

Recreating the Trabecular Outflow Tissue on Implantable, Micropatterned, Ultrathin, Porous Polycaprolactone Scaffolds
 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
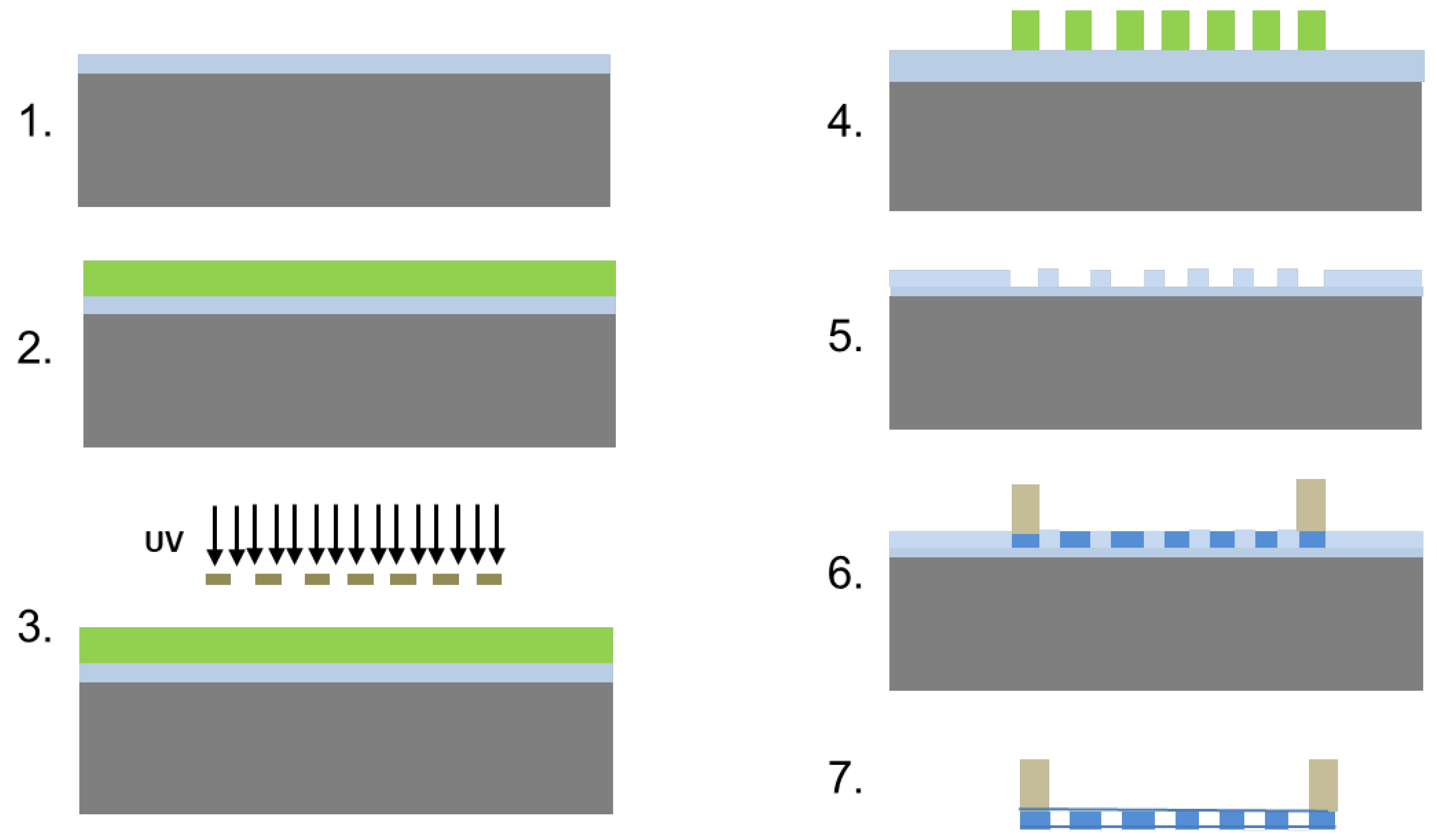
2.1. Microfabrication of Patterned, Ultrathin, Porous Polycaprolactone (PCL) Scaffolds
2.2. Human Trabecular Meshwork (HTM) Cell Culture
2.3. Culture of HTM Cells on PCL Scaffolds
2.4. Scanning Electron Microscopy (SEM)
2.5. MTT Assay
2.6. Optical Imaging
2.7. Cytoskeleton Staining and Immunocytochemistry Followed by Confocal Imaging
2.8. Quantitative Polymerase Chain Reaction (qPCR) Analysis
2.9. Statistical Analysis
3. Results
3.1. Micropatterned, Ultrathin, Porous PCL Scaffolds Support Better Trabecular Meshwork Cell Growth Than 2D Glass Coverslips
3.2. Gelatin-Coated, Micropatterned PCL Scaffolds Support Trabecular Meshwork Cell Morphology and Cytoskeletal Protein Expression
3.3. Gelatin-Coated PCL Scaffolds Support Higher Gene Expression of ECM Molecules and α-SMA in HTM Cells Than 2D Glass Coverslips
3.4. Gelatin-Coated Grid-Patterned PCL Scaffolds Support Higher Expression of ECM Proteins and HTM Markers Than Hexagon-Patterned Scaffolds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Taylor, A.W. Primary Open-Angle Glaucoma: A transforming growth factor-β pathway-mediated disease. Am. J. Pathol. 2012, 180, 2201–2204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapetanakis, V.V.; Chan, M.P.Y.; Foster, P.J.; Cook, D.G.; Owen, C.G.; Rudnicka, A.R. Global variations and time trends in the prevalence of primary open angle glaucoma (POAG): A systematic review and meta-analysis. Br. J. Ophthalmol. 2016, 100, 86–93. [Google Scholar] [CrossRef] [Green Version]
- Allison, K.; Patel, D.; Alabi, O. Epidemiology of Glaucoma: The Past, Present, and Predictions for the Future. Cureus 2020, 12, e11686. [Google Scholar] [CrossRef] [PubMed]
- Ferrer, E. Trabecular meshwork as a new target for the treatment of glaucoma. Drug News Perspect. 2006, 19, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, P.L.; Rasmussen, C.A. Advances in Glaucoma Treatment and Management: Outflow Drugs. Investig. Ophthalmol. Vis. Sci. 2012, 53, 2495–2500. [Google Scholar] [CrossRef] [Green Version]
- O’Callaghan, J.; Cassidy, P.S.; Humphries, P. Open-angle glaucoma: Therapeutically targeting the extracellular matrix of the conventional outflow pathway. Expert Opin. Ther. Targets 2017, 21, 1037–1050. [Google Scholar] [CrossRef] [Green Version]
- Castro, A.; Du, Y. Trabecular Meshwork Regeneration—A Potential Treatment for Glaucoma. Curr. Ophthalmol. Rep. 2019, 7, 80–88. [Google Scholar] [CrossRef] [PubMed]
- Coulon, S.J.; Schuman, J.S.; Du, Y.; Bahrani Fard, M.R.; Ethier, C.R.; Stamer, W.D. A novel glaucoma approach: Stem cell regeneration of the trabecular meshwork. Prog. Retin. Eye Res. 2022, 90, 101063. [Google Scholar] [CrossRef]
- Abu-Hassan, D.W.; Acott, T.S.; Kelley, M.J. The Trabecular Meshwork: A Basic Review of Form and Function. J. Ocul. Biol. 2014, 2, 9. [Google Scholar] [CrossRef]
- Weinreb, R.N.; Aung, T.; Medeiros, F.A. The Pathophysiology and Treatment of Glaucoma: A Review. JAMA 2014, 311, 1901–1911. [Google Scholar] [CrossRef] [Green Version]
- Kelley, M.J.; Rose, A.Y.; Keller, K.E.; Hessle, H.; Samples, J.R.; Acott, T.S. Stem cells in the trabecular meshwork: Present and future promises. Exp. Eye Res. 2009, 88, 747–751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, W.; Gramlich, O.W.; Laboissonniere, L.; Jain, A.; Sheffield, V.C.; Trimarchi, J.M.; Tucker, B.A.; Kuehn, M.H. Transplantation of iPSC-derived TM cells rescues glaucoma phenotypes in vivo. Proc. Natl. Acad. Sci. USA 2016, 113, E3492–E3500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, W.; Jain, A.; Gramlich, O.W.; Tucker, B.A.; Sheffield, V.C.; Kuehn, M.H. Restoration of Aqueous Humor Outflow Following Transplantation of iPSC-Derived Trabecular Meshwork Cells in a Transgenic Mouse Model of Glaucoma. Investig. Ophthalmol. Vis. Sci. 2017, 58, 2054–2062. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, S.; Kumar, A.; Tian, S.; Taher, E.E.; Yang, E.; Kinchington, P.R.; Xia, X.; Du, Y. Stem cell transplantation rescued a primary open-angle glaucoma mouse model. eLife 2021, 10, e63677. [Google Scholar] [CrossRef] [PubMed]
- Mallick, S.; Sharma, M.; Kumar, A.; Du, Y. Cell-Based Therapies for Trabecular Meshwork Regeneration to Treat Glaucoma. Biomolecules 2021, 11, 1258. [Google Scholar] [CrossRef]
- Acott, T.S.; Kelley, M.J. Extracellular matrix in the trabecular meshwork. Exp. Eye Res. 2008, 86, 543–561. [Google Scholar] [CrossRef] [Green Version]
- Alvarado, J.A.; Wood, I.; Polansky, J.R. Human trabecular cells. II. Growth pattern and ultrastructural characteristics. Investig. Ophthalmol. Vis. Sci. 1982, 23, 464–478. [Google Scholar]
- Gasiorowski, J.Z.; Russell, P. Biological properties of trabecular meshwork cells. Exp. Eye Res. 2009, 88, 671–675. [Google Scholar] [CrossRef] [Green Version]
- Fautsch, M.P.; Howell, K.G.; Vrabel, A.M.; Charlesworth, M.C.; Muddiman, D.C.; Johnson, D.H. Primary trabecular meshwork cells incubated in human aqueous humor differ from cells incubated in serum supplements. Investig. Ophthalmol. Vis. Sci. 2005, 46, 2848–2856. [Google Scholar] [CrossRef] [Green Version]
- Goffin, J.M.; Pittet, P.; Csucs, G.; Lussi, J.W.; Meister, J.-J.; Hinz, B. Focal adhesion size controls tension-dependent recruitment of alpha-smooth muscle actin to stress fibers. J. Cell Biol. 2006, 172, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Osmond, M.J.; Krebs, M.D.; Pantcheva, M.B. Human trabecular meshwork cell behavior is influenced by collagen scaffold pore architecture and glycosaminoglycan composition. Biotechnol. Bioeng. 2020, 117, 3150–3159. [Google Scholar] [CrossRef] [PubMed]
- Soto, D.; Comes, N.; Ferrer, E.; Morales, M.; Escalada, A.; Palés, J.; Solsona, C.; Gual, A.; Gasull, X. Modulation of aqueous humor outflow by ionic mechanisms involved in trabecular meshwork cell volume regulation. Investig. Ophthalmol. Vis. Sci. 2004, 45, 3650–3661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torrejon, K.Y.; Pu, D.; Bergkvist, M.; Danias, J.; Sharfstein, S.T.; Xie, Y. Recreating a human trabecular meshwork outflow system on microfabricated porous structures. Biotechnol. Bioeng. 2013, 110, 3205–3218. [Google Scholar] [CrossRef] [PubMed]
- Dautriche, C.N.; Tian, Y.; Xie, Y.; Sharfstein, S.T. A Closer Look at Schlemm’s Canal Cell Physiology: Implications for Biomimetics. J. Funct. Biomater. 2015, 6, 963–985. [Google Scholar] [CrossRef] [Green Version]
- Torrejon, K.Y.; Papke, E.L.; Halman, J.R.; Bergkvist, M.; Danias, J.; Sharfstein, S.T.; Xie, Y. TGFβ2-induced outflow alterations in a bioengineered trabecular meshwork are offset by a rho-associated kinase inhibitor. Sci. Rep. 2016, 6, 38319. [Google Scholar] [CrossRef] [Green Version]
- Tian, Y.I.; Zhang, X.; Torrejon, K.; Danias, J.; Gindina, S.; Nayyar, A.; Du, Y.; Xie, Y. A bioengineering approach to Schlemm’s canal-like stem cell differentiation for in vitro glaucoma drug screening. Acta Biomater. 2020, 105, 203–213. [Google Scholar] [CrossRef]
- Tian, Y.I.; Zhang, X.; Torrejon, K.; Danias, J.; Du, Y.; Xie, Y. A Biomimetic, Stem Cell-Derived In Vitro Ocular Outflow Model. Adv. Biosyst. 2020, 4, 2000004. [Google Scholar] [CrossRef]
- Williams, J.M.; Adewunmi, A.; Schek, R.M.; Flanagan, C.L.; Krebsbach, P.H.; Feinberg, S.E.; Hollister, S.J.; Das, S. Bone tissue engineering using polycaprolactone scaffolds fabricated via selective laser sintering. Biomaterials 2005, 26, 4817–4827. [Google Scholar] [CrossRef]
- Eshraghi, S.; Das, S. Mechanical and microstructural properties of polycaprolactone scaffolds with one-dimensional, two-dimensional, and three-dimensional orthogonally oriented porous architectures produced by selective laser sintering. Acta Biomater. 2010, 6, 2467–2476. [Google Scholar] [CrossRef] [Green Version]
- Zein, I.; Hutmacher, D.W.; Tan, K.C.; Teoh, S.H. Fused deposition modeling of novel scaffold architectures for tissue engineering applications. Biomaterials 2002, 23, 1169–1185. [Google Scholar] [CrossRef]
- Shahriari, D.; Koffler, J.Y.; Tuszynski, M.H.; Campana, W.M.; Sakamoto, J.S. Hierarchically Ordered Porous and High-Volume Polycaprolactone Microchannel Scaffolds Enhanced Axon Growth in Transected Spinal Cords. Tissue Eng. Part A 2017, 23, 415–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, S.; Lee, G.Y.; Wong, J.Y.; Desai, T.A. Development and characterization of a porous micro-patterned scaffold for vascular tissue engineering applications. Biomaterials 2006, 27, 4775–4782. [Google Scholar] [CrossRef] [PubMed]
- Aldemir Dikici, B.; Malayeri, A.; Sherborne, C.; Dikici, S.; Paterson, T.; Dew, L.; Hatton, P.; Ortega Asencio, I.; MacNeil, S.; Langford, C.; et al. Thiolene- and Polycaprolactone Methacrylate-Based Polymerized High Internal Phase Emulsion (PolyHIPE) Scaffolds for Tissue Engineering. Biomacromolecules 2022, 23, 720–730. [Google Scholar] [CrossRef] [PubMed]
- Aldemir Dikici, B.; Sherborne, C.; Reilly, G.C.; Claeyssens, F. Emulsion templated scaffolds manufactured from photocurable polycaprolactone. Polymer 2019, 175, 243–254. [Google Scholar] [CrossRef]
- Dong, L.; Wang, S.-J.; Zhao, X.-R.; Zhu, Y.-F.; Yu, J.-K. 3D-Printed Poly(ε-caprolactone) Scaffold Integrated with Cell-laden Chitosan Hydrogels for Bone Tissue Engineering. Sci. Rep. 2017, 7, 13412. [Google Scholar] [CrossRef] [Green Version]
- Hollister, S.J. Porous scaffold design for tissue engineering. Nat. Mater. 2005, 4, 518–524. [Google Scholar] [CrossRef] [PubMed]
- Salerno, A.; Guarnieri, D.; Iannone, M.; Zeppetelli, S.; Di Maio, E.; Iannace, S.; Netti, P.A. Engineered mu-bimodal poly(epsilon-caprolactone) porous scaffold for enhanced hMSC colonization and proliferation. Acta Biomater. 2009, 5, 1082–1093. [Google Scholar] [CrossRef]
- Yoshimoto, H.; Shin, Y.M.; Terai, H.; Vacanti, J.P. A biodegradable nanofiber scaffold by electrospinning and its potential for bone tissue engineering. Biomaterials 2003, 24, 2077–2082. [Google Scholar] [CrossRef]
- Beardslee, L.A.; Stolwijk, J.; Khaladj, D.A.; Trebak, M.; Halman, J.; Torrejon, K.Y.; Niamsiri, N.; Bergkvist, M. A sacrificial process for fabrication of biodegradable polymer membranes with submicron thickness. J. Biomed. Mater. Res. Part B Appl. Biomater. 2016, 104, 1192–1201. [Google Scholar] [CrossRef]
- Sodha, S.; Wall, K.; Redenti, S.; Klassen, H.; Young, M.J.; Tao, S.L. Microfabrication of a Three-Dimensional Polycaprolactone Thin-Film Scaffold for Retinal Progenitor Cell Encapsulation. J. Biomater. Sci. Polym. Ed. 2011, 22, 443–456. [Google Scholar] [CrossRef]
- Kweon, H.; Yoo, M.K.; Park, I.K.; Kim, T.H.; Lee, H.C.; Lee, H.-S.; Oh, J.-S.; Akaike, T.; Cho, C.S. A novel degradable polycaprolactone networks for tissue engineering. Biomaterials 2003, 24, 801–808. [Google Scholar] [CrossRef] [PubMed]
- De Kater, A.W.; Shahsafaei, A.; Epstein, D.L. Localization of smooth muscle and nonmuscle actin isoforms in the human aqueous outflow pathway. Investig. Ophthalmol. Vis. Sci. 1992, 33, 424–429. [Google Scholar]
- Yun, A.J.; Murphy, C.G.; Polansky, J.R.; Newsome, D.A.; Alvarado, J.A. Proteins secreted by human trabecular cells. Glucocorticoid and other effects. Investig. Ophthalmol. Vis. Sci. 1989, 30, 2012–2022. [Google Scholar]
- Fuchshofer, R.; Tamm, E.R. Modulation of extracellular matrix turnover in the trabecular meshwork. Exp. Eye Res. 2009, 88, 683–688. [Google Scholar] [CrossRef]
- Goel, M.; Picciani, R.G.; Lee, R.K.; Bhattacharya, S.K. Aqueous humor dynamics: A review. Open Ophthalmol. J. 2010, 4, 52–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, H.; Tripathi, R.C.; Tripathi, B.J. Morphology of the aqueous outflow pathway. Microsc. Res. Technol. 1996, 33, 336–367. [Google Scholar] [CrossRef]
- Cherng, S.; Young, J.; Ma, H. Alpha-Smooth Muscle Actin (α-SMA). J. Am. Sci. 2008, 4, 7–9. [Google Scholar]
- Hinz, B.; Celetta, G.; Tomasek, J.J.; Gabbiani, G.; Chaponnier, C. Alpha-Smooth Muscle Actin Expression Upregulates Fibroblast Contractile Activity. Mol. Biol. Cell 2001, 12, 2730–2741. [Google Scholar] [CrossRef] [Green Version]
- Horwitz, J. Alpha-crystallin can function as a molecular chaperone. Proc. Natl. Acad. Sci. USA 1992, 89, 10449–10453. [Google Scholar] [CrossRef] [Green Version]
- Tomarev, S.I.; Wistow, G.; Raymond, V.; Dubois, S.; Malyukova, I. Gene Expression Profile of the Human Trabecular Meshwork: NEIBank Sequence Tag Analysis. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2588–2596. [Google Scholar] [CrossRef] [Green Version]
- Nazir, S.; Mukhtar, M.; Shahnawaz, M.; Farooqi, S.; Fatima, N.; Mehmood, R.; Sheikh, N. A novel single nucleotide polymorphism in exon 3 of MYOC enhances the risk of glaucoma. PLoS ONE 2018, 13, e0195157. [Google Scholar] [CrossRef] [PubMed]
- Torrejon, K.Y.; Papke, E.L.; Halman, J.R.; Stolwijk, J.; Dautriche, C.N.; Bergkvist, M.; Danias, J.; Sharfstein, S.T.; Xie, Y. Bioengineered glaucomatous 3D human trabecular meshwork as an in vitro disease model. Biotechnol. Bioeng. 2016, 113, 1357–1368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dautriche, C.N.; Xie, Y.; Sharfstein, S.T. Walking through trabecular meshwork biology: Toward engineering design of outflow physiology. Biotechnol. Adv. 2014, 32, 971–983. [Google Scholar] [CrossRef]
- Ryskamp, D.A.; Frye, A.M.; Phuong, T.T.T.; Yarishkin, O.; Jo, A.O.; Xu, Y.; Lakk, M.; Iuso, A.; Redmon, S.N.; Ambati, B.; et al. TRPV4 regulates calcium homeostasis, cytoskeletal remodeling, conventional outflow and intraocular pressure in the mammalian eye. Sci. Rep. 2016, 6, 30583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastia, E.; Toris, C.B.; Brambilla, S.; Galli, C.; Almirante, N.; Bergamini, M.V.W.; Masini, E.; Sgambellone, S.; Unser, A.M.; Ahmed, F.; et al. NCX 677, a Novel Nitric Oxide Donor, Lowers Introcular Pressure in RBBITS, Dogs and Non-Human Primates and Enhances TGFβ2-Induced Outflow in HTM/HSC Constructs. Investig. Ophthalmol. Vis. Sci. 2021, 62, 17. [Google Scholar] [CrossRef]
- Kim, J.; Kudisch, M.; da Silva, N.R.K.; Asada, H.; Aya-Shibuya, E.; Bloomer, M.M.; Mudumba, S.; Bhisitkul, R.B.; Desai, T.A. Long-term intraocular pressure reduction with intracameral polycaprolactone glaucoma devices that deliver a novel anti-glaucoma agent. J. Control. Release 2018, 269, 45–51. [Google Scholar] [CrossRef]
- Kim, J.; Kudisch, M.; Mudumba, S.; Asada, H.; Aya-Shibuya, E.; Bhisitkul, R.B.; Desai, T.A. Biocompatibility and Pharmacokinetic Analysis of an Intracameral Polycaprolactone Drug Delivery Implant for Glaucoma. Investig. Ophthalmol. Vis. Sci. 2016, 57, 4341–4346. [Google Scholar] [CrossRef] [Green Version]
- Lance, K.D.; Good, S.D.; Mendes, T.S.; Ishikiriyama, M.; Chew, P.; Estes, L.S.; Yamada, K.; Mudumba, S.; Bhisitkul, R.B.; Desai, T.A. In Vitro and In Vivo Sustained Zero-Order Delivery of Rapamycin (Sirolimus) from a Biodegradable Intraocular Device. Investig. Ophthalmol. Vis. Sci. 2015, 56, 7331–7337. [Google Scholar] [CrossRef] [Green Version]
- Bernards, D.A.; Bhisitkul, R.B.; Wynn, P.; Steedman, M.R.; Lee, O.-T.; Wong, F.; Thoongsuwan, S.; Desai, T.A. Ocular biocompatibility and structural integrity of micro- and nanostructured poly(caprolactone) films. J. Ocul. Pharmacol. Ther. Off. J. Assoc. Ocul. Pharmacol. Ther. 2013, 29, 249–257. [Google Scholar] [CrossRef] [Green Version]
- Xiang, P.; Wu, K.-C.; Zhu, Y.; Xiang, L.; Li, C.; Chen, D.-L.; Chen, F.; Xu, G.; Wang, A.; Li, M.; et al. A novel Bruch’s membrane-mimetic electrospun substrate scaffold for human retinal pigment epithelium cells. Biomaterials 2014, 35, 9777–9788. [Google Scholar] [CrossRef] [Green Version]
- Kashanian, S.; Harding, F.; Irani, Y.; Klebe, S.; Marshall, K.; Loni, A.; Canham, L.; Fan, D.; Williams, K.A.; Voelcker, N.H.; et al. Evaluation of mesoporous silicon/polycaprolactone composites as ophthalmic implants. Acta Biomater. 2010, 6, 3566–3572. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse |
|---|---|---|
| α-SMA | 5′-GGA TTA AGT TCA TAA GAT TCC ATG CT-3′ | 5′-TGT TAT GGA GAG TGG CAG AAA G-3′ |
| Fibronectin | 5′-GTC CTT GTG TCC TGA TCG TTG-3′ | 5′-AGG CTG GAT GAT GGT AGA TTG-3′ |
| Collagen IV | 5′-CCT TTG TGC CAT TGC ATC C-3′ | 5′-GAA CAA AAG GGA CAA GAG GAC-3′ |
| GAPDH | 5′-TGT AGT TGA GGT CAA TGA AGG D-3′ | 5′-ACA TCG CTC AGA CAC CAT G-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Beardslee, L.A.; Halman, J.R.; Unser, A.M.; Xie, Y.; Danias, J.; Bergkvist, M.; Sharfstein, S.T.; Torrejon, K.Y. Recreating the Trabecular Outflow Tissue on Implantable, Micropatterned, Ultrathin, Porous Polycaprolactone Scaffolds. Bioengineering 2023, 10, 679. https://doi.org/10.3390/bioengineering10060679
Beardslee LA, Halman JR, Unser AM, Xie Y, Danias J, Bergkvist M, Sharfstein ST, Torrejon KY. Recreating the Trabecular Outflow Tissue on Implantable, Micropatterned, Ultrathin, Porous Polycaprolactone Scaffolds. Bioengineering. 2023; 10(6):679. https://doi.org/10.3390/bioengineering10060679
Chicago/Turabian StyleBeardslee, Luke A., Justin R. Halman, Andrea M. Unser, Yubing Xie, John Danias, Magnus Bergkvist, Susan T. Sharfstein, and Karen Y. Torrejon. 2023. "Recreating the Trabecular Outflow Tissue on Implantable, Micropatterned, Ultrathin, Porous Polycaprolactone Scaffolds" Bioengineering 10, no. 6: 679. https://doi.org/10.3390/bioengineering10060679