a two-dimensional safety-relief valve in balancing specific amino acids in rice under hexavalent chromium stress. J. Agric. Food Chem. 2021, 69, 11185–11195. [Google Scholar] [CrossRef] [PubMed]
- Novair, S.B.; Cheraghi, M.; Faramarzi, F.; Lajayer, B.A.; Senapathi, V.; Astatkie, T.; Price, G.W. Reviewing the role of biochar in paddy soils: An agricultural and environmental perspective. Ecotox Environ. Saf. 2023, 263, 115228. [Google Scholar] [CrossRef]
- Tian, P.; Feng, Y.X.; Li, C.Z.; Zhang, P.; Yu, X.Z. Transcriptional analysis of heavy metal P1B-ATPases (HMAs) elucidates competitive interaction in metal transport between cadmium and mineral elements in rice plants. Environ. Sci. Pollut. Res. 2023, 30, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Ling, Q.L.; Feng, Y.X.; Lu, C.J.; Lin, Y.J.; Yu, X.Z. Genetic variation and gene expression of anthocyanin synthesis and transport related enzymes in Oryza sativa against thiocyanate. Plant Physiol. Biochem. 2021, 160, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Abubakar, A.S.; Chen, J.K.; Chen, P.; Chen, K.M.; Yu, C.M.; Wang, X.F.; Qiu, X.J.; Huang, X.Y.; Shao, D.Y.; et al. Comparative genome and metabolome analyses uncover the evolution and flavonoid biosynthesis between Apocynum venetum and Apocynum hendersonii. Iscience 2023, 26, 106772. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.J.; Feng, Y.X.; Li, Y.H.; Lin, Y.J.; Yu, X.Z. Unraveling genes promoting ROS metabolism in subcellular organelles of Oryza sativa in response to trivalent and hexavalent chromium. Sci. Total Environ. 2020, 744, 140951. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Feng, Y.X.; Lin, Y.J.; Yu, X.Z. Mathematical quantification of interactive complexity of transcription factors involved in proline-mediated regulative strategies in Oryza sativa under chromium stress. Plant Physiol. Biochem. 2022, 182, 36–44. [Google Scholar] [CrossRef]
- Yu, X.Z.; Chu, Y.P.; Zhang, H.; Lin, Y.J.; Tian, P. Jasmonic acid and hydrogen sulfide modulate transcriptional and enzymatic changes of plasma membrane NADPH oxidases (NOXs) and decrease oxidative damage in Oryza sativa L. during thiocyanate exposure. Ecotoxicology 2021, 30, 1511–1520. [Google Scholar] [CrossRef]
- Sytar, O.; Kumari, P.; Yadav, S.; Brestic, M.; Rastogi, A. Phytohormone priming: Regulator for heavy metal stress in plants. J. Plant Growth Regul. 2019, 38, 739–752. [Google Scholar] [CrossRef]
- Feng, Y.X.; Li, C.Z.; Yang, L.; Yu, X.Z. Spatial-temporal variations of proline and related amino acids elucidate different nitrogenous utilization strategies in rice during detoxification of exogenous cyanide. Chem. Biol. Interact. 2023, 369, 110267. [Google Scholar] [CrossRef]
- Dong, N.Q.; Lin, H.X. Contribution of phenylpropanoid metabolism to plant development and plant–environment interactions. J. Integr. Plant Biol. 2021, 63, 180–209. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.A.; Grayer, R.J. Anthocyanins and other flavonoids. Nat. Product. Rep. 2004, 21, 539–573. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.X.; Liu, H.; Guo, H.Y.; Musante, C.; Coskun, S.H.; Nelson, B.C.; White, J.C.; Dhankher, O.P. Defense mechanisms and nutrient displacement in Arabidopsis thaliana upon exposure to CeO2 and In2O3 nanoparticles. Environ. Sci. Nano 2016, 3, 1369–1379. [Google Scholar] [CrossRef]
- Wohl, J.; Petersen, M. Functional expression and characterization of cinnamic acid 4-hydroxylase from the hornwort Anthoceros agrestis in Physcomitrella patens. Plant Cell Rep. 2020, 39, 597–607. [Google Scholar] [CrossRef]
- Mizutani, M.; Ohta, D.; Sato, R. Isolation of a cDNA and a genomic clone encoding cinnamate 4-hydroxylase from Arabidopsis and its expression manner in planta. Plant Physiol. 1997, 113, 755–763. [Google Scholar] [CrossRef]
- Alvarez-Rivera, G.; Sanz, A.; Cifuentes, A.; Ibánez, E.; Paape, T.; Lucas, M.M.; Pueyo, J.J. Flavonoid accumulation varies in Medicago truncatula in response to mercury stress. Front. Plant Sci. 2022, 13, 933209. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Fokou, P.V.T.; Sharifi-Rad, M.; Zucca, P.; Pezzani, R.; Martins, N.; Sharifi-Rad, J. The therapeutic potential of naringenin: A review of clinical trials. Pharmaceuticals 2019, 12, 11. [Google Scholar] [CrossRef]
- Wang, F.B.; Ren, G.L.; Li, F.S.; Qi, S.T.; Xu, Y.; Wang, B.W.; Yang, Y.L.; Ye, Y.X.; Zhou, Q.; Chen, X.H. A chalcone synthase gene AeCHS from Abelmoschus esculentus regulates flavonoid accumulation and abiotic stress tolerance in transgenic Arabidopsis. Acta Physiol. Plant 2018, 40, 97. [Google Scholar] [CrossRef]
- Hodaei, M.; Rahimmalek, M.; Arzani, A.; Talebi, M. The effect of water stress on phytochemical accumulation, bioactive compounds and expression of key genes involved in flavonoid biosynthesis in Chrysanthemum morifolium L. Ind. Crop Prod. 2018, 120, 295–304. [Google Scholar] [CrossRef]
- Mekawy, A.M.M.; Abdelaziz, M.N.; Ueda, A. Apigenin pretreatment enhances growth and salinity tolerance of rice plants. Plant Physiol. Biochem. 2018, 130, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Seelinger, G.; Merfort, I.; Schempp, C.M. Antioxidant, anti-inflammatory and anti-allergic activities of luteolin. Planta Med. 2008, 74, 1667–1677. [Google Scholar] [CrossRef] [PubMed]
- Song, X.Y.; Diao, J.J.; Ji, J.; Wang, G.; Guan, C.F.; Jin, C.; Wang, Y.R. Molecular cloning and identification of a flavanone 3-hydroxylase gene from Lycium chinense, and its overexpression enhances drought stress in tobacco. Plant Physiol. Biochem. 2016, 98, 89–100. [Google Scholar] [CrossRef]
- Falcone Ferreyra, M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2022, 3, 222. [Google Scholar] [CrossRef]
- Gai, Z.S.; Wang, Y.; Ding, Y.Q.; Qian, W.J.; Qiu, C.; Xie, H.; Sun, L.T.; Jiang, Z.W.; Ma, Q.P.; Wang, L.J.; et al. Exogenous abscisic acid induces the lipid and flavonoid metabolism of tea plants under drought stress. Sci. Rep. 2020, 10, 12275. [Google Scholar] [CrossRef]
- Yang, W.L.; Li, N.; Fan, Y.X.; Dong, B.Y.; Song, Z.H.; Cao, H.Y.; Du, T.T.; Liu, T.Y.; Qi, M.; Niu, L.L.; et al. Transcriptome analysis reveals abscisic acid enhancing drought resistance by regulating genes related to flavonoid metabolism in pigeon pea. Environ. Exp. Bot. 2021, 191, 104627. [Google Scholar] [CrossRef]
- Malla, A.M.; Dar, B.A.; Isaev, A.B.; Lone, Y.; Banday, M.R. Flavonoids: A reservoir of drugs from nature. Mini Rev. Med. Chem. 2023, 23, 772–786. [Google Scholar] [CrossRef]
- Hayyan, M.; Hashim, M.A.; AlNashef, I.M. Superoxide ion: Generation and chemical implications. Chem. Rev. 2016, 116, 3029–3085. [Google Scholar] [CrossRef]
- Gulcin, İ. Antioxidants and antioxidant methods: An updated overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, P.; Feng, Y.-X.; Li, Y.-H. SOS! Hydrogen Sulfide Enhances the Flavonoid Early Warning System in Rice Plants to Cope with Thiocyanate Pollution. Toxics 2024, 12, 591. https://doi.org/10.3390/toxics12080591
Tian P, Feng Y-X, Li Y-H. SOS! Hydrogen Sulfide Enhances the Flavonoid Early Warning System in Rice Plants to Cope with Thiocyanate Pollution. Toxics. 2024; 12(8):591. https://doi.org/10.3390/toxics12080591
Chicago/Turabian StyleTian, Peng, Yu-Xi Feng, and Yan-Hong Li. 2024. "SOS! Hydrogen Sulfide Enhances the Flavonoid Early Warning System in Rice Plants to Cope with Thiocyanate Pollution" Toxics 12, no. 8: 591. https://doi.org/10.3390/toxics12080591
APA StyleTian, P., Feng, Y.-X., & Li, Y.-H. (2024). SOS! Hydrogen Sulfide Enhances the Flavonoid Early Warning System in Rice Plants to Cope with Thiocyanate Pollution. Toxics, 12(8), 591. https://doi.org/10.3390/toxics12080591