
Zinc, Zinc Transporters, and Cadmium Cytotoxicity in a Cell Culture Model of Human Urothelium


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cells and Culture Maintenance
2.2. Quantitative Cell Viability Assay
2.3. Quantification of ZIP and ZnT Zinc Transporter Gene Expression
2.4. Statistical Analysis
3. Results
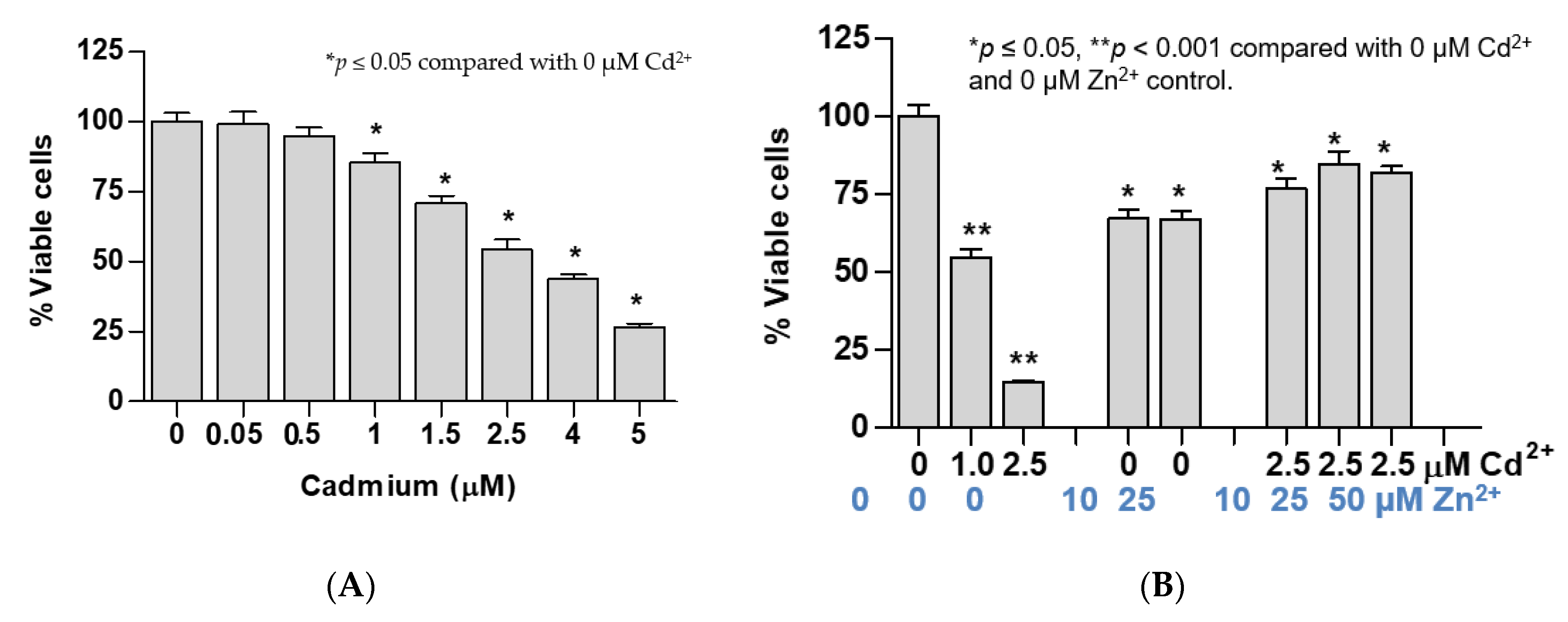
3.1. Dose-Response Analysis of Effects of Cadmium on Expression Levels of Zinc Transporters
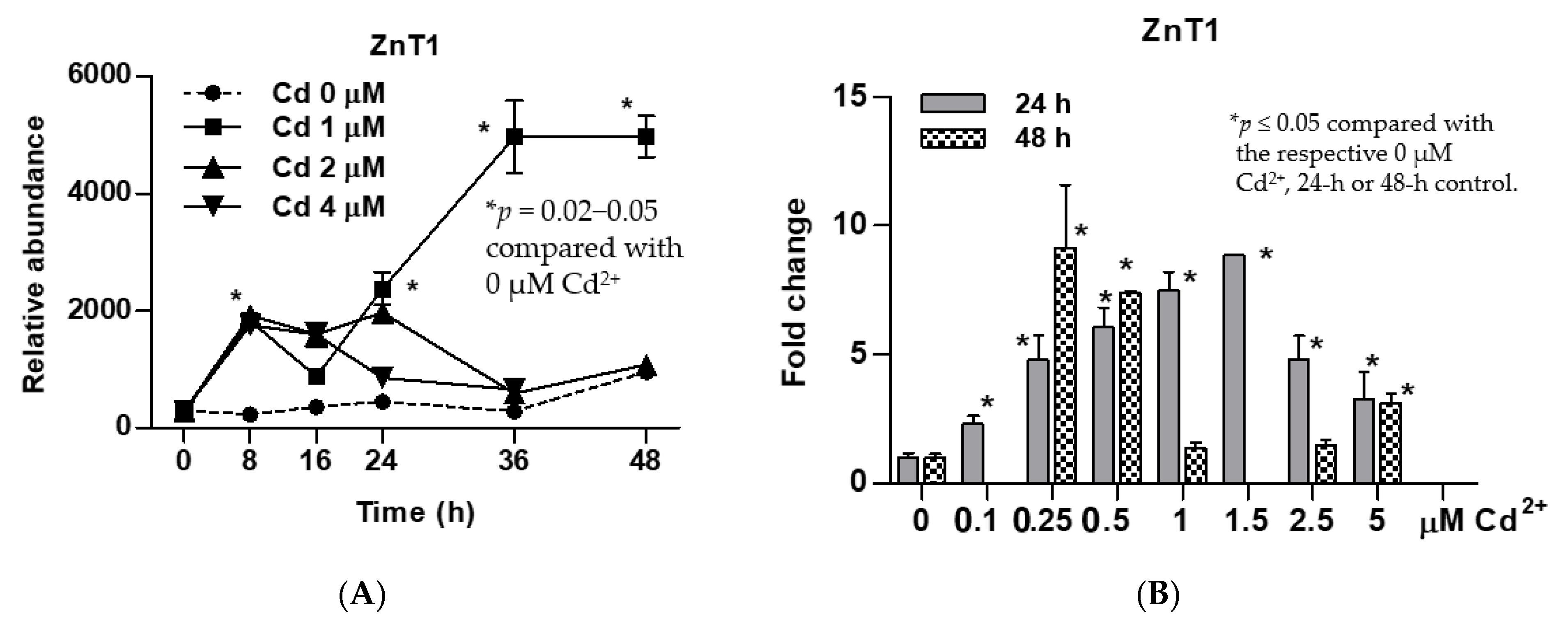
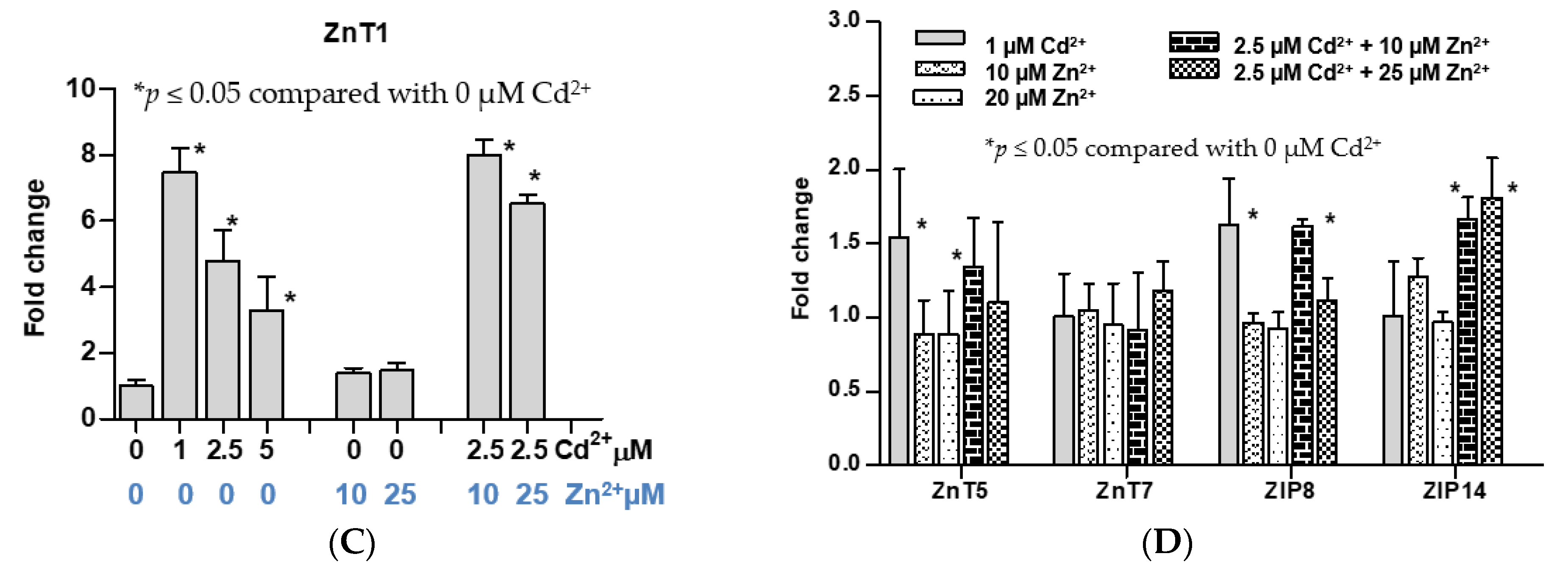
3.2. Time-Course and Dose-Response Analyses for Induced Expression of the ZnT1 Gene by Cadmium
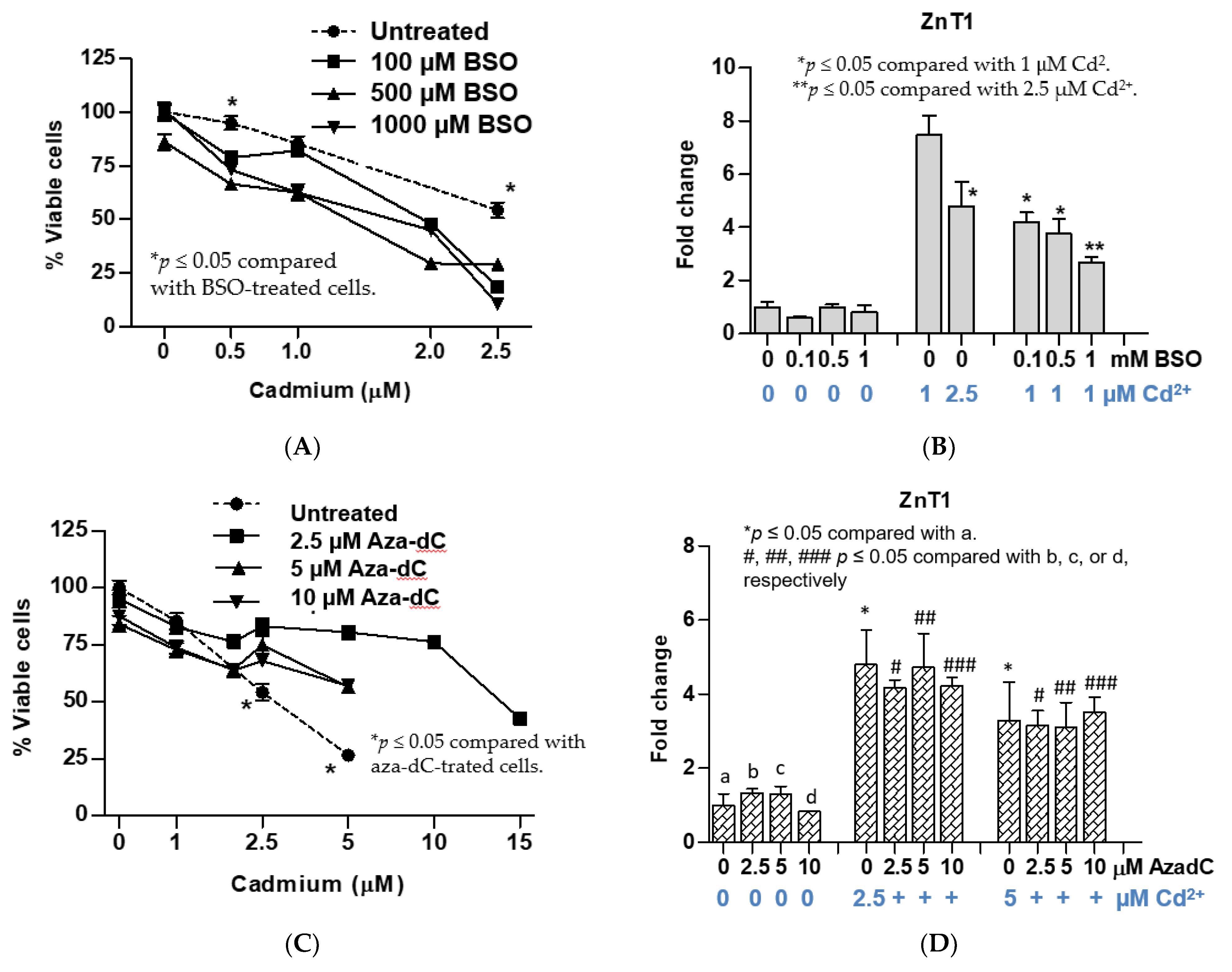
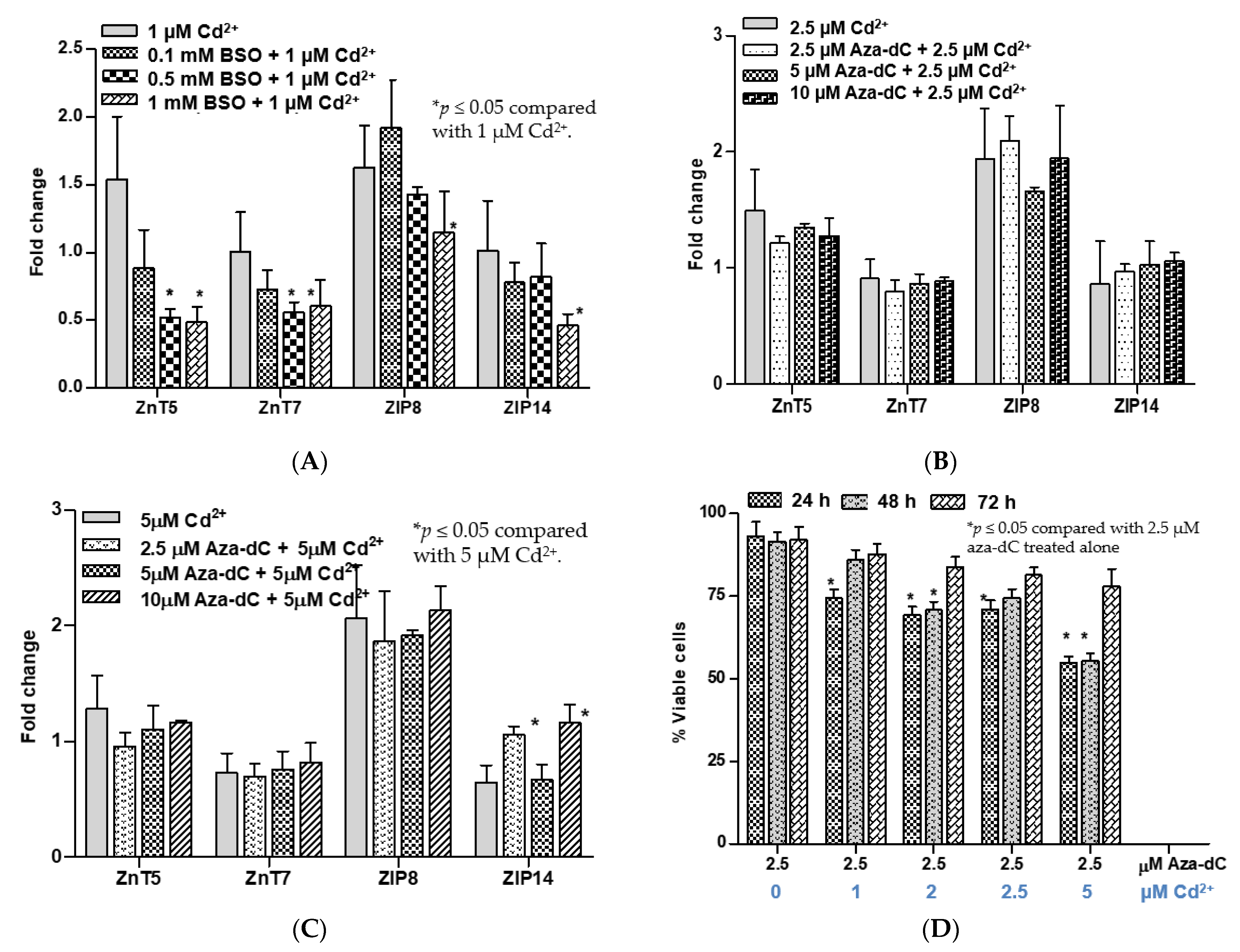
3.3. Effects of Pretreatment with BSO and Aza-dC on Cytotoxicity of Cadmium and Expression of ZnT1 Gene
3.4. Other Notable Effects of BSO or Aza-dC Pretreatment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Satarug, S.; Phelps, K.R. Cadmium Exposure and Toxicity. In Metal Toxicology Handbook; Bagchi, D., Bagchi, M., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 219–274. [Google Scholar]
- Satarug, S.; Gobe, G.C.; Vesey, D.A.; Phelps, K.R. Cadmium and lead exposure, nephrotoxicity, and mortality. Toxics 2020, 8, 86. [Google Scholar] [CrossRef]
- Satarug, S.; Vesey, D.A.; Gobe, G.C. Current health risk assessment practice for dietary cadmium: Data from different countries. Food Chem. Toxicol. 2017, 106, 430–445. [Google Scholar] [CrossRef]
- Thevenod, F.; Fels, J.; Lee, W.-K.; Zarbock, R. Channels, transporters and receptors for cadmium and cadmium complexes in eukaryotic cells: Myths and facts. Biometals 2019, 32, 469–489. [Google Scholar] [CrossRef]
- Flanagan, P.R.; McLellan, J.S.; Haist, J.; Cherian, M.G.; Chamberlain, M.J.; Valberg, L.S. Increased dietary cadmium absorption in mice and human subjects with iron deficiency. Gastroenterology 1978, 46, 609–623. [Google Scholar] [CrossRef]
- Finley, J.W. Manganese absorption and retention by young women is associated with serum ferritin concentration. Am. J. Clin. Nutr. 1999, 70, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Fujishiro, H.; Yano, Y.; Takada, Y.; Tanihara, M.; Himeno, S. Roles of ZIP8, ZIP14, and DMT1 in transport of cadmium and manganese in mouse kidney proximal tubule cells. Metallomics 2012, 4, 700–708. [Google Scholar] [CrossRef] [PubMed]
- Fujishiro, H.; Hamao, S.; Tanaka, R.; Kambe, T.; Himeno, S. Concentration-dependent roles of DMT1 and ZIP14 in cadmium absorption in Caco-2 cells. J. Toxicol. Sci. 2017, 42, 559–567. [Google Scholar] [CrossRef] [Green Version]
- Fujishiro, H.; Himeno, S. New insights into the roles of ZIP8, a cadmium and manganese transporter, and its relation to human diseases. Biol. Pharm. Bull. 2019, 42, 1076–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkitkasemwong, S.; Wang, C.Y.; Mackenzie, B.; Knutson, M.D. Physiologic implications of metal-ion transport by ZIP14 and ZIP8. Biometals 2012, 25, 643–655. [Google Scholar] [CrossRef] [Green Version]
- Schneider, S.N.; Liu, Z.; Wang, B.; Miller, M.L.; Afton, S.E.; Soleimani, M.; Nebert, D.W. Oral cadmium in mice carrying 5 versus 2 copies of the Slc39a8 gene: Comparison of uptake, distribution, metal content, and toxicity. Int. J. Toxicol. 2014, 33, 14–20. [Google Scholar] [CrossRef]
- Zhang, R.; Witkowska, K.; Afonso Guerra-Assunção, J.; Ren, M.; Ng, F.L.; Mauro, C.; Tucker, A.T.; Caulfield, M.J.; Shu Ye, S. A blood pressure-associated variant of the SLC39A8 gene influences cellular cadmium accumulation and toxicity. Hum. Mol. Genet. 2016, 25, 4117–4126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aydemir, T.B.; Cousins, R.J. The multiple faces of the metal transporter ZIP14 (SLC39A14). J. Nutr. 2018, 148, 174–184. [Google Scholar] [CrossRef]
- Rentschler, G.; Kippler, M.; Axmon, A.; Raqib, R.; Skerfving, S.; Vahter, M.; Broberg, K. Cadmium concentrations in human blood and urine are associated with polymorphisms in zinc transporter genes. Metallomics 2014, 6, 885–891. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.S.; Caffrey, J.L.; Lin, J.W.; Bayliss, D.; Faramawi, M.F.; Bateson, T.F.; Sonawane, B. Increased risk of cancer mortality associated with cadmium exposures in older Americans with low zinc intake. J. Toxicol. Environ. Health A 2013, 76, 1–15. [Google Scholar] [CrossRef]
- Min, K.S.; Sano, E.; Ueda, H.; Sakazaki, F.; Yamada, K.; Takano, M.; Tanaka, K. Dietary deficiency of calcium and/or iron, an age-related risk factor for renal accumulation of cadmium in mice. Biol. Pharm. Bull. 2015, 38, 1557–1563. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.S.; Ho, W.C.; Caffrey, J.L.; Sonawane, B. Low serum zinc is associated with elevated risk of cadmium nephrotoxicity. Environ. Res. 2014, 134, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.S.; Caffrey, J.L.; Chang, M.H.; Dowling, N.; Lin, J.W. Cigarette smoking, cadmium exposure, and zinc intake on obstructive lung disorder. Respir. Res. 2010, 11, 53. [Google Scholar] [CrossRef] [Green Version]
- Petzoldt, L.; Leigh, I.M.; Duffy, P.G.; Sexton, C.; Masters, R.W. Immortalisation of human urothelial cells. Urol. Res. 1995, 23, 377–380. [Google Scholar] [CrossRef]
- Rossi, M.R.; Masters, J.R.W.; Park, S.; Todd, J.H.; Garrett, S.H.; Sens, M.A.; Somji, S.; Nath, J.; Sens, D.A. The immortalized UROtsa cell line as a potential cell culture model of human urothelium. Environ. Health Perspect. 2001, 109, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Sens, D.A.; Park, S.; Gurel, V.; Sens, M.A.; Garrett, S.H.; Somji, S. Inorganic cadmium- and arsenite-induced malignant transformation of human bladder urothelial cells. Toxicol. Sci. 2004, 79, 56–63. [Google Scholar] [CrossRef] [Green Version]
- Hoggarth, Z.E.; Osowski, D.B.; Freeberg, B.A.; Garrett, S.H.; Sens, D.A.; Sens, M.A.; Zhou, X.D.; Zhang, K.K.; Somji, S. The urothelial cell line UROtsa transformed by arsenite and cadmium display basal characteristics associated with muscle invasive urothelial cancers. PLoS ONE 2018, 13, e0207877. [Google Scholar] [CrossRef] [Green Version]
- Ohana, E.; Sekler, I.; Kaisman, T.; Kahn, N.; Cove, J.; Silverman, W.F.; Amsterdam, A.; Hershfinkel, M. Silencing of ZnT-1 expression enhances heavy metal influx and toxicity. J. Mol. Med. 2006, 84, 753–763. [Google Scholar] [CrossRef]
- Nishito, Y.; Kambe, T. Zinc transporter 1 (ZNT1) expression on the cell surface is elaborately controlled by cellular zinc levels. J. Biol. Chem. 2019, 294, 15686–15697. [Google Scholar] [CrossRef]
- Aiba, I.; Hossain, A.; Kuo, M.T. Elevated GSH level increases cadmium resistance through down-regulation of Sp1-dependent expression of the cadmium transporter ZIP8. Mol. Pharmacol. 2008, 74, 823–833. [Google Scholar] [CrossRef] [Green Version]
- Sens, D.; Rossi, M.; Park, S.; Gurel, V.; Nath, J.; Garrett, S.; Sens, M.A.; Somji, S. Metallothionein isoform 1 and 2 gene expression in a human urothelial cell line (UROtsa) exposed to CdCl2 and NaAsO2. J. Toxicol. Environ. Health A 2003, 66, 2031–2046. [Google Scholar] [CrossRef]
- Lertratanangkoon, K.; Wu, C.J.; Savaraj, N.; Thomas, M.L. Alterations of DNA methylation by glutathione depletion. Cancer Lett. 1997, 120, 149–156. [Google Scholar] [CrossRef]
- Waalkes, M.P.; Miller, M.S.; Wilson, M.J.; Bare, R.M.; McDowell, A.E. Increased metallothionein gene expression in 5-aza-2’-deoxycytidine-induced resistance to cadmium cytotoxicity. Chem. Biol. Interact. 1998, 66, 189–204. [Google Scholar]
- Garrett, S.H.; Somji, S.; Sens, D.A.; Zhang, K.K. Prediction of the number of activated genes in multiple independent Cd+2 and As+3-induced malignant transformations of human urothelial cells (UROtsa). PLoS ONE 2014, 9, e85614. [Google Scholar] [CrossRef]
- Berridge, M.V.; Herst, P.M.; Tan, A.S. Tetrazolium dyes as tools in cell biology: New insights into their cellular reduction, Biotech. Ann. Rev. 2005, 11, 127–152. [Google Scholar]
- Albrecht, A.L.; Somji, S.; Sens, M.A.; Sens, D.A.; Garrett, S.H. Zinc transporter mRNA expression in the RWPE-1 human prostate epithelial cell line. Biometals 2008, 21, 405–416. [Google Scholar] [CrossRef]
- Ajjimaporn, A.; Botsford, T.; Garrett, S.H.; Sens, M.A.; Zhou, X.D.; Dunlevy, J.R.; Sens, D.A.; Somji, S. ZIP8 expression in human proximal tubule cells, human urothelial cells transformed by Cd+2 and As+3 and in specimens of normal human urothelium and urothelial cancer. Cancer Cell Int. 2012, 12, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takiguchi, M.; Cherrington, N.J.; Hartley, D.P.; Klaassen, C.D.; Waalkes, M.P. Cyproterone acetate induces a cellular tolerance to cadmium in rat liver epithelial cells involving reduced cadmium accumulation. Toxicology 2001, 165, 13–25. [Google Scholar] [CrossRef]
- Fujishiro, H.; Okugaki, S.; Yasumitsu, S.; Enomoto, S.; Himeno, S. Involvement of DNA hypermethylation in down-regulation of the zinc transporter ZIP8 in cadmium-resistant metallothionein-null cells. Toxicol. Appl. Pharmacol. 2009, 241, 195–201. [Google Scholar] [CrossRef]
- Fujishiro, H.; Ohashi, T.; Takuma, M.; Himeno, S. Suppression of ZIP8 expression is a common feature of cadmium-resistant and manganese-resistant RBL-2H3 cells. Metallomics 2013, 5, 437–444. [Google Scholar] [CrossRef]
- Satarug, S.; Kikuchi, M.; Wisedpanichkij, R.; Li, B.; Takeda, K.; Na-Bangchang, K.; Moore, M.R.; Hirayama, K.; Shibahara, S. Prevention of cadmium accumulation in retinal pigment epithelium with manganese and zinc. Exp. Eye Res. 2008, 87, 587–593. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zinc Transporters | Cadmium Concentration, µM | p-Values | |||
|---|---|---|---|---|---|
| 0 | 1 | 2 | 4 | ||
| SLC30A family | |||||
| ZnT1 | 365 ± 38 | 3007 ± 465 | 1434 ± 146 | 1216 ± 153 | <0.001 |
| ZnT2 | 0.06 ± 0.01 | 73 ± 15 | 16 ± 1.9 | 11 ± 1.5 | <0.001 |
| ZnT3 | 0.15 ± 0.01 | 0.24 ± 0.05 | 0.10 ± 0.02 | 0.10 ± 0.02 | 0.03 |
| ZnT4 | 11.4 ± 0.8 | 10 ± 1 | 8.7 ± 1 | 6.2 ± 0.5 | 0.001 |
| ZnT5 | 510 ± 30 | 1038 ± 132 | 495 ± 54 | 568 ± 91 | 0.001 |
| ZnT6 | 65 ± 8 | 77± 6 | 63 ± 13 | 57 ± 12 | 0.174 |
| ZnT7 | 758 ± 76 | 1007 ±136 | 706 ± 44 | 488 ± 63 | 0.02 |
| ZnT10 | 1.1 ± 0.2 | 2.4 ± 0.2 | 1.7 ± 0.2 | 1.1 ± 0.1 | <0.001 |
| SLC39A family | |||||
| ZIP1 | 82 ± 9 | 99 ± 15 | 55 ± 10 | 59 ± 12 | 0.02 |
| ZIP2 | 1.2 ± 0.1 | 0.8 ± 0.2 | 0.4 ± 0.1 | 0.2 ± 0.03 | <0.001 |
| ZIP3A | 19 ± 1 | 23 ± 2.3 | 17 ± 1.7 | 14 ± 1.3 | 0.01 |
| ZIP3B | 4.1 ± 0.2 | 6.2 ± 0.7 | 4.4 ± 0.2 | 4.2 ± 0.4 | 0.02 |
| ZIP4 | 0.06 ± 0.01 | 0.06 ± 0.01 | 0.04 ± 0.01 | 0.06 ± 0.01 | 0.40 |
| ZIP5 | 0.01 ± 0.001 | 0.01 ± 0.003 | 0.01 ± 0.002 | 0.01 ±.002 | 0.28 |
| ZIP6 | 92 ± 8 | 133 ± 12 | 80 ± 9 | 75 ± 10 | 0.001 |
| ZIP7 | 204 ± 25 | 342 ± 69 | 149 ± 32 | 94 ± 21 | <0.001 |
| ZIP8 | 2.1 ± 0.2 | 2.6 ± 0.3 | 2.0 ± 0.2 | 2.7 ± 0.4 | 0.50 |
| ZIP10 | 54 ± 4 | 30 ± 8 | 14 ± 3 | 14 ± 3 | <0.001 |
| ZIP14 | 146 ± 19 | 218 ± 24 | 158 ± 26 | 128 ± 19 | 0.01 |
| SLC30A Family | Concentration of Cd2+ (0, 1, 2, 4 µM) | SLC39A Family | Concentration of Cd2+ (0, 1, 2, 4 µM) | ||
|---|---|---|---|---|---|
| r | p-Values | r | p-Values | ||
| ZnT1 | 0.51 | <0.001 | ZIP1 | −0.29 | 0.02 |
| ZnT2 | 0.10 | 0.33 | ZIP2 | −0.69 | <0.001 |
| ZnT3 | −0.28 | 0.03 | ZIP3A | −0.33 | 0.01 |
| ZnT4 | −0.52 | <0.001 | ZIP3B | 0.10 | 0.44 |
| ZnT5 | −0.01 | 0.92 | ZIP4 | −0.08 | 0.52 |
| ZnT6 | −0.07 | 0.56 | ZIP5 | −0.27 | 0.05 |
| ZnT7 | −0.24 | 0.06 | ZIP6 | −0.19 | 0.13 |
| ZnT10 | 0.11 | 0.40 | ZIP7 | −0.42 | 0.001 |
| ZIP8 | 0.04 | 0.622 | |||
| ZIP10 | −0.68 | <0.001 | |||
| ZIP14 | −0.03 | 0.84 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Satarug, S.; Garrett, S.H.; Somji, S.; Sens, M.A.; Sens, D.A. Zinc, Zinc Transporters, and Cadmium Cytotoxicity in a Cell Culture Model of Human Urothelium. Toxics 2021, 9, 94. https://doi.org/10.3390/toxics9050094
Satarug S, Garrett SH, Somji S, Sens MA, Sens DA. Zinc, Zinc Transporters, and Cadmium Cytotoxicity in a Cell Culture Model of Human Urothelium. Toxics. 2021; 9(5):94. https://doi.org/10.3390/toxics9050094
Chicago/Turabian StyleSatarug, Soisungwan, Scott H. Garrett, Seema Somji, Mary Ann Sens, and Donald A. Sens. 2021. "Zinc, Zinc Transporters, and Cadmium Cytotoxicity in a Cell Culture Model of Human Urothelium" Toxics 9, no. 5: 94. https://doi.org/10.3390/toxics9050094
APA StyleSatarug, S., Garrett, S. H., Somji, S., Sens, M. A., & Sens, D. A. (2021). Zinc, Zinc Transporters, and Cadmium Cytotoxicity in a Cell Culture Model of Human Urothelium. Toxics, 9(5), 94. https://doi.org/10.3390/toxics9050094