Modeling the Potential of Submarine Groundwater Discharge to Facilitate Growth of Vibrio cholerae Bacteria


Abstract
1. Introduction
2. Materials and Methods
2.1. Setup
2.2. Governing Equations
2.3. Parametrization
2.4. Model Evaluation
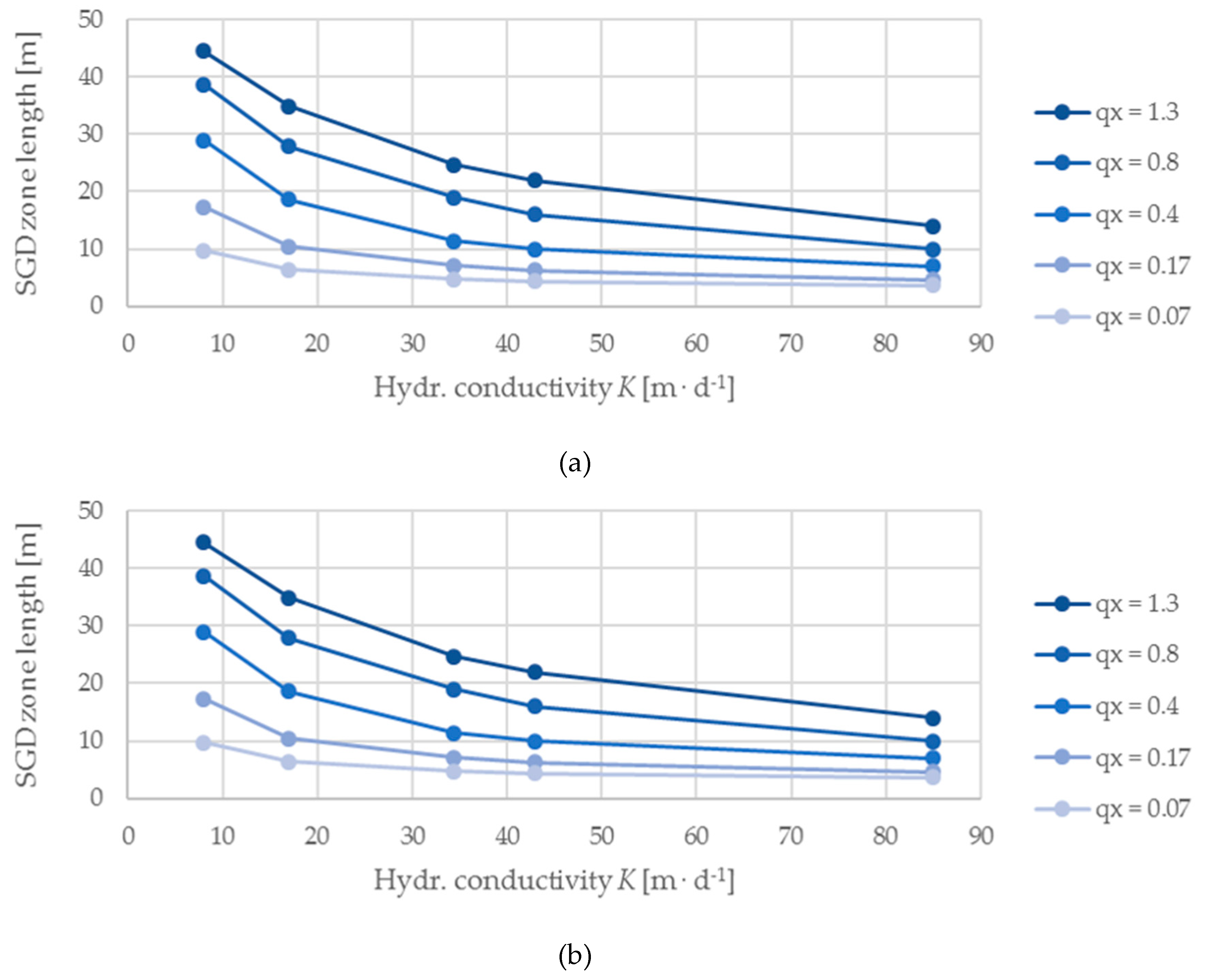
3. Results
4. Discussion
5. Conclusions
- Sufficient fresh groundwater inflow and hydraulic conductivity of a coastal aquifer are the most important controls that govern V. cholerae growth. Demanding a high freshwater supply (CS ≤ 100 mg L−1) to the coastal slope and thus a high hydraulic gradient between the hinterland and sea to displace saltwater intrusion, a preferably high groundwater inflow and low hydraulic conductivity within the bounds of a highly conductive material (e.g., sand) create the most favorable conditions for V. cholerae growth, which is not necessarily linked to the specific SGD flux.
- Although not significantly affecting the SGD volume, dispersion facilitates non-halophilic bacterial expansion at decreasing values, keeping the fresh groundwater flow low in salt by preventing mixing processes between fresh and saline groundwaters.
- Coastal slope had substantially less impact on the estimated habitat areas than the other analyzed parameters.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Burnett, W.C.; Bokuniewicz, H.; Huettel, M.; Moore, W.S.; Taniguchi, M. Groundwater and pore water inputs to the coastal zone. Biogeochemistry 2003, 66, 3–33. [Google Scholar] [CrossRef]
- Moore, W.S. The effect of submarine groundwater discharge on the ocean. Annu. Rev. Mar. Sci. 2010, 2, 59–88. [Google Scholar] [CrossRef] [PubMed]
- Burnett, W.C.; Aggarwal, P.K.; Aureli, A.; Bokuniewicz, H.; Cable, J.E.; Charette, M.A.; Moore, W.S.; Krupa, S.; Kulkarni, K.M.; Loveless, A.; et al. Quantifying submarine groundwater discharge in the coastal zone via multiple methods. Sci. Total Environ. 2006, 367, 498–543. [Google Scholar] [CrossRef] [PubMed]
- da Rocha, C.M.; Barboza, E.G.; Niencheski, L.F.H. Radon activity and submarine groundwater discharge in different geological regions of a coastal barrier in Southern Brazil. Environ. Earth Sci. 2018, 77, 527. [Google Scholar] [CrossRef]
- Taniguchi, M.; Burnett, W.C.; Cable, J.E.; Turner, J.V. Investigation of submarine groundwater discharge. Hydrol. Process. 2002, 16, 2115–2129. [Google Scholar] [CrossRef]
- Johannes, R.E. Ecological significance of the submarine discharge of groundwater. Mar. Ecol. Prog. Ser. 1980, 3, 365–373. [Google Scholar] [CrossRef]
- Li, X.; Hu, B.X.; Burnett, W.C.; Santos, I.R.; Chanton, J.P. Submarine ground water discharge driven by tidal pumping in a heterogeneous aquifer. Groundwater 2009, 47, 558–568. [Google Scholar] [CrossRef]
- Antonellini, M.; Mollema, P.; Giambastiani, B.; Bishop, K.; Caruso, L.; Minchio, A.; Pellegrini, L.; Sabia, M.; Ulazzi, E.; Gabbianelli, G. Salt water intrusion in the coastal aquifer of the southern Po Plain, Italy. Hydrogeol. J. 2008, 16, 1541. [Google Scholar] [CrossRef]
- Michael, H.A.; Mulligan, A.E.; Harvey, C.F. Seasonal oscillations in water exchange between aquifers and the coastal ocean. Nature 2005, 436, 1145. [Google Scholar] [CrossRef]
- Mulligan, A.E.; Charette, M.A. Intercomparison of submarine groundwater discharge estimates from a sandy unconfined aquifer. J. Hydrol. 2006, 327, 411–425. [Google Scholar] [CrossRef]
- Cable, J.E.; Burnett, W.C.; Chanton, J.P.; Corbett, D.R.; Cable, P.H. Field evaluation of seepage meters in the coastal marine environment. Estuar. Coast. Shelf Sci. 1997, 45, 367–375. [Google Scholar] [CrossRef]
- Walther, M.; Graf, T.; Kolditz, O.; Liedl, R.; Post, V. How significant is the slope of the sea-side boundary for modelling seawater intrusion in coastal aquifers? J. Hydrol. 2017, 551, 648–659. [Google Scholar] [CrossRef]
- Wilson, A.M. Fresh and saline groundwater discharge to the ocean: A regional perspective. Water Resour. Res. 2005, 41. [Google Scholar] [CrossRef]
- Santos, I.R.; Eyre, B.D.; Huettel, M. The driving forces of porewater and groundwater flow in permeable coastal sediments: A review. Estuar. Coast. Shelf Sci. 2012, 98, 1–15. [Google Scholar] [CrossRef]
- Adolf, J.E.; Burns, J.; Walker, J.K.; Gamiao, S. Near shore distributions of phytoplankton and bacteria in relation to submarine groundwater discharge-fed fishponds, Kona coast, Hawai‘i, USA. Estuar. Coast. Shelf Sci. 2019, 219, 341–353. [Google Scholar] [CrossRef]
- Lecher, A.; Mackey, K. Synthesizing the effects of submarine groundwater discharge on marine biota. Hydrology 2018, 5, 60. [Google Scholar] [CrossRef]
- Kopprio, G.A.; Streitenberger, M.E.; Okuno, K.; Baldini, M.; Biancalana, F.; Fricke, A.; Martínez, A.; Neogi, S.B.; Koch, B.P.; Yamasaki, S.; et al. Biogeochemical and hydrological drivers of the dynamics of Vibrio species in two Patagonian estuaries. Sci. Total Environ. 2017, 579, 646–656. [Google Scholar] [CrossRef]
- Cook, D.W. Effect of time and temperature on multiplication of Vibrio vulnificus in postharvest Gulf Coast shellstock oysters. Appl. Environ. Microb. 1994, 60, 3483–3484. [Google Scholar]
- Blackwell, K.D.; Oliver, J.D. The ecology of Vibrio vulnificus, Vibrio cholerae, and Vibrio parahaemolyticus in North Carolina estuaries. J. Microb. 2008, 46, 146–153. [Google Scholar] [CrossRef]
- Esteves, K.; Mosser, T.; Aujoulat, F.; Hervio-Heath, D.; Monfort, P.; Jumas-Bilak, E. Highly diverse recombining populations of Vibrio cholerae and Vibrio parahaemolyticus in French Mediterranean coastal lagoons. Front. Microb. 2015, 6, 708. [Google Scholar] [CrossRef]
- Adyasari, D.; Hassenrück, C.; Oehler, T.; Sabdaningsih, A.; Moosdorf, N. Microbial community structure associated with submarine groundwater discharge in northern Java (Indonesia). Sci. Total Environ. 2019. under review. [Google Scholar] [CrossRef]
- Morris, J.J. Non-O group 1 Vibrio cholerae: A look at the epidemiology of an occasional pathogen. Epidemiol. Rev. 1990, 12, 179–191. [Google Scholar] [CrossRef] [PubMed]
- Johnston, M.D.; Brown, M.H. An investigation into the changed physiological state of Vibrio bacteria as a survival mechanism in response to cold temperatures and studies on their sensitivity to heating and freezing. J. Appl. Microbiol. 2002, 92, 1066–1077. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.D. Wound infections caused by Vibrio vulnificus and other marine bacteria. Epidemiol. Infect. 2005, 133, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Howard, R.J.; Bennett, N.T. Infections caused by halophilic marine Vibrio bacteria. Ann. Surg. 1993, 217, 525. [Google Scholar] [CrossRef] [PubMed]
- Fleming, L.E.; Broad, K.; Clement, A.; Dewailly, E.; Elmir, S.; Knap, A.; Pomponi, S.A.; Smith, S.; Solo Gabriele, H.; Walsh, P. Oceans and human health: Emerging public health risks in the marine environment. Mar. Pollut. Bull. 2006, 53, 545–560. [Google Scholar] [CrossRef] [PubMed]
- Feglo, P.K.; Sewurah, M. Characterization of highly virulent multidrug resistant Vibrio cholerae isolated from a large cholera outbreak in Ghana. BMC Res. Notes 2018, 11, 45. [Google Scholar] [CrossRef] [PubMed]
- Dupke, S.; Akinsinde, K.A.; Grunow, R.; Iwalokun, B.A.; Olukoya, D.K.; Oluwadun, A.; Thirumalaisamy, P.V.; Jacob, D. Characterization of Vibrio cholerae strains isolated from the Nigerian cholera outbreak in 2010. J. Clin. Microbiol. 2016, 54.10, 2618–2621. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chowdhury, G.; Bhadra, R.K.; Bag, S.; Pazhani, G.P.; Das, B.; Basu, P.; Nagamani, K.; Nandy, R.K.; Mukhopadhyay, A.K.; Ramamurthy, T. Rugose atypical Vibrio cholerae O1 El Tor responsible for 2009 cholera outbreak in India. J. Med.l Microbiol. 2016, 65, 1130–1136. [Google Scholar] [CrossRef]
- Kohout, F.A. Ground-water flow and the geothermal regime of the floridian plateau (1). GCAGS Trans. 1967, 17, 339–354. [Google Scholar]
- Bratton, J.F. The three scales of submarine groundwater flow and discharge across passive continental margins. J. Geol. 2010, 118, 565–575. [Google Scholar] [CrossRef]
- Kim, G.; Swarzenski, P.W. Submarine groundwater discharge (SGD) and associated nutrient fluxes to the coastal ocean. In Carbon and Nutrient Fluxes in Continental Margins: A Global Synthesis, Part III. Arising Issues and New Approaches; Liu, K.-K., Atkinson, L., Quinones, R., Talaue-McManus, L., Eds.; Springer: New York, NY, USA, 2010; pp. 529–538. [Google Scholar]
- Sugimoto, R.; Kitagawa, K.; Nishi, S.; Honda, H.; Yamada, M.; Kobayashi, S.; Shoji, J.; Ohsawa, S.; Taniguchi, M.; Tominaga, O. Phytoplankton primary productivity around submarine groundwater discharge in nearshore coasts. Mar. Ecol. Prog. Ser. 2017, 563, 25–33. [Google Scholar] [CrossRef]
- Schlüter, M.; Sauter, E.J.; Andersen, C.E.; Dahlgaard, H.; Dando, P.R. Spatial distribution and budget for submarine groundwater discharge in Eckernförde Bay (Western Baltic Sea). Limnol. Oceanogr. 2004, 49, 157–167. [Google Scholar] [CrossRef]
- Oehler, T.; Mogollón, J.M.; Moosdorf, N.; Winkler, A.; Kopf, A.; Pichler, T. Submarine groundwater discharge within a landslide scar at the French Mediterranean coast. Estuar. Coast. Shelf Sci. 2017, 198, 128–137. [Google Scholar] [CrossRef]
- Heiss, J.W.; Michael, H.A. Saltwater-freshwater mixing dynamics in a sandy beach aquifer over tidal, spring-neap, and seasonal cycles. Water Resour. Res. 2014, 50, 6747–6766. [Google Scholar] [CrossRef]
- Park, C.H.; Aral, M.M. Saltwater intrusion hydrodynamics in a tidal aquifer. J. Hydrol. Eng. 2008, 13, 863–872. [Google Scholar] [CrossRef]
- Righetti, C.; Gigliuto, A.; Chini, A.; Rossetto, R. Saltwater intrusion in a coastal contaminated aquifer density-dependent finite element model of flow and transport to assess remediation strategies and saltwater intrusion at a coastal gas plant site. In Proceedings of the 2nd International FEFLOW User Conference, Potsdam, Germany, 14–18 September 2009. [Google Scholar]
- Essink, G.H.O. Salt water intrusion in a three-dimensional groundwater system in the Netherlands: A numerical study. Transport Porous Med. 2001, 43.1, 137–158. [Google Scholar] [CrossRef]
- Giambastiani, B.M.; Antonellini, M.; Essink, G.H.O.; Stuurman, R.J. Saltwater intrusion in the unconfined coastal aquifer of Ravenna (Italy): A numerical model. J. Hydrol. 2007, 340, 91–104. [Google Scholar] [CrossRef]
- Bobba, A.G. Mathematical models for saltwater intrusion in coastal aquifers. Water Resour. Manag. 1993, 7, 3–37. [Google Scholar] [CrossRef]
- Ghassemi, F.; Jakeman, A.J.; Jacobson, G.; Howard, K.W.F. Simulation of seawater intrusion with 2D and 3D models: Nauru Island case study. Hydrogeol. J. 1996, 4.3, 4–22. [Google Scholar] [CrossRef]
- Diersch, H.J.G. FEFLOW-Finite Element Modeling of Flow, Mass and Heat Transport in Porous and Fractured Media; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Smith, A.J. Mixed convection and density-dependent seawater circulation in coastal aquifers. Water Resour. Res. 2004, 40. [Google Scholar] [CrossRef]
- Kolditz, O.; Ratke, R.; Diersch, H.J.G.; Zielke, W. Coupled groundwater flow and transport: 1. Verification of variable density flow and transport models. Adv. Water Resour. 1998, 21, 27–46. [Google Scholar] [CrossRef]
- Diersch, H.J.G. Modeling variable-density problems in 2D horizontally schematized aquifers using projected gravity. White Pap. 2004, 3, 5–12. [Google Scholar]
- Diersch, H.J.G. An efficient method for computing groundwater residence times. White Pap. 2002, 1, 141–150. [Google Scholar]
- Diersch, H.J.G. Consistent velocity approximation in the finite-element simulation of density dependent mass and heat transport processes. White Pap. 2002, 1, 141–150. [Google Scholar]
- Diersch, H.J.G. Nonlinear dispersion in density-dependent mass transport. White Pap. 2002, 1, 277–282. [Google Scholar]
- Walther, M.; Delfs, J.O.; Grundmann, J.; Kolditz, O.; Liedl, R. Saltwater intrusion modeling: Verification and application to an agricultural coastal arid region in Oman. J. Comput. Appl. Math. 2012, 236, 4798–4809. [Google Scholar] [CrossRef]
- Huo, Z.L.; Feng, S.Y.; Kang, S.Z.; Cen, S.J.; Ma, Y. Simulation of effects of agricultural activities on groundwater level by combining FEFLOW and GIS. New Zeal. J. Agr. Res. 2007, 50, 839–846. [Google Scholar] [CrossRef]
- Lu, J. Identification of forensic information from existing conventional site-investigation data. Introduction to Environ. Forensics 2015, 3, 149–164. [Google Scholar]
- Grant, S.A. Hydraulic properties, temperature effects. In Encyclopedia of Soils in the Environment; Hillel, D., Ed.; Elesevier: Amsterdam, The Netherlands, 2005; Volume 2, pp. 207–211. [Google Scholar]
- Kaleris, V.; Lagas, G.; Marczinek, S.; Piotrowski, J.A. Modelling submarine groundwater discharge: An example from the western Baltic Sea. J. Hydrol. 2002, 265, 76–99. [Google Scholar] [CrossRef]
- Bear, J.; Verruijt, A. Modeling Groundwater Flow and Pollution, 2nd ed.; Reidel, D., Ed.; Springer: Dordrecht, the Netherlands, 1987. [Google Scholar]
- Datta, B.; Vennalakanti, H.; Dhar, A. Modeling and control of saltwater intrusion in a coastal aquifer of Andhra Pradesh, India. J. Hydro-Environ. Res. 2009, 3, 148–159. [Google Scholar] [CrossRef]
- Robinson, C.; Gibbes, B.; Li, L. Driving mechanisms for groundwater flow and salt transport in a subterranean estuary. Geophys. Res. Lett. 2006, 33. [Google Scholar] [CrossRef]
- Narayan, K.A.; Schleeberger, C.; Bristow, K.L. Modelling seawater intrusion in the Burdekin Delta irrigation area, North Queensland, Australia. Agr. Water Manag. 2007, 89, 217–228. [Google Scholar] [CrossRef]
- Abarca, E.; Carrera, J.; Sanchez-Vila, X.; Dentz, M. Anisotropic dispersive Henry problem. Adv. Water Resour. 2007, 30, 913–926. [Google Scholar] [CrossRef]
- Doran, K.S.; Long, J.W.; Overbeck, J.R. A method for determining average beach slope and beach slope variability for US sandy coastlines. USGS 2015, 2015-1053, 5. [Google Scholar] [CrossRef]
- Boufadel, M.C. A mechanistic study of nonlinear solute transport in a groundwater-surface water system under steady state and transient hydraulic conditions. Water Resour. Res. 2000, 36, 2549–2565. [Google Scholar] [CrossRef]
- Robinson, C.; Li, L.; Prommer, H. Tide-induced recirculation across the aquifer-ocean interface. Water Resour. Res. 2007, 43. [Google Scholar] [CrossRef]
- Ghabayen, S.M.; McKee, M.; Kemblowski, M. Ionic and isotopic ratios for identification of salinity sources and missing data in the Gaza aquifer. J. Hydrol. 2006, 318, 360–373. [Google Scholar] [CrossRef]
- Davis, R.A. The Evolving Coast; Scientific American Library: New York, NY, USA, 1997. [Google Scholar]
- Houben, G.J.; Stoeckl, L.; Mariner, K.E.; Choudhury, A.S. The influence of heterogeneity on coastal groundwater flow-physical and numerical modeling of fringing reefs, dykes and structured conductivity fields. Adv. Water Resour. 2018, 113, 155–166. [Google Scholar] [CrossRef]
- Weinstein, Y.; Burnett, W.C.; Swarzenski, P.W.; Shalem, Y.; Yechieli, Y.; Herut, B. Role of aquifer heterogeneity in fresh groundwater discharge and seawater recycling: An example from the Carmel coast, Israel. J. Geophys. Res. 2007, 112. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Varying Parameter | Symbol | Unit | Value |
| Groundwater inflow | qX | m d−1 | 0.07; 0.17; 0.4; 0.8; 1.3 [12,51] |
| Hydr. conductivity | K | m d−1 | 8; 17; 34.4; 43; 85 [36,38,52,53,54] |
| Long. dispersivity | αL | m | 2.5; 5; 10; 15; 25; 35; 50; 75 [12,55] |
| Trans. dispersivity | αT | m | αL × 0.1 [12,55] |
| Coastal slope angle | β | ° | 0; 11.3; 28.1; 53.8; 69.7; 76.1 |
| Constant Parameter | Symbol | Unit | Value |
| Porosity aquifer | ϕS | – | 0.3 [56,57] |
| Salinity groundwater | CF | mg L−1 | 100 [44] |
| Salinity seawater | CS | mg L−1 | 35000 [44] |
| Density groundwater | ρF | kg m−3 | 1000 [58] |
| Density seawater | ρS | kg m−3 | 1026 [58] |
| Density ratio | d | – | 0.026 |
| Molecular diffusion coefficient | DM | m2 s−1 | 1 × 10−9 [58] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vollberg, F.; Walther, M.; Gärdes, A.; Moosdorf, N. Modeling the Potential of Submarine Groundwater Discharge to Facilitate Growth of Vibrio cholerae Bacteria. Hydrology 2019, 6, 39. https://doi.org/10.3390/hydrology6020039
Vollberg F, Walther M, Gärdes A, Moosdorf N. Modeling the Potential of Submarine Groundwater Discharge to Facilitate Growth of Vibrio cholerae Bacteria. Hydrology. 2019; 6(2):39. https://doi.org/10.3390/hydrology6020039
Chicago/Turabian StyleVollberg, Felix, Marc Walther, Astrid Gärdes, and Nils Moosdorf. 2019. "Modeling the Potential of Submarine Groundwater Discharge to Facilitate Growth of Vibrio cholerae Bacteria" Hydrology 6, no. 2: 39. https://doi.org/10.3390/hydrology6020039
APA StyleVollberg, F., Walther, M., Gärdes, A., & Moosdorf, N. (2019). Modeling the Potential of Submarine Groundwater Discharge to Facilitate Growth of Vibrio cholerae Bacteria. Hydrology, 6(2), 39. https://doi.org/10.3390/hydrology6020039