Elastin-Dependent Aortic Heart Valve Leaflet Curvature Changes During Cyclic Flexure

 , ,
, , 


Abstract
1. Introduction
2. Materials and Methods
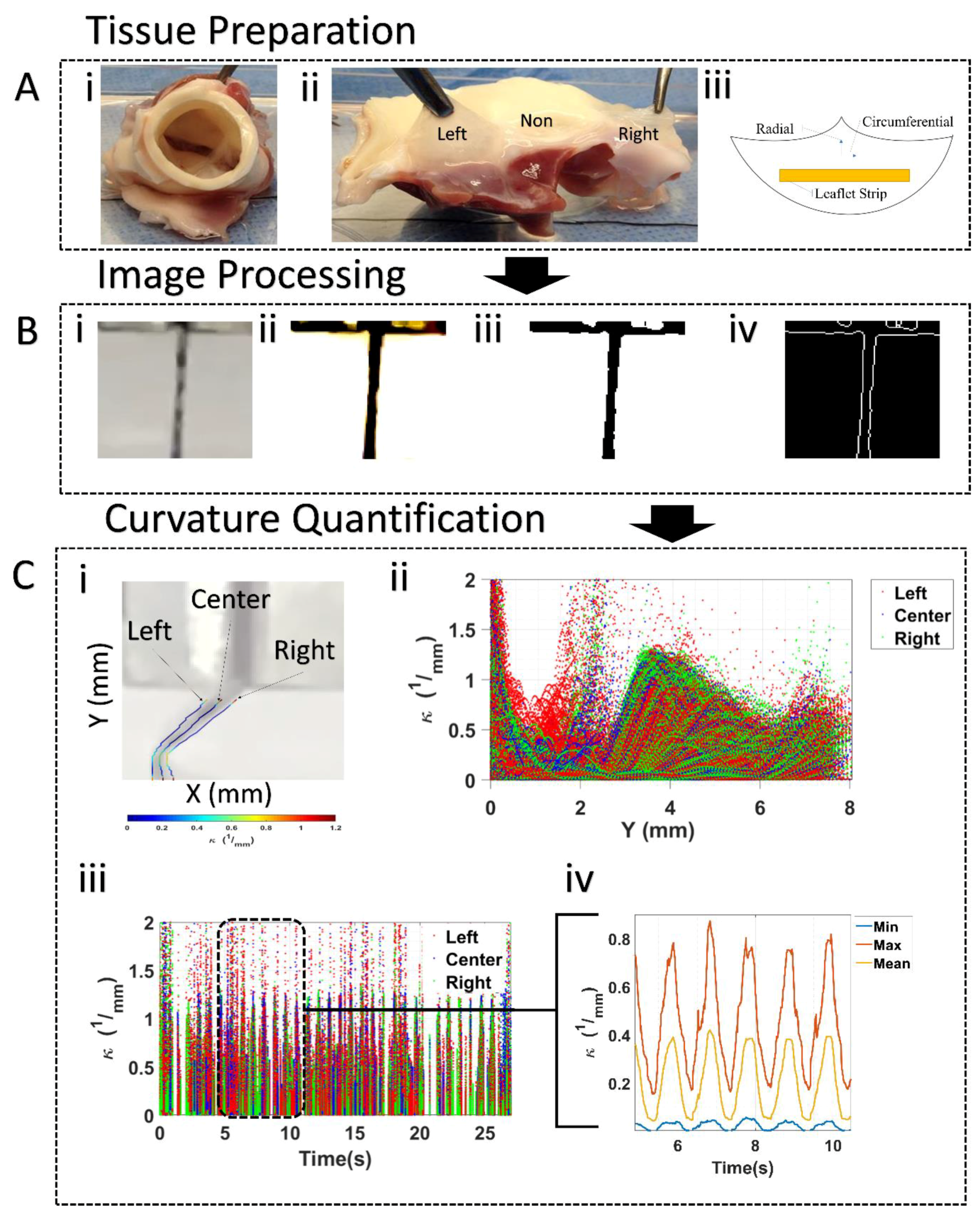
2.1. Tissue Sample Preparation
Elastin Degradation
2.2. Cyclic Flexure Experiments
Curvature Assessment
2.3. Elastin Assay
2.4. Elastin Structure
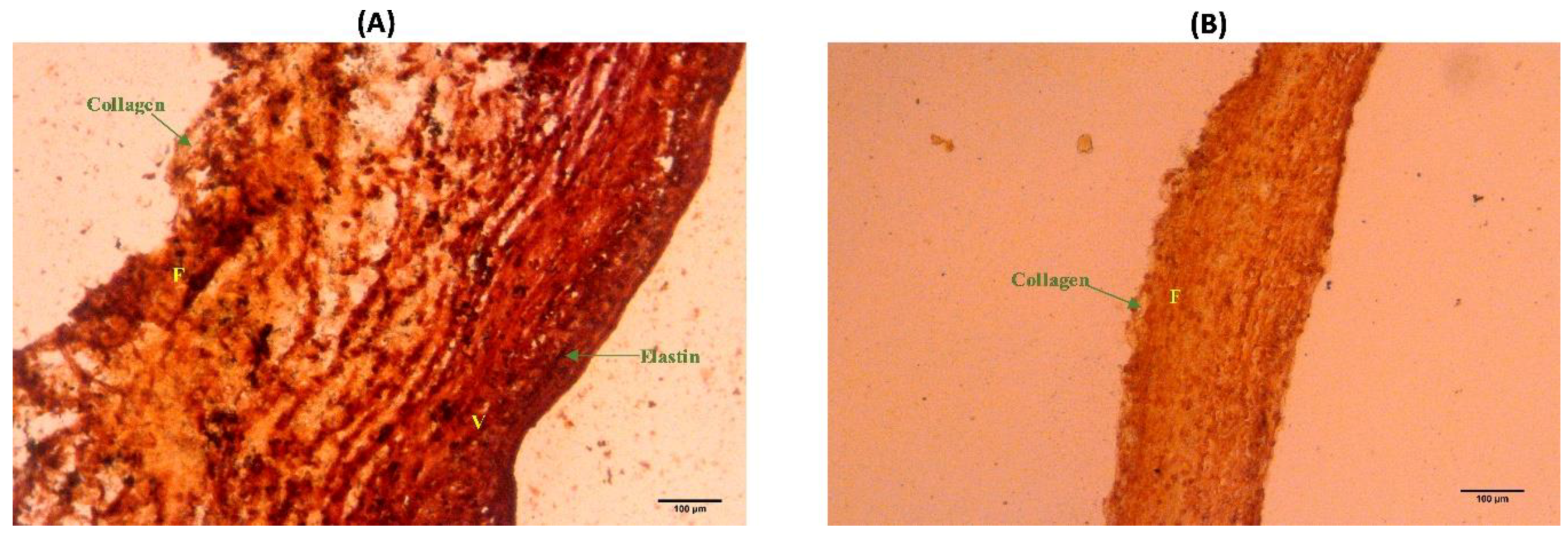
2.5. Histological Staining
2.6. Statistical Analysis
3. Results
3.1. Leaflet Shape Changes
Curvature Computation
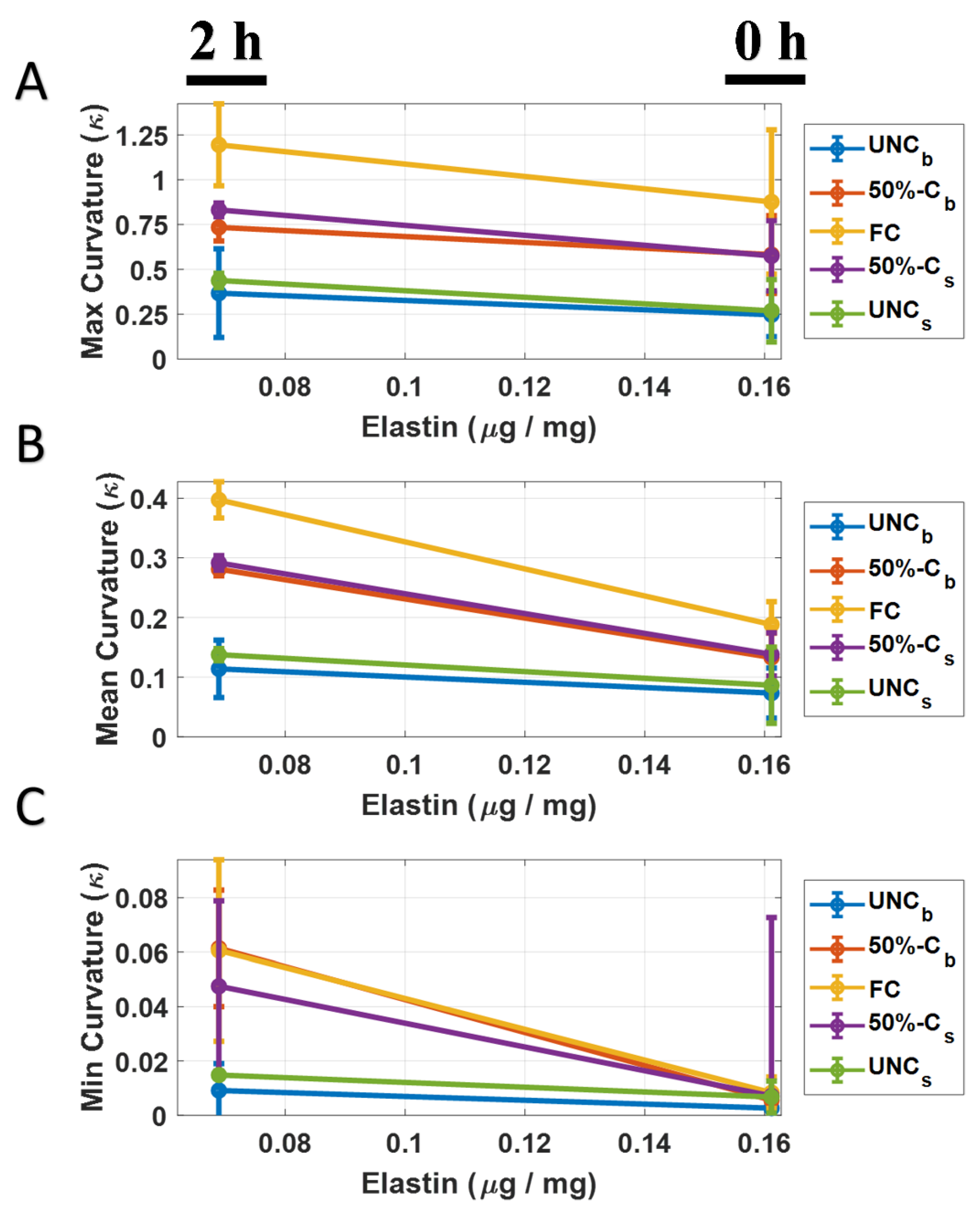
3.2. Leaflet Elastin Loss
3.3. Elastin Curvature Comparison
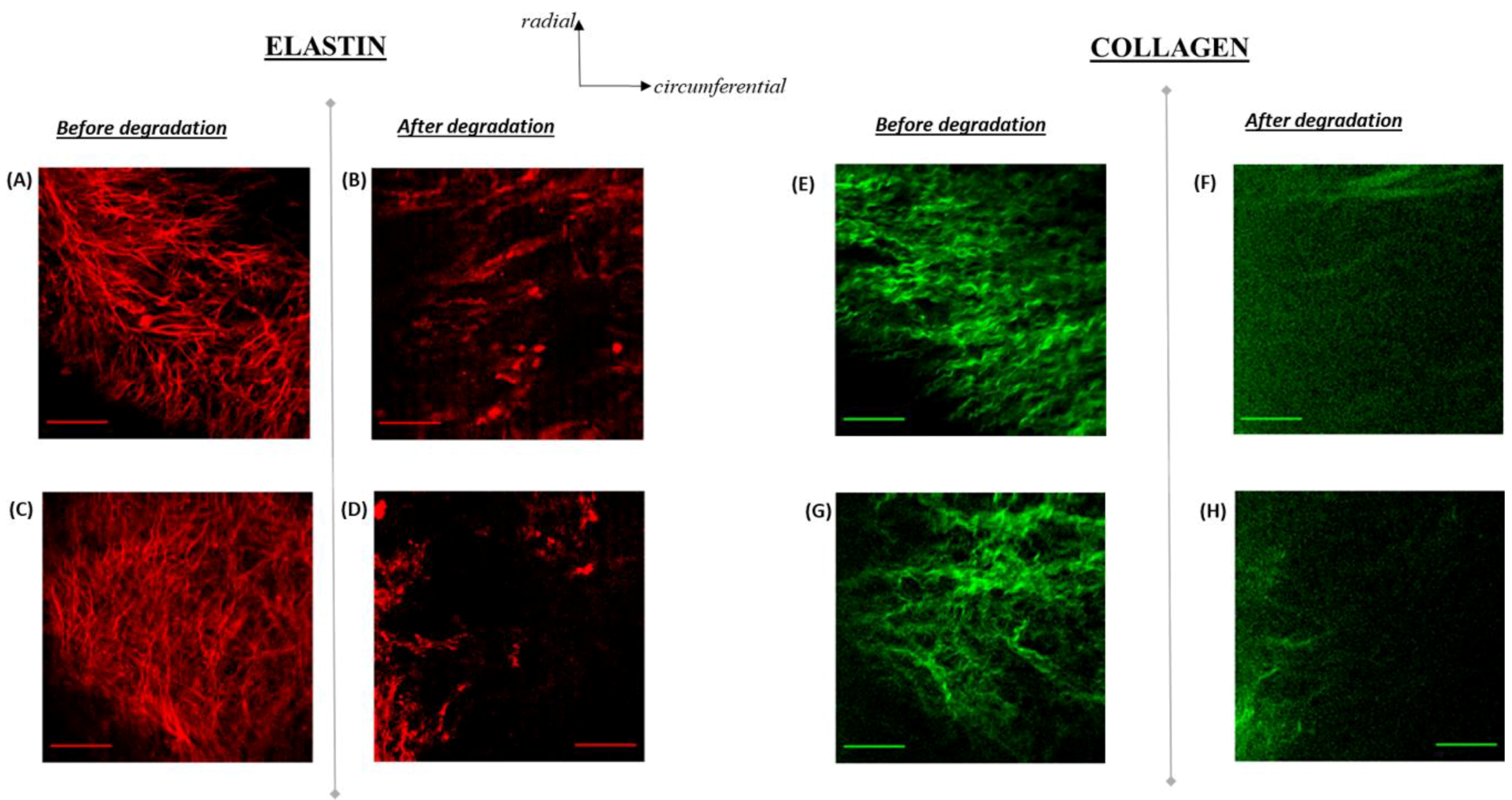
3.4. Elastin Structure
3.5. Elastin Distribution
4. Discussion and Conclusions
5. Limitations
Author Contributions
Funding
Conflicts of Interest
References
- Perrotta, I.; Camastra, C.; Filice, G.; Di Mizio, G.; Colosimo, F.; Ricci, P.; Tripepi, S.; Amorosi, A.; Triumbari, F.; Donato, G.; et al. New evidence for a critical role of elastin in calcification of native heart valves: Immunohistochemical and ultrastructural study with literature review. Histopathology 2011, 59, 504–513. [Google Scholar] [CrossRef]
- Passik, C.S.; Ackermann, D.M.; Pluth, J.R.; Edwards, W.D. Temporal changes in the causes of aortic stenosis: A surgical pathologic study of 646 cases. Mayo Clin. Proc. 1987, 62, 119–123. [Google Scholar] [CrossRef]
- Rutkovskiy, A.; Malashicheva, A.; Sullivan, G.; Bogdanova, M.; Kostareva, A.; Stensløkken, K.; Fiane, A.; Vaage, J. Valve interstitial cells: The key to understanding the pathophysiology of heart valve calcification. J. Am. Heart Assoc. 2017, 6, 6. [Google Scholar] [CrossRef]
- Yutzey, K.E.; Demer, L.L.; Body, S.C.; Huggins, G.S.; Towler, D.A.; Giachelli, C.M.; Hofmann-Bowman, M.A.; Mortlock, D.P.; Rogers, M.B.; Sadeghi, M.M.; et al. Calcific aortic valve disease: A consensus summary from the Alliance of Investigators on Calcific Aortic Valve Disease. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 2387–2393. [Google Scholar] [CrossRef]
- Engelmayr, G.C.; Sales, V.L.; Mayer, J.E.; Sacks, M.S. Cyclic flexure and laminar flow synergistically accelerate mesenchymal stem cell-mediated engineered tissue formation: Implications for engineered heart valve tissues. Biomaterials 2006, 27, 6083–6095. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Sun, R.; Li, X.; Liu, M.; Chen, S.; Zhang, P. Pathophysiology of valvular heart disease. Exp. Ther. Med. 2016, 11, 1184–1188. [Google Scholar] [CrossRef]
- van der Ven, C.F.; Wu, P.J.; Tibbitt, M.W.; van Mil, A.; Sluijter, J.P.; Langer, R.; Aikawa, E. In vitro 3D model and miRNA drug delivery to target calcific aortic valve disease. Clin. Sci. (Lond.) 2017, 131, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Barbarash, L.; Rutkovskaya, N.; Barbarash, O.; Odarenko, Y.; Stasev, A.; Uchasova, E. Prosthetic heart valve selection in women of childbearing age with acquired heart disease: A case report. J. Med. Case Rep. 2016, 10, 51. [Google Scholar] [CrossRef] [PubMed]
- Hinton, R.B.; Lincoln, J.; Deutsch, G.H.; Osinska, H.; Manning, P.B.; Benson, D.W.; Yutzey, K.E. Extracellular matrix remodeling and organization in developing and diseased aortic valves. Circ. Res. 2006, 98, 1431–1438. [Google Scholar] [CrossRef]
- Hinton, R.B.; Adelman-Brown, J.; Witt, S.; Krishnamurthy, V.K.; Osinska, H.; Sakthivel, B.; James, J.F.; Li, D.Y.; Narmoneva, D.A.; Mecham, R.P.; et al. Elastin Haploinsufficiency Results in Progressive Aortic Valve Malformation and Latent Valve Disease in a Mouse Model Novelty and Significance. Circ. Res. 2010, 107, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Bleakley, C.; Monaghan, M.J. The Pivotal Role of Imaging in TAVR Procedures. Curr. Cardiol. Rep. 2018, 12, 9. [Google Scholar] [CrossRef]
- Gould, R.A.; Jonathan, T.B. Isolation of valvular endothelial cells. J. Vis. Exp. 2010, 46, e2158. [Google Scholar] [CrossRef]
- Roach, M.R.; Alan, C.B. The reason for the shape of the distensibility curves of arteries. Can. J. Biochem. Physiol. 1957, 35, 681–690. [Google Scholar] [CrossRef]
- Salinas, M.; Rath, S.; Villegas, A.; Unnikrishnan, V.; Ramaswamy, S. Relative effects of fluid oscillations and nutrient transport in the in vitro growth of valvular tissues. Cardiov. Eng. Technol. 2016, 7, 170–181. [Google Scholar] [CrossRef]
- Chen, X.; Nadiarynkh, O.; Plotnikov, S.; Campagnola, P. Second Harmonic Generation Microscopy for Quantitative Analysis of Collagen Fibrillar Structure. Nat. Protoc. 2012, 7, 654–669. [Google Scholar] [CrossRef] [PubMed]
- König, K.; Schenke-Layland, K.; Riemann, I.; Stock, U. Multiphoton autofluorescence imaging of intratissue elastic fibers. Biomaterials 2005, 5, 495–500. [Google Scholar] [CrossRef]
- Hinton, R.B.; Katherine, E.Y. Heart valve structure and function in development and disease. Ann. Rev. Physiol. 2011, 73, 29–46. [Google Scholar] [CrossRef]
- Aikawa, E.; Aikawa, M.; Libby, P.; Figueiredo, J.-L.; Rusanescu, G.; Iwamoto, Y.; Fukuda, D.; Kohler, R.H.; Shi, G.-P.; Jaffer, F.A.; et al. Arterial and aortic valve calcification abolished by elastolytic cathepsin S deficiency in chronic renal disease. Circulation 2009, 119, 1785–1794. [Google Scholar] [CrossRef] [PubMed]
- Hutcheson, J.D.; Aikawa, E.; Merryman, W.D. Potential drug targets for calcific aortic valve disease. Nat. Rev. Cardiol. 2014, 11. [Google Scholar] [CrossRef]
- Yap, C.H.; Saikrishnan, N.; Tamilselvan, G.; Yoganathan, A.P. Experimental measurement of dynamic fluid shear stress on the aortic surface of the aortic valve leaflet. Biomech. Model. Mechanobiol. 2012, 11, 171–182. [Google Scholar] [CrossRef]
- Chow, M.J.; Turcotte, R.; Lin, C.P.; Zhang, Y. Arterial Extracellular Matrix: A Mechanobiological Study of The Contributions and Interactions of Elastin and Collagen. Biophys. J. 2014, 106, 2684–2692. [Google Scholar] [CrossRef] [PubMed]
- Mirnajafi, A.; Raymer, J.M.; McClure, L.R.; Sacks, M.S. The flexural rigidity of the aortic valve leaflet in the commissural region. J. Biomech. 2006, 39, 2966–2973. [Google Scholar] [CrossRef] [PubMed]
- Bell, E.D.; Converse, M.; Mao, H.; Unnikrishnan, G.; Reifman, J.; Monson, K.L. Material Properties of Rat Middle Cerebral Arteries at High Strain Rates. J. Biomech. Eng. 2018. [Google Scholar] [CrossRef]
- Karunaratne, A.; Li, S.; Bull, A.M.J. Nano-scale Mechanisms Explain the Stiffening and Strengthening of Ligament Tissue with Increasing Strain Rate. Sci. Rep. 2018, 8. [Google Scholar] [CrossRef]
- Anssari-Benam, A.; Bucchi, A.; Screen, H.R.C.; Evans, S.L. A Transverse Isotropic Viscoelastic Constitutive Model for Aortic Valve Tissue. R. Soc. Open Sci. 2017. [Google Scholar] [CrossRef]
- Anssari-Benam, A.; Tseng, Y.-T.; Holzapfel, G.A.; Bucchi, A. Rate-dependency of The Mechanical Behavior of Semilunar Heart Valves under Biaxial Deformation. Acta Biomater. 2019, 88, 120–130. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | UNCb | 50%-Cb | FC | 50%-Cs | UNCs |
|---|---|---|---|---|---|
| Control 0 h |  |  |  |  |  |
| 2 h |  |  |  |  |  |
| Group | Amount of Bending | n | ||||
|---|---|---|---|---|---|---|
| Elastin Degradation | UNCb | 50%-Cb * | FC * | 50%-Cs * | UNCs | |
| 0 h (Control) | 0.07 ± 0.04 | 0.13 ± 0.04 | 0.19 ± 0.04 | 0.14 ± 0.04 | 0.09 ± 0.06 | 9 |
| 2 h | 0.11 ± 0.05 | 0.28 ± 0.01 | 0.40 ± 0.03 | 0.29 ± 0.01 | 0.14 ± 0.01 | 6 |
| p-Value | p > 0.05 | p < 0.05 | p < 0.05 | p < 0.05 | p > 0.05 | |
| Group | Amount of Bending | n | ||||
|---|---|---|---|---|---|---|
| Elastin Degradation | UNCb | 50%-Cb * | FC | 50%-Cs | UNCs | |
| 0 h(Control) | 0.0026 ± 0.0006 | 0.0055 ± 0.0017 | 0.0083 ± 0.0058 | 0.0071 ± 0.0656 | 0.0067 ± 0.0059 | 9 |
| 2 h | 0.0091 ± 0.0099 | 0.0614 ± 0.0214 | 0.0606 ± 0.0334 | 0.0475 ± 0.0313 | 0.0148 ± 0.0001 | 6 |
| p-Value | p > 0.05 | p < 0.05 | p >0.05 | p >0.05 | p > 0.05 | |
| Group | Amount of Bending | n | ||||
|---|---|---|---|---|---|---|
| Elastin Degradation | UNCb | 50%-Cb | FC | 50%-UCs | UNCs | |
| 0 h (Control) | 0.25 ± 0.12 | 0.58 ± 0.22 | 0.88 ± 0.40 | 0.58 ± 0.20 | 0.27 ± 0.17 | 9 |
| 2 h | 0.37 ± 0.25 | 0.73 ± 0.08 | 1.20 ± 0.23 | 0.83 ± 0.04 | 0.44 ± 0.04 | 6 |
| p-Value | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | p > 0.05 | |
| Group | Elastin Degradation | |
|---|---|---|
| Elastin (µg/mg) | 0 h (Control) | 2 h |
| 0.161 ± 0.07 | 0.069 ± 0.016 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tesfamariam, M.D.; Mirza, A.M.; Chaparro, D.; Ali, A.Z.; Montalvan, R.; Saytashev, I.; Gonzalez, B.A.; Barreto, A.; Ramella-Roman, J.; Hutcheson, J.D.; et al. Elastin-Dependent Aortic Heart Valve Leaflet Curvature Changes During Cyclic Flexure. Bioengineering 2019, 6, 39. https://doi.org/10.3390/bioengineering6020039
Tesfamariam MD, Mirza AM, Chaparro D, Ali AZ, Montalvan R, Saytashev I, Gonzalez BA, Barreto A, Ramella-Roman J, Hutcheson JD, et al. Elastin-Dependent Aortic Heart Valve Leaflet Curvature Changes During Cyclic Flexure. Bioengineering. 2019; 6(2):39. https://doi.org/10.3390/bioengineering6020039
Chicago/Turabian StyleTesfamariam, Melake D., Asad M. Mirza, Daniel Chaparro, Ahmed Z. Ali, Rachel Montalvan, Ilyas Saytashev, Brittany A. Gonzalez, Amanda Barreto, Jessica Ramella-Roman, Joshua D. Hutcheson, and et al. 2019. "Elastin-Dependent Aortic Heart Valve Leaflet Curvature Changes During Cyclic Flexure" Bioengineering 6, no. 2: 39. https://doi.org/10.3390/bioengineering6020039
APA StyleTesfamariam, M. D., Mirza, A. M., Chaparro, D., Ali, A. Z., Montalvan, R., Saytashev, I., Gonzalez, B. A., Barreto, A., Ramella-Roman, J., Hutcheson, J. D., & Ramaswamy, S. (2019). Elastin-Dependent Aortic Heart Valve Leaflet Curvature Changes During Cyclic Flexure. Bioengineering, 6(2), 39. https://doi.org/10.3390/bioengineering6020039