Skeletal Muscle Tissue Engineering: Biomaterials-Based Strategies for the Treatment of Volumetric Muscle Loss


Abstract
:1. Clinical Need: Volumetric Muscle Loss
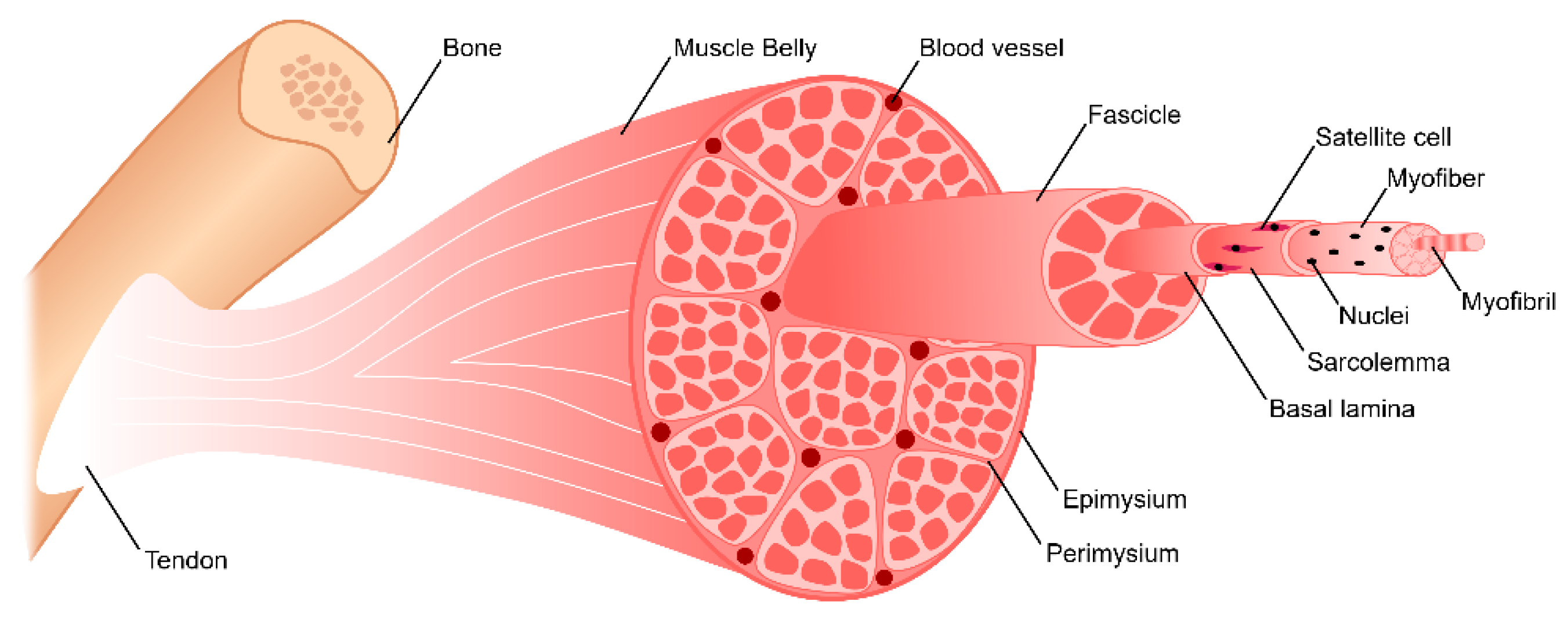
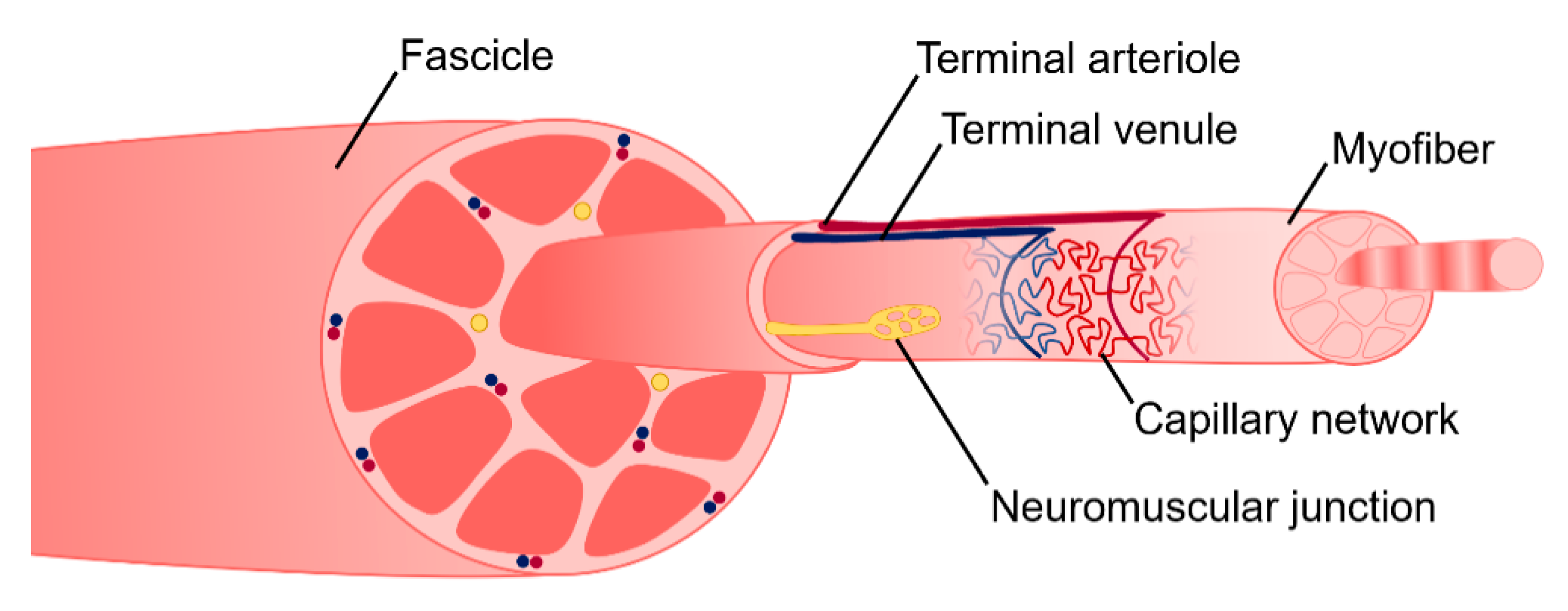
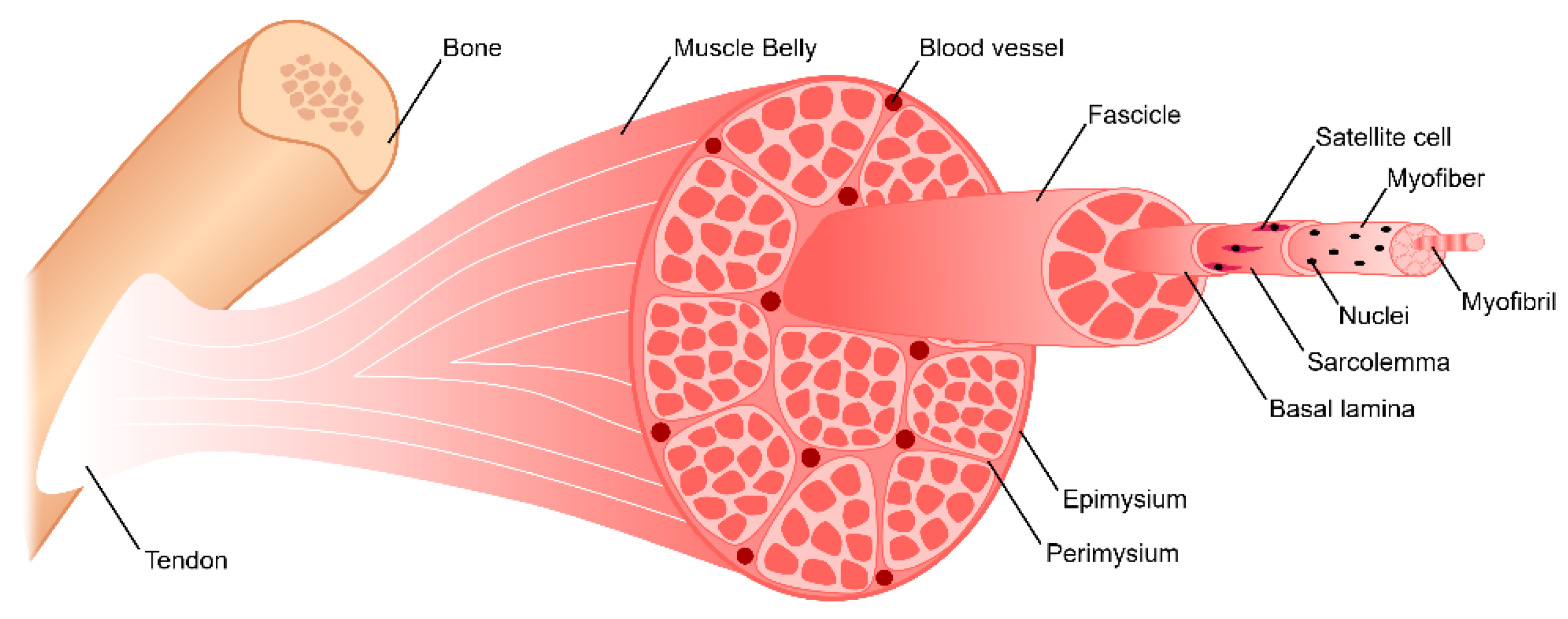
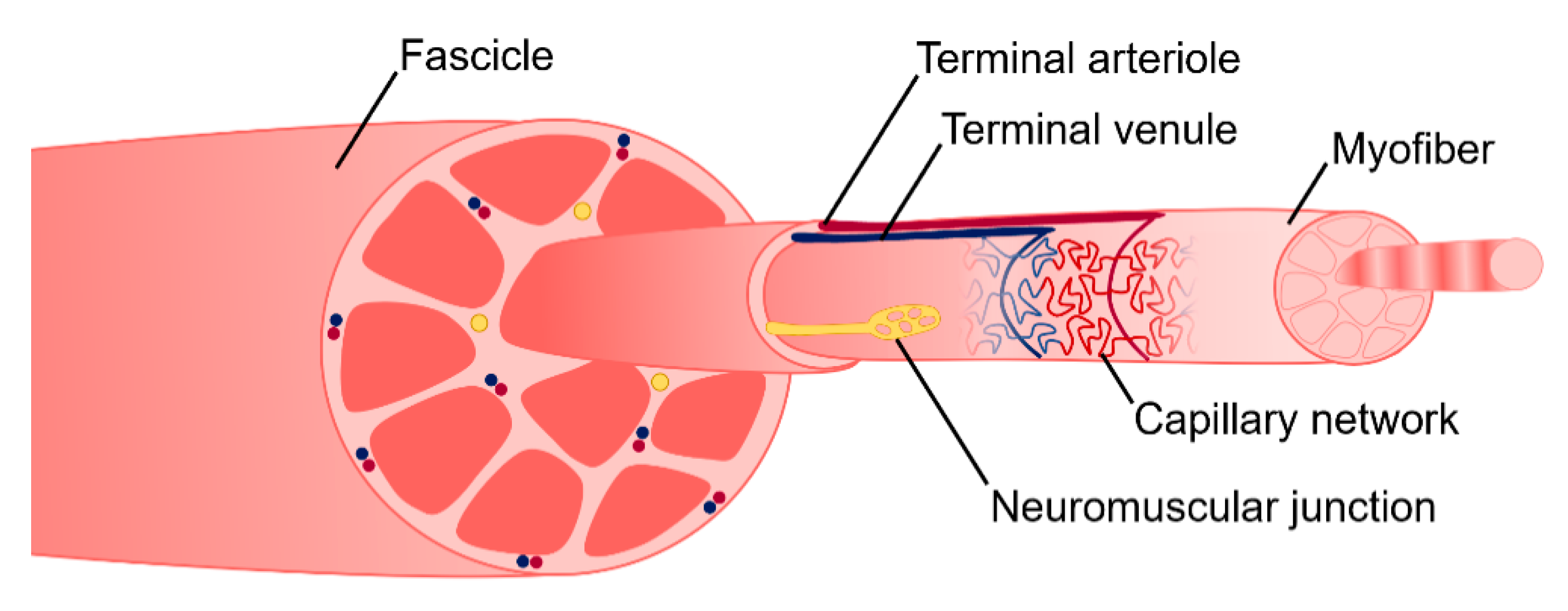
2. Skeletal Muscle Anatomy
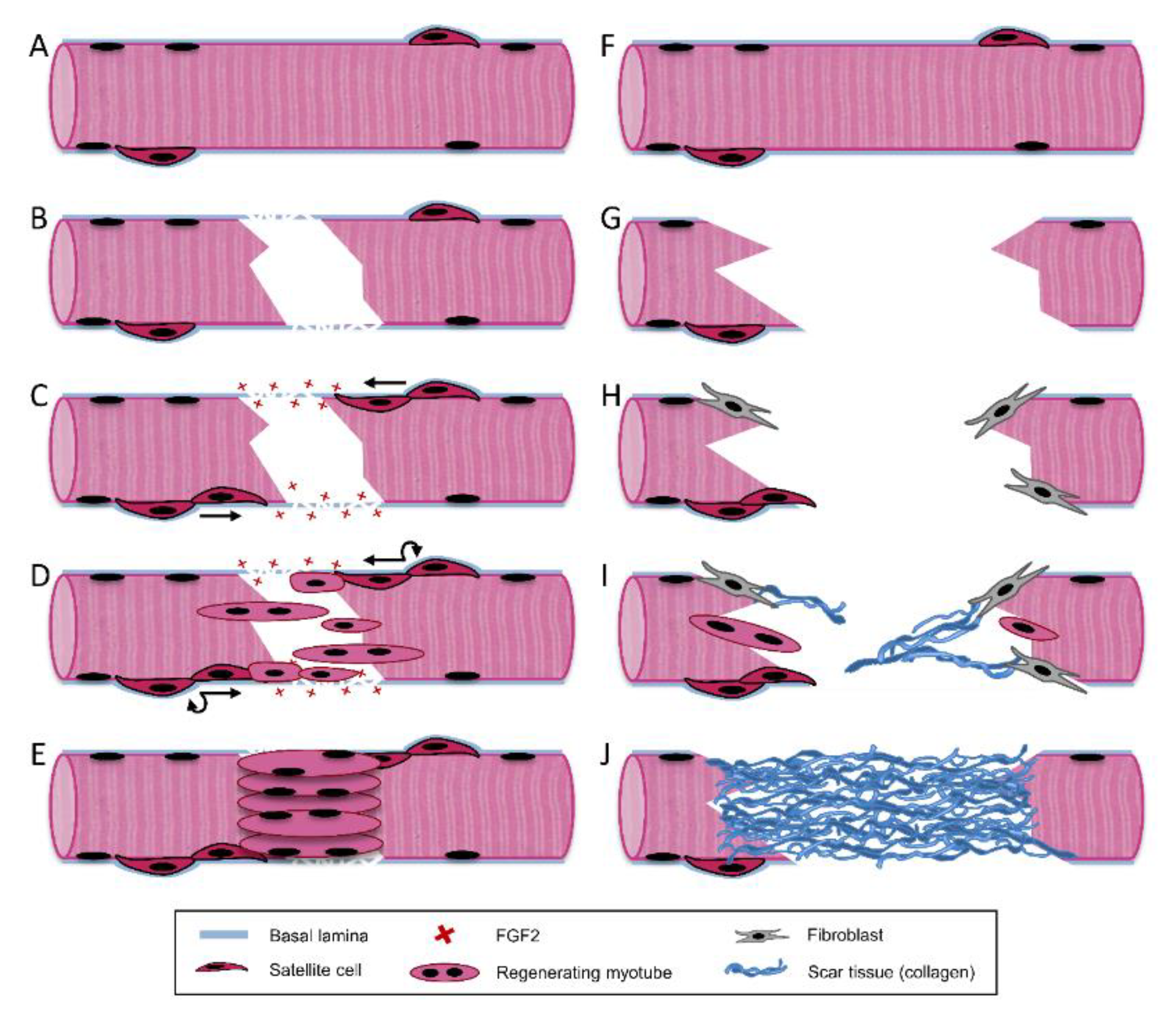
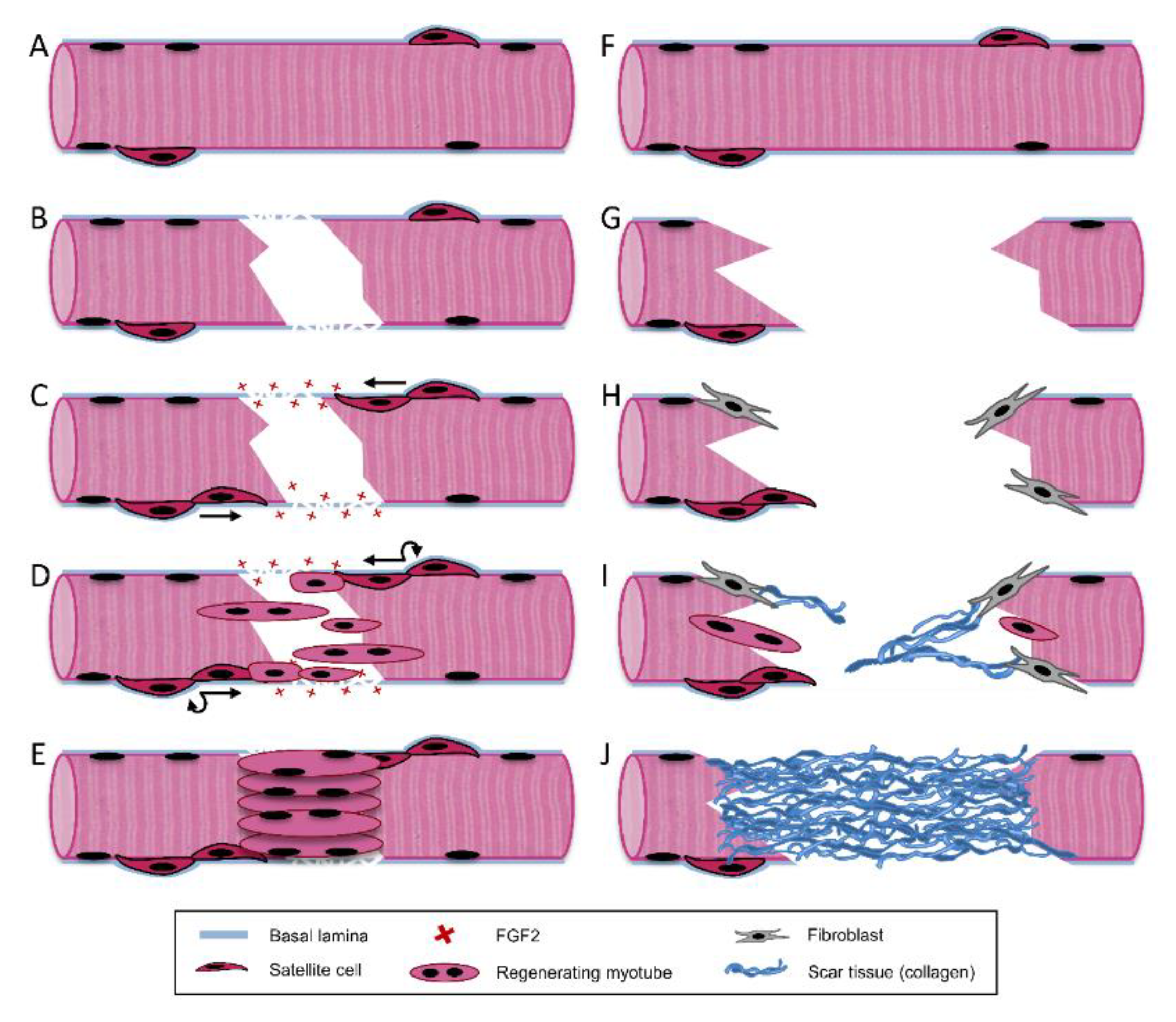
3. Skeletal Muscle Regeneration
3.1. Destruction/Inflammatory Phase
3.2. Repair Phase
3.3. Remodeling Phase
3.4. Limited Capacity for Regeneration in VML Injuries
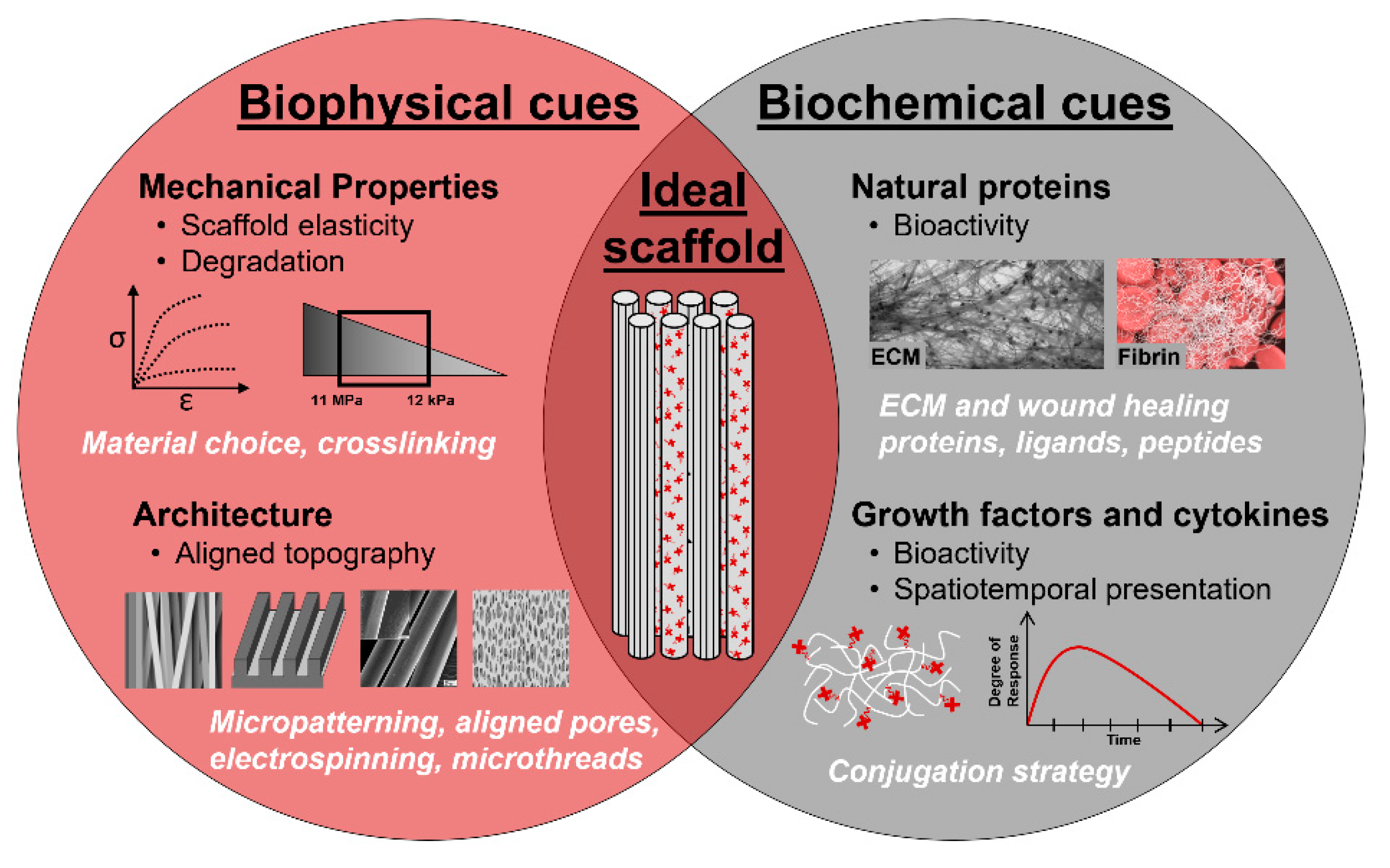
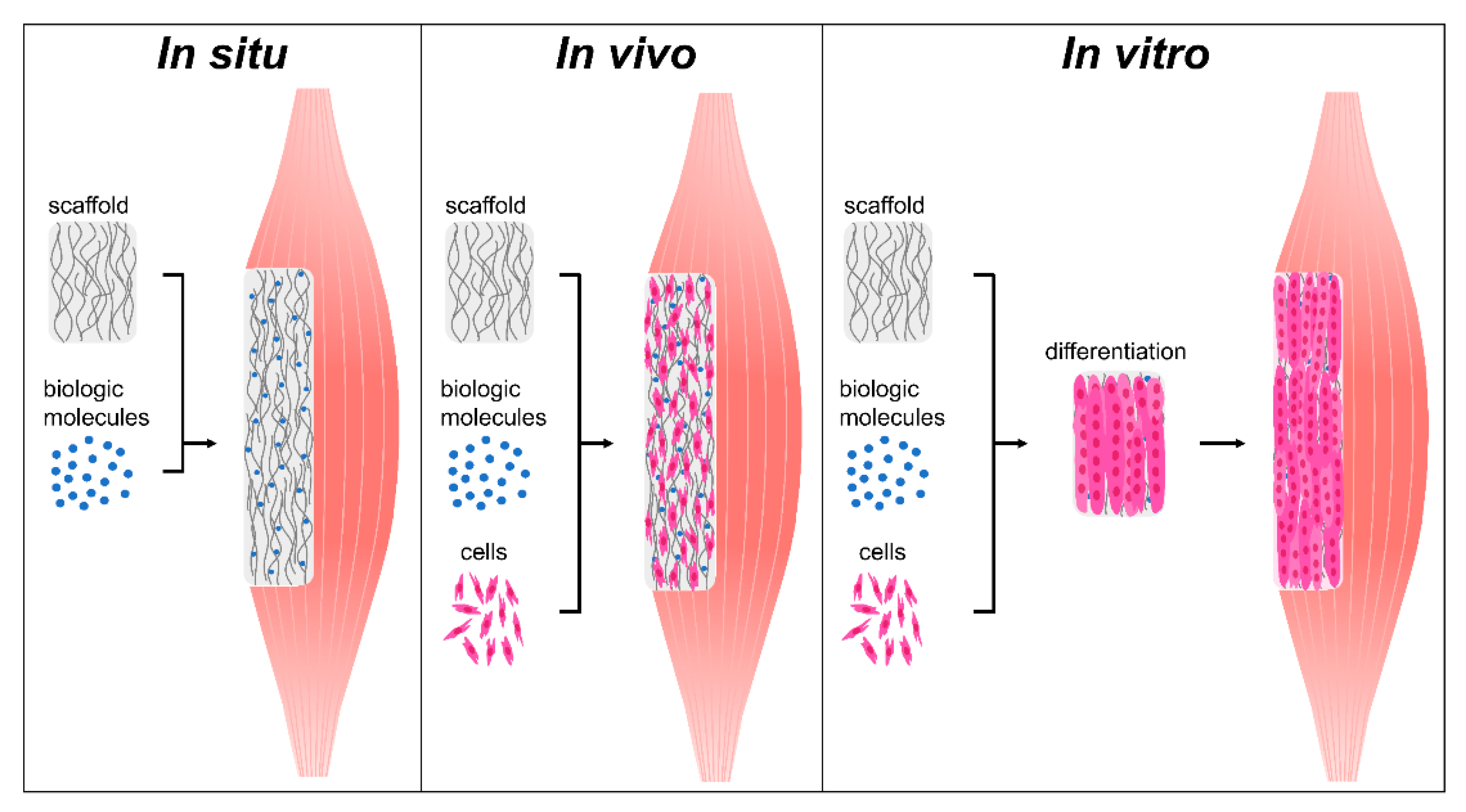
4. Biomaterial Strategies for Skeletal Muscle Regeneration
4.1. In Situ Strategies: Acellular Scaffolds to Promote Endogenous Regeneration
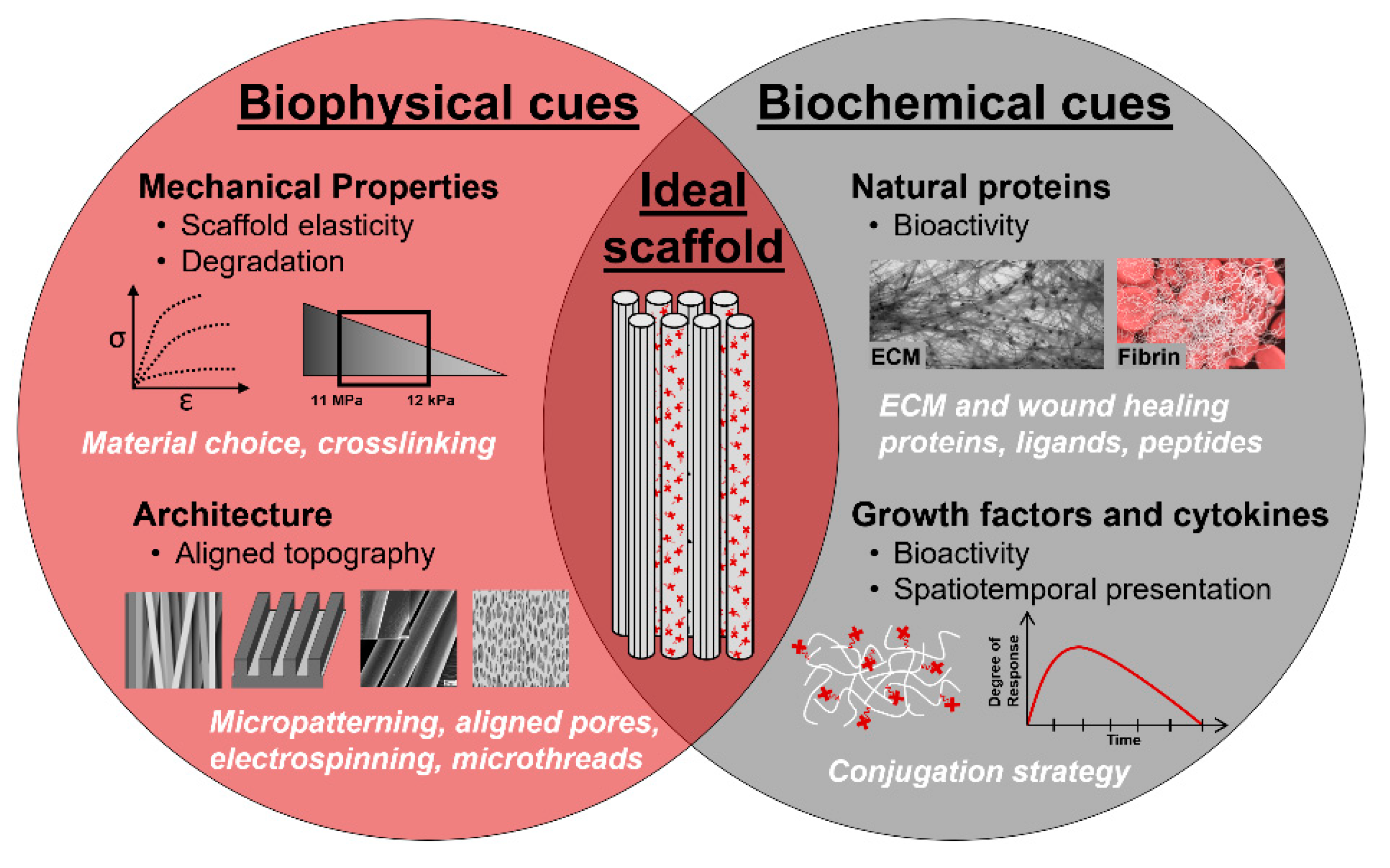
4.1.1. Biomaterial Selection: Synthetic, Natural, and Hybrid Polymers
4.1.2. Biophysical Cues
Decellularized ECM
Hydrogels, Sponges, and Meshes
Aligned Scaffolds
4.1.3. Biochemical Cues
Growth Factors and Cytokines
Genetic Substances
Small Molecules
4.2. In Vivo Strategies: Utilizing Biomaterials to Improve Cell Delivery
4.2.1. Cell Source
4.2.2. Hydrogel-Based Delivery
4.2.3. Decellularized ECM-Based Delivery
4.2.4. Microfiber-Based Delivery
4.2.5. Growth Factor-Loaded Scaffolds
4.2.6. Genetically Modified Cells
4.3. In Vitro Strategies: Developing Mature Tissue Constructs Prior to Implantation
4.3.1. Aligned Scaffolds
4.3.2. Mechanical and Electrical Stimulation
4.3.3. Angiogenesis and Innervation
5. Conclusions and Future Directions
Funding
Conflicts of Interest
References
- Center for Disease Control. National Hospital Discharge Survey (NHDS); Center for Disease Control: Atlanta, GA, USA, 2010.
- Center for Disease Control. National Hospital Ambulatory Medical Care Survey Outpatient Department (NHAMCS_OP); Center for Disease Control: Atlanta, GA, USA, 2010.
- Center for Disease Control. National Hospital Ambulatory Medical Care Survey Emergency Department (NHAMCS_ED); Center for Disease Control: Atlanta, GA, USA, 2010.
- Center for Disease Control. National Ambulatory Medical Care Survey (NAMCS); Center for Disease Control: Atlanta, GA, USA, 2010.
- Agency for Healthcare Research and Quality, Department of Health and Human Services. Medical Expenditures Panel Survey (MEPS) 1996–2011; Agency for Healthcare Research and Quality: Rockville, MD, USA, 2011.
- Disease, G.B.D.; Injury, I.; Prevalence, C. Global, regional, and national incidence, prevalence, and years lived with disability for 328 diseases and injuries for 195 countries, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet 2017, 390, 1211–1259. [Google Scholar] [CrossRef] [Green Version]
- Corso, P.; Finkelstein, E.; Miller, T.; Fiebelkorn, I.; Zaloshnja, E. Incidence and lifetime costs of injuries in the United States. Inj. Prev. 2015, 21, 434–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devore, D.I.; Walters, T.J.; Christy, R.J.; Rathbone, C.R.; Hsu, J.R.; Baer, D.G.; Wenke, J.C. For combat wounded: Extremity trauma therapies from the USAISR. Mil. Med. 2011, 176, 660–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- American Society of Plastic Surgeons. Plastic Surgery Statistics Report; National Clearinghouse of Plastic Surgery: Arlington Heights, IL, USA, 2014. [Google Scholar]
- Masini, B.D.; Waterman, S.M.; Wenke, J.C.; Owens, B.D.; Hsu, J.R.; Ficke, J.R. Resource utilization and disability outcome assessment of combat casualties from Operation Iraqi Freedom and Operation Enduring Freedom. J. Orthop. Trauma 2009, 23, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Greer, M.A.; Miklos-Essenberg, M.E.; Harrison-Weaver, S. A review of 41 upper extremity war injuries and the protective gear worn during Operation Enduring Freedom and Operation Iraqi Freedom. Mil. Med. 2006, 171, 595–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Owens, B.D.; Kragh, J.F., Jr.; Wenke, J.C.; Macaitis, J.; Wade, C.E.; Holcomb, J.B. Combat wounds in operation Iraqi Freedom and operation Enduring Freedom. J. Trauma 2008, 64, 295–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rivera, J.C.; Corona, B.T. Muscle-related disability following combat injury increases with time. US Army Med. Dep. J. 2016, 30–34. [Google Scholar]
- Corona, B.T.; Rivera, J.C.; Owens, J.G.; Wenke, J.C.; Rathbone, C.R. Volumetric muscle loss leads to permanent disability following extremity trauma. J. Rehabil. Res. Dev. 2015, 52, 785–792. [Google Scholar] [CrossRef]
- Cross, J.D.; Ficke, J.R.; Hsu, J.R.; Masini, B.D.; Wenke, J.C. Battlefield orthopaedic injuries cause the majority of long-term disabilities. J. Am. Acad. Orthop. Surg. 2011, 19 (Suppl. 1), S1–S7. [Google Scholar] [CrossRef]
- Garg, K.; Ward, C.L.; Hurtgen, B.J.; Wilken, J.M.; Stinner, D.J.; Wenke, J.C.; Owens, J.G.; Corona, B.T. Volumetric muscle loss: Persistent functional deficits beyond frank loss of tissue. J. Orthop. Res. 2015, 33, 40–46. [Google Scholar] [CrossRef]
- Mase, V.J., Jr.; Hsu, J.R.; Wolf, S.E.; Wenke, J.C.; Baer, D.G.; Owens, J.; Badylak, S.F.; Walters, T.J. Clinical application of an acellular biologic scaffold for surgical repair of a large, traumatic quadriceps femoris muscle defect. Orthopedics 2010, 33, 511. [Google Scholar] [CrossRef]
- Aurora, A.; Garg, K.; Corona, B.T.; Walters, T.J. Physical rehabilitation improves muscle function following volumetric muscle loss injury. BMC Sports Sci. Med. Rehabil. 2014, 6, 41. [Google Scholar] [CrossRef] [Green Version]
- Grogan, B.F.; Hsu, J.R.; Consortium, S.T.R. Volumetric muscle loss. J. Am. Acad. Orthop. Sur. 2011, 19, S35–S37. [Google Scholar] [CrossRef] [PubMed]
- Doi, K.; Hattori, Y.; Tan, S.H.; Dhawan, V. Basic science behind functioning free muscle transplantation. Clin. Plast. Surg. 2002, 29, 483–495. [Google Scholar] [CrossRef]
- Eckardt, A. Microsurgical reconstruction in the head and neck region: An 18-year experience with 500 consecutive cases. J. Cranio Maxillofac. Surg. 2003, 31, 197–201. [Google Scholar] [CrossRef]
- Lin, C.H.; Lin, Y.T.; Yeh, J.T.; Chen, C.T. Free functioning muscle transfer for lower extremity posttraumatic composite structure and functional defect. Plast. Reconstr. Surg. 2007, 119, 2118–2126. [Google Scholar] [CrossRef]
- Riccio, M.; Zingaretti, N.; Verdini, F.; Marchesini, A.; De Francesco, F.; Parodi, P.C. Functional donor-site morbidity after soleus muscle-flap procedure in the treatment of lower limb severe injuries. Handchir. Mikrochir. Plast. Chir. 2019, 51, 453–463. [Google Scholar] [CrossRef] [Green Version]
- Pochini, A.D.; Andreoli, C.V.; Belangero, P.S.; Figueiredo, E.A.; Terra, B.B.; Cohen, C.; Andrade, M.D.; Cohen, M.; Ejnisman, B. Clinical considerations for the surgical treatment of pectoralis major muscle ruptures based on 60 cases: A prospecitve study and literature review. Am. J. Sports Med. 2014, 42, 95–102. [Google Scholar] [CrossRef]
- Äärimaa, V.; Rantanen, J.; Heikkilä, J.; Helttula, I.; Orava, S. Rupture of the pectoralis major muscle. Am. J. Sports Med. 2004, 32, 1256–1262. [Google Scholar] [CrossRef]
- Diwan, A.; Eberlin, K.R.; Smith, R.M. The principles and practice of open fracture care, 2018. Chin. J. Traumatol. 2018, 21, 187–192. [Google Scholar] [CrossRef]
- Bianchi, B.; Copelli, C.; Ferrari, S.; Ferri, A.; Sesenna, E. Free flaps: Outcomes and complications in head and neck reconstructions. J. Craniomaxillofac. Surg. 2009, 37, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Lawson, R.; Levin, L.S. Principles of free tissue transfer in orthopaedic practice. J. Am. Acad. Orthop. Surg. 2007, 15, 290–299. [Google Scholar] [CrossRef] [PubMed]
- Qazi, T.H.; Mooney, D.J.; Pumberger, M.; Geißler, S.; Duda, G.N. Biomaterials based strategies for skeletal muscle tissue engineering: Existing technologies and future trends. Biomaterials 2015, 53, 502–521. [Google Scholar] [CrossRef]
- Jarvinen, T.A.; Jarvinen, T.L.; Kaariainen, M.; Kalimo, H.; Jarvinen, M. Muscle injuries: Biology and treatment. Am. J. Sports Med. 2005, 33, 745–764. [Google Scholar] [CrossRef] [PubMed]
- Jarvinen, T.A.; Jarvinen, T.L.; Kaariainen, M.; Aarimaa, V.; Vaittinen, S.; Kalimo, H.; Jarvinen, M. Muscle injuries: Optimising recovery. Best Pract. Res. Clin. Rheumatol. 2007, 21, 317–331. [Google Scholar] [CrossRef]
- Sanes, J.R. The basement membrane/basal lamina of skeletal muscle. J. Biol. Chem. 2003, 278, 12601–12604. [Google Scholar] [CrossRef] [Green Version]
- Gillies, A.R.; Lieber, R.L. Structure and function of the skeletal muscle extracellular matrix. Muscle Nerve 2011, 44, 318–331. [Google Scholar] [CrossRef] [Green Version]
- Folkman, J.; Klagsbrun, M.; Sasse, J.; Wadzinski, M.; Ingber, D.; Vlodavsky, I. A heparin-binding angiogenic protein—Basic fibroblast growth factor—Is stored within basement membrane. Am. J. Pathol. 1988, 130, 393–400. [Google Scholar]
- Naka, D.; Ishii, T.; Shimomura, T.; Hishida, T.; Hara, H. Heparin modulates the receptor-binding and mitogenic activity of hepatocyte growth factor on hepatocytes. Exp. Cell Res. 1993, 209, 317–324. [Google Scholar] [CrossRef]
- Roghani, M.; Moscatelli, D. Basic fibroblast growth factor is internalized through both receptor-mediated and heparan sulfate-mediated mechanisms. J. Biol. Chem. 1992, 267, 22156–22162. [Google Scholar]
- Rapraeger, A.C. Syndecan-regulated receptor signaling. J. Cell Biol. 2000, 149, 995–998. [Google Scholar] [CrossRef]
- Mauro, A. Satellite cell of skeletal muscle fibers. J. Biophys. Biochem. Cytol. 1961, 9, 493–495. [Google Scholar] [CrossRef] [PubMed]
- Rudnicki, M.A.; Le Grand, F.; McKinnell, I.; Kuang, S. The molecular regulation of muscle stem cell function. Cold Spring Harb. Symp. Quant. Biol. 2008, 73, 323–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Maltzahn, J.; Jones, A.E.; Parks, R.J.; Rudnicki, M.A. Pax7 is critical for the normal function of satellite cells in adult skeletal muscle. Proc. Natl. Acad. Sci. USA 2013, 110, 16474–16479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sambasivan, R.; Yao, R.; Kissenpfennig, A.; Van Wittenberghe, L.; Paldi, A.; Gayraud-Morel, B.; Guenou, H.; Malissen, B.; Tajbakhsh, S.; Galy, A. Pax7-expressing satellite cells are indispensable for adult skeletal muscle regeneration. Development 2011, 138, 3647–3656. [Google Scholar] [CrossRef] [Green Version]
- Lepper, C.; Partridge, T.A.; Fan, C.M. An absolute requirement for Pax7-positive satellite cells in acute injury-induced skeletal muscle regeneration. Development 2011, 138, 3639–3646. [Google Scholar] [CrossRef] [Green Version]
- Murphy, M.M.; Lawson, J.A.; Mathew, S.J.; Hutcheson, D.A.; Kardon, G. Satellite cells, connective tissue fibroblasts and their interactions are crucial for muscle regeneration. Development 2011, 138, 3625–3637. [Google Scholar] [CrossRef] [Green Version]
- Cosgrove, B.D.; Sacco, A.; Gilbert, P.M.; Blau, H.M. A home away from home: Challenges and opportunities in engineering in vitro muscle satellite cell niches. Differentiation 2009, 78, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Moss, F.P.; Leblond, C.P. Satellite cells as the source of nuclei in muscles of growing rats. Anat. Rec. 1971, 170, 421–435. [Google Scholar] [CrossRef]
- Hermansen, L.; Wachtlova, M. Capillary density of skeletal muscle in well-trained and untrained men. J. Appl. Physiol. 1971, 30, 860–863. [Google Scholar] [CrossRef]
- Grefte, S.; Kuijpers-Jagtman, A.M.; Torensma, R.; Von den Hoff, J.W. Skeletal muscle development and regeneration. Stem Cells Dev. 2007, 16, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Grasman, J.M.; Zayas, M.J.; Page, R.L.; Pins, G.D. Biomimetic scaffolds for regeneration of volumetric muscle loss in skeletal muscle injuries. Acta Biomater. 2015, 25, 2–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert-Honick, J.; Grayson, W. Vascularized and Innervated Skeletal Muscle Tissue Engineering. Adv. Healthc. Mater. 2020, 9, e1900626. [Google Scholar] [CrossRef] [PubMed]
- Ciciliot, S.; Schiaffino, S. Regeneration of mammalian skeletal muscle. Basic mechanisms and clinical implications. Curr. Pharm. Des. 2010, 16, 906–914. [Google Scholar] [CrossRef] [Green Version]
- Do, M.K.; Suzuki, T.; Gerelt, B.; Sato, Y.; Mizunoya, W.; Nakamura, M.; Ikeuchi, Y.; Anderson, J.E.; Tatsumi, R. Time-coordinated prevalence of extracellular HGF, FGF2 and TGF-beta3 in crush-injured skeletal muscle. Anim. Sci. J. 2012, 83, 712–717. [Google Scholar] [CrossRef]
- Tidball, J.G. Inflammatory cell response to acute muscle injury. Med. Sci. Sports Exerc. 1995, 27, 1022–1032. [Google Scholar] [CrossRef]
- Fielding, R.A.; Manfredi, T.J.; Ding, W.; Fiatarone, M.A.; Evans, W.J.; Cannon, J.G. Acute phase response in exercise. III. Neutrophil and IL-1 beta accumulation in skeletal muscle. Am. J. Physiol. 1993, 265, R166–R172. [Google Scholar] [CrossRef]
- Husmann, I.; Soulet, L.; Gautron, J.; Martelly, I.; Barritault, D. Growth factors in skeletal muscle regeneration. Cytokine Growth Factor Rev. 1996, 7, 249–258. [Google Scholar] [CrossRef]
- Charge, S.B.P.; Rudnicki, M.A. Cellular and molecular regulation of muscle regeneration. Physiol. Rev. 2004, 84, 209–238. [Google Scholar] [CrossRef]
- Toumi, H.; Best, T.M. The inflammatory response: Friend or enemy for muscle injury? Br. J. Sports Med. 2003, 37, 284–286. [Google Scholar] [CrossRef]
- Wahl, S.M.; Hunt, D.A.; Wakefield, L.M.; McCartney-Francis, N.; Wahl, L.M.; Roberts, A.B.; Sporn, M.B. Transforming growth factor type beta induces monocyte chemotaxis and growth factor production. Proc. Natl. Acad. Sci. USA 1987, 84, 5788–5792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mantovani, A.; Sozzani, S.; Locati, M.; Allavena, P.; Sica, A. Macrophage polarization: Tumor-associated macrophages as a paradigm for polarized M2 mononuclear phagocytes. Trends Immunol. 2002, 23, 549–555. [Google Scholar] [CrossRef]
- Martinez, F.O.; Sica, A.; Mantovani, A.; Locati, M. Macrophage activation and polarization. Front. Biosci. 2008, 13, 453–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saclier, M.; Yacoub-Youssef, H.; Mackey, A.L.; Arnold, L.; Ardjoune, H.; Magnan, M.; Sailhan, F.; Chelly, J.; Pavlath, G.K.; Mounier, R.; et al. Differentially activated macrophages orchestrate myogenic precursor cell fate during human skeletal muscle regeneration. Stem Cells 2013, 31, 384–396. [Google Scholar] [CrossRef] [Green Version]
- Philippou, A.; Maridaki, M.; Theos, A.; Koutsilieris, M. Cytokines in muscle damage. Adv. Clin. Chem. 2012, 58, 49–87. [Google Scholar] [CrossRef]
- Tidball, J.G.; Dorshkind, K.; Wehling-Henricks, M. Shared signaling systems in myeloid cell-mediated muscle regeneration. Development 2014, 141, 1184–1196. [Google Scholar] [CrossRef] [Green Version]
- Badylak, S.F.; Dziki, J.L.; Sicari, B.M.; Ambrosio, F.; Boninger, M.L. Mechanisms by which acellular biologic scaffolds promote functional skeletal muscle restoration. Biomaterials 2016, 103, 128–136. [Google Scholar] [CrossRef]
- Cantini, M.; Giurisato, E.; Radu, C.; Tiozzo, S.; Pampinella, F.; Senigaglia, D.; Zaniolo, G.; Mazzoleni, F.; Vitiello, L. Macrophage-secreted myogenic factors: A promising tool for greatly enhancing the proliferative capacity of myoblasts in vitro and in vivo. Neurol. Sci. 2002, 23, 189–194. [Google Scholar] [CrossRef]
- Huard, J.; Lu, A.; Mu, X.; Guo, P.; Li, Y. Muscle injuries and repair: What’s new on the horizon! Cells Tissues Organs 2016, 202, 227–236. [Google Scholar] [CrossRef]
- Bashkin, P.; Doctrow, S.; Klagsbrun, M.; Svahn, C.M.; Folkman, J.; Vlodavsky, I. Basic fibroblast growth factor binds to subendothelial extracellular matrix and is released by heparitinase and heparin-like molecules. Biochemistry 1989, 28, 1737–1743. [Google Scholar] [CrossRef]
- Cornelison, D.D.; Olwin, B.B.; Rudnicki, M.A.; Wold, B.J. MyoD(-/-) satellite cells in single-fiber culture are differentiation defective and MRF4 deficient. Dev. Biol. 2000, 224, 122–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wozniak, A.C.; Anderson, J.E. Nitric oxide-dependence of satellite stem cell activation and quiescence on normal skeletal muscle fibers. Dev. Dyn. 2007, 236, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Tatsumi, R.; Liu, X.; Pulido, A.; Morales, M.; Sakata, T.; Dial, S.; Hattori, A.; Ikeuchi, Y.; Allen, R.E. Satellite cell activation in stretched skeletal muscle and the role of nitric oxide and hepatocyte growth factor. Am. J. Physiol. Cell Physiol. 2006, 290, C1487–C1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatsumi, R.; Allen, R.E. Active hepatocyte growth factor is present in skeletal muscle extracellular matrix. Muscle Nerve 2004, 30, 654–658. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.E.; Wozniak, A.C. Satellite cell activation on fibers: Modeling events in vivo—An invited review. Can. J. Physiol. Pharmacol. 2004, 82, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.E.; Sheehan, S.M.; Taylor, R.G.; Kendall, T.L.; Rice, G.M. Hepatocyte growth factor activates quiescent skeletal muscle satellite cells in vitro. J. Cell Physiol. 1995, 165, 307–312. [Google Scholar] [CrossRef]
- Gal-Levi, R.; Leshem, Y.; Aoki, S.; Nakamura, T.; Halevy, O. Hepatocyte growth factor plays a dual role in regulating skeletal muscle satellite cell proliferation and differentiation. Biochim. Biophys. Acta 1998, 1402, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Tatsumi, R.; Anderson, J.E.; Nevoret, C.J.; Halevy, O.; Allen, R.E. HGF/SF is present in normal adult skeletal muscle and is capable of activating satellite cells. Dev. Biol. 1998, 194, 114–128. [Google Scholar] [CrossRef] [Green Version]
- Tatsumi, R.; Hattori, A.; Ikeuchi, Y.; Anderson, J.E.; Allen, R.E. Release of hepatocyte growth factor from mechanically stretched skeletal muscle satellite cells and role of pH and nitric oxide. Mol. Biol. Cell 2002, 13, 2909–2918. [Google Scholar] [CrossRef] [Green Version]
- Sheehan, S.M.; Allen, R.E. Skeletal muscle satellite cell proliferation in response to members of the fibroblast growth factor family and hepatocyte growth factor. J. Cell Physiol. 1999, 181, 499–506. [Google Scholar] [CrossRef]
- Cornelison, D.D.; Filla, M.S.; Stanley, H.M.; Rapraeger, A.C.; Olwin, B.B. Syndecan-3 and syndecan-4 specifically mark skeletal muscle satellite cells and are implicated in satellite cell maintenance and muscle regeneration. Dev. Biol. 2001, 239, 79–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornelison, D.D.; Wilcox-Adelman, S.A.; Goetinck, P.F.; Rauvala, H.; Rapraeger, A.C.; Olwin, B.B. Essential and separable roles for Syndecan-3 and Syndecan-4 in skeletal muscle development and regeneration. Genes Dev. 2004, 18, 2231–2236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mourikis, P.; Relaix, F. Activated muscle satellite cells chase ghosts. Cell Stem Cell 2016, 18, 160–162. [Google Scholar] [CrossRef] [PubMed]
- Webster, M.T.; Manor, U.; Lippincott-Schwartz, J.; Fan, C.M. Intravital Imaging Reveals Ghost Fibers as Architectural Units Guiding Myogenic Progenitors during Regeneration. Cell Stem Cell 2016, 18, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Serrano, A.L.; Munoz-Canoves, P. Regulation and dysregulation of fibrosis in skeletal muscle. Exp. Cell Res. 2010, 316, 3050–3058. [Google Scholar] [CrossRef]
- Yablonka-Reuveni, Z.; Seger, R.; Rivera, A.J. Fibroblast growth factor promotes recruitment of skeletal muscle satellite cells in young and old rats. J. Histochem. Cytochem. 1999, 47, 23–42. [Google Scholar] [CrossRef] [Green Version]
- Yeh, H.J.; Jin, J.J.; Wang, Y.X.; Zhou, J.Q.; Lin, X.H.; Mu, X.J.; Li, W.Y. Effect of expression of exogenous PDGF-A chain on growth and transformation of CHO cells. Shi Yan Sheng Wu Xue Bao 1989, 22, 455–465. [Google Scholar]
- Lafyatis, R.; Lechleider, R.; Roberts, A.B.; Sporn, M.B. Secretion and transcriptional regulation of transforming growth factor-beta 3 during myogenesis. Mol. Cell Biol. 1991, 11, 3795–3803. [Google Scholar] [CrossRef] [Green Version]
- Clegg, C.H.; Linkhart, T.A.; Olwin, B.B.; Hauschka, S.D. Growth factor control of skeletal muscle differentiation: Commitment to terminal differentiation occurs in G1 phase and is repressed by fibroblast growth factor. J. Cell Biol. 1987, 105, 949–956. [Google Scholar] [CrossRef]
- Allen, R.E.; Boxhorn, L.K. Regulation of skeletal muscle satellite cell proliferation and differentiation by transforming growth factor-beta, insulin-like growth factor I, and fibroblast growth factor. J. Cell Physiol. 1989, 138, 311–315. [Google Scholar] [CrossRef]
- Engert, J.C.; Berglund, E.B.; Rosenthal, N. Proliferation precedes differentiation in IGF-I-stimulated myogenesis. J. Cell Biol. 1996, 135, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Coolican, S.A.; Samuel, D.S.; Ewton, D.Z.; McWade, F.J.; Florini, J.R. The mitogenic and myogenic actions of insulin-like growth factors utilize distinct signaling pathways. J. Biol Chem. 1997, 272, 6653–6662. [Google Scholar] [CrossRef] [Green Version]
- Slater, C.R.; Schiaffino, S. Innervation of regenerating muscle. In Skeletal Muscle Repair and Regeneration; Springer: Dordrecht, The Netherlands, 2008; pp. 303–334. [Google Scholar]
- Kalhovde, J.M.; Jerkovic, R.; Sefland, I.; Cordonnier, C.; Calabria, E.; Schiaffino, S.; Lomo, T. “Fast” and “slow” muscle fibres in hindlimb muscles of adult rats regenerate from intrinsically different satellite cells. J. Physiol. 2005, 562, 847–857. [Google Scholar] [CrossRef] [PubMed]
- Christov, C.; Chretien, F.; Abou-Khalil, R.; Bassez, G.; Vallet, G.; Authier, F.J.; Bassaglia, Y.; Shinin, V.; Tajbakhsh, S.; Chazaud, B.; et al. Muscle satellite cells and endothelial cells: Close neighbors and privileged partners. Mol. Biol. Cell 2007, 18, 1397–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nillesen, S.T.; Geutjes, P.J.; Wismans, R.; Schalkwijk, J.; Daamen, W.F.; van Kuppevelt, T.H. Increased angiogenesis and blood vessel maturation in acellular collagen-heparin scaffolds containing both FGF2 and VEGF. Biomaterials 2007, 28, 1123–1131. [Google Scholar] [CrossRef]
- Baird, A. Fibroblast growth factors: Activities and significance of non-neurotrophin neurotrophic growth factors. Curr. Opin. Neurobiol. 1994, 4, 78–86. [Google Scholar] [CrossRef]
- Unsicker, K.; Reichert-Preibsch, H.; Wewetzer, K. Stimulation of neuron survival by basic FGF and CNTF is a direct effect and not mediated by non-neuronal cells: Evidence from single cell cultures. Brain Res. Dev. Brain Res. 1992, 65, 285–288. [Google Scholar] [CrossRef]
- Sakiyama-Elbert, S.E.; Hubbell, J.A. Development of fibrin derivatives for controlled release of heparin-binding growth factors. J. Control Release 2000, 65, 389–402. [Google Scholar] [CrossRef]
- Jungnickel, J.; Haase, K.; Konitzer, J.; Timmer, M.; Grothe, C. Faster nerve regeneration after sciatic nerve injury in mice over-expressing basic fibroblast growth factor. J. Neurobiol. 2006, 66, 940–948. [Google Scholar] [CrossRef]
- Schmalbruch, H. The morphology of regeneration of skeletal muscles in the rat. Tissue Cell 1976, 8, 673–692. [Google Scholar] [CrossRef]
- Ignotz, R.A.; Massague, J. Transforming growth factor-beta stimulates the expression of fibronectin and collagen and their incorporation into the extracellular matrix. J. Biol. Chem. 1986, 261, 4337–4345. [Google Scholar] [PubMed]
- Roberts, A.B.; Joyce, M.E.; Bolander, M.E.; Sporn, M.B. Transforming Growth-Factor-Beta (Tgf-Beta)—A multifunctional effector of both soft and hard tissue regeneration. Int. Congr. Ser. 1990, 925, 89–101. [Google Scholar]
- Edwards, D.R.; Murphy, G.; Reynolds, J.J.; Whitham, S.E.; Docherty, A.J.; Angel, P.; Heath, J.K. Transforming growth factor beta modulates the expression of collagenase and metalloproteinase inhibitor. EMBO J. 1987, 6, 1899–1904. [Google Scholar] [CrossRef] [PubMed]
- Cornwell, K.G.; Pins, G.D. Enhanced proliferation and migration of fibroblasts on the surface of fibroblast growth factor-2-loaded fibrin microthreads. Tissue Eng. Part A 2010, 16, 3669–3677. [Google Scholar] [CrossRef] [PubMed]
- Lehto, M.; Jarvinen, M.; Nelimarkka, O. Scar formation after skeletal muscle injury. A histological and autoradiographical study in rats. Arch. Orthop. Trauma Surg. 1986, 104, 366–370. [Google Scholar]
- Segawa, M.; Fukada, S.; Yamamoto, Y.; Yahagi, H.; Kanematsu, M.; Sato, M.; Ito, T.; Uezumi, A.; Hayashi, S.; Miyagoe-Suzuki, Y.; et al. Suppression of macrophage functions impairs skeletal muscle regeneration with severe fibrosis. Exp. Cell Res. 2008, 314, 3232–3244. [Google Scholar] [CrossRef]
- Kharraz, Y.; Guerra, J.; Mann, C.J.; Serrano, A.L.; Munoz-Canoves, P. Macrophage plasticity and the role of inflammation in skeletal muscle repair. Mediat. Inflamm. 2013, 2013, 491497. [Google Scholar] [CrossRef] [Green Version]
- Watt, D.J.; Morgan, J.E.; Clifford, M.A.; Partridge, T.A. The movement of muscle precursor cells between adjacent regenerating muscles in the mouse. Anat. Embryol. 1987, 175, 527–536. [Google Scholar] [CrossRef]
- Bian, W.; Bursac, N. Tissue engineering of functional skeletal muscle: Challenges and recent advances. IEEE Eng. Med. Biol. Mag. 2008, 27, 109–113. [Google Scholar] [CrossRef] [Green Version]
- Turner, N.J.; Badylak, S.F. Regeneration of skeletal muscle. Cell Tissue Res. 2012, 347, 759–774. [Google Scholar] [CrossRef]
- Corona, B.T.; Flanagan, K.E.; Brininger, C.M.; Goldman, S.M.; Call, J.A.; Greising, S.M. Impact of volumetric muscle loss injury on persistent motoneuron axotomy. Muscle Nerve 2018, 57, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Chakkalakal, J.V. The composition, development, and regeneration of neuromuscular junctions. Curr. Top. Dev. Biol. 2018, 126, 99–124. [Google Scholar] [CrossRef] [PubMed]
- Grounds, M.D. Towards understanding skeletal muscle regeneration. Pathol. Res. Pract. 1991, 187, 1–22. [Google Scholar] [CrossRef]
- Nguyen, F.; Guigand, L.; Goubault-Leroux, I.; Wyers, M.; Cherel, Y. Microvessel density in muscles of dogs with golden retriever muscular dystrophy. Neuromuscul. Disord. 2005, 15, 154–163. [Google Scholar] [CrossRef]
- Juhas, M.; Bursac, N. Engineering skeletal muscle repair. Curr. Opin. Biotechnol. 2013, 24, 880–886. [Google Scholar] [CrossRef] [Green Version]
- Langer, R.; Vacanti, J.P. Tissue engineering. Science 1993, 260, 920–926. [Google Scholar] [CrossRef] [Green Version]
- Nakayama, K.H.; Shayan, M.; Huang, N.F. Engineering biomimetic materials for skeletal muscle repair and regeneration. Adv. Healthc. Mater. 2019, 8, e1801168. [Google Scholar] [CrossRef]
- Stern-Straeter, J.; Riedel, F.; Bran, G.; Hormann, K.; Goessler, U.R. Advances in skeletal muscle tissue engineering. In Vivo 2007, 21, 435–444. [Google Scholar]
- Sicari, B.M.; Londono, R.; Badylak, S.F. Strategies for skeletal muscle tissue engineering: Seed vs. soil. J. Mater. Chem. B 2015, 3, 7881–7895. [Google Scholar] [CrossRef]
- Tedesco, F.S.; Cossu, G. Stem cell therapies for muscle disorders. Curr. Opin. Neurol. 2012, 25, 597–603. [Google Scholar] [CrossRef]
- Corona, B.T.; Wenke, J.C.; Ward, C.L. Pathophysiology of volumetric muscle loss injury. Cells Tissues Organs 2016, 202, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, D.; Waldman, S.D.; Li, S. From in vitro to in situ tissue engineering. Ann. Biomed. Eng. 2014, 42, 1537–1545. [Google Scholar] [CrossRef] [PubMed]
- Sicari, B.M.; Rubin, J.P.; Dearth, C.L.; Wolf, M.T.; Ambrosio, F.; Boninger, M.; Turner, N.J.; Weber, D.J.; Simpson, T.W.; Wyse, A.; et al. An acellular biologic scaffold promotes skeletal muscle formation in mice and humans with volumetric muscle loss. Sci. Transl. Med. 2014, 6, 234ra58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, R.; Ma, P.X.; Guo, B. Conductive biomaterials for muscle tissue engineering. Biomaterials 2020, 229, 119584. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.C.; Sun, Y.C.; Chen, Y.H. Electrically conductive nanofibers with highly oriented structures and their potential application in skeletal muscle tissue engineering. Acta Biomater. 2013, 9, 5562–5572. [Google Scholar] [CrossRef] [PubMed]
- Ku, S.H.; Lee, S.H.; Park, C.B. Synergic effects of nanofiber alignment and electroactivity on myoblast differentiation. Biomaterials 2012, 33, 6098–6104. [Google Scholar] [CrossRef] [PubMed]
- Engler, A.J.; Griffin, M.A.; Sen, S.; Bonnemann, C.G.; Sweeney, H.L.; Discher, D.E. Myotubes differentiate optimally on substrates with tissue-like stiffness: Pathological implications for soft or stiff microenvironments. J. Cell Biol. 2004, 166, 877–887. [Google Scholar] [CrossRef] [Green Version]
- Lanza, R.; Langer, R.; Vacanti, J.P.; Atala, A. Principles of Tissue Engineering; Academic Press: Cambridge, MA, USA, 2020. [Google Scholar]
- Cezar, C.A.; Mooney, D.J. Biomaterial-based delivery for skeletal muscle repair. Adv. Drug Deliv. Rev. 2015, 84, 188–197. [Google Scholar] [CrossRef] [Green Version]
- Badylak, S.F. The extracellular matrix as a biologic scaffold material. Biomaterials 2007, 28, 3587–3593. [Google Scholar] [CrossRef]
- Badylak, S.F.; Freytes, D.O.; Gilbert, T.W. Extracellular matrix as a biological scaffold material: Structure and function. Acta Biomater. 2009, 5, 1–13. [Google Scholar] [CrossRef]
- Garg, K.; Ward, C.L.; Corona, B.T. Asynchronous inflammation and myogenic cell migration limit muscle tissue regeneration mediated by a cellular scaffolds. Inflamm. Cell Signal. 2014, 1. [Google Scholar] [CrossRef]
- Wolf, M.T.; Dearth, C.L.; Sonnenberg, S.B.; Loboa, E.G.; Badylak, S.F. Naturally derived and synthetic scaffolds for skeletal muscle reconstruction. Adv. Drug Deliv. Rev. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- VoytikHarbin, S.L.; Brightman, A.O.; Kraine, M.R.; Waisner, B.; Badylak, S.F. Identification of extractable growth factors from small intestinal submucosa. J. Cell. Biochem. 1997, 67, 478–491. [Google Scholar] [CrossRef]
- Brown, B.; Lindberg, K.; Reing, J.; Stolz, D.B.; Badylak, S.F. The basement membrane component of biologic scaffolds derived from extracellular matrix. Tissue Eng. 2006, 12, 519–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcal, H.; Ahmed, T.; Badylak, S.F.; Tottey, S.; Foster, L.J.R. A comprehensive protein expression profile of extracellular matrix biomaterial derived from porcine urinary bladder. Regen. Med. 2012, 7, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Wolf, M.T.; Daly, K.A.; Reing, J.E.; Badylak, S.F. Biologic scaffold composed of skeletal muscle extracellular matrix. Biomaterials 2012, 33, 2916–2925. [Google Scholar] [CrossRef] [Green Version]
- Brown, B.N.; Freund, J.M.; Han, L.; Rubin, J.P.; Reing, J.E.; Jeffries, E.M.; Wolf, M.T.; Tottey, S.; Barnes, C.A.; Ratner, B.D.; et al. Comparison of three methods for the derivation of a biologic scaffold composed of adipose tissue extracellular matrix. Tissue Eng. Part C Methods 2011, 17, 411–421. [Google Scholar] [CrossRef]
- Faulk, D.M.; Carruthers, C.A.; Warner, H.J.; Kramer, C.R.; Reing, J.E.; Zhang, L.; D’Amore, A.; Badylak, S.F. The effect of detergents on the basement membrane complex of a biologic scaffold material. Acta Biomater. 2014, 10, 183–193. [Google Scholar] [CrossRef] [Green Version]
- Reing, J.E.; Brown, B.N.; Daly, K.A.; Freund, J.M.; Gilbert, T.W.; Hsiong, S.X.; Huber, A.; Kullas, K.E.; Tottey, S.; Wolf, M.T.; et al. The effects of processing methods upon mechanical and biologic properties of porcine dermal extracellular matrix scaffolds. Biomaterials 2010, 31, 8626–8633. [Google Scholar] [CrossRef] [Green Version]
- Turner, N.J.; Yates, A.J.; Weber, D.J.; Qureshi, I.R.; Stolz, D.B.; Gilbert, T.W.; Badylak, S.F. Xenogeneic Extracellular Matrix as an Inductive Scaffold for Regeneration of a Functioning Musculotendinous Junction. Tissue Eng. Part A 2010, 16, 3309–3317. [Google Scholar] [CrossRef]
- Valentin, J.E.; Turner, N.J.; Gilbert, T.W.; Badylak, S.F. Functional skeletal muscle formation with a biologic scaffold. Biomaterials 2010, 31, 7475–7484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merritt, E.K.; Hammers, D.W.; Tierney, M.; Suggs, L.J.; Walters, T.J.; Farrar, R.P. Functional assessment of skeletal muscle regeneration utilizing homologous extracellular matrix as scaffolding. Tissue Eng. Part A 2010, 16, 1395–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sicari, B.M.; Agrawal, V.; Siu, B.F.; Medberry, C.J.; Dearth, C.L.; Turner, N.J.; Badylak, S.F. A Murine model of volumetric muscle loss and a regenerative medicine approach for tissue replacement. Tissue Eng. Part A 2012, 18, 1941–1948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corona, B.T.; Wu, X.; Ward, C.L.; McDaniel, J.S.; Rathbone, C.R.; Walters, T.J. The promotion of a functional fibrosis in skeletal muscle with volumetric muscle loss injury following the transplantation of muscle-ECM. Biomaterials 2013, 34, 3324–3335. [Google Scholar] [CrossRef] [PubMed]
- Garg, K.; Ward, C.L.; Rathbone, C.R.; Corona, B.T. Transplantation of devitalized muscle scaffolds is insufficient for appreciable de novo muscle fiber regeneration after volumetric muscle loss injury. Cell Tissue Res. 2014, 358, 857–873. [Google Scholar] [CrossRef] [PubMed]
- Aurora, A.; Roe, J.L.; Corona, B.T.; Walters, T.J. An acellular biologic scaffold does not regenerate appreciable de novo muscle tissue in rat models of volumetric muscle loss injury. Biomaterials 2015, 67, 393–407. [Google Scholar] [CrossRef]
- Greising, S.M.; Rivera, J.C.; Goldman, S.M.; Watts, A.; Aguilar, C.A.; Corona, B.T. Unwavering Pathobiology of Volumetric Muscle Loss Injury. Sci. Rep. 2017, 7, 13179. [Google Scholar] [CrossRef] [Green Version]
- Han, N.; Yabroudi, M.A.; Stearns-Reider, K.; Helkowski, W.; Sicari, B.M.; Rubin, J.P.; Badylak, S.F.; Boninger, M.L.; Ambrosio, F. electrodiagnostic evaluation of individuals implanted with extracellular matrix for the treatment of volumetric muscle injury: Case Series. Phys. Ther. 2016, 96, 540–549. [Google Scholar] [CrossRef] [Green Version]
- Dziki, J.; Badylak, S.; Yabroudi, M.; Sicari, B.; Ambrosio, F.; Stearns, K.; Turner, N.; Wyse, A.; Boninger, M.L.; Brown, E.H.; et al. An acellular biologic scaffold treatment for volumetric muscle loss: Results of a 13-patient cohort study. Nat. Regen. Med. 2016, 1, 16008. [Google Scholar] [CrossRef]
- Hill, E.; Boontheekul, T.; Mooney, D.J. Designing scaffolds to enhance transplanted myoblast survival and migration. Tissue Eng. 2006, 12, 1295–1304. [Google Scholar] [CrossRef] [Green Version]
- DeQuach, J.A.; Lin, J.E.; Cam, C.; Hu, D.; Salvatore, M.A.; Sheikh, F.; Christman, K.L. Injectable skeletal muscle matrix hydrogel promotes neovascularization and muscle cell infiltration in a hindlimb ischemia model. Eur. Cells Mater. 2012, 23, 400. [Google Scholar] [CrossRef] [PubMed]
- Marcinczyk, M.; Dunn, A.; Haas, G.; Madsen, J.; Scheidt, R.; Patel, K.; Talovic, M.; Garg, K. The Effect of Laminin-111 Hydrogels on Muscle Regeneration in a Murine Model of Injury. Tissue Eng. Part A 2019, 25, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Kin, S.; Hagiwara, A.; Nakase, Y.; Kuriu, Y.; Nakashima, S.; Yoshikawa, T.; Sakakura, C.; Otsuji, E.; Nakamura, T.; Yamagishi, H. Regeneration of skeletal muscle using in situ tissue engineering on an acellular collagen sponge scaffold in a rabbit model. ASAIO J. 2007, 53, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Haas, G.J.; Dunn, A.J.; Marcinczyk, M.; Talovic, M.; Schwartz, M.; Scheidt, R.; Patel, A.D.; Hixon, K.R.; Elmashhady, H.; McBride-Gagyi, S.H. Biomimetic sponges for regeneration of skeletal muscle following trauma. J. Biomed. Mater. Res. A 2019, 107, 92–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, Y.M.; Atala, A.; Yoo, J.J.; Lee, S.J. In situ regeneration of skeletal muscle tissue through host cell recruitment. Acta Biomater. 2014, 10, 4332–4339. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.F.; Lee, R.J.; Li, S. Engineering of aligned skeletal muscle by micropatterning. Am. J. Transl. Res. 2010, 2, 43–55. [Google Scholar]
- Huang, N.F.; Patel, S.; Thakar, R.G.; Wu, J.; Hsiao, B.S.; Chu, B.; Lee, R.J.; Li, S. Myotube assembly on nanofibrous and micropatterned polymers. Nano Lett. 2006, 6, 537–542. [Google Scholar] [CrossRef]
- Charest, J.L.; Garcia, A.J.; King, W.P. Myoblast alignment and differentiation on cell culture substrates with microscale topography and model chemistries. Biomaterials 2007, 28, 2202–2210. [Google Scholar] [CrossRef]
- Wang, P.Y.; Yu, H.T.; Tsai, W.B. Modulation of alignment and differentiation of skeletal myoblasts by submicron ridges/grooves surface structure. Biotechnol. Bioeng. 2010, 106, 285–294. [Google Scholar] [CrossRef]
- Monge, C.; Ren, K.; Berton, K.; Guillot, R.; Peyrade, D.; Picart, C. Engineering muscle tissues on microstructured polyelectrolyte multilayer films. Tissue Eng. Part A 2012, 18, 1664–1676. [Google Scholar] [CrossRef] [Green Version]
- Flemming, R.G.; Murphy, C.J.; Abrams, G.A.; Goodman, S.L.; Nealey, P.F. Effects of synthetic micro- and nano-structures surfaces on cell behavior. Biomaterials 1999, 20, 573–588. [Google Scholar] [CrossRef]
- Zhao, Y.; Zeng, H.; Nam, J.; Agarwal, S. Fabrication of skeletal muscle constructs by topographic activation of cell alignment. Biotechnol. Bioeng. 2009, 102, 624–631. [Google Scholar] [CrossRef] [Green Version]
- Bian, W.; Liau, B.; Badie, N.; Bursac, N. Mesoscopic hydrogel molding to control the 3D geometry of bioartificial muscle tissues. Nat. Protoc. 2009, 4, 1522–1534. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.S.; Lee, S.J.; Christ, G.J.; Atala, A.; Yoo, J.J. The influence of electrospun aligned poly(epsilon-caprolactone)/collagen nanofiber meshes on the formation of self-aligned skeletal muscle myotubes. Biomaterials 2008, 29, 2899–2906. [Google Scholar] [CrossRef]
- Li, W.; Mauck, R.L.; Cooper, J.A.; Yuan, X.; Tuan, R.S. Engineering controllable anisotropy in electrospun biodegradable nanofibrous scaffolds for musculoskeletal tissue engineering. J. Biomech. 2007, 40, 1686–1693. [Google Scholar] [CrossRef] [Green Version]
- Aviss, K.J.; Gough, J.E.; Downes, S. Aligned electrospun polymer fibres for skeletal muscle regeneration. Eur. Cells Mater. 2010, 19, 193–204. [Google Scholar] [CrossRef]
- Zhang, S.; Liu, X.; Barreto-Ortiz, S.F.; Yu, Y.; Ginn, B.P.; DeSantis, N.A.; Hutton, D.L.; Grayson, W.L.; Cui, F.Z.; Korgel, B.A.; et al. Creating polymer hydrogel microfibres with internal alignment via electrical and mechanical stretching. Biomaterials 2014, 35, 3243–3251. [Google Scholar] [CrossRef] [Green Version]
- Cornwell, K.G.; Pins, G.D. Discrete crosslinked fibrin microthread scaffolds for tissue regeneration. J. Biomed. Mater. Res. A 2007, 82, 104–112. [Google Scholar] [CrossRef]
- O’Brien, M.P.; Carnes, M.E.; Page, R.L.; Gaudette, G.R.; Pins, G.D. Designing biopolymer microthreads for tissue engineering and regenerative medicine. Curr. Stem Cell Rep. 2016, 2, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Pins, G.D.; Silver, F.H. A self-assembled collagen scaffold suitable for use in soft and hard tissue replacement. Mater. Sci. Eng. C 1995, 3, 101–107. [Google Scholar] [CrossRef]
- Jana, S.; Cooper, A.; Zhang, M. Chitosan scaffolds with unidirectional microtubular pores for large skeletal myotube generation. Adv. Healthc. Mater. 2013, 2, 557–561. [Google Scholar] [CrossRef]
- Kroehne, V.; Heschel, I.; Schugner, F.; Lasrich, D.; Bartsch, J.W.; Jockusch, H. Use of a novel collagen matrix with oriented pore structure for muscle cell differentiation in cell culture and in grafts. J. Cell. Mol. Med. 2008, 12, 1640–1648. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.X.; Zhang, R. Microtubular architecture of biodegradable polymer scaffolds. J. Biomed. Mater. Res. 2001, 56, 469–477. [Google Scholar] [CrossRef] [Green Version]
- Jana, S.; Levengood, S.K.; Zhang, M. Anisotropic materials for skeletal-muscle-tissue engineering. Adv. Mater. 2016, 28, 10588–10612. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, K.H.; Alcazar, C.; Yang, G.; Quarta, M.; Paine, P.; Doan, L.; Davies, A.; Rando, T.A.; Huang, N.F. Rehabilitative exercise and spatially patterned nanofibrillar scaffolds enhance vascularization and innervation following volumetric muscle loss. NPJ Regen. Med. 2018, 3, 16. [Google Scholar] [CrossRef]
- Simons, M.; Annex, B.H.; Laham, R.J.; Kleiman, N.; Henry, T.; Dauerman, H.; Udelson, J.E.; Gervino, E.V.; Pike, M.; Whitehouse, M.J.; et al. Pharmacological treatment of coronary artery disease with recombinant fibroblast growth factor-2: Double-blind, randomized, controlled clinical trial. Circulation 2002, 105, 788–793. [Google Scholar] [CrossRef]
- Simons, M.; Ware, J.A. Therapeutic angiogenesis in cardiovascular disease. Nat. Rev. Drug Discov. 2003, 2, 863–871. [Google Scholar] [CrossRef]
- Henry, T.D.; Annex, B.H.; McKendall, G.R.; Azrin, M.A.; Lopez, J.J.; Giordano, F.J.; Shah, P.K.; Willerson, J.T.; Benza, R.L.; Berman, D.S.; et al. The VIVA trial: Vascular endothelial growth factor in Ischemia for Vascular Angiogenesis. Circulation 2003, 107, 1359–1365. [Google Scholar] [CrossRef]
- Simons, M.; Bonow, R.O.; Chronos, N.A.; Cohen, D.J.; Giordano, F.J.; Hammond, H.K.; Laham, R.J.; Li, W.; Pike, M.; Sellke, F.W.; et al. Clinical trials in coronary angiogenesis: Issues, problems, consensus: An expert panel summary. Circulation 2000, 102, E73–E86. [Google Scholar] [CrossRef]
- Lee, K.; Silva, E.A.; Mooney, D.J. Growth factor delivery-based tissue engineering: General approaches and a review of recent developments. J. R. Soc. Interface 2011, 8, 153–170. [Google Scholar] [CrossRef] [Green Version]
- Silva, E.A.; Mooney, D.J. Spatiotemporal control of vascular endothelial growth factor delivery from injectable hydrogels enhances angiogenesis. J. Thromb. Haemost. 2007, 5, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Shvartsman, D.; Storrie-White, H.; Lee, K.; Kearney, C.; Brudno, Y.; Ho, N.; Cezar, C.; McCann, C.; Anderson, E.; Koullias, J.; et al. Sustained delivery of VEGF maintains innervation and promotes reperfusion in ischemic skeletal muscles via NGF/GDNF signaling. Mol. Ther. J. Am. Soc. Gene Ther. 2014, 22, 1243–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Bhang, S.H.; Park, H.; Kim, B.S.; Lee, K.Y. Active blood vessel formation in the ischemic hindlimb mouse model using a microsphere/hydrogel combination system. Pharm. Res. Dordr. 2010, 27, 767–774. [Google Scholar] [CrossRef] [PubMed]
- Frey, S.P.; Jansen, H.; Raschke, M.J.; Meffert, R.H.; Ochman, S. VEGF improves skeletal muscle regeneration after acute trauma and reconstruction of the limb in a rabbit model. Clin. Orthop. Relat. Res. 2012, 470, 3607–3614. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammers, D.W.; Sarathy, A.; Pham, C.B.; Drinnan, C.T.; Farrar, R.P.; Suggs, L.J. Controlled release of IGF-1 from a biodegradable matrix improves functional recovery of skeletal muscle from ischemia/reperfusion. Biotechnol. Bioeng. 2011, 109, 1051–1059. [Google Scholar] [CrossRef] [Green Version]
- Doi, K.; Ikeda, T.; Marui, A.; Kushibiki, T.; Arai, Y.; Hirose, K.; Soga, Y.; Iwakura, A.; Ueyama, K.; Yamahara, K.; et al. Enhanced angiogenesis by gelatin hydrogels incorporating basic fibroblast growth factor in rabbit model of hind limb ischemia. Heart Vessel. 2007, 22, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Layman, H.; Spiga, M.G.; Brooks, T.; Pham, S.; Webster, K.A.; Andreopoulos, F.M. The effect of the controlled release of basic fibroblast growth factor from ionic gelatin-based hydrogels on angiogenesis in a murine critical limb ischemic model. Biomaterials 2007, 28, 2646–2654. [Google Scholar] [CrossRef] [Green Version]
- Yasuda, Y.; Koyama, H.; Tabata, Y.; Fujihara, Y.; Oba, M.; Uchinuma, E.; Takato, T. Controlled delivery of bFGF remodeled vascular network in muscle flap and increased perfusion capacity via minor pedicle. J. Surg. Res. 2008, 147, 132–137. [Google Scholar] [CrossRef]
- Grasman, J.M.; Do, D.M.; Page, R.L.; Pins, G.D. Rapid release of growth factors regenerates force output in volumetric muscle loss injuries. Biomaterials 2015, 72, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Borselli, C.; Storrie, H.; Benesch-Lee, F.; Shvartsman, D.; Cezar, C.; Lichtman, J.W.; Vandenburgh, H.H.; Mooney, D.J. Functional muscle regeneration with combined delivery of angiogenesis and myogenesis factors. Proc. Natl. Acad. Sci. USA 2010, 107, 3287–3292. [Google Scholar] [CrossRef] [Green Version]
- Rybalko, V.Y.; Pham, C.B.; Hsieh, P.L.; Hammers, D.W.; Merscham-Banda, M.; Suggs, L.J.; Farrar, R.P. Controlled delivery of SDF-1alpha and IGF-1: CXCR4(+) cell recruitment and functional skeletal muscle recovery. Biomater. Sci. 2015, 3, 1475–1486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, M.; Tabata, Y. Enhanced angiogenesis by multiple release of platelet-rich plasma contents and basic fibroblast growth factor from gelatin hydrogels. Acta Biomater. 2012, 8, 1792–1801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passipieri, J.A.; Christ, G.J. The potential of combination therapeutics for more complete repair of volumetric muscle loss injuries: The role of exogenous growth factors and/or progenitor cells in implantable skeletal muscle tissue engineering technologies. Cells Tissues Organs 2016, 202, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.M.; Zhang, M.; Wu, Z.F. Toward delivery of multiple growth factors in tissue engineering. Biomaterials 2010, 31, 6279–6308. [Google Scholar] [CrossRef]
- Stilhano, R.S.; Madrigal, J.L.; Wong, K.; Williams, P.A.; Martin, P.K.; Yamaguchi, F.S.; Samoto, V.Y.; Han, S.W.; Silva, E.A. Injectable alginate hydrogel for enhanced spatiotemporal control of lentivector delivery in murine skeletal muscle. J. Control Release 2016, 237, 42–49. [Google Scholar] [CrossRef]
- Falco, E.E.; Wang, M.O.; Thompson, J.A.; Chetta, J.M.; Yoon, D.M.; Li, E.Z.; Kulkami, M.M.; Shah, S.; Pandit, A.; Roth, J.S.; et al. Porous EH and EH-PEG scaffolds as gene delivery vehicles to skeletal muscle. Pharm. Res. 2011, 28, 1306–1316. [Google Scholar] [CrossRef]
- Doukas, J.; Blease, K.; Craig, D.; Ma, C.; Chandler, L.A.; Sosnowski, B.A.; Pierce, G.F. Delivery of FGF genes to wound repair cells enhances arteriogenesis and myogenesis in skeletal muscle. Mol. Ther. J. Am. Soc. Gene Ther. 2002, 5, 517–527. [Google Scholar] [CrossRef]
- Kasahara, H.; Tanaka, E.; Fukuyama, N.; Sato, E.; Sakamoto, H.; Tabata, Y.; Ando, K.; Iseki, H.; Shinozaki, Y.; Kimura, K.; et al. Biodegradable gelatin hydrogel potentiates the angiogenic effect of fibroblast growth factor 4 plasmid in rabbit hindlimb ischemia. J. Am. Coll. Cardiol. 2003, 41, 1056–1062. [Google Scholar] [CrossRef] [Green Version]
- Zaitseva, T.S.; Alcazar, C.; Zamani, M.; Hou, L.; Sawamura, S.; Yakubov, E.; Hopkins, M.; Woo, Y.J.; Paukshto, M.V.; Huang, N.F. Aligned Nanofibrillar scaffolds for controlled delivery of modified mRNA. Tissue Eng. Part A 2019, 25, 121–130. [Google Scholar] [CrossRef]
- Jung, D.W.; Williams, D.R. Reawakening atlas: Chemical approaches to repair or replace dysfunctional musculature. ACS Chem. Biol. 2012, 7, 1773–1790. [Google Scholar] [CrossRef]
- San Emeterio, C.L.; Olingy, C.E.; Chu, Y.; Botchwey, E.A. Selective recruitment of non-classical monocytes promotes skeletal muscle repair. Biomaterials 2017, 117, 32–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awojoodu, A.O.; Ogle, M.E.; Sefcik, L.S.; Bowers, D.T.; Martin, K.; Brayman, K.L.; Lynch, K.R.; Peirce-Cottler, S.M.; Botchwey, E. Sphingosine 1-phosphate receptor 3 regulates recruitment of anti-inflammatory monocytes to microvessels during implant arteriogenesis. Proc. Natl. Acad. Sci. USA 2013, 110, 13785–13790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, L.; Yang, G.; Tang, S.; Alcazar, C.; Joshi, P.; Strassberg, Z.; Kim, M.; Kawamura, M.; Woo, Y.J.; Shrager, J.; et al. Small molecule derived from carboxyethylpyrrole protein adducts promotes angiogenesis in a mouse model of peripheral arterial disease. J. Am. Heart Assoc. 2018, 7, e009234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corona, B.T.; Greising, S.M. Challenges to acellular biological scaffold mediated skeletal muscle tissue regeneration. Biomaterials 2016, 104, 238–246. [Google Scholar] [CrossRef]
- Smythe, G.M.; Hodgetts, S.I.; Grounds, M.D. Problems and solutions in myoblast transfer therapy. J. Cell. Mol. Med. 2001, 5, 33–47. [Google Scholar] [CrossRef]
- Boldrin, L.; Elvassore, N.; Malerba, A.; Flaibani, M.; Cimetta, E.; Piccoli, M.; Baroni, M.D.; Gazzola, M.V.; Messina, C.; Gamba, P.; et al. Satellite cells delivered by micro-patterned scaffolds: A new strategy for cell transplantation in muscle diseases. Tissue Eng. 2007, 13, 253–262. [Google Scholar] [CrossRef]
- Sicari, B.M.; Dearth, C.L.; Badylak, S.F. Tissue engineering and regenerative medicine approaches to enhance the functional response to skeletal muscle injury. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2014, 297, 51–64. [Google Scholar] [CrossRef]
- Pantelic, M.N.; Larkin, L.M. Stem cells for skeletal muscle tissue engineering. Tissue Eng. Part B Rev. 2018, 24, 373–391. [Google Scholar] [CrossRef]
- McCullagh, K.J.; Perlingeiro, R.C. Coaxing stem cells for skeletal muscle repair. Adv. Drug Deliv. Rev. 2015, 84, 198–207. [Google Scholar] [CrossRef] [Green Version]
- Montarras, D.; Morgan, J.; Collins, C.; Relaix, F.; Zaffran, S.; Cumano, A.; Partridge, T.; Buckingham, M. Direct isolation of satellite cells for skeletal muscle regeneration. Science 2005, 309, 2064–2067. [Google Scholar] [CrossRef]
- Swijnenburg, R.-J.; Schrepfer, S.; Govaert, J.A.; Cao, F.; Ransohoff, K.; Sheikh, A.Y.; Haddad, M.; Connolly, A.J.; Davis, M.M.; Robbins, R.C. Immunosuppressive therapy mitigates immunological rejection of human embryonic stem cell xenografts. Proc. Natl. Acad. Sci. USA 2008, 105, 12991–12996. [Google Scholar] [CrossRef] [Green Version]
- Van Wachem, P.B.; Brouwer, L.A.; van Luyn, M.J. Absence of muscle regeneration after implantation of a collagen matrix seeded with myoblasts. Biomaterials 1999, 20, 419–426. [Google Scholar] [CrossRef]
- Beier, J.P.; Stern-Straeter, J.; Foerster, V.T.; Kneser, U.; Stark, G.B.; Bach, A.D. Tissue engineering of injectable muscle: Three-dimensional myoblast-fibrin injection in the syngeneic rat animal model. Plast Reconstr. Surg. 2006, 118, 1113–1121. [Google Scholar] [CrossRef]
- Matthias, N.; Hunt, S.D.; Wu, J.B.; Lo, J.; Callahan, L.A.S.; Li, Y.; Huard, J.; Darabi, R. Volumetric muscle loss injury repair using in situ fibrin gel cast seeded with muscle-derived stem cells (MDSCs). Stem Cell Res. 2018, 27, 65–73. [Google Scholar] [CrossRef]
- Rossi, C.A.; Flaibani, M.; Blaauw, B.; Pozzobon, M.; Figallo, E.; Reggiani, C.; Vitiello, L.; Elvassore, N.; De Coppi, P. In vivo tissue engineering of functional skeletal muscle by freshly isolated satellite cells embedded in a photopolymerizable hydrogel. FASEB J. 2011, 25, 2296–2304. [Google Scholar] [CrossRef]
- Kim, J.H.; Ko, I.K.; Atala, A.; Yoo, J.J. Progressive Muscle Cell Delivery as a Solution for Volumetric Muscle Defect Repair. Sci. Rep. UK 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Ward, C.L.; Ji, L.; Corona, B.T. An autologous muscle tissue expansion approach for the treatment of volumetric muscle loss. Biores Open Access 2015, 4, 198–208. [Google Scholar] [CrossRef]
- Goldman, S.M.; Henderson, B.E.P.; Walters, T.J.; Corona, B.T. Co-delivery of a laminin-111 supplemented hyaluronic acid based hydrogel with minced muscle graft in the treatment of volumetric muscle loss injury. PLoS ONE 2018, 13, e0191245. [Google Scholar] [CrossRef] [Green Version]
- Pereira, T.; Armada-da Silva, P.A.S.; Amorim, I.; Rema, A.; Caseiro, A.R.; Gartner, A.; Rodrigues, M.; Lopes, M.A.; Bartolo, P.J.; Santos, J.D.; et al. effects of human mesenchymal stem cells isolated from Wharton’s jelly of the umbilical cord and conditioned media on skeletal muscle regeneration using a myectomy model. Stem Cells Int. 2014, 2014. [Google Scholar] [CrossRef]
- Aurora, A.; Wrice, N.; Walters, T.J.; Christy, R.J.; Natesan, S. A PEGylated platelet free plasma hydrogel based composite scaffold enables stable vascularization and targeted cell delivery for volumetric muscle loss. Acta Biomater. 2018, 65, 150–162. [Google Scholar] [CrossRef]
- Huang, H.; Liu, J.; Hao, H.; Chen, D.; Zhizhong, L.; Li, M.; Song, H.; Xiang, R.; Jiang, C.; Fu, X.; et al. Preferred M2 Polarization by ASC-Based hydrogel accelerated angiogenesis and Myogenesis in volumetric muscle loss rats. Stem Cells Int. 2017, 2017, 2896874. [Google Scholar] [CrossRef] [Green Version]
- Goldman, S.M.; Henderson, B.E.P.; Corona, B.T. Evaluation of bone marrow mononuclear cells as an adjunct therapy to minced muscle graft for the treatment of volumetric muscle loss injuries. Stem Cell Res. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Lesman, A.; Koffler, J.; Atlas, R.; Blinder, Y.J.; Kam, Z.; Levenberg, S. Engineering vessel-like networks within multicellular fibrin-based constructs. Biomaterials 2011, 32, 7856–7869. [Google Scholar] [CrossRef]
- Merritt, E.K.; Cannon, M.V.; Hammers, D.W.; Le, L.N.; Gokhale, R.; Sarathy, A.; Song, T.J.; Tierney, M.T.; Suggs, L.J.; Walters, T.J.; et al. Repair of traumatic skeletal muscle injury with bone-marrow-derived mesenchymal stem cells seeded on extracellular matrix. Tissue Eng. Part A 2010, 16, 2871–2881. [Google Scholar] [CrossRef] [Green Version]
- Conconi, M.T.; De Coppi, P.; Bellini, S.; Zara, G.; Sabatti, M.; Marzaro, M.; Zanon, G.F.; Gamba, P.G.; Parnigotto, P.P.; Nussdorfer, G.G. Homologous muscle acellular matrix seeded with autologous myoblasts as a tissue-engineering approach to abdominal wall-defect repair. Biomaterials 2005, 26, 2567–2574. [Google Scholar] [CrossRef]
- De Coppi, P.; Bellini, S.; Conconi, M.T.; Sabatti, M.; Simonato, E.; Gamba, P.G.; Nussdorfer, G.G.; Parnigotto, P.P. Myoblast-acellular skeletal muscle matrix constructs guarantee a long-term repair of experimental full-thickness abdominal wall defects. Tissue Eng. 2006, 12, 1929–1936. [Google Scholar] [CrossRef]
- Kasukonis, B.; Kim, J.; Brown, L.; Jones, J.; Ahmadi, S.; Washington, T.; Wolchok, J. Codelivery of Infusion Decellularized Skeletal Muscle with Minced Muscle Autografts Improved Recovery from Volumetric Muscle Loss Injury in a Rat Model. Tissue Eng. Part A 2016, 22, 1151–1163. [Google Scholar] [CrossRef] [Green Version]
- Goldman, S.M.; Corona, B.T. Co-delivery of micronized urinary bladder matrix damps regenerative capacity of minced muscle grafts in the treatment of volumetric muscle loss injuries. PLoS ONE 2017, 12, e0186593. [Google Scholar] [CrossRef] [Green Version]
- Quarta, M.; Cromie, M.; Chacon, R.; Blonigan, J.; Garcia, V.; Akimenko, I.; Hamer, M.; Paine, P.; Stok, M.; Shrager, J.B.; et al. Bioengineered constructs combined with exercise enhance stem cell-mediated treatment of volumetric muscle loss. Nat. Commun. 2017, 8, 15613. [Google Scholar] [CrossRef]
- Quarta, M.; Cromie Lear, M.J.; Blonigan, J.; Paine, P.; Chacon, R.; Rando, T.A. Biomechanics show stem cell necessity for effective treatment of volumetric muscle loss using bioengineered constructs. NPJ Regen. Med. 2018, 3, 18. [Google Scholar] [CrossRef]
- Qiu, X.; Liu, S.; Zhang, H.; Zhu, B.; Su, Y.; Zheng, C.; Tian, R.; Wang, M.; Kuang, H.; Zhao, X.; et al. Mesenchymal stem cells and extracellular matrix scaffold promote muscle regeneration by synergistically regulating macrophage polarization toward the M2 phenotype. Stem Cell Res. 2018, 9, 88. [Google Scholar] [CrossRef] [PubMed]
- Page, R.L.; Malcuit, C.; Vilner, L.; Vojtic, I.; Shaw, S.; Hedblom, E.; Hu, J.; Pins, G.D.; Rolle, M.W.; Dominko, T. Restoration of Skeletal Muscle Defects with Adult Human Cells Delivered on Fibrin Microthreads. Tissue Eng. Part A 2011, 17, 2629–2640. [Google Scholar] [CrossRef] [PubMed]
- Greising, S.M.; Corona, B.T.; McGann, C.; Frankum, J.K.; Warren, G.L. Therapeutic Approaches for Volumetric Muscle Loss Injury: A Systematic Review and Meta-Analysis. Tissue Eng. Part B Rev. 2019, 25, 510–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagiwara, K.; Chen, G.; Kawazoe, N.; Tabata, Y.; Komuro, H. Promotion of muscle regeneration by myoblast transplantation combined with the controlled and sustained release of bFGFcpr. J. Tissue Eng. Regen. Med. 2016, 10, 325–333. [Google Scholar] [CrossRef]
- Borselli, C.; Cezar, C.A.; Shvartsman, D.; Vandenburgh, H.H.; Mooney, D.J. The role of multifunctional delivery scaffold in the ability of cultured myoblasts to promote muscle regeneration. Biomaterials 2011, 32, 8905–8914. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Cao, L.; Shansky, J.; Wang, Z.; Mooney, D.; Vandenburgh, H. Minimally invasive approach to the repair of injured skeletal muscle with a shape-memory scaffold. Mol. Ther. J. Am. Soc. Gene Ther. 2014, 22, 1441–1449. [Google Scholar] [CrossRef] [Green Version]
- Pumberger, M.; Qazi, T.H.; Ehrentraut, M.C.; Textor, M.; Kueper, J.; Stoltenburg-Didinger, G.; Winkler, T.; von Roth, P.; Reinke, S.; Borselli, C.; et al. Synthetic niche to modulate regenerative potential of MSCs and enhance skeletal muscle regeneration. Biomaterials 2016, 99, 95–108. [Google Scholar] [CrossRef] [Green Version]
- Hill, E.; Boontheekul, T.; Mooney, D.J. Regulating activation of transplanted cells controls tissue regeneration. Proc. Natl. Acad. Sci. USA 2006, 103, 2494–2499. [Google Scholar] [CrossRef] [Green Version]
- Passipieri, J.A.; Baker, H.B.; Siriwardane, M.; Ellenburg, M.D.; Vadhavkar, M.; Saul, J.M.; Tomblyn, S.; Burnett, L.; Christ, G.J. Keratin hydrogel enhances in vivo skeletal muscle function in a rat model of volumetric muscle loss. Tissue Eng. Part A 2017. [Google Scholar] [CrossRef]
- Baker, H.B.; Passipieri, J.A.; Siriwardane, M.; Ellenburg, M.D.; Vadhavkar, M.; Bergman, C.R.; Saul, J.M.; Tomblyn, S.; Burnett, L.; Christ, G.J. Cell and growth factor-loaded keratin hydrogels for treatment of volumetric muscle loss in a mouse model. Tissue Eng. Part A 2017. [Google Scholar] [CrossRef]
- Fuoco, C.; Rizzi, R.; Biondo, A.; Longa, E.; Mascaro, A.; Shapira-Schweitzer, K.; Kossovar, O.; Benedetti, S.; Salvatori, M.L.; Santoleri, S.; et al. In vivo generation of a mature and functional artificial skeletal muscle. EMBO Mol. Med. 2015, 7, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Stratos, I.; Madry, H.; Rotter, R.; Weimer, A.; Graff, J.; Cucchiarini, M.; Mittlmeier, T.; Vollmar, B. Fibroblast growth factor-2-overexpressing myoblasts encapsulated in alginate spheres increase proliferation, reduce apoptosis, induce adipogenesis, and enhance regeneration following skeletal muscle injury in rats. Tissue Eng. Part A 2011, 17, 2867–2877. [Google Scholar] [CrossRef]
- Zhou, W.; He, D.Q.; Liu, J.Y.; Feng, Y.; Zhang, X.Y.; Hua, C.G.; Tang, X.F. Angiogenic gene-modified myoblasts promote vascularization during repair of skeletal muscle defects. J. Tissue Eng. Regen. Med. 2015, 9, 1404–1416. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, E.K.; Makarevich, P.I.; Tsokolaeva, Z.I.; Boldyreva, M.A.; Sysoeva, V.Y.; Tkachuk, V.A.; Parfyonova, Y.V. Transplantation of modified human adipose derived stromal cells expressing VEGF165 results in more efficient angiogenic response in ischemic skeletal muscle. J. Transl. Med. 2013, 11, 138. [Google Scholar] [CrossRef] [Green Version]
- Madden, L.; Juhas, M.; Kraus, W.E.; Truskey, G.A.; Bursac, N. Bioengineered human myobundles mimic clinical responses of skeletal muscle to drugs. Elife 2015, 4, e04885. [Google Scholar] [CrossRef] [Green Version]
- Vandenburgh, H.; Shansky, J.; Benesch-Lee, F.; Barbata, V.; Reid, J.; Thorrez, L.; Valentini, R.; Crawford, G. Drug-screening platform based on the contractility of tissue-engineered muscle. Muscle Nerve 2008, 37, 438–447. [Google Scholar] [CrossRef] [Green Version]
- Gilbert-Honick, J.; Ginn, B.; Zhang, Y.; Salehi, S.; Wagner, K.R.; Mao, H.Q.; Grayson, W.L. Adipose-derived stem/stromal cells on electrospun fibrin microfiber bundles enable moderate muscle reconstruction in a volumetric muscle loss model. Cell Transpl. 2018. [Google Scholar] [CrossRef] [Green Version]
- Gilbert-Honick, J.; Iyer, S.R.; Somers, S.M.; Lovering, R.M.; Wagner, K.; Mao, H.Q.; Grayson, W.L. Engineering functional and histological regeneration of vascularized skeletal muscle. Biomaterials 2018, 164, 70–79. [Google Scholar] [CrossRef]
- Nakayama, K.H.; Quarta, M.; Paine, P.; Alcazar, C.; Karakikes, I.; Garcia, V.; Abilez, O.J.; Calvo, N.S.; Simmons, C.S.; Rando, T.A.; et al. Treatment of volumetric muscle loss in mice using nanofibrillar scaffolds enhances vascular organization and integration. Commun. Biol. 2019, 2. [Google Scholar] [CrossRef]
- Rangarajan, S.; Madden, L.; Bursac, N. Use of flow, electrical, and mechanical stimulation to promote engineering of striated muscles. Ann. Biomed. Eng. 2014, 42, 1391–1405. [Google Scholar] [CrossRef] [Green Version]
- Booth, F. Time course of muscular atrophy during immobilization of hindlimbs in rats. J. Appl. Physiol. 1977, 43, 656–661. [Google Scholar] [CrossRef]
- Herbison, G.; Jaweed, M.; Ditunno, J. Muscle atrophy in rats following denervation, casting, inflammation, and tenotomy. Arch. Phys. Med. Rehabil. 1979, 60, 401–404. [Google Scholar]
- Serena, E.; Flaibani, M.; Carnio, S.; Boldrin, L.; Vitiello, L.; De Coppi, P.; Elvassore, N. Electrophysiologic stimulation improves myogenic potential of muscle precursor cells grown in a 3D collagen scaffold. Neurol. Res. 2008, 30, 207–214. [Google Scholar] [CrossRef]
- Moon du, G.; Christ, G.; Stitzel, J.D.; Atala, A.; Yoo, J.J. Cyclic mechanical preconditioning improves engineered muscle contraction. Tissue Eng. Part A 2008, 14, 473–482. [Google Scholar] [CrossRef]
- Machingal, M.A.; Corona, B.T.; Walters, T.J.; Kesireddy, V.; Koval, C.N.; Dannahower, A.; Zhao, W.X.; Yoo, J.J.; Christ, G.J. A Tissue-Engineered Muscle Repair Construct for Functional Restoration of an Irrecoverable Muscle Injury in a Murine Model. Tissue Eng. Part A 2011, 17, 2291–2303. [Google Scholar] [CrossRef] [Green Version]
- Corona, B.T.; Machingal, M.A.; Criswell, T.; Vadhavkar, M.; Dannahower, A.C.; Bergman, C.; Zhao, W.X.; Christ, G.J. Further development of a tissue engineered muscle repair construct in vitro for enhanced functional recovery following implantation in vivo in a murine model of volumetric muscle loss injury. Tissue Eng. Part A 2012, 18, 1213–1228. [Google Scholar] [CrossRef] [Green Version]
- Passipieri, J.A.; Hu, X.; Mintz, E.; Dienes, J.; Baker, H.B.; Wallace, C.H.; Blemker, S.S.; Christ, G.J. In silico and in vivo studies detect functional repair mechanisms in a volumetric muscle loss injury. Tissue Eng. Part A 2019, 25, 1272–1288. [Google Scholar] [CrossRef]
- Corona, B.T.; Ward, C.L.; Baker, H.B.; Walters, T.J.; Christ, G.J. Implantation of in vitro tissue engineered muscle repair constructs and bladder acellular matrices partially restore in vivo skeletal muscle function in a rat model of volumetric muscle loss injury. Tissue Eng. Part A 2014, 20, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Mintz, E.L.; Passipieri, J.A.; Franklin, I.R.; Toscano, V.M.; Afferton, E.C.; Sharma, P.R.; Christ, G.J. Long-term evaluation of functional outcomes following rat volumetric muscle loss injury and repair. Tissue Eng. Part A 2020, 26, 140–156. [Google Scholar] [CrossRef]
- Wu, X.; Corona, B.T.; Chen, X.; Walters, T.J. A standardized rat model of volumetric muscle loss injury for the development of tissue engineering therapies. Biores. Open Access. 2012, 1, 280–290. [Google Scholar] [CrossRef]
- Juhas, M.; Engelmayr, G.C.; Fontanella, A.N.; Palmer, G.M.; Bursac, N. Biomimetic engineered muscle with capacity for vascular integration and functional maturation in vivo. Proc. Natl. Acad. Sci. USA 2014, 111, 5508–5513. [Google Scholar] [CrossRef] [Green Version]
- Morimoto, Y.; Kato-Negishi, M.; Onoe, H.; Takeuchi, S. Three-dimensional neuron-muscle constructs with neuromuscular junctions. Biomaterials 2013, 34, 9413–9419. [Google Scholar] [CrossRef]
- Wang, L.; Shansky, J.; Vandenburgh, H. Induced formation and maturation of acetylcholine receptor clusters in a defined 3D Bio-artificial muscle. Mol. Neurobiol. 2013, 48, 397–403. [Google Scholar] [CrossRef] [Green Version]
- Gholobova, D.; Decroix, L.; Van Muylder, V.; Desender, L.; Gerard, M.; Carpentier, G.; Vandenburgh, H.; Thorrez, L. Endothelial network formation within human tissue-engineered skeletal muscle. Tissue Eng. Part A 2015, 21, 2548–2558. [Google Scholar] [CrossRef] [Green Version]
- Ko, I.K.; Lee, B.K.; Lee, S.J.; Andersson, K.E.; Atala, A.; Yoo, J.J. The effect of in vitro formation of acetylcholine receptor (AChR) clusters in engineered muscle fibers on subsequent innervation of constructs in vivo. Biomaterials 2013, 34, 3246–3255. [Google Scholar] [CrossRef]
- Li, M.T.; Ruehle, M.; Stevens, H.; Servies, N.; Willett, N.; Karthikeyakannan, S.; Warren, G.L.; Guldberg, R.; Krishnan, L.N. Skeletal myoblast-seeded vascularized tissue scaffolds in the treatment of a large volumetric muscle defect in the rat biceps femoris muscle. Tissue Eng. Part A 2017. [Google Scholar] [CrossRef]
- Koffler, J.; Kaufman-Francis, K.; Shandalov, Y.; Egozi, D.; Pavlov, D.A.; Landesberg, A.; Levenberg, S. Improved vascular organization enhances functional integration of engineered skeletal muscle grafts. Proc. Natl. Acad. Sci. USA 2011, 108, 14789–14794. [Google Scholar] [CrossRef] [Green Version]
- Shandalov, Y.; Egozi, D.; Koffler, J.; Dado-Rosenfeld, D.; Ben-Shimol, D.; Freiman, A.; Shor, E.; Kabala, A.; Levenberg, S. An engineered muscle flap for reconstruction of large soft tissue defects. Proc. Natl. Acad. Sci. USA 2014, 111, 6010–6015. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Jun, I.; Park, H.J.; Kang, T.J.; Shin, H.; Cho, S.W. Genetically engineered myoblast sheet for therapeutic angiogenesis. Biomacromolecules 2014, 15, 361–372. [Google Scholar] [CrossRef]
- Ward, C.L.; Pollot, B.E.; Goldman, S.M.; Greising, S.M.; Wenke, J.C.; Corona, B.T. Autologous minced muscle grafts improve muscle strength in a porcine model of volumetric muscle loss injury. J. Orthop. Trauma 2016, 30, e396–e403. [Google Scholar] [CrossRef]
- Sarrafian, T.L.; Bodine, S.C.; Murphy, B.; Grayson, J.K.; Stover, S.M. Extracellular matrix scaffolds for treatment of large volume muscle injuries: A review. Vet. Surg. 2018, 47, 524–535. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| In Situ | In Vivo | In Vitro | |
|---|---|---|---|
| Off-the-shelf availability | Likely | Possible | Not possible |
| Scalability | Easier | Difficult | Most difficult |
| Ease of clinical translation | Easier | Complex | Complex |
| Cost-effectiveness | More | Less | Least |
| Disease modeling | No | No | Yes |
| Drug screening modeling | No | No | Yes |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carnes, M.E.; Pins, G.D. Skeletal Muscle Tissue Engineering: Biomaterials-Based Strategies for the Treatment of Volumetric Muscle Loss. Bioengineering 2020, 7, 85. https://doi.org/10.3390/bioengineering7030085
Carnes ME, Pins GD. Skeletal Muscle Tissue Engineering: Biomaterials-Based Strategies for the Treatment of Volumetric Muscle Loss. Bioengineering. 2020; 7(3):85. https://doi.org/10.3390/bioengineering7030085
Chicago/Turabian StyleCarnes, Meagan E., and George D. Pins. 2020. "Skeletal Muscle Tissue Engineering: Biomaterials-Based Strategies for the Treatment of Volumetric Muscle Loss" Bioengineering 7, no. 3: 85. https://doi.org/10.3390/bioengineering7030085
APA StyleCarnes, M. E., & Pins, G. D. (2020). Skeletal Muscle Tissue Engineering: Biomaterials-Based Strategies for the Treatment of Volumetric Muscle Loss. Bioengineering, 7(3), 85. https://doi.org/10.3390/bioengineering7030085