

Human Periodontal Ligament Stem Cells (hPDLSCs) Spontaneously Differentiate into Myofibroblasts to Repair Diabetic Wounds



Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Isolation and Cultivation of hPDLSCs
2.2. MTT Assays Were Used to Detect the Inhibitory Effect of AGEs on the Proliferation of Periodontal Ligament Stem Cells
2.3. Real-Time Quantitative Fluorescent Polymerase Chain Reaction (qRT-PCR) Was Used to Detect the Expressions of α-SMA, COL1, and Vimentin
2.4. Western Blot Analysis Was Used to Detect Protein Expression
2.5. STZ-Induced Diabetic Whole-Skin Defect Experiment to Verify Wound Healing
2.6. Immunohistochemistry (IHC) Detects α-SMA and CD31 Expression
2.7. Immunofluorescence (IF) Assay
2.8. Fluorescence In Situ Hybridization (FISH)
2.9. Statistical Analysis
3. Results
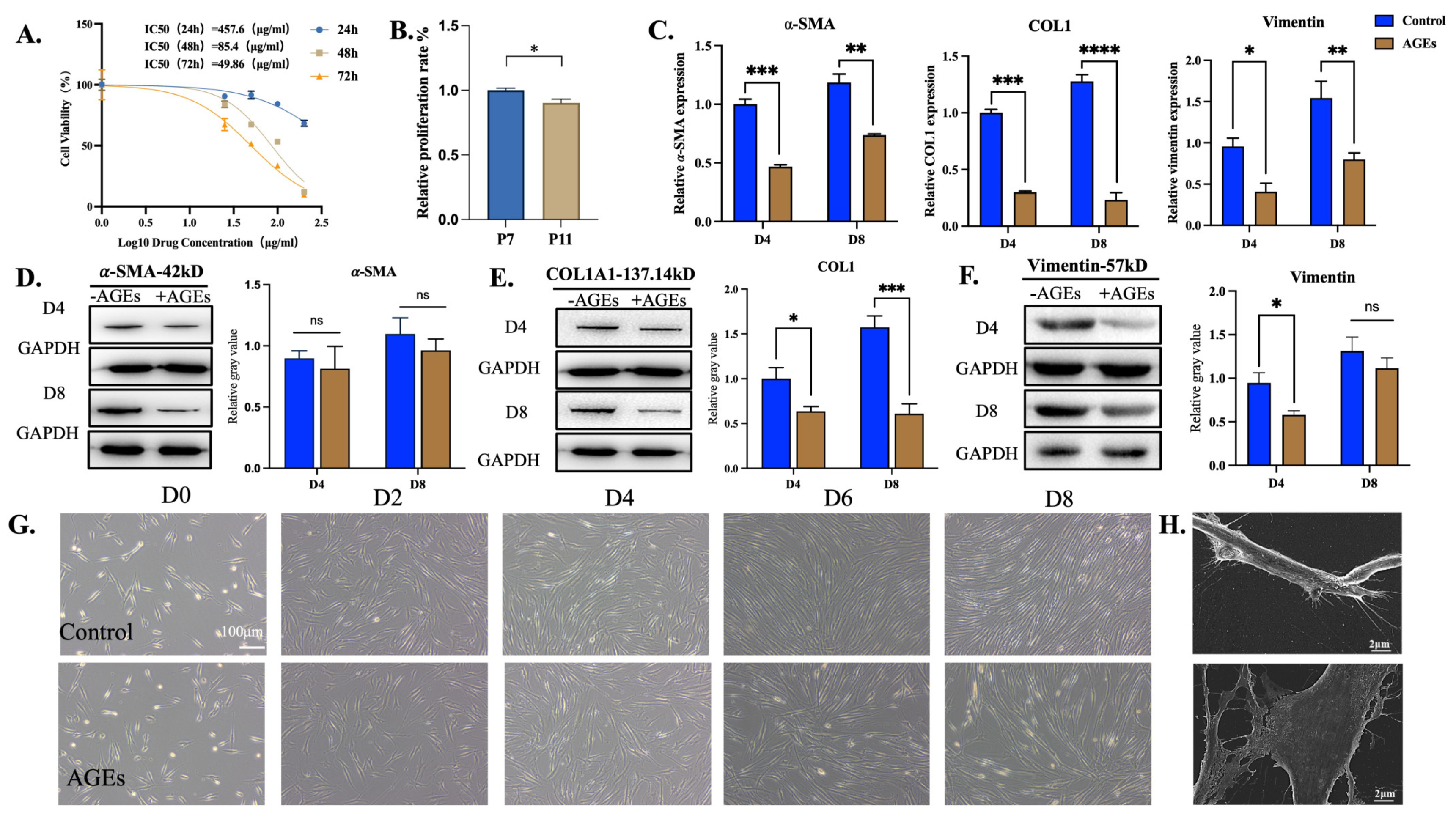
3.1. Spontaneous Differentiation of hPDLSCs into Myofibroblasts Was Associated with Conditioned Medium
3.2. hPDLSCs Resist the Damage Caused by AGEs
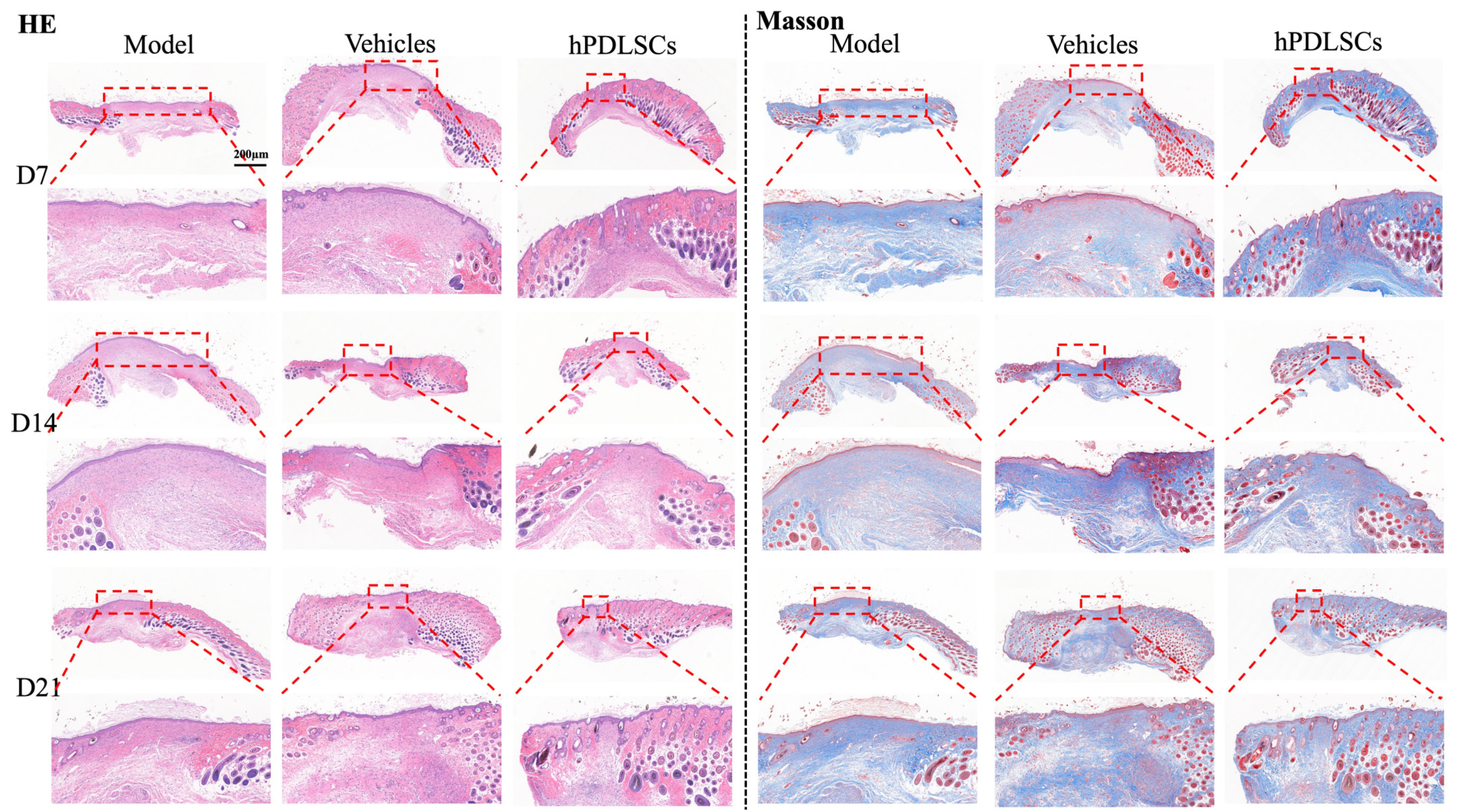
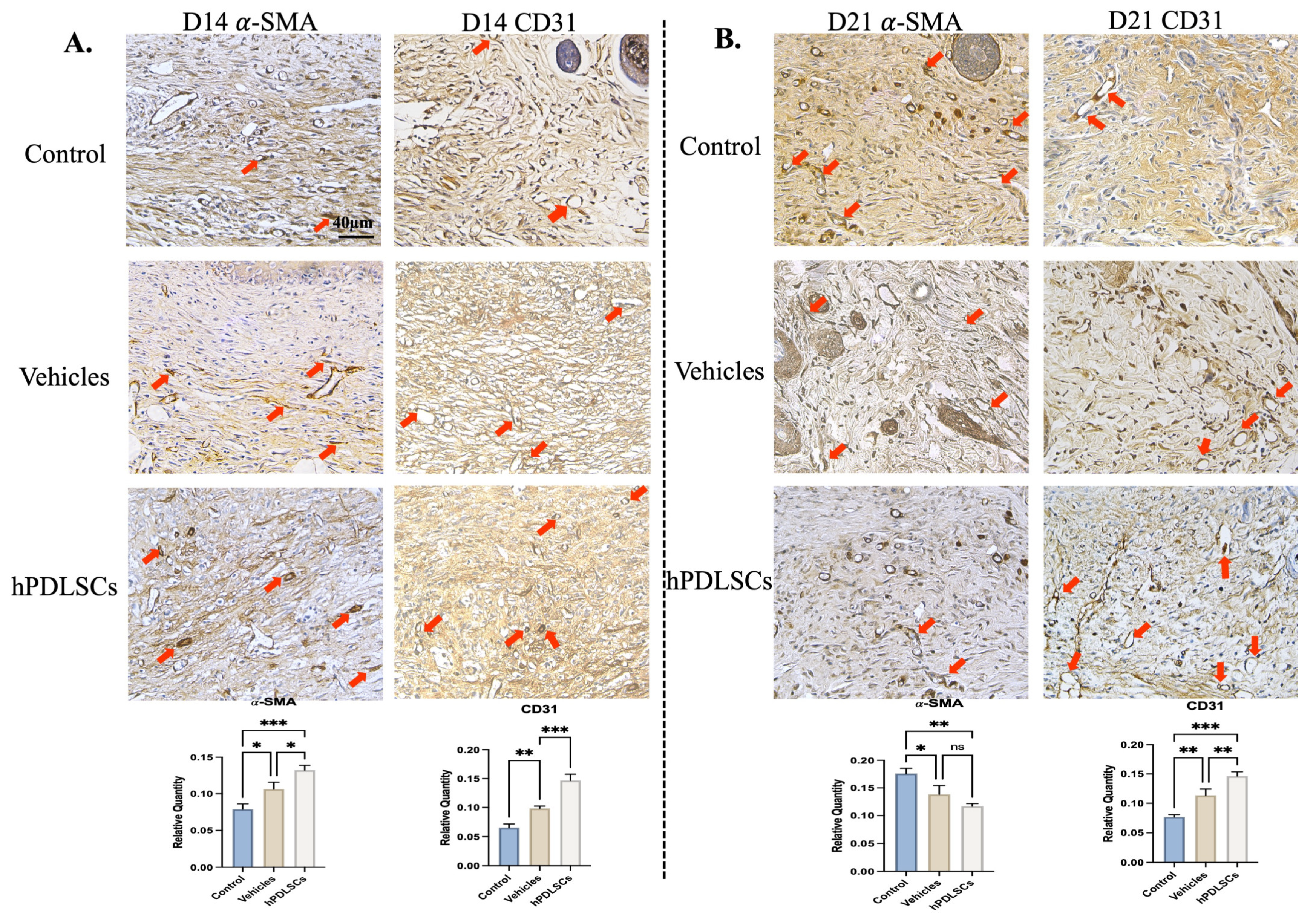
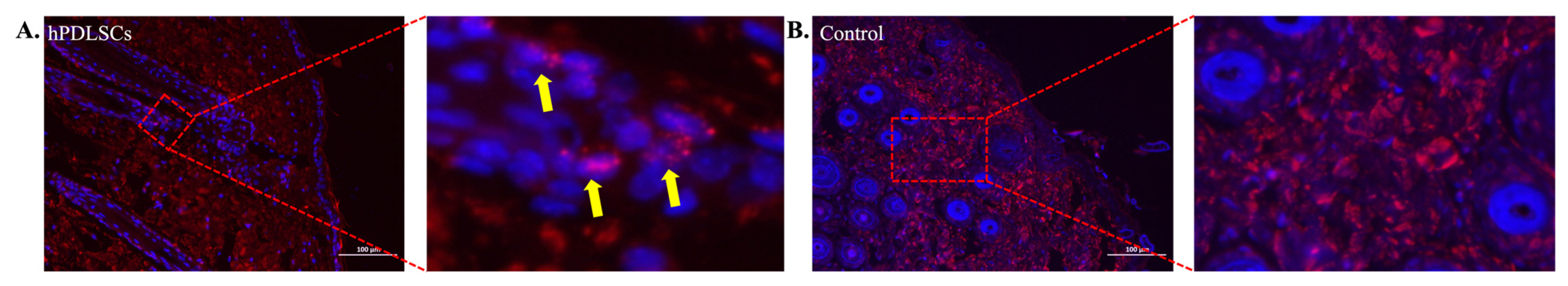
3.3. hPDLSCs Repaired Full-Thickness Skin Defects in Diabetic Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Asadipooya, K.; Uy, E.M. Advanced glycation end products (ages), receptor for ages, diabetes, and bone: Review of the literature. J. Endocr. Soc. 2019, 3, 1799–1818. [Google Scholar] [CrossRef] [PubMed]
- Khalid, M.; Petroianu, G.; Adem, A. Advanced glycation end products and diabetes mellitus: Mechanisms and perspectives. Biomolecules 2022, 12, 542. [Google Scholar] [CrossRef] [PubMed]
- Cole, J.B.; Florez, J.C. Genetics of diabetes mellitus and diabetes complications. Nat. Rev. Nephrol. 2020, 16, 377–390. [Google Scholar] [CrossRef] [PubMed]
- Cano Sanchez, M.; Lancel, S.; Boulanger, E.; Neviere, R. Targeting oxidative stress and mitochondrial dysfunction in the treatment of impaired wound healing: A systematic review. Antioxidants 2018, 7, 98. [Google Scholar] [CrossRef] [PubMed]
- Hinz, B. Formation and function of the myofibroblast during tissue repair. J. Investig. Dermatol. 2007, 127, 526–537. [Google Scholar] [CrossRef]
- Teng, Y.; Zou, M.; Zhou, X.; Wu, J.; Liu, S.; Yuan, Z.; Jia, Y.; Zhang, K.; Li, X.; Ye, J.; et al. Novel prospects for scarless wound healing: The roles of myofibroblasts and adipocytes. J. Cell. Mol. Med. 2022, 26, 5113–5121. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Mack, J.A.; Hascall, V.C.; Maytin, E.V. Transforming growth factor-β receptor–mediated, p38 mitogen-activated protein kinase–dependent signaling drives enhanced myofibroblast differentiation during skin wound healing in mice lacking hyaluronan synthases 1 and 3. Am. J. Pathol. 2022, 192, 1683–1698. [Google Scholar] [CrossRef]
- Jagadeeshaprasad, M.G.; Govindappa, P.K.; Nelson, A.M.; Noble, M.D.; Elfar, J.C. 4-Aminopyridine induces nerve growth factor to improve skin wound healing and tissue regeneration. Biomedicines 2023, 10, 1649. [Google Scholar] [CrossRef]
- Schuster, R.; Younesi, F.; Ezzo, M.; Hinz, B. The Role of myofibroblasts in physiological and pathological tissue repair. Cold Spring Harb. Perspect. Biol. 2022, 15, a041231. [Google Scholar] [CrossRef]
- Desmoulière, A.; Chaponnier, C.; Gabbiani, G. Perspective article: Tissue repair, contraction, and the myofibroblast. Wound Repair Regen. 2005, 13, 7–12. [Google Scholar] [CrossRef]
- Ghorbani, A.; Mojarrad, M.; Hatami, A.; Hoseini, S.; Ghazavi, H.; Hosseini, A. Effects of streptozotocin-induced diabetes on proliferation and differentiation abilities of mesenchymal stem cells derived from subcutaneous and visceral adipose tissues. Exp. Clin. Endocrinol. Diabetes 2017, 125, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Jumabay, M.; Moon, J.H.; Yeerna, H.; Boström, K.I. Effect of diabetes mellitus on adipocyte-derived stem cells in rat. J. Cell. Physiol. 2015, 230, 2821–2828. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-C.; Kim, J.-M.; Jung, I.-H.; Kim, J.C.; Choi, S.-H.; Cho, K.-S.; Kim, C.-S. Isolation and characterization of human periodontal ligament (PDL) stem cells (PDLSCs) from the inflamed PDL tissue: In vitro and in vivo evaluations. J. Clin. Periodontol. 2011, 38, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Lei, T.; Wang, J.; Liu, Y.; Chen, P.; Zhang, Z.; Zhang, X.; Guo, W.; Wang, X.; Li, Q.; Du, H. Proteomic profile of human stem cells from dental pulp and periodontal ligament. J. Proteom. 2021, 245, 104280. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhao, Z.; Ruan, J.; Weir, M.D.; Ma, T.; Ren, K.; Schneider, A.; Oates, T.W.; Li, A.; Zhao, L.; et al. Stem cells in the periodontal ligament differentiated into osteogenic, fibrogenic and cementogenic lineages for the regeneration of the periodontal complex. J. Dent. 2020, 92, 103259. [Google Scholar] [CrossRef] [PubMed]
- Onizuka, S.; Iwata, T. Application of periodontal ligament-derived multipotent mesenchymal stromal cell sheets for periodontal regeneration. Int. J. Mol. Sci. 2019, 20, 2796. [Google Scholar] [CrossRef] [PubMed]
- Tassi, S.A.; Sergio, N.Z.; Misawa, M.Y.O.; Villar, C.C. Efficacy of stem cells on periodontal regeneration: Systematic review of pre-clinical studies. J. Periodontal Res. 2017, 52, 793–812. [Google Scholar] [CrossRef] [PubMed]
- Iwata, T.; Yamato, M.; Zhang, Z.; Mukobata, S.; Washio, K.; Ando, T.; Feijen, J.; Okano, T.; Ishikawa, I. Validation of human periodontal ligament-derived cells as a reliable source for cytotherapeutic use. J. Clin. Periodontol. 2010, 37, 1088–1099. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Xie, J.; Wang, C.; Zhong, D.; Xie, L.; Fang, H. Immunomodulatory properties of stem cells in periodontitis: Current status and future prospective. Stem Cells Int. 2020, 2020, 9836518. [Google Scholar] [CrossRef]
- Menicanin, D.; Mrozik, K.M.; Wada, N.; Marino, V.; Shi, S.; Bartold, P.M.; Gronthos, S. Periodontal-Ligament-Derived stem cells exhibit the capacity for long-term survival, self-renewal, and regeneration of multiple tissue types in vivo. Stem Cells Dev. 2014, 23, 1001–1011. [Google Scholar] [CrossRef]
- Guo, J.; Weng, J.; Rong, Q.; Zhang, X.; Zhu, S.; Huang, D.; Li, X.; Chen, S.L. Investigation of multipotent postnatal stem cells from human maxillary sinus membrane. Sci. Rep. 2015, 5, srep11660. [Google Scholar] [CrossRef] [PubMed]
- Vandana, K.L.; Shalini, H.S. Direct application of autologous periodontal ligament stem cell niche in treatment of periodontal osseous defects: A randomized controlled trial. J. Indian Soc. Periodontol. 2018, 22, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Iwata, T.; Yamato, M.; Washio, K.; Yoshida, T.; Tsumanuma, Y.; Yamada, A.; Onizuka, S.; Izumi, Y.; Ando, T.; Okano, T.; et al. Periodontal regeneration with autologous periodontal ligament-derived cell sheets—A safety and efficacy study in ten patients. Regen. Ther. 2018, 9, 38–44. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, K.; Komaki, M.; Akazawa, K.; Nagata, M.; Yokoyama, N.; Watabe, T.; Morita, I. Spontaneous differentiation of periodontal ligament stem cells into myofibroblast during ex vivo expansion. J. Cell. Physiol. 2019, 234, 20377–20391. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Yu, F.; Cheng, Y.; Li, Y.; Chen, Y.; Tang, J.; Bei, Y.; Tang, Q.; Zhao, Y.; Huang, Y.; et al. Transforming growth factor-β3/recombinant human-like collagen/chitosan freeze-dried sponge primed with human periodontal ligament stem cells promotes bone regeneration in calvarial defect rats. Front. Pharmacol. 2021, 12, 678322. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Geng, D.; Kuang, Z.; Huang, S.; Cheng, Y.; Chen, Y.; Leng, F.; Bei, Y.; Zhao, Y.; Tang, Q.; et al. Sequentially releasing self-healing hydrogel fabricated with TGFβ3-microspheres and bFGF to facilitate rat alveolar bone defect repair. Asian J. Pharm. Sci. 2022, 17, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Shi, S.; Bartold, P.; Miura, M.; Seo, B.; Robey, P.; Gronthos, S. The efficacy of mesenchymal stem cells to regenerate and repair dental structures. Orthod. Craniofacial Res. 2005, 8, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Feng, F.; Akiyama, K.; Liu, Y.; Yamaza, T.; Wang, T.-M.; Chen, J.-H.; Wang, B.; Huang, G.T.-J.; Wang, S.; Shi, S. Utility of PDL progenitors for in vivo tissue regeneration: A report of 3 cases. Oral Dis. 2010, 16, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Hinz, B. The role of myofibroblasts in wound healing. Curr. Res. Transl. Med. 2016, 64, 171–177. [Google Scholar] [CrossRef]
- Al-Habeeb, F.; Aloufi, N.; Traboulsi, H.; Liu, X.; Nair, P.; Haston, C.; Azuelos, I.; Huang, S.K.; White, E.S.; Gallouzi, I.E.; et al. Human antigen R promotes lung fibroblast differentiation to myofibroblasts and increases extracellular matrix production. J. Cell. Physiol. 2021, 236, 6836–6851. [Google Scholar] [CrossRef]
- Čater, M.; Majdič, G. In vitro culturing of adult stem cells: The importance of serum and atmospheric oxygen. Adv. Exp. Med. Biol. 2022, 1376, 101–118. [Google Scholar] [CrossRef] [PubMed]
- Jung, I.; Kwon, B.; Kim, S.; Shim, H.; Jun, C.; Yun, J. Optimal medium formulation for the long-term expansion and maintenance of human periodontal ligament stem cells. J. Periodontol. 2013, 84, 1434–1444. [Google Scholar] [CrossRef] [PubMed]
- Dworzański, J.; Strycharz-Dudziak, M.; Kliszczewska, E.; Kiełczykowska, M.; Dworzańska, A.; Drop, B.; Polz-Dacewicz, M. Glutathione peroxidase (GPx) and superoxide dismutase (SOD) activity in patients with diabetes mellitus type 2 infected with Epstein-Barr virus. PLoS ONE 2020, 15, e0230374. [Google Scholar] [CrossRef] [PubMed]
- Fijany, A.; Sayadi, L.R.; Khoshab, N.; Banyard, D.A.; Shaterian, A.; Alexander, M.; Lakey, J.R.T.; Paydar, K.Z.; Evans, G.R.D.; Widgerow, A.D. Mesenchymal stem cell dysfunction in diabetes. Mol. Biol. Rep. 2019, 46, 1459–1475. [Google Scholar] [CrossRef] [PubMed]
- Qi, Y.; Ma, J.; Li, S.; Liu, W. Applicability of adipose-derived mesenchymal stem cells in treatment of patients with type 2 diabetes. Stem Cell Res. Ther. 2019, 10, 274. [Google Scholar] [CrossRef] [PubMed]
- Grohová, A.; Dáňová, K.; Špíšek, R.; Palová-Jelínková, L. Cell based therapy for type 1 diabetes: Should we take hyperglycemia into account? Front. Immunol. 2019, 10, 79. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, X.; Wang, Z.; Li, K.; Xu, B.; Zhang, J. Berberine improves advanced glycation end products-induced osteogenic differentiation responses in human periodontal ligament stem cells through the canonical Wnt/β-catenin pathway. Mol. Med. Rep. 2019, 19, 5440–5452. [Google Scholar] [CrossRef]
- Oyebode, O.A.; Jere, S.W.; Houreld, N.N. Current therapeutic modalities for the management of chronic diabetic wounds of the foot. J. Diabetes Res. 2023, 2023, 1359537. [Google Scholar] [CrossRef]
- Singh, W.R.; Sharma, A.; Devi, H.S.; Bhatia, A.; Patel, M.R.; Kumar, D. Icariin improves cutaneous wound healing in streptozotocin-induced diabetic rats. J. Tissue Viability 2021, 31, 197–206. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Y.; He, W.; Mu, X.; Wu, X.; Deng, J.; Nie, X. Fibroblasts: Immunomodulatory factors in refractory diabetic wound healing. Front. Immunol. 2022, 13, 918223. [Google Scholar] [CrossRef]
- Santonocito, S.; Ferlito, S.; Polizzi, A.; Ronsivalle, V.; Sclafani, R.; Valletta, A.; Giudice, A.L.; Cavalcanti, R.; Spagnuolo, G.; Isola, G. Therapeutic and metagenomic potential of the biomolecular therapies against periodontitis and the oral microbiome: Current evidence and future perspectives. Int. J. Mol. Sci. 2022, 23, 13708. [Google Scholar] [CrossRef] [PubMed]
- Baik, H.-S.; Park, J.; Lee, K.-J.; Chung, C. Local application of periodontal ligament stromal cells promotes soft tissue regeneration. Oral Dis. 2014, 20, 574–581. [Google Scholar] [CrossRef] [PubMed]
- Retamal, I.; Hernandez, R.; Velarde, V.; Oyarzun, A.; Martinez, C.; Gonzalez, M.J.; Martinez, J.; Smith, P.C. Diabetes alters the involvement of myofibroblasts during periodontal wound healing. Oral Dis. 2020, 26, 1062–1071. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.C. Role of myofibroblasts in normal and pathological periodontal wound healing. Oral Dis. 2018, 24, 26–29. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Su, Q.; Tao, Z.; Cai, X.; Zhao, Y.; Zhou, Z.; Huang, Y.; Xiang, Q. Human Periodontal Ligament Stem Cells (hPDLSCs) Spontaneously Differentiate into Myofibroblasts to Repair Diabetic Wounds. Bioengineering 2024, 11, 602. https://doi.org/10.3390/bioengineering11060602
Li Y, Su Q, Tao Z, Cai X, Zhao Y, Zhou Z, Huang Y, Xiang Q. Human Periodontal Ligament Stem Cells (hPDLSCs) Spontaneously Differentiate into Myofibroblasts to Repair Diabetic Wounds. Bioengineering. 2024; 11(6):602. https://doi.org/10.3390/bioengineering11060602
Chicago/Turabian StyleLi, Yuxiao, Qi Su, Zhaoyu Tao, Xiang Cai, Yueping Zhao, Zhiying Zhou, Yadong Huang, and Qi Xiang. 2024. "Human Periodontal Ligament Stem Cells (hPDLSCs) Spontaneously Differentiate into Myofibroblasts to Repair Diabetic Wounds" Bioengineering 11, no. 6: 602. https://doi.org/10.3390/bioengineering11060602
APA StyleLi, Y., Su, Q., Tao, Z., Cai, X., Zhao, Y., Zhou, Z., Huang, Y., & Xiang, Q. (2024). Human Periodontal Ligament Stem Cells (hPDLSCs) Spontaneously Differentiate into Myofibroblasts to Repair Diabetic Wounds. Bioengineering, 11(6), 602. https://doi.org/10.3390/bioengineering11060602