Lactic Fermented Fruit or Vegetable Juices: Past, Present and Future




Abstract
:1. Introduction
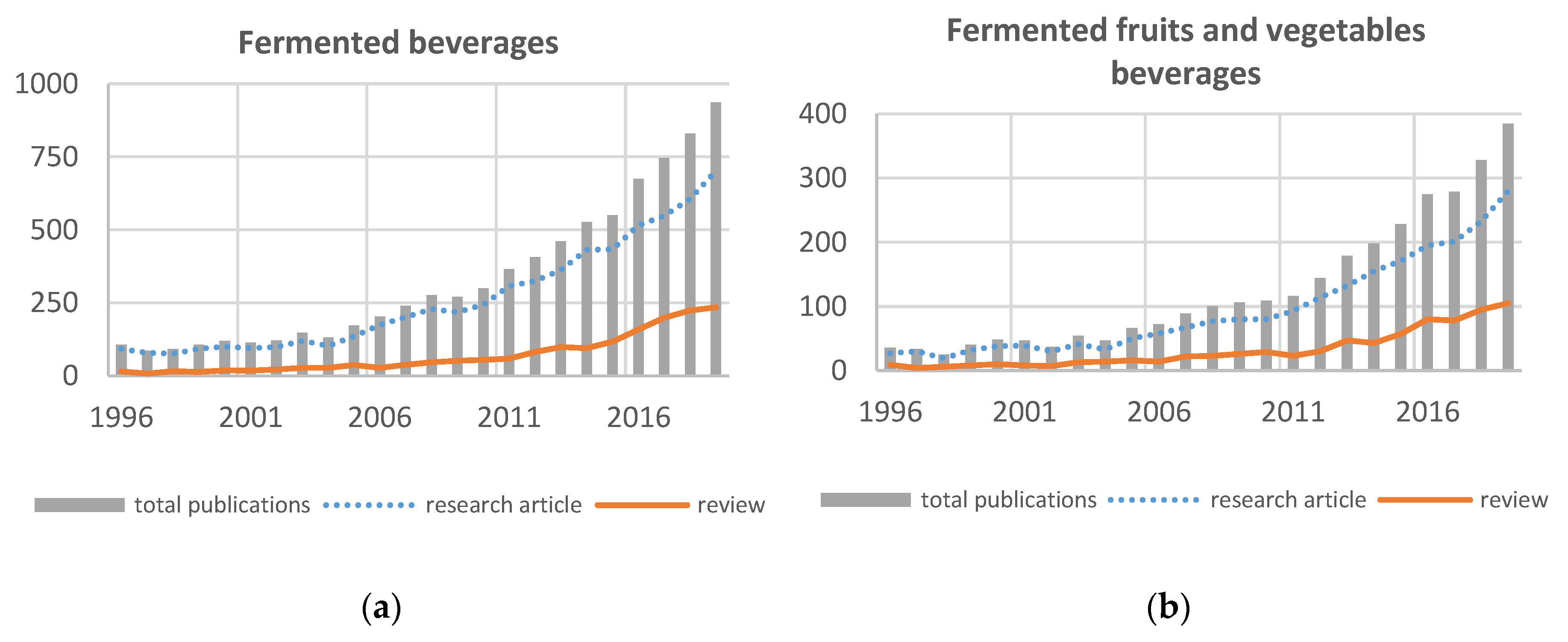
2. Actual Market
3. Traditional Lactic Fermented Beverages
4. Recently Developed Lactic acid Fermented Juices
4.1. Overview
4.2. Juices Stabilization Treatments
4.2.1. Conventional Thermal Treatment of Juices
4.2.2. Alternative Physical Treatment
4.2.3. Pre-Treatments Combined with Fermentation
4.3. Microbiota of Traditional and Recently Developed Lactic Acid Fermented Beverages
4.4. Directing Fermentation
4.5. Nutritional Changes and Functional Effects Mediated by LAB Activity
5. Probiotication
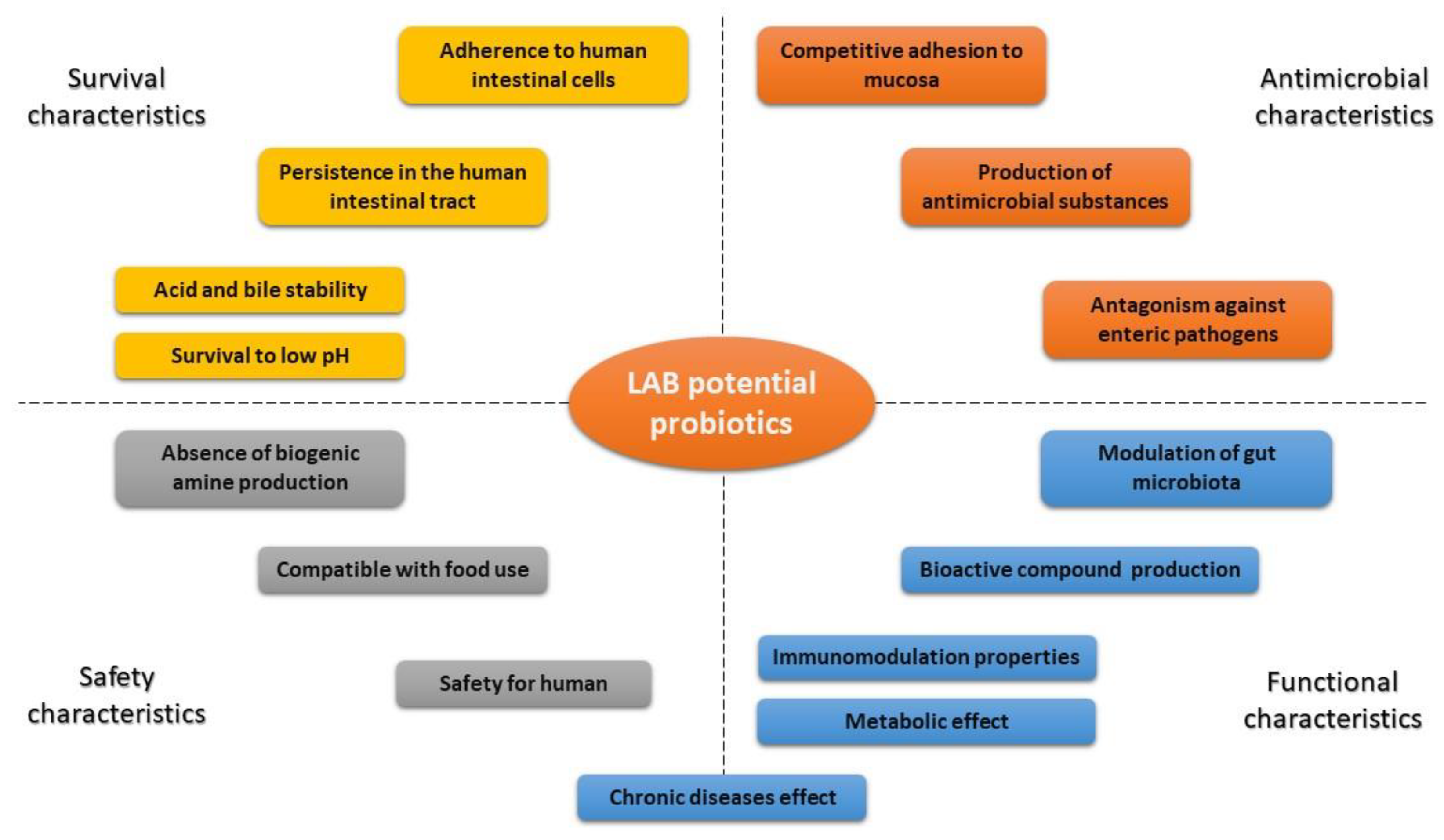
5.1. Probiotic Characteristics and Health Benefits
5.2. Optimized Inoculation to Ensure LAB Viability
5.2.1. Immobilization
5.2.2. Antioxidants
5.2.3. Co-Inoculation
6. Perspectives Related to LAB Physiological Traits and Metabolism of Phenolic Compounds
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tamang, J.P.; Watanabe, K.; Holzapfel, W.H. Review: Diversity of microorganisms in global fermented foods and beverages. Front. Microbiol. 2016, 7, 377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charlton, K.; Kowal, P.; Soriano, M.M.; Williams, S.; Banks, E.; Vo, K.; Byles, J. Fruit and vegetable intake and body mass index in a large sample of middle-aged Australian men and women. Nutrients 2014, 6, 2305–2319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamang, J. Plant-based fermented foods and beverages of Asia. In Handbook of Plant-Based Fermented Food and Beverage Technology, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2012; pp. 49–90. [Google Scholar]
- Karovičová, J.; Kohajdová, Z. Lactic acid fermented vegetable juices. Hortic. Sci. 2011, 30, 152–158. [Google Scholar] [CrossRef] [Green Version]
- Fessard, A.; Kapoor, A.; Patche, J.; Assemat, S.; Hoarau, M.; Bourdon, E.; Bahorun, T.; Remize, F. Lactic fermentation as an efficient tool to enhance the antioxidant activity of tropical fruit juices and teas. Microorganisms 2017, 5, 2. [Google Scholar] [CrossRef] [PubMed]
- Mokoena, M.P.; Mutanda, T.; Olaniran, A.O. Perspectives on the probiotic potential of lactic acid bacteria from African traditional fermented foods and beverages. Food Nutr. Res. 2016, 60, 29630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legras, J.-L.; Merdinoglu, D.; Cornuet, J.-M.; Karst, F. Bread, beer and wine: Saccharomyces cerevisiae diversity reflects human history. Mol. Ecol. 2007, 16, 2091–2102. [Google Scholar] [CrossRef]
- Cavanagh, D.; Fitzgerald, G.F.; McAuliffe, O. From field to fermentation: The origins of Lactococcus lactis and its domestication to the dairy environment. Food Microbiol. 2015, 47, 45–61. [Google Scholar] [CrossRef]
- Campbell-Sills, H.; el Khoury, M.; Favier, M.; Romano, A.; Biasioli, F.; Spano, G.; Sherman, D.J.; Bouchez, O.; Coton, E.; Coton, M.; et al. Phylogenomic Analysis of Oenococcus oeni reveals specific domestication of strains to cider and wines. Genome Biol. 2015, E7, 1506–1518. [Google Scholar] [CrossRef] [Green Version]
- Azam, M.; Mohsin, M.; Ijaz, H.; Tulain, U.R.; Ashraf, M.A.; Fayyaz, A.; Abadeen, Z.; Kamran, Q. Review-Lactic acid bacteria in traditional fermented Asian foods. Pak. J. Pharm. Sci. 2017, 30, 1803–1814. [Google Scholar]
- di Cagno, R.; Coda, R.; de Angelis, M.; Gobbetti, M. Exploitation of vegetables and fruits through lactic acid fermentation. Food Microbiol. 2013, 33, 1–10. [Google Scholar] [CrossRef]
- Corbo, M.R.; Bevilacqua, A.; Petruzzi, L.; Casanova, F.P.; Sinigaglia, M. Functional beverages: The emerging side of functional foods. Compr. Rev. Food Sci. Food Saf. 2014, 13, 1192–1206. [Google Scholar] [CrossRef]
- Marsh, A.J.; O’Sullivan, O.; Hill, C.; Ross, R.P.; Cotter, P.D. Sequence-based analysis of the microbial composition of water kefir from multiple sources. FEMS Microbiol. Lett. 2013, 348, 79–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espirito-Santo, A.P.; Carlin, F.; Renard, C.M.G.C. Apple, grape or orange juice: Which one offers the best substrate for lactobacilli growth?—A screening study on bacteria viability, superoxide dismutase activity, folates production and hedonic characteristics. Food Res. Int. 2015, 78, 352–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Les Grands Prix SIAL Innovation. Available online: https://www.sialparis.fr/Programme/SIAL-Innovation/Les-gagnants-du-prix-SIAL-Innovation/GRAND-PRIX-OR-Namaste-water-kefirs (accessed on 13 December 2019).
- Maldonado, R.R.; da Costa Araújo, L.; da Silva Dariva, L.C.; Rebac, K.N.; de Souza Pinto, I.A.; Prado, J.P.R.; Saeki, J.K.; Silva, T.S.; Takematsu, E.K.; Tiene, N.V.; et al. Potential application of four types of tropical fruits in lactic fermentation. LWT 2017, 86, 254–260. [Google Scholar] [CrossRef]
- Marsh, A.J.; O’Sullivan, O.; Hill, C.; Ross, R.P.; Cotter, P.D. Sequence-based analysis of the bacterial and fungal compositions of multiple kombucha (tea fungus) samples. Food Microbiol. 2014, 38, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Altay, F.; Karbancıoglu-Güler, F.; Daskaya-Dikmen, C. A review on traditional Turkish fermented non-alcoholic beverages: Microbiota, fermentation process and quality characteristics. Int. J. Food Microbiol. 2013, 167, 44–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de la Fuente-Salcido, N.M.; Castañeda-Ramírez, J.C.; García-Almendárez, B.E.; Bideshi, D.K.; Salcedo-Hernández, R.; Barboza-Corona, J.E. Isolation and characterization of bacteriocinogenic lactic bacteria from M-Tuba and Tepache, two traditional fermented beverages in México. Food Sci. Nutr. 2015, 3, 434–442. [Google Scholar] [CrossRef]
- Sõukand, R.; Pieroni, A.; Biró, M.; Dénes, A.; Dogan, Y.; Hajdari, A.; Kallea, R.; Reade, B.; Mustafa, B.; Nedelcheva, A.; et al. An ethnobotanical perspective on traditional fermented plant foods and beverages in Eastern Europe. J. Ethnopharmacol. 2015, 170, 284–296. [Google Scholar] [CrossRef]
- Pidoux, M. The microbial flora of sugary kefir grain (the gingerbeer plant): Biosynthesis of the grain from Lactobacillus hilgardii producing a polysaccharide gel. MIRCEN J. Appl. Microbiol. Biotechnol. 1989, 5, 223–238. [Google Scholar] [CrossRef]
- Magalhães, K.T.; de M. Pereira, G.V.; Dias, D.R.; Schwan, R.F. Microbial communities and chemical changes during fermentation of sugary Brazilian kefir. World J. Microbiol. Biotechnol. 2010, 26, 1241–1250. [Google Scholar] [CrossRef]
- Laureys, D.; de Vuyst, L. Microbial species diversity, community dynamics, and metabolite kinetics of water kefir fermentation. Appl. Environ. Microbiol. 2014, 80, 2564–2572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da CP Miguel, M.G.; Cardoso, P.G.; Magalhães, K.T.; Schwan, R.F. Profile of microbial communities present in tibico (sugary kefir) grains from different Brazilian States. World J. Microbiol. Biotechnol. 2011, 27, 1875–1884. [Google Scholar] [CrossRef]
- Chakravorty, S.; Bhattacharya, S.; Chatzinotas, A.; Chakraborty, W.; Bhattacharya, D.; Gachhui, R. Kombucha tea fermentation: Microbial and biochemical dynamics. Int. J. Food Microbiol. 2016, 220, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Baschali, A.; Tsakalidou, E.; Kyriacou, A.; Karavasiloglou, N.; Matalas, A.-L. Traditional low-alcoholic and non-alcoholic fermented beverages consumed in European countries: A neglected food group. Nutr. Res. Rev. 2017, 30, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Sireswar, S.; Montet, D.; Dey, G. Principal component analysis for clustering probiotic-fortified beverage matrices efficient in elimination of Shigella sp. Fermentation 2018, 4, 34. [Google Scholar] [CrossRef] [Green Version]
- di Cagno, R.; Surico, R.F.; Paradiso, A.; de Angelis, M.; Salmon, J.-C.; Buchin, S.; de Gara, L.; Gobbetti, M. Effect of autochthonous lactic acid bacteria starters on health-promoting and sensory properties of tomato juices. Int. J. Food Microbiol. 2009, 128, 473–483. [Google Scholar] [CrossRef]
- di Cagno, R.; Surico, R.F.; Minervini, G.; de Angelis, M.; Rizzello, C.G.; Gobbetti, M. Use of autochthonous starters to ferment red and yellow peppers (Capsicum annum L.) to be stored at room temperature. Int. J. Food Microbiol. 2009, 130, 108–116. [Google Scholar] [CrossRef]
- Fessard, A.; Remize, F. Genetic and technological characterization of lactic acid bacteria isolated from tropically grown fruits and vegetables. Int. J. Food Microbiol. 2019, 301, 61–72. [Google Scholar] [CrossRef]
- Filannino, P.; Bai, Y.; di Cagno, R.; Gobbetti, M.; Gänzle, M.G. Metabolism of phenolic compounds by Lactobacillus spp. during fermentation of cherry juice and broccoli puree. Food Microbiol. 2015, 46, 272–279. [Google Scholar] [CrossRef]
- Martins, E.M.F.; Ramos, A.M.; Vanzela, E.S.L.; Stringheta, P.C.; de Oliveira Pinto, C.L.; Martins, J.M. Products of vegetable origin: A new alternative for the consumption of probiotic bacteria. Food Res. Int. 2013, 51, 764–770. [Google Scholar] [CrossRef]
- Septembre-Malaterre, A.; Remize, F.; Poucheret, P. Fruits and vegetables, as a source of nutritional compounds and phytochemicals: Changes in bioactive compounds during lactic fermentation. Food Res. Int. 2018, 104, 86–99. [Google Scholar] [CrossRef] [PubMed]
- Capozzi, V.; Fragasso, M.; Romaniello, R.; Berbegal, C.; Russo, P.; Spano, G. Spontaneous food fermentations and potential risks for human health. Fermentation 2017, 3, 49. [Google Scholar] [CrossRef]
- Fessard, A.; Bourdon, E.; Payet, B.; Remize, F. Identification, stress tolerance, and antioxidant activity of lactic acid bacteria isolated from tropically grown fruits and leaves. Can. J. Microbiol. 2016, 62, 550–561. [Google Scholar] [CrossRef] [PubMed]
- Buruleanu, L.; Nicolescu, C.L.; Avram, D.; Bratu, M.G.; Manea, I. Survival of probiotic bacteria during lactic acid fermentation of vegetable juices. J. Agroaliment. Process. Technol. 2009, 15, 132–139. [Google Scholar]
- Yoon, K.Y.; Woodams, E.E.; Hang, Y.D. Fermentation of beet juice by beneficial lactic acid bacteria. LWT-Food Sci. Technol. 2005, 38, 73–75. [Google Scholar] [CrossRef]
- Yoon, K.Y.; Woodams, E.E.; Hang, Y.D. Production of probiotic cabbage juice by lactic acid bacteria. Bioresour. Technol. 2006, 97, 1427–1430. [Google Scholar] [CrossRef]
- Yoon, K.Y.; Woodams, E.E.; Hang, Y.D. Probiotication of tomato juice by lactic acid bacteria. J. Microbiol. 2004, 42, 315–318. [Google Scholar]
- Swain, M.R.; Anandharaj, M.; Ray, R.C.; Rani, R.P. Fermented fruits and vegetables of Asia: A potential source of probiotics. Biotechnol. Res. Int. 2014, 2014, 250424. [Google Scholar] [CrossRef]
- Bergsveinson, J.; Kajala, I.; Ziola, B. Next-generation sequencing approaches for improvement of lactic acid bacteria-fermented plant-based beverages. AIMS Microbiol. 2017, 3, 8–24. [Google Scholar] [CrossRef]
- Corcoran, B.M.; Stanton, C.; Fitzgerald, G.F.; Ross, R.P. Survival of probiotic Lactobacilli in acidic environments is enhanced in the presence of metabolizable sugars. Appl. Environ. Microbiol. 2005, 71, 3060–3067. [Google Scholar] [CrossRef] [Green Version]
- Kang, M.-S.; Kim, Y.-S.; Lee, H.-C.; Lim, H.-S.; Oh, J.-S. Comparison of temperature and additives affecting the stability of the probiotic Weissella cibaria. Chonnam Med. J. 2012, 48, 159–163. [Google Scholar] [CrossRef] [PubMed]
- Beganović, J.; Kos, B.; Pavunc, A.L.; Uroić, K.; Jokić, M.; Šušković, J. Traditionally produced sauerkraut as source of autochthonous functional starter cultures. Microbiol. Res. 2014, 169, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Verón, H.E.; di Risio, H.D.; Isla, M.I.; Torres, S. Isolation and selection of potential probiotic lactic acid bacteria from Opuntia ficus-indica fruits that grow in Northwest Argentina. LWT 2017, 84, 231–240. [Google Scholar] [CrossRef]
- de Souza, J.V.; Dias, F.S. Protective, technological, and functional properties of select autochthonous lactic acid bacteria from goat dairy products. Curr. Opin. Food Sci. 2017, 13, 1–9. [Google Scholar] [CrossRef]
- Fessard, A.; Remize, F. Why are Weissella spp. not used as commercial starter cultures for food fermentation? Fermentation 2017, 3, 38. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Luo, D.; Bao, Y.; Liao, X.; Wu, J. Characterization of diversity and probiotic efficiency of the autochthonous lactic acid bacteria in the fermentation of selected raw fruit and vegetable juices. Front. Microbiol. 2018, 9, 2539. [Google Scholar] [CrossRef] [Green Version]
- Passerini, D.; Beltramo, C.; Coddeville, M.; Quentin, Y.; Ritzenthaler, P.; Daveran-Mingot, M.L.; le Bourgeois, P. Genes but not genomes reveal bacterial domestication of Lactococcus lactis. PLoS ONE 2010, 5, e15306. [Google Scholar] [CrossRef] [Green Version]
- Manno, M.T.; Zuljan, F.; Alarcon, S.; Esteban, L.; Blancato, V.; Espariz, M.; Magni, C. Genetic and phenotypic features defining industrial relevant Lactococcus lactis, L. cremoris and L. lactis biovar. diacetylactis strains. J. Biotechnol. 2018, 282, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Passerini, D.; Laroute, V.; Coddeville, M.; le Bourgeois, P.; Loubière, P.; Ritzenthaler, P.; Cocaign-Bousquet, M.; Daveran-Mingot, M.-L. New insights into Lactococcus lactis diacetyl- and acetoin-producing strains isolated from diverse origins. Int. J. Food Microbiol. 2013, 160, 329–336. [Google Scholar] [CrossRef]
- Endo, A.; Tanizawa, Y.; Tanaka, N.; Maeno, S.; Kumar, H.; Shiwa, Y.; Okada, S.; Yoshikawa, H.; Dicks, L.; Nakagawa, J.; et al. Comparative genomics of Fructobacillus spp. and Leuconostoc spp. reveals niche-specific evolution of Fructobacillus spp. BMC Genom. 2015, 16, 1117. [Google Scholar] [CrossRef] [Green Version]
- Martino, M.E.; Bayjanov, J.R.; Caffrey, B.E.; Wels, M.; Joncour, P.; Hughes, S.; Gillet, B.; Kleerebeze, M.; van Hijum, S.A.F.T.; Leulier, F. Nomadic lifestyle of Lactobacillus plantarum revealed by comparative genomics of 54 strains isolated from different habitats. Environ. Microbiol. 2016, 18, 4974–4989. [Google Scholar] [CrossRef] [PubMed]
- Sivudu, S.N.; Umamahesh, K.; Reddy, O.V.S. A comparative study on probiotication of mixed watermelon and tomato juice by using probiotic strains of Lactobacilli. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 977–984. [Google Scholar]
- Koh, J.-H.; Kim, Y.; Oh, J.-H. Chemical characterization of tomato juice fermented with bifidobacteria. J. Food Sci. 2010, 75, C428–C432. [Google Scholar] [CrossRef] [PubMed]
- Corona, O.; Randazzo, W.; Miceli, A.; Guarcello, R.; Francesca, N.; Erten, H.; Moschettia, G.; Settanni, L. Characterization of kefir-like beverages produced from vegetable juices. LWT Food Sci. Technol. 2016, 66, 572–581. [Google Scholar] [CrossRef] [Green Version]
- Gumienna, M.; Szwengiel, A.; Górna, B. Bioactive components of pomegranate fruit and their transformation by fermentation processes. Eur. Food Res. Technol. 2016, 242, 631–640. [Google Scholar] [CrossRef] [Green Version]
- Jeong, S.-Y.; Velmurugan, P.; Lim, J.-M.; Oh, B.-T.; Jeong, D.-Y. Photobiological (LED light)-mediated fermentation of blueberry (Vaccinium corymbosum L.) fruit with probiotic bacteria to yield bioactive compounds. LWT 2018, 93, 158–166. [Google Scholar] [CrossRef]
- Costa, M.G.M.; Fonteles, T.V.; de Jesus, A.L.T.; Rodrigues, S. Sonicated pineapple juice as substrate for L. casei cultivation for probiotic beverage development: Process optimisation and product stability. Food Chem. 2013, 139, 261–266. [Google Scholar] [CrossRef] [Green Version]
- Zheng, X.; Yu, Y.; Xiao, G.; Xu, Y.; Wu, J.; Tang, D.; Zhang, Y. Comparing product stability of probiotic beverages using litchi juice treated by high hydrostatic pressure and heat as substrates. Innov. Food Sci. Emerg. Technol. 2014, 23, 61–67. [Google Scholar] [CrossRef]
- Chen, H.; Xiao, G.; Xu, Y.; Yu, Y.; Wu, J.; Zou, B. High hydrostatic pressure and co-fermentation by Lactobacillus rhamnosus and Gluconacetobacter xylinus improve flavor of yacon-litchi-longan juice. Foods 2019, 8, 8. [Google Scholar] [CrossRef] [Green Version]
- Miller, F.A.; Silva, C.L.M. Thermal treatment effects in fruit juices. In Advances in Fruit Processing Technologies; Rodrigues, S., Fernandes, F.A.N., Eds.; CRC Press: Boca Raton, FA, USA, 2012; pp. 363–383. ISBN 978-1-4398-5153-1. [Google Scholar]
- Marszałek, K.; Krzyżanowska, J.; Woźniak, Ł.; Skąpska, S. Kinetic modelling of polyphenol oxidase, peroxidase, pectin esterase, polygalacturonase, degradation of the main pigments and polyphenols in beetroot juice during high pressure carbon dioxide treatment. LWT-Food Sci. Technol. 2017, 85, 412–417. [Google Scholar] [CrossRef]
- Mercali, G.D.; Gurak, P.D.; Schmitz, F.; Marczak, L.D.F. Evaluation of non-thermal effects of electricity on anthocyanin degradation during ohmic heating of jaboticaba (Myrciaria cauliflora) juice. Food Chem. 2015, 171, 200–205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Profir, A.G.; Vizireanu, C. Effect of the preservation processes on the storage stability of juice made from carrot, celery and beetroot. J. Agroaliment. Process. Technol. 2013, 19, 99–104. [Google Scholar]
- Dima, F.; Istrati, D.; Garnai, M.C.; Serea, V.; Vizireanu, C. Study on obtaining vegetables juices with high antioxidant potential, preserved by ohmic pasteurization. J. Agroaliment. Process. Technol. 2015, 21, 67–74. [Google Scholar]
- Bhat, S.; Saini, C.S.; Kumar, M.; Sharma, H.K. Effect of thermal and alternate thermal processing on bottle gourd (L agenaria siceraria) juice. J. Food Process. Preserv. 2017, 41, e12911. [Google Scholar] [CrossRef]
- Achir, N.; Dhuique-Mayer, C.; Hadjal, T.; Madani, K.; Pain, J.-P.; Dornier, M. Pasteurization of citrus juices with ohmic heating to preserve the carotenoid profile. Innov. Food Sci. Emerg. Technol. 2016, 33, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Tian, X.; Yu, Q.; Wu, W.; Dai, R. Inactivation of microorganisms in foods by ohmic heating: A review. J. Food Prot. 2018, 7, 1093–1107. [Google Scholar] [CrossRef]
- Arjmandi, M.; Otón, M.; Artés, F.; Artés-Hernández, F.; Gómez, P.A.; Aguayo, E. Semi-industrial microwave treatments positively affect the quality of orange-colored smoothies. J. Food Sci. Technol. 2016, 10, 3695–3703. [Google Scholar] [CrossRef] [Green Version]
- Saikia, S.; Mahnot, N.K.; Mahanta, C.L. A comparative study on the effect of conventional thermal pasteurisation, microwave and ultrasound treatments on the antioxidant activity of five fruit juices. Food Sci. Technol. Int. 2016, 22, 288–301. [Google Scholar] [CrossRef]
- Stratakos, A.C.; Delgado-Pando, G.; Linton, M.; Patterson, M.F.; Koidis, A. Industrial scale microwave processing of tomato juice using a novel continuous microwave system. Food Chem. 2016, 190, 622–628. [Google Scholar] [CrossRef] [Green Version]
- Math, R.G.; Nagender, A.; Nayani, A.S.; Satyanarayana, A. Continuous microwave processing and preservation ofacidic and non acidic juice blends. IJAFST 2014, 2, 81–90. [Google Scholar]
- Salazar-González, C.; Martín-González, M.F.S.; Vergara-Balderas, F.T.; López-Malo, A.; Sosa-Morales, M.E. Physical-chemical and microbiological stability during refrigerated storage of microwave-pasteurized guava nectar. Focus. Mod. Food Ind. 2014, 3, 43. [Google Scholar] [CrossRef]
- Pérez-Grijalva, B.; Herrera-Sotero, M.; Mora-Escobedo, R.; Zebadùa-Garcià, J.C.; Silva-Hernàndez, E.; Oliart-Ros, R.; Pérez-Cruz, C.; Guzmàn-Geronimo, R. Effect of microwaves and ultrasound on bioactive compounds and microbiological quality of blackberry juice. LWT 2018, 87, 47–53. [Google Scholar] [CrossRef]
- Gonçalves, L.C.P.; di Genova, B.M.; Dörr, F.A.; Pinto, E.; Bastos, E.L. Effect of dielectric microwave heating on the color and antiradical capacity of betanin. J. Food Eng. 2013, 118, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Abdullah, N. Application of thermosonication treatment in processing and production of high quality and safe-to-drink fruit juices. Agric. Agric. Sci. Procedia 2014, 2, 320–327. [Google Scholar] [CrossRef] [Green Version]
- Abid, M.; Jabbar, S.; Hu, B.; Hashim, M.M.; Wu, T.; Lei, S.; Kan, M.A.; Zheng, X. Thermosonication as a potential quality enhancement technique of apple juice. Ultrason. Sonochem. 2014, 21, 984–990. [Google Scholar] [CrossRef]
- el Kantar, S.; Boussetta, N.; Lebovka, N.I.; Foucart, F.; Rajha, H.N.; Maroun, R.G.; Louka, N.; Vorobiev, E. Pulsed electric field treatment of citrus fruits: Improvement of juice and polyphenols extraction. Innov. Food Sci. Emerg. Technol. 2018, 46, 153–161. [Google Scholar] [CrossRef]
- Mesquita, T.C.; Schiassi, M.C.E.V.; Lago, A.M.T.; Careli-Grondim, I.; Silva, L.M.; de Azevedo Lira, N.; Carvalho, E.E.N.; de Oliveira Lima, L.C. Grape juice blends treated with gamma irradiation evaluated during storage. Radiat. Phys. Chem. 2020, 168, 108570. [Google Scholar] [CrossRef]
- Mestry, A.P.; Mujumdar, A.S.; Thorat, B.N. Optimization of spray drying of an innovative functional food: Fermented mixed juice of carrot and watermelon. Dry. Technol. 2011, 29, 1121–1131. [Google Scholar] [CrossRef]
- Fiorda, F.A.; de Melo Pereira, G.V.; Thomaz-Soccol, V.; Rakshit, S.K.; Pagnoncelli, M.G.B.; Vandenberghe, L.P.S.; Soccol, C.R. Microbiological, biochemical, and functional aspects of sugary kefir fermentation—A review. Food Microbiol. 2017, 66, 86–95. [Google Scholar] [CrossRef]
- Gulitz, A.; Stadie, J.; Wenning, M.; Ehrmann, M.A.; Vogel, R.F. The microbial diversity of water kefir. Int. J. Food Microbiol. 2011, 151, 284–288. [Google Scholar] [CrossRef]
- Gulitz, A.; Stadie, J.; Ehrmann, M.A.; Ludwig, W.; Vogel, R.F. Comparative phylobiomic analysis of the bacterial community of water kefir by 16S rRNA gene amplicon sequencing and ARDRA analysis. J. Appl. Microbiol. 2013, 114, 1082–1091. [Google Scholar] [CrossRef] [PubMed]
- Stadie, J.; Gulitz, A.; Ehrmann, M.A.; Vogel, R.F. Metabolic activity and symbiotic interactions of lactic acid bacteria and yeasts isolated from water kefir. Food Microbiol. 2013, 35, 92–98. [Google Scholar] [CrossRef]
- Bogdan, M.; Justine, S.; Filofteia, D.; Petruta, C.; Gabriela, L.; Roxana, U.; Florentina, M. Lactic acid bacteria strains isolated from Kombucha with potential probiotic effect. Rom. Biotechnol. Lett. 2018, 23, 13592–13598. [Google Scholar]
- Tanguler, H.; Erten, H. Occurrence and growth of lactic acid bacteria species during the fermentation of shalgam (salgam), a traditional Turkish fermented beverage. LWT-Food Sci. Technol. 2012, 46, 36–41. [Google Scholar] [CrossRef]
- Tanguler, H.; Erten, H. Chemical and microbiological characteristics of shalgam (salgam), a traditional Turkish lactic acid fermented beverage. J. Food Qual. 2012, 35, 298–306. [Google Scholar] [CrossRef]
- Touret, T.; Oliveira, M.; Semedo-Lemsaddek, T. Putative probiotic lactic acid bacteria isolated from sauerkraut fermentations. PLoS ONE 2018, 13, e0203501. [Google Scholar] [CrossRef] [Green Version]
- Plengvidhya, V.; Breidt, F.; Lu, Z.; Fleming, H.P.; Fleming, H.P. DNA fingerprinting of lactic acid bacteria in sauerkraut fermentations. Appl. Environ. Microbiol. 2007, 73, 7697–7702. [Google Scholar] [CrossRef] [Green Version]
- Wuyts, S.; van Beeck, W.; Oerlemans, E.; Wittouck, S.; Claes, I.; de Boeck, I.; Weckx, S.; Lievens, B.; de Vuyst, L.; Lebeer, S. Carrot juice fermentations as man-made microbial ecosystems dominated by lactic acid bacteria. Appl. Environ. Microbiol. 2018, 84, e00134-18. [Google Scholar] [CrossRef] [Green Version]
- Puerari, C.; Magalhães, K.T.; Schwan, R.F. New cocoa pulp-based kefir beverages: Microbiological, chemical composition and sensory analysis. Food Res. Int. 2012, 48, 634–640. [Google Scholar] [CrossRef] [Green Version]
- Randazzo, W.; Corona, O.; Guarcello, R.; Francesca, N.; Germanà, M.; Erten, H.; Moschetti, G.; Settanni, L. Development of new non-dairy beverages from Mediterranean fruit juices fermented with water kefir microorganisms. Food Microbiol. 2016, 54, 40–51. [Google Scholar] [CrossRef] [Green Version]
- Moraru, D.; Bleoanca, I.; Segal, R. Probiotic vegetable juices. The Annals of the University Dunarea de Jos of Galati–Fascicle IV. Food Technol. 2007, 4, 87–91. [Google Scholar]
- Kun, S.; Rezessy-Szabó, J.M.; Nguyen, Q.D.; Hoschke, Á. Changes of microbial population and some components in carrot juice during fermentation with selected Bifidobacterium strains. Process Biochem. 2008, 43, 816–821. [Google Scholar] [CrossRef]
- Sharma, V.; Mishra, H.N. Unstructured kinetic modeling of growth and lactic acid production by Lactobacillus plantarum NCDC 414 during fermentation of vegetable juices. LWT Food Sci. Technol. 2014, 59, 1123–1128. [Google Scholar] [CrossRef]
- di Cagno, R.; Cardinali, G.; Minervini, G.; Antonielli, L.; Rizzello, C.G.; Ricciuti, P.; Gobetti, M. Taxonomic structure of the yeasts and lactic acid bacteria microbiota of pineapple (Ananas comosus L. Merr.) and use of autochthonous starters for minimally processing. Food Microbiol. 2010, 27, 381–389. [Google Scholar] [CrossRef]
- Mousavi, Z.E.; Mousavi, S.M.; Razavi, S.H.; Emam-Djomeh, Z.; Kiani, H. Fermentation of pomegranate juice by probiotic lactic acid bacteria. World J. Microbiol. Biotechnol. 2011, 27, 123–128. [Google Scholar] [CrossRef]
- Mousavi, Z.E.; Mousavi, S.M.; Razavi, S.H.; Hadinejad, M.; Emam-Djomeh, Z.; Mirzapour, M. Effect of fermentation of pomegranate juice by Lactobacillus plantarum and Lactobacillus acidophilus on the antioxidant activity and metabolism of sugars, organic acids and phenolic compounds. Food Biotechnol. 2013, 27, 1–13. [Google Scholar] [CrossRef]
- Filannino, P.; Azzi, L.; Cavoski, I.; Vincentini, O.; Rizzello, C.; Gobbetti, M.; di Cagno, R. Exploitation of the health-promoting and sensory properties of organic pomegranate (Punica granatum L.) juice through lactic acid fermentation. Int. J. Food Microbiol. 2013, 163, 184–192. [Google Scholar] [CrossRef]
- Valero-Cases, E.; Nuncio-Jáuregui, N.; Frutos, M.J. Influence of fermentation with different lactic acid bacteria and in vitro digestion on the biotransformation of phenolic compounds in fermented pomegranate juices. J. Agric. Food Chem. 2017, 65, 6488–6496. [Google Scholar] [CrossRef]
- Rahman, N.; Xiaohong, C.; Meiqin, F.; Mingsheng, D. Characterization of the dominant microflora in naturally fermented camel milk shubat. World J. Microbiol. Biotechnol. 2009, 25, 1941–1946. [Google Scholar] [CrossRef]
- Wang, C.-Y.; Ng, C.-C.; Su, H.; Tzeng, W.-S.; Shyu, Y.-T. Probiotic potential of noni juice fermented with lactic acid bacteria and bifidobacteria. Int. J. Food Sci. Nutr. 2009, 60, 98–106. [Google Scholar] [CrossRef]
- di Cagno, R.; Minervini, G.; Rizzello, C.G.; de Angelis, M.; Gobbetti, M. Effect of lactic acid fermentation on antioxidant, texture, color and sensory properties of red and green smoothies. Food Microbiol. 2011, 28, 1062–1071. [Google Scholar] [CrossRef]
- Ankolekar, C.; Pinto, M.; Greene, D.; Shetty, K. In vitro bioassay based screening of antihyperglycemia and antihypertensive activities of Lactobacillus acidophilus fermented pear juice. Innov. Food Sci. Emerg. Technol. 2012, 13, 221–230. [Google Scholar] [CrossRef]
- Prado, F.C.; Lindner, J.D.D.; Inaba, J.; Thomaz-Soccol, V.; Brar, S.K.; Soccol, C.R. Development and evaluation of a fermented coconut water beverage with potential health benefits. J. Funct. Foods 2015, 12, 489–497. [Google Scholar] [CrossRef]
- Reddy, L.V.; Min, J.-H.; Wee, Y.-J. Production of probiotic mango juice by fermentation of lactic acid bacteria. Microbiol. Biotechnol. Lett. 2015, 43, 120–125. [Google Scholar] [CrossRef] [Green Version]
- di Cagno, R.; Filannino, P.; Vincentini, O.; Lanera, A.; Cavoski, I.; Gobbetti, M. Exploitation of Leuconostoc mesenteroides strains to improve shelf life, rheological, sensory and functional features of prickly pear (Opuntia ficus-indica L.) fruit puree. Food Microbiol. 2016, 59, 176–189. [Google Scholar] [CrossRef]
- Panda, S.K.; Behera, S.K.; Qaku, X.W.; Sekar, S.; Ndinteh, D.T.; Nanjundaswamy, H.M.; Ray, R.C.; Kayitesi, E. Quality enhancement of prickly pears (Opuntia sp.) juice through probiotic fermentation using Lactobacillus fermentum-ATCC 9338. LWT 2017, 75, 453–459. [Google Scholar] [CrossRef]
- Verón, H.E.; Cano, P.G.; Fabersani, E.; Sanz, Y.; Isla, M.I.; Espinar, M.T.F.; Ponce, J.V.G.; Torres, S. Cactus pear (Opuntia ficus-indica) juice fermented with autochthonous Lactobacillus plantarum S-811. Food Funct. 2019, 10, 1085–1097. [Google Scholar] [CrossRef]
- Hashemi, S.M.B.; Khaneghah, A.M.; Barba, F.J.; Nemati, Z.; Shokofti, S.S.; Alizadeh, F. Fermented sweet lemon juice (Citrus limetta) using Lactobacillus plantarum LS5: Chemical composition, antioxidant and antibacterial activities. J. Funct. Foods 2017, 38, 409–414. [Google Scholar] [CrossRef]
- Kaprasob, R.; Kerdchoechuen, O.; Laohakunjit, N.; Sarkar, D.; Shetty, K. Fermentation-based biotransformation of bioactive phenolics and volatile compounds from cashew apple juice by select lactic acid bacteria. Process Biochem. 2017, 59, 141–149. [Google Scholar] [CrossRef]
- Vivek, K.; Mishra, S.; Pradhan, R.C.; Jayabalan, R. Effect of probiotification with Lactobacillus plantarum MCC 2974 on quality of Sohiong juice. LWT 2019, 108, 55–60. [Google Scholar] [CrossRef]
- Cui, S.; Zhao, N.; Lu, W.; Zhao, F.; Zheng, S.; Wang, W.; Chen, W. Effect of different Lactobacillus species on volatile and nonvolatile flavor compounds in juices fermentation. Food Sci. Nutr. 2019, 7, 2214–2223. [Google Scholar] [CrossRef] [Green Version]
- di Cagno, R.; Surico, R.F.; Siragusa, S.; de Angelis, M.; Paradiso, A.; Minervini, F.; de Gara, L.; Gobbetti, M. Selection and use of autochthonous mixed starter for lactic acid fermentation of carrots, French beans or marrows. Int. J. Food Microbiol. 2008, 127, 220–228. [Google Scholar] [CrossRef]
- Subramaniyam, R.S.; Vimala, R. Solid state and submerged fermentation for the production of bioactive substances: A comparative study. Int. J. Sci. Nat. 2012, 3, 480–486. [Google Scholar]
- Mantzourani, I.; Kazakos, S.; Terpou, A.; Alexopoulos, A.; Bezirtzoglou, E.; Bekatorou, A.; Plessas, S. Potential of the probiotic Lactobacillus plantarum ATCC 14917 strain to produce functional fermented pomegranate juice. Foods 2018, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Pereira, A.L.F.; Feitosa, W.S.C.; Abreu, V.K.G.; Lemos, T.d.; Gomes, W.F.; Narain, N.; Rodrigues, S. Impact of fermentation conditions on the quality and sensory properties of a probiotic cupuassu (Theobroma grandiflorum) beverage. Food Res. Int. 2017, 100, 603–611. [Google Scholar] [CrossRef]
- Garcia, E.F.; de Oliveira Araújo, A.; Luciano, W.A.; de Albuquerque, T.M.R.; de Oliveira Arcanjo, N.M.; Madruga, M.S.; dos Santos Lima, M.; Magnani, M.; Saarela, M.; de Souza, E.L. The performance of five fruit-derived and freeze-dried potentially probiotic Lactobacillus strains in apple, orange, and grape juices. J. Sci. Food Agric. 2018, 13, 5000–5010. [Google Scholar] [CrossRef]
- Cirlini, M.; Ricci, A.; Galaverna, G.; Lazzi, C. Application of lactic acid fermentation to elderberry juice: Changes in acidic and glucidic fractions. LWT 2020, 118, 108779. [Google Scholar] [CrossRef]
- Markkinen, N.; Laaksonen, O.; Nahku, R.; Kuldjärv, R.; Yang, B. Impact of lactic acid fermentation on acids, sugars, and phenolic compounds in black chokeberry and sea buckthorn juices. Food Chem. 2019, 286, 204–215. [Google Scholar] [CrossRef]
- di Cagno, R.; Surico, R.F.; Minervini, G.; Rizello, C.G.; Lovino, R.; Servili, M.; Taticchi, A.; Urbani, S.; Gobetti, M. Exploitation of sweet cherry (Prunus avium L.) puree added of stem infusion through fermentation by selected autochthonous lactic acid bacteria. Food Microbiol. 2011, 28, 900–909. [Google Scholar] [CrossRef]
- Gao, H.; Wen, J.-J.; Hu, J.-L.; Nie, Q.-X.; Chen, H.-H.; Nie, S.-P.; Xiong, T.; Xie, M.-Y. Momordica charantia juice with Lactobacillus plantarum fermentation: Chemical composition, antioxidant properties and aroma profile. Food Biosci. 2019, 29, 62–72. [Google Scholar] [CrossRef]
- Kwaw, E.; Ma, Y.; Tchabo, W.; Apaliya, M.T.; Wu, M.; Sackey, A.S.; Xiao, L.; Tahir, H.E. Effect of Lactobacillus strains on phenolic profile, color attributes and antioxidant activities of lactic-acid-fermented mulberry juice. Food Chem. 2018, 250, 148–154. [Google Scholar] [CrossRef]
- Li, Z.; Teng, J.; Lyu, Y.; Hu, X.; Zhao, Y.; Wang, M. Enhanced antioxidant activity for apple juice fermented with Lactobacillus plantarum ATCC14917. Molecules 2018, 24, 51. [Google Scholar] [CrossRef] [Green Version]
- Yan, Y.; Zhang, F.; Chai, Z.; Liu, M.; Battino, M.; Meng, X. Mixed fermentation of blueberry pomace with L. rhamnosus GG and L. plantarum-1: Enhance the active ingredient, antioxidant activity and health-promoting benefits. Food Chem. Toxicol. 2019, 131, 110541. [Google Scholar] [CrossRef]
- Hur, S.J.; Lee, S.Y.; Kim, Y.-C.; Choi, I.; Kim, G.-B. Effect of fermentation on the antioxidant activity in plant-based foods. Food Chem. 2014, 160, 346–356. [Google Scholar] [CrossRef]
- Ye, J.-H.; Huang, L.-Y.; Terefe, N.S.; Augustin, M.A. Fermentation-based biotransformation of glucosinolates, phenolics and sugars in retorted broccoli puree by lactic acid bacteria. Food Chem. 2019, 286, 616–623. [Google Scholar] [CrossRef]
- Ricci, A.; Cirlini, M.; Calani, L.; Bernini, V.; Neviani, E.; del Rio, D.; Galaverna, G.; Lazzi, C. In vitro metabolism of elderberry juice polyphenols by lactic acid bacteria. Food Chem. 2019, 276, 692–699. [Google Scholar] [CrossRef]
- Scalbert, A.; Manach, C.; Morand, C.; Rémésy, C.; Jiménez, L. Dietary polyphenols and the prevention of diseases. Crit. Rev. Food Sci. Nutr. 2005, 45, 287–306. [Google Scholar] [CrossRef]
- Costa, C.; Tsatsakis, A.; Mamoulakis, C.; Teodoro, M.; Briguglio, G.; Caruso, E.; Tsoukalas, D.; Margina, D.; Dardiotis, E.; Kouretas, D.; et al. Current evidence on the effect of dietary polyphenols intake on chronic diseases. Food Chem. Toxicol. 2017, 110, 286–299. [Google Scholar] [CrossRef]
- Silva, F.; Borges, F.; Guimarães, C.; Lima, J.; Matos, C.; Reis, S. Phenolic acids and derivatives: Studies on the relationship among structure, radical scavenging activity, and physicochemical parameters. J. Agric. Food Chem. 2000, 48, 2122–2126. [Google Scholar] [CrossRef]
- Huang, J.; de Paulis, T.; May, J.M. Antioxidant effects of dihydrocaffeic acid in human EA.hy926 endothelial cells. J. Nutr. Biochem. 2004, 12, 722–729. [Google Scholar] [CrossRef]
- Lin, D.; Xiao, M.; Zhao, J.; Li, Z.; Xing, B.; Li, X.; Kong, M.; Li, L.; Zhang, Q.; Liu, Y.; et al. An overview of plant phenolic compounds and their importance in human nutrition and management of type 2 diabetes. Molecules 2016, 21, 10. [Google Scholar] [CrossRef]
- Nagappan, H.; Pee, P.P.; Kee, S.H.Y.; Ow, J.T.; Yan, S.W.; Chew, L.Y.; Kong, K.W. Malaysian brown seaweeds Sargassum siliquosum and Sargassum polycystum: Low density lipoprotein (LDL) oxidation, angiotensin converting enzyme (ACE), α-amylase, and α-glucosidase inhibition activities. Food Res. Int. 2017, 99, 950–958. [Google Scholar] [CrossRef]
- Tan, Y.; Chang, S.K.C.; Zhang, Y. Comparison of α-amylase, α-glucosidase and lipase inhibitory activity of the phenolic substances in two black legumes of different genera. Food Chem. 2017, 214, 259–268. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Hogan, S.; Li, J.; Sun, S.; Canning, C.; Zheng, S.J.; Zhou, K. Grape skin extract inhibits mammalian intestinal α-glucosidase activity and suppresses postprandial glycemic response in streptozocin-treated mice. Food Chem. 2011, 126, 466–471. [Google Scholar] [CrossRef]
- He, Q.; Lv, Y.; Yao, K. Effects of tea polyphenols on the activities of α-amylase, pepsin, trypsin and lipase. Food Chem. 2007, 101, 1178–1182. [Google Scholar] [CrossRef]
- Kostinek, M.; Spechta, I.; Edward, V.A.; Schillinger, U.; Hertel, C.; Holzapfel, W.H.; Franza, C.M.A.P. Diversity and technological properties of predominant lactic acid bacteria from fermented cassava used for the preparation of Gari, a traditional African food. Syst. Appl. Microbiol. 2005, 28, 527–540. [Google Scholar] [CrossRef]
- Kohajdová, Z.; Karovičová, J. Fermentation of cereals for specific purpose. J. Food Nutr. Res. 2007, 46, 51–57. [Google Scholar]
- Roger, T.; Léopold, T.N.; Funtong, M.C.M. Nutritional properties and antinutritional factors of corn paste (kutukutu) fermented by different strains of lactic acid bacteria. Int. J. Food Sci. 2015, 2015, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Ripari, V. Techno-functional role of exopolysaccharides in cereal-based, yogurt-like beverages. Beverages 2019, 5, 16. [Google Scholar] [CrossRef] [Green Version]
- Juvonen, R.; Honkapää, K.; Maina, N.H.; Shi, Q.; Viljanen, K.; Maaheimo, H.; Virkki, L.; Tenkanen, M.; Lantto, R. The impact of fermentation with exopolysaccharide producing lactic acid bacteria on rheological, chemical and sensory properties of pureed carrots (Daucus carota L.). Int. J. Food Microbiol. 2015, 207, 109–118. [Google Scholar] [CrossRef]
- Lynch, K.M.; Zannini, E.; Coffey, A.; Arendt, E.K. Lactic acid bacteria exopolysaccharides in foods and beverages: Isolation, properties, characterization, and health benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 155–176. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, C.; Li, D.; Zhao, Y.; Zhang, X.; Zeng, X.; Yang, Z.; Li, S. Antioxidant activity of an exopolysaccharide isolated from Lactobacillus plantarum C88. Int. J. Biol. Macromol. 2013, 54, 270–275. [Google Scholar] [CrossRef]
- Pan, D.; Mei, X. Antioxidant activity of an exopolysaccharide purified from Lactococcus lactis subsp. lactis 12. Carbohydr. Polym. 2010, 80, 908–914. [Google Scholar] [CrossRef]
- Liu, C.-F.; Tseng, K.-C.; Chiang, S.-S.; Lee, B.-H.; Hsu, W.-H.; Pan, T.-M. Immunomodulatory and antioxidant potential of Lactobacillus exopolysaccharides. J. Sci. Food Agric. 2011, 91, 2284–2291. [Google Scholar] [CrossRef]
- Caggianiello, G.; Kleerebezem, M.; Spano, G. Exopolysaccharides produced by lactic acid bacteria: From health-promoting benefits to stress tolerance mechanisms. Appl. Microbiol. Biotechnol. 2016, 100, 3877–3886. [Google Scholar] [CrossRef]
- Rivera-Espinoza, Y.; Gallardo-Navarro, Y. Non-dairy probiotic products. Food Microbiol. 2010, 27, 1–11. [Google Scholar] [CrossRef]
- Pinto, S.S.; Verruck, S.; Vieira, C.R.W.; Prudêncio, E.S.; Amante, E.R.; Amboni, R.D.M.C. Influence of microencapsulation with sweet whey and prebiotics on the survival of Bifidobacterium-BB-12 under simulated gastrointestinal conditions and heat treatments. LWT-Food Sci. Technol. 2015, 64, 1004–1009. [Google Scholar] [CrossRef]
- Chaudhary, A. Probiotic fruit and vegetable juices: Approach towards a healthy gut. Int. J. Curr. Microbiol. Appl. Sci. 2019, 8, 1265–1279. [Google Scholar] [CrossRef]
- Parvez, S.; Malik, K.A.; Kang, S.A.; Kim, H.-Y. Probiotics and their fermented food products are beneficial for health. J. Appl. Microbiol. 2006, 100, 1171–1185. [Google Scholar] [CrossRef]
- Nagpal, R.; Kumar, A.; Kumar, M.; Behare, P.V.; Jain, S.; Yadav, H. Probiotics, their health benefits and applications for developing healthier foods: A review. FEMS Microbiol. Lett. 2012, 334, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.-M.; Guo, J.; Zeng, X.-A.; Sun, D.-W.; Brennan, C.S.; Zhou, Q.-X.; Zhou, J.-S. The probiotic role of Lactobacillus plantarum in reducing risks associated with cardiovascular disease. Int. J. Food Sci. Technol. 2017, 52, 127–136. [Google Scholar] [CrossRef]
- Wuyts, S.; van Beeck, W.; Allonsius, C.N.; van den Broek, M.F.; Lebeer, S. Applications of plant-based fermented foods and their microbes. Curr. Opin. Biotechnol. 2020, 61, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Caro, G.; Oliver, C.M.; Weerakkody, R.; Singh, T.; Conlon, M.; Borges, G.; Sanguansri, L.; Lockett, T.; Roberts, S.A.; Augustin, A.C.M.A. Chronic administration of a microencapsulated probiotic enhances the bioavailability of orange juice flavanones in humans. Free Radic. Biol. Med. 2015, 84, 206–214. [Google Scholar] [CrossRef]
- Valero-Cases, E.; Roy, N.C.; Frutos, M.J.; Anderson, R.C. Influence of the fruit juice carriers on the ability of Lactobacillus plantarum DSM20205 to improve in vitro intestinal barrier integrity and its probiotic properties. J. Agric. Food Chem. 2017, 65, 5632–5638. [Google Scholar] [CrossRef]
- Ren, T.; Zhu, J.; Zhu, L.; Cheng, M. The combination of blueberry juice and probiotics ameliorate non-alcoholic steatohepatitis (NASH) by affecting SREBP-1c/PNPLA-3 pathway via PPAR-α. Nutrients 2017, 9, 198. [Google Scholar] [CrossRef] [Green Version]
- Gaanappriya, M.; Guhankumar, P.; Kiruththica, V.; Santhiya, N.; Anita, S. Probiotication of fruit juices by Lactobacillus acidophilus. Int. J. Adv. Biotechnol. Res. 2013, 4, 72–77. [Google Scholar]
- Shah, N.P.; Ding, W.K.; Fallourd, M.J.; Leyer, G. Improving the stability of probiotic bacteria in model fruit juices using vitamins and antioxidants. J. Food Sci. 2010, 75, 5. [Google Scholar] [CrossRef]
- Rúa, J.; López-Rodríguez, I.; Sanz, J.; García-Fernández, M.C.; del Valle, M.P.; García-Armesto, M.R. Improving functional properties of ‘Piel de Sapo’ melon juice by addition of a Lippia citriodora natural extract and probiotic-type lactic acid bacteria. LWT 2018, 96, 75–81. [Google Scholar] [CrossRef]
- Haffner, F.B.; Pasc, A. Freeze-dried alginate-silica microparticles as carriers of probiotic bacteria in apple juice and beer. LWT 2018, 91, 175–179. [Google Scholar] [CrossRef]
- Vivek, K.; Mishra, S.; Pradhan, R.C. Characterization of spray dried probiotic Sohiong fruit powder with Lactobacillus plantarum. LWT 2020, 117, 108699. [Google Scholar] [CrossRef]
- Tsen, J.-H.; Lin, Y.-P.; Huang, H.-Y.; King, V.A.-E. Studies on the fermentation of tomato juice by using κ-carrageenan immobilized Lactobacillus acidophilus. J. Food Process. Preserv. 2008, 32, 178–189. [Google Scholar] [CrossRef]
- Olivares, A.; Soto, C.; Caballero, E.; Altamirano, C. Survival of microencapsulated Lactobacillus casei (prepared by vibration technology) in fruit juice during cold storage. Electron. J. Biotechnol. 2019, 42, 42–48. [Google Scholar] [CrossRef]
- Šárka, H.; Kristýna, R.; Kristina, B.; Iveta, K.; Marcela, S. Fruit juices with probiotics—New type of functional foods. Czech J. Food Sci. 2018, 36, 284–288. [Google Scholar] [CrossRef] [Green Version]
- Sivudu, S.N.; Ramesh, B.; Umamahesh, K.; Vijaya Sarathi Reddy, O. Probiotication of tomato and carrot juices for shelf-life enhancement using micro-encapsulation. J. Food Biosci. Technol. 2016, 6, 13–22. [Google Scholar]
- Dias, C.O.; de Almeida, J.D.S.O.; Pinto, S.S.; de Oliveira Santana, F.C.; Verruck, S.; Müller, C.M.O.; Schwinden Prudêncio, E.; de Mello Castanho Amboni, R.D. Development and physico-chemical characterization of microencapsulated bifidobacteria in passion fruit juice: A functional non-dairy product for probiotic delivery. Food Biosci. 2018, 24, 26–36. [Google Scholar] [CrossRef]
- Santos, E.; Andrade, R.; Gouveia, E. Utilization of the pectin and pulp of the passion fruit from Caatinga as probiotic food carriers. Food Biosci. 2017, 20, 56–61. [Google Scholar] [CrossRef]
- Miranda, R.F.; de Paula, M.M.; da Costa, G.M.; Barão, C.E.; da Silva, A.C.R.; Raices, R.S.L.; Gomes, R.G.; Pimentel, T.C. Colombo Pimentel, Orange juice added with L. casei: Is there an impact of the probiotic addition methodology on the quality parameters? LWT 2019, 106, 186–193. [Google Scholar] [CrossRef]
- Gerardi, C.; Tristezza, M.; Giordano, L.; Rampino, P.; Perrotta, C.; Baruzzi, F.; Capozzi, V.; Mita, G.; Grieco, F. Exploitation of Prunus mahaleb fruit by fermentation with selected strains of Lactobacillus plantarum and Saccharomyces cerevisiae. Food Microbiol. 2019, 84, 103262. [Google Scholar] [CrossRef]
- Lu, Y.; Putra, S.D.; Liu, S.-Q. A novel non-dairy beverage from durian pulp fermented with selected probiotics and yeast. Int. J. Food Microbiol. 2018, 265, 1–8. [Google Scholar] [CrossRef]
- Ávila, M.; Hidalgo, M.; Sánchez-Moreno, C.; Pelaez, C.; Requena, T.; de Pascual-Teresa, S. Bioconversion of anthocyanin glycosides by Bifidobacteria and Lactobacillus. Food Res. Int. 2009, 42, 1453–1461. [Google Scholar] [CrossRef]
- Esteban-Torres, M.; Landete, J.M.; Reverón, I.; Santamaría, L.; de las Rivas, B.; Muñoz, R. A Lactobacillus plantarum esterase active on a broad range of phenolic esters. Appl. Environ. Microbiol. 2015, 81, 3235–3242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santamaría, L.; Reverón, I.; de Felipe, F.L.; de las Rivas, B.; Muñoz, R. Unravelling the reduction pathway as an alternative metabolic route to hydroxycinnamate decarboxylation in Lactobacillus plantarum. Appl. Environ. Microbiol. 2018, 84, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santamaría, L.; Reverón, I.; de Felipe, F.L.; de las Rivas, B.; Muñoz, R. Ethylphenol formation by Lactobacillus plantarum: Identification of the enzyme involved in the reduction of vinylphenols. Appl. Environ. Microbiol. 2018, 84, 17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mozzi, F.; Ortiz, M.E.; Bleckwedel, J.; de Vuyst, L.; Pescuma, M. Metabolomics as a tool for the comprehensive understanding of fermented and functional foods with lactic acid bacteria. Food Res. Int. 2013, 54, 1152–1161. [Google Scholar] [CrossRef]
- Singh, D.; Lee, S.; Lee, C.H. Metabolomics for empirical delineation of the traditional Korean fermented foods and beverages. Trends Food Sci. Technol. 2017, 61, 103–115. [Google Scholar] [CrossRef]
- Tomita, S.; Saito, K.; Nakamura, T.; Sekiyama, Y.; Kikuchi, J. Rapid discrimination of strain-dependent fermentation characteristics among Lactobacillus strains by NMR-based metabolomics of fermented vegetable juice. PLoS ONE 2017, 12, e0182229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chun, B.H.; Kim, K.H.; Jeon, H.H.; Lee, S.H.; Jeon, C.O. Pan-genomic and transcriptomic analyses of Leuconostoc mesenteroides provide insights into its genomic and metabolic features and roles in kimchi fermentation. Sci. Rep. 2017, 7, 11504. [Google Scholar] [CrossRef] [Green Version]
- Filannino, P.; di Cagno, R.; Gobbetti, M. Metabolic and functional paths of lactic acid bacteria in plant foods: Get out of the labyrinth. Curr. Opin. Biotechnol. 2018, 49, 64–72. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Processing Conditions | Raw Material | Outputs | Reference |
|---|---|---|---|
| Thermal Treatments Prior to Fermentation | |||
| Sterilization (120 °C/5 min); fermentation (30 or 37 °C) Lactobacillus fermentum and Lactobacillus casei; cold storage (4 weeks) | Watermelon and tomato | Different survival of bacteria during the cold storage period depending on fermentation temperature | [54] |
| Homogenization; heating (100 °C/5, 30, 60, 90 and 120 min); fermentation (Bifidobacterium brevis, Bifidobacterium longum, and Bifidobacterium infantis) | Fresh tomato | No heating impact on pH, Brix, acidity of juice; redness increases until 60 min heating; heating increases lycopene content; fermentation did not affect lycopene content | [55] |
| Commercial tomato juice (pasteurized); fermentation (Lactobacillus acidophilus LA39, L. casei A4, Lactobacillus. delbrueckii D7, Lactobacillus plantarum); storage (4 °C/4 weeks) | Commercial tomato juice | Viability of L. acidophilus and L. delbrueckii did not decrease during cold storage of fermented juice (106 to 108 CFU/mL) | [39] |
| Extraction; autoclave sterilization (121 °C/15 min); fermentation; cold storage for 4 weeks | Red beets | The fermented juice contained a significant load of beneficial viable cells for LAB (109 CFU/mL) | [37] |
| Centrifugal juice extractor; pasteurization (75 °C/5 min); fermentation (25 °C/48 h) by commercial water kefir preparation of 109 UFC/mL containing (Lactobacillus, Lactococcus, Leuconostoc and Saccharomyces) | Carrots, fennels, melons, onions, tomatoes | Thermal treatment reduced all microbial groups at levels below the detection limits; physicochemical and organoleptic attributes of fermented juice were acceptable; changes in antioxidant activities | [56] |
| Pre-treatments (filtration connected or not with thermal treatment and/or enzymatic processing) of juices prior to fermentation | Pomegranate | A short pasteurization at 95 °C/30–60 s eliminated all microorganisms found in juice ➔ Preservation of bioactive molecules + less anthocyanin degradation + improved antioxidant activities; clarification was related to reduction of phenol content and lower antioxidant activity | [57] |
| Combination between Thermal and Non-Thermal Treatments | |||
| Drying of raw material (40 °C/3 days); sterilization of powder suspension (121 °C/15 min); fermentation by Bacillus amyloliquefaciens and Lactobacillus brevis separately; light illumination (LED, sunlight and dark) | Blueberry | Light illumination enhanced metabolic activity + maximal LAB growth + highest TPC and flavonoids; blue LED mediated fermentation ➔ an extract with antibacterial activity only; Red LED mediated fermentation ➔ extract with antioxidant (phenols and flavonoids) and cytotoxic activities | [58] |
| Ultrasonicated homogenization (376 W/cm2, 500 W, 10 min); of L. casei); fermentation (24 h; 10 41 °C); cold storage (4 °C/42 days) | Pineapple | Ultrasonication of pineapple juice improved LAB viability and prevented browning; fermented juice exhibited acceptance for color and sensory evaluation | [59] |
| Heating (95 °C/1 min); High Hydrostatic Pressure: HHP (500 MP/2 min; 25 °C); fermentation of L. casei (105 CFU/mL) | Litchi | HHP fermented juice exhibited better color, flavor and overall acceptance than heated one; both HHP and Heated fermented litchi juice showed enhanced cell viability | [60] |
| Bleaching (100 °C/2 min), squeezing, filtering; thermal processing TP (100 °C/30 s); HHP (300, 400, 500 MP/15 min; 25 °C); storage (20 °C) until use; fermentation (Lactobacillus rhamnosus (30 °C/18h), Gluconacetobacter xylinus (30 °C/24 h; 160 rpm)) | Yacon, Litchi and Longan (YLL) | TP (100 °C/30 s) and HHP (500 MPa/15 min) of juice complied with aseptic standards and ensured microbial safety; TP reduced the taste score of juice and reduced flavor molecules (alcohols, esters,…); HPP preserved the nutritional and aroma characteristics of juice; fermentation of HHP juice improved the taste (aminoacids, carbohydrates,…) and flavors of YLL juice; HHP cofermentation improved the quality of YLL juice | [61] |
| Raw Material | Pre-Fermentative Treatment | Microorganism | Nutritional Impact | Reference |
|---|---|---|---|---|
| Water kefir beverages | ||||
| Cocoa pulp | - | Kefir grains | - | [92] |
| Kiwifruit, quince, pomegranate, grape, apple, cactus pear | Heat treatment 75 °C 5 min | Kefir grains | Decrease in DPPH 1 activity | [93] |
| Carrot, fennel, melon, onion, tomato, strawberry | Heat treatment 75 °C 5 min | Kefir grains | - | [56] |
| Vegetable-based beverages | ||||
| Tomato juice | Pasteurization (commercial) | L. acidophilus, L. casei, L. delbrueckii, L. plantarum | - | [39] |
| Tomato juice | Sucrose addition, Heat treatment 80 °C 5 min | L. plantarum, W. cibaria/confusa, L. brevis, P. pentosaceus, Lactobacillus sp., Enterococcus faecium/faecalis | Lower decrease in ascorbic acid content, glutathione content and total antioxidant activity in juices started with L. plantarum compared to other conditions | [28] |
| Red beetroot juice | ||||
| Heat treatment 121 °C 15 min | L. acidophilus, L. casei, L. delbrueckii, L. plantarum | - | [37] | |
| Cabbage juice | Heat treatment 121 °C 15 min | L. plantarum, L. casei, L. delbrueckii | - | [38] |
| Celery and beetroot juice | Heat treatment 120 °C 15 min | Bifidobacterium animalis subsp. lactis | - | [94] |
| Carrot | Heat treatment 80 °C 20 min | B. lactis, B. bifidum | Decrease of 10–20% of β-carotene and 5–25% of α-carotene | [95] |
| Bitter gourd, bottle gourd, carrot | Heat treatment 80 °C 20 min | L. plantarum | - | [96] |
| Fruit-based juices | ||||
| Pineapple | Filtration 0.22 µm, heat treatment 75 °C 15 s | L. plantarum, L. rossiae | Slight increase in antioxidant DPPH activity for started juices | [97] |
| Pineapple | Sonication | L. casei | - | [59] |
| Pineapple, papaya, mango, green tea, black tea | Commercial pasteurized pineapple juice; 75 °C 5 min for papaya and mango; infusion 5 min in boiling water for tea | W. cibaria, W. confusa, W. paramesenteroides, W. soli, L. mesenteroides, L. pseudomesenteroides, L. citreum, Leuconostoc lactis, L. paraplantarum, L. plantarum | Increase in total phenolic content and DPPH activity with several strains on papaya and pineapple | [5] |
| Pomegranate | Heat treatment 80 °C 5 min | L. plantarum, L. acidophilus, L. paracasei, L. delbrueckii | Increase in DPPH radical scavenging activity; decrease in anthocyanin content; increase in antioxidant activity more marked with L. acidophilus than with L. plantarum | [98,99] |
| Pomegranate | Filtration 0.22 µm | L. plantarum | Total phenolic compounds concentration and free radical scavenging activity were better maintained in fermented juice than in control; fermented juice modulated the synthesis of the immune mediators into peripheral blood mononuclear cells | [100] |
| Pomegranate | - | L. acidophilus, L. plantarum, B. longum subsp. infantis, B. bifidum | Identification of catechin and α-punicalagin in fermented juices; increase in antioxidant activity and concentration of phenolic derivatives during simulated gastrointestinal digestion of fermented juices; fermentation increased bioaccessibility of phenolic compounds | [101] |
| Pomegranate | Heat treatment 70 °C 5 min | L. plantarum, L. acidophilus, L. delbrueckii | Higher antioxidant activity and total phenolic compounds in juices fermented with L. plantarum compared to unfermented | [102] |
| Noni | Heat treatment 121 °C 15 min | L. casei subsp. casei, B. longum, L. plantarum | Antioxidant activity of juice fermented with B. longum was better maintained than with other strains and remained similar to initial non fermented juice | [103] |
| Cherries, tomato, blackberry, prune, white grape juice and aloe vera Kiwifruit, fennels, spinach, papaya, white grape juice and aloe vera | Heat treatment 80 °C 10 min | L. plantarum, L. pentosus, Lactobacillus sp. | Better maintain of radical scavenging activity and total phenolic compound concentration in started juices | [104] |
| Pear | - | L. acidophilus | Significant increase in α-glucosidase inhibition for most samples of fermented juice | [105] |
| Cherry juice and broccoli purée | Cherry juice was heat treated at 121 °C for 10 min | L. plantarum | Fermented cherry juice and broccoli puree were enriched in phenolic derivative with high human bioavailability and biological activity | [31] |
| Coconut water | - | L. plantarum, B. animalis subsp. lactis | - | [106] |
| Apple, grape or orange juices | Heat treatment 80 °C 20 min | L. acidophilus, L. casei, L. rhamnosus, L. paracasei | - | [14] |
| Mango | Addition of 50 mg/L potassium metabisulfite and pectinolytic enzymes, pasteurization | L. acidophilus, L. delbrueckii, L. plantarum, L. casei | - | [107] |
| Cactus pear | Heat treatment 70 °C 5 min | L. mesenteroides | Increase in free radical scavenging activity; started fermented juice markedly inhibited the inflammatory status of Caco-2/TC7 cells; started fermented juice scavenged the reactive oxygen species generated by H2O2 on Caco-2 cells; fermented juices variously affected the immunomodulatory activity towards anti- and pro-inflammatory cytokines | [108] |
| Cactus pear | Dilution 2:1 in water | L. fermentum | No change in total phenolic content during fermentation; decrease in antioxidant activity during fermentation | [109] |
| Cactus pear | Heat treatment 64 °C 30 min | L. plantarum | Maintain of health-promoting properties in fermented cactus pear juice; fermented juice markedly promoted antioxidant mechanisms in yeast cells; significant decrease in the body weight gain and amelioration of the insulin resistance, hyperglycemia, and hyperlipemia after feeding obese mice with fermented juice | [110] |
| Carambola, guava, mango, pitaya | Heat treatment 80 °C 5 min | L. casei, S. thermophilus, L. bulgaricus | - | [16] |
| Sweet lemon juice | Heat treatment 80 °C 5 min | L. plantarum | No effect of fermented juice on ascorbic acid; decrease in total phenolic compound concentration; higher antioxidant activity (DPPH and FRAP 2) in fermented juice during 28 days of storage compared to unfermented juice | [111] |
| Cashew apple juice | Heat treatment 70 °C 15 min | L. plantarum, L. casei, L. acidophilus | Decrease in hydrolysable tannin content during fermentation; juice fermented with L. plantarum retained better free radical scavenging activity (DPPH and ABTS 3) than with other strains | [112] |
| Sohiong juice | Heat treatment 70 °C 5 min | L. acidophilus, L. plantarum | - | [113] |
| Apple, carrot, tomato, cucumber, haw (40:25:15:15:5) | Heat treatment 80 °C 20 min | L. plantarum, L. casei, L. acidophilus, L. fermentum, L. rhamnosus | - | [114] |
| Autochthonous Strains | Allochthonous Strains |
|---|---|
| Highest viscosity of the juice [10] Highest values of ascorbic acid, glutathione and total antioxidant activity during storage [10] Rapid decrease in pH [28] Marked consumption of fermentable carbohydrates [28] Inhibition of Enterobacteriaceae and yeasts [28] Positively affected firmness and color [28] | Longer latency phases of growth and acidification [10] |
| Juices | Compounds Metabolized | Compounds Released | LAB Species | Ref. |
|---|---|---|---|---|
| Apple | QA, Q, Q-3-gal, Q-3-glc, phlorizin | 5-O-caffeoylquinic acid, quercetin, phloretin | L. plantarum | [125] |
| Broccoli | CGA, QA | PA, sinapic acid | L. reuteri | [31] |
| Sinapic acid | PA | L. plantarum or L. mesenteroides | [128] | |
| Black chokeberry | CA, CGA, neochlorogenic acid | Anthocyanins: C-3-gal, C-3-glc, C-3-xyl | L. plantarum | [121] |
| Cherry | PTA, CA | catechol, DCA | Lactobacillus spp. | [31] |
| Elderberry | PTA, CA | DCA, catechol, flavonols glycosides (Q-3-rut, Q-3-glc) and anthocyanins (C-3-samb, C-3-glc) | L. plantarum, L. rhamnosus and L. casei | [129] |
| Momordica charantia L. | CA, p-CA, p-HBA, vinylphenol, catechol, PTA, BA, vanillin | DCA, phloretic acid, vinylcatechol, pyrogallol | L. plantarum | [123] |
| Mulberry | Flavonols (catechin, rutin, morin, quercetin, kaempferol) Anthocyanins (C-3-glc, C-3-O-rut, esculin) Phenolic acids (gallic acid, syringic acid, vanillic acid, quinic acid, ferulic acid, cinnamic acid, PTA, CGA, CA, p-HBA, p-CA) | L. plantarum, L. acidophilus or L. paracasei | [124] | |
| Sea buckthorn | Q-3-glc, I-3-glc | PTA | L. plantarum | [121] |
| Processing Conditions | Raw Material | Mains Findings | Reference |
|---|---|---|---|
| Bleaching (60 °C/20 min), extraction, clarification (muslin cloth); autoclave sterilization (121 °C/15 min); encapsulation of Lactobacillus spp and S. cerevisiae using alginate coated chitosan beads; cold storage (6 weeks) | Tomato and carrot | Survivability of encapsulated probiotic cells in a blended juice of tomato-carrot fruit was improved; overall sensory acceptance (taste profile, mouthfeel, acidity, flavor, aroma, color) | [54] |
| Filter sterilization; immobilization of L. acidophilus in an alginate CaCl2 beads | Grapes, orange, watermelon | Immobilized cultures remained viable over a long period of time; probiotically drinks were potentially inhibiting the pathogenic growth | [159] |
| Model juice (sucrose, sodium citrate, citric acid powder, distilled water); water bath pasteurization (85 °C/10 min); enrichment by antioxidants (grape seed and green tea extracts) and vitamins (B, C); probiotication by (L. rhamnosus, B. lactis, L. paracasei); storage (6 weeks) | Model juice | Turbidity and viscosity of model juice increases ➔ do not support growth of probiotics; combined effect of probiotics and bioactive ingredients ➔ improve cells viability on model juice | [160] |
| Sterilized filtration (stainless steel strainer) of juice pulp; polyphenol-fortified melon juice; inoculation by LAB probiotics and incubation (4 °C/4 days) | Melon | Herbal extract based additive ➔ increases antioxidant activity; delivering viable LAB + survivability and growth into refrigerated enriched juice | [161] |
| Electrospraying assisted microencapsulation of L. rhamnosus ➔ Silica-coated alginate beads; freeze drying (85 °C; 0.2 mbar; 20 h) of microcapsules; addition of these microcarriers to pasteurized apple juice | Apple | Silica-coated beads diminished the leakage of L. rhamnosus when compared to alone alginate beads ➔ More protection and survival | [162] |
| Inoculation L. acidophilus of juice mixture (70:30 v/v); air spray drying (Tair: 120–160 °C; flow rate: 25 mL/min); with maltodextrin | Watermelon and carrot | Higher Tair + longer time + lower flow ➔ Reducing of cell viability, lycopene and β-carotene of dried juice (powder); increased atomization pressure ➔ Better cohesiveness but less flowability of juice powder | [81] |
| Ultrasonic (90%/9.95 min), pectinase (0.05% w/w) extraction of juice; sonicated pasteurization (15 min); fermentation by L. plantarum; Anticaking agent + maltodextrin; spray drying (concurrent, 120 °C, 201 mL/h); storage (25 °C, 50% RH) | Sohiong fruit (Indian blackberry fruit) | A decreasing trend of probiotics viability during storage ➔ This might be the result of stress during spray drying + thermal shock, oxygen and humidity exposure at ambient storage; dried probiotic juice could be maintained up to 36 days without packaging | [163] |
| Mixing (45 s); short pasteurization (95 °C/15 s); skin and seeds removal (mesh screen); juice was kept (88 °C/2 min); L. acidophilus cell immobilization k carrageenan beads; free and cell-immobilized fermentation of tomato juice; cold storage (4 °C/10 weeks) | Fresh raw tomato | Fermentation was improved by immobilized cells ➔ protection effect against adverse environment of tomato juice; immobilization increased growth and viability of L. acidophilus during fermentation and storage compared to the free cells; high overall palatability score (before and during cold storage) of juice | [164] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia, C.; Guerin, M.; Souidi, K.; Remize, F. Lactic Fermented Fruit or Vegetable Juices: Past, Present and Future. Beverages 2020, 6, 8. https://doi.org/10.3390/beverages6010008
Garcia C, Guerin M, Souidi K, Remize F. Lactic Fermented Fruit or Vegetable Juices: Past, Present and Future. Beverages. 2020; 6(1):8. https://doi.org/10.3390/beverages6010008
Chicago/Turabian StyleGarcia, Cyrielle, Marie Guerin, Kaies Souidi, and Fabienne Remize. 2020. "Lactic Fermented Fruit or Vegetable Juices: Past, Present and Future" Beverages 6, no. 1: 8. https://doi.org/10.3390/beverages6010008
APA StyleGarcia, C., Guerin, M., Souidi, K., & Remize, F. (2020). Lactic Fermented Fruit or Vegetable Juices: Past, Present and Future. Beverages, 6(1), 8. https://doi.org/10.3390/beverages6010008