Tamarisk and Russian Olive Occurrence and Absence Dataset Collected in Select Tributaries of the Colorado River for 2017

 , , , ,
, , , , 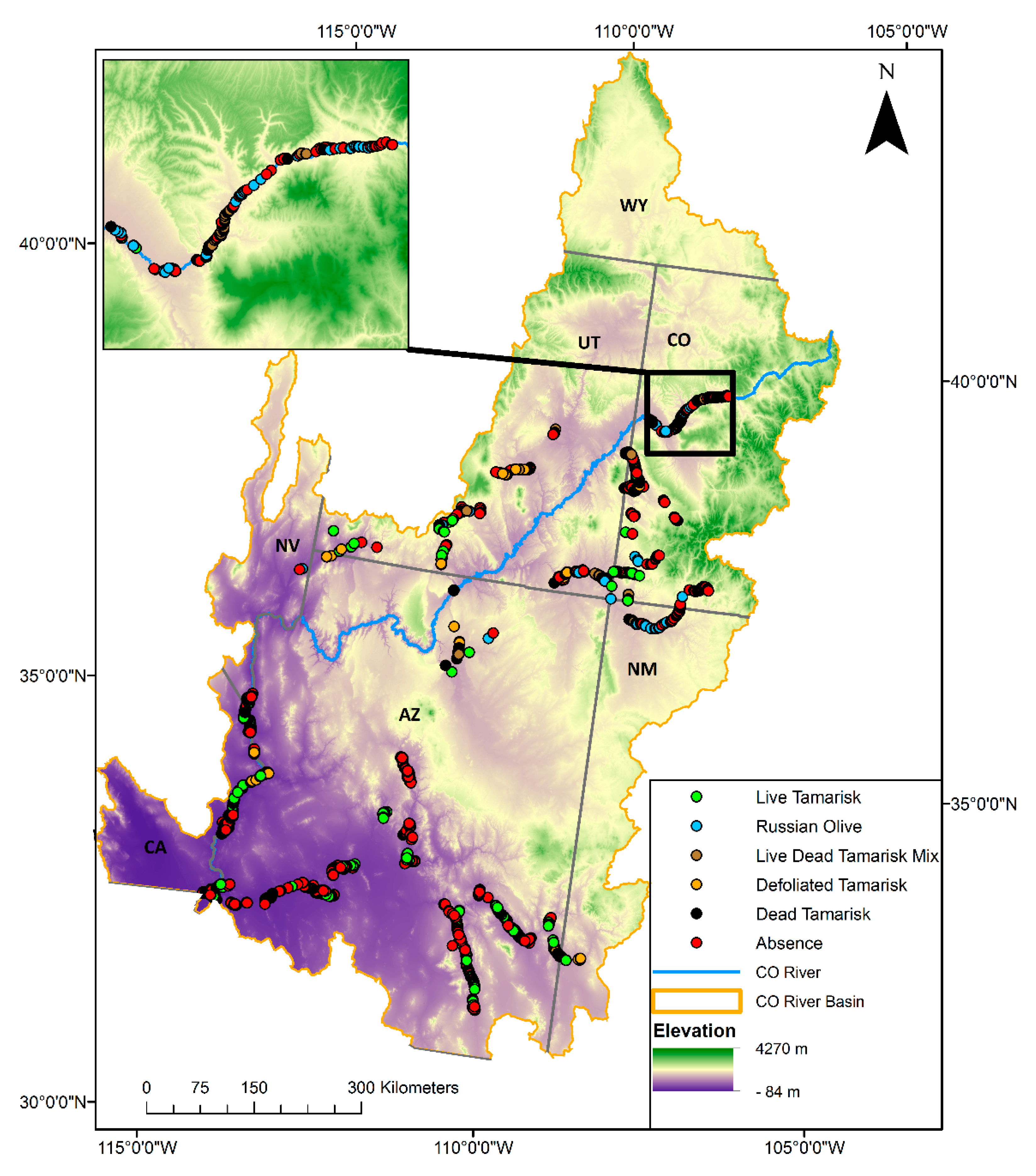{kind=link}
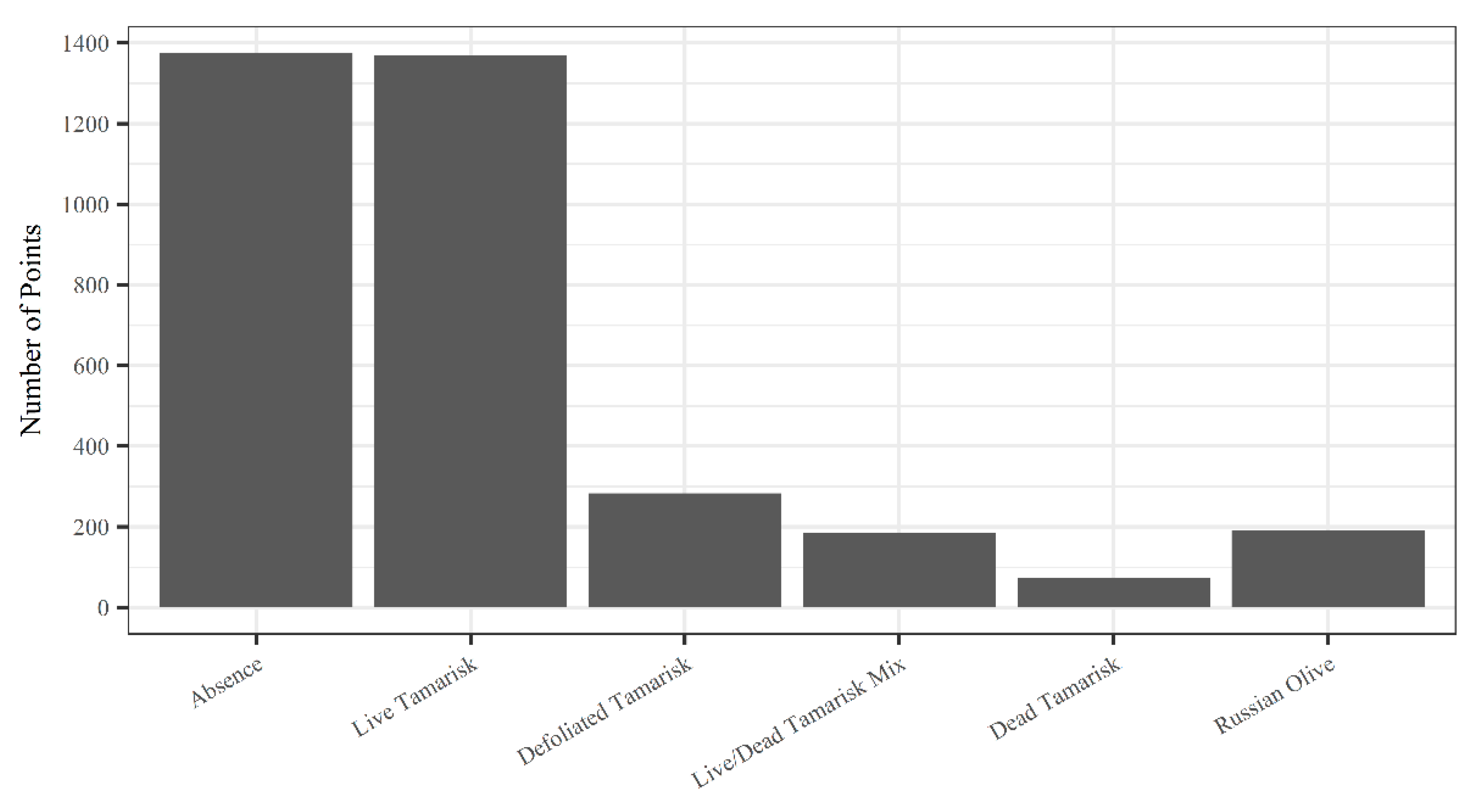{kind=link}
{kind=link}
Abstract
:1. Summary
2. Data Description
3. Methods
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Friedman, J.M.; Auble, G.T.; Shafroth, P.B.; Scott, M.L.; Merigliano, M.F.; Freehling, M.D.; Griffin, E.R. Dominance of non-native riparian trees in western USA. Biol. Invasions 2005, 7, 747–751. [Google Scholar] [CrossRef]
- Nagler, P.L.; Glenn, E.P.; Jarnevich, C.S.; Shafroth, P.B. Distribution and abundance of Saltcedar and Russian Olive in the Western United States. Crit. Rev. Plant Sci. 2011, 30, 508–523. [Google Scholar] [CrossRef]
- Carman, J.G.; Brotherson, J.D. Comparisons of Sites Infested and Not Infested with Saltcedar (Tamarix pentandra) and Russian Olive (Elaeagnus angustifolia). Weed Sci. 1982, 30, 360–364. [Google Scholar] [CrossRef]
- Sher, A.A.; Marshall, D.L.; Taylor, J.P. Establishment patterns of native Populus and Salix in the presence of invasive nonnative Tamarix. Ecol. Appl. 2002, 12, 760–772. [Google Scholar] [CrossRef]
- Meinhardt, K.A.; Gehring, C.A. Disrupting mycorrhizal mutualisms: A potential mechanism by which exotic tamarisk outcompetes native cottonwoods. Ecol. Appl. 2012, 22, 532–549. [Google Scholar] [CrossRef] [PubMed]
- Stromberg, J.C. Functional equivalency of saltcedar (Tamarix chinensis) and fremont cottonwood (Populus fremontii) along a free-flowing river. Wetlands 1998, 18, 675–686. [Google Scholar] [CrossRef]
- Nagler, P.L.; Morino, K.; Didan, K.; Erker, J.; Osterberg, J.; Hultine, K.R.; Glenn, E.P. Wide-area estimates of saltcedar (Tamarix spp.) evapotranspiration on the lower Colorado River measured by heat balance and remote sensing methods. Ecohydrology 2009, 2, 18–33. [Google Scholar] [CrossRef]
- Fischer, R.A.; Valente, J.J.; Guilfoyle, M.P.; Michael, D.; Jackson, S.S.; Ratti, J.T. Bird Community Response to Vegetation Cover and Composition in Riparian Habitats Dominated by Russian Olive (Elaeagnus angustifolia). Northwest Sci. 2012, 86, 39–52. [Google Scholar] [CrossRef]
- Nelson, S.M.; Wydoski, R. Butterfly Assemblages Associated with Invasive Tamarisk (Tamarix spp.) Sites: Comparisons with Tamarisk Control and Native Vegetation Reference Sites. J. Insects 2013, 2013, 10. [Google Scholar] [CrossRef]
- Katz, G.L.; Shafroth, P.B. Biology, ecology and management of Elaeagnus angustifolia L. (Russian olive) in western North America. Wetlands 2003, 23, 763–777. [Google Scholar] [CrossRef]
- Merritt, D.M.; Leroy Poff, N. Shifting dominance of riparian Populus and Tamarix along gradients of flow alteration in western North American rivers. Ecol. Appl. 2010, 20, 135–152. [Google Scholar] [CrossRef] [PubMed]
- Bloodworth, B.R.; Shafroth, P.B.; Sher, A.A.; Manners, R.B.; Bean, D.W.; Johnson, M.J.; Hinojosa-Huerta, O. Tamarisk beetle (Diorhabda spp.) in the Colorado River Basin: Synthesis of an Expert Panel Forum; Scientific and Technical Report No. 1; Colorado Mesa University Ruth Powell Hutchins Water Center: Grand Junction, CO, USA, 2016. [Google Scholar]
- Hultine, K.R.; Dudley, T.L.; Koepke, D.F.; Bean, D.W.; Glenn, E.P.; Lambert, A.M. Patterns of herbivory-induced mortality of a dominant non-native tree/shrub (Tamarix spp.) in a southwestern US watershed. Biol. Invasions 2015, 17, 1729–1742. [Google Scholar] [CrossRef]
- Kennard, D.; Louden, N.; Gemoets, D.; Ortega, S.; González, E.; Bean, D.; Cunningham, P.; Johnson, T.; Rosen, K.; Stahlke, A. Tamarix dieback and vegetation patterns following release of the northern tamarisk beetle (Diorhabda carinulata) in western Colorado. Biol. Control 2016, 101, 114–122. [Google Scholar] [CrossRef]
- Morisette, J.T.; Jarnevich, C.S.; Ullah, A.; Cai, W.; Pedelty, J.A.; Gentle, J.E.; Stohlgren, T.J.; Schnase, J.L. A tamarisk habitat suitability map for the continental United States. Front. Ecol. Environ. 2006, 4, 11–17. [Google Scholar] [CrossRef]
- Jarnevich, C.S.; Reynolds, L.V. Challenges of predicting the potential distribution of a slow-spreading invader: A habitat suitability map for an invasive riparian tree. Biol. Invasions 2010, 13, 153–163. [Google Scholar] [CrossRef]
- Environmental Systems Research Institute (ESRI). World Imagery; ESRI: Redlands, CA, USA, 2017. [Google Scholar]
- Environmental Systems Research Institute (ESRI). Collector for ArcGIS Desktop; ESRI: Redlands, CA, USA, 2016. [Google Scholar]
- Stohlgren, T.J.; Barnett, D.T.; Simonson, S.E. Beyond North American Weed Management Association Standards. 2010. Available online: http://irmaservices.nps.gov/datastore/v4/rest/DownloadFile/434136?accessType=DOWNLOAD (accessed on 17 October 2018).
- West, A.M.; Evangelista, P.H.; Jarnevich, C.S.; Kumar, S.; Swallow, A.; Luizza, M.; Chignell, S. Using multi-date satellite imagery to monitor invasive grass species distribution in post-wildfire landscapes; an iterative, adaptable approach that employs open-source data and software. Int. J. Appl. Earth Obs. Geoinf. 2017, 59, 135–146. [Google Scholar] [CrossRef]
- Gaskin, J.F.; Schaal, B.A. Hybrid Tamarix widespread in U.S. invasion and undetected in native Asian range. Proc. Natl. Acad. Sci. USA 2002, 99, 11256–11259. [Google Scholar] [CrossRef] [PubMed]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vorster, A.G.; Woodward, B.D.; West, A.M.; Young, N.E.; Sturtevant, R.G.; Mayer, T.J.; Girma, R.K.; Evangelista, P.H. Tamarisk and Russian Olive Occurrence and Absence Dataset Collected in Select Tributaries of the Colorado River for 2017. Data 2018, 3, 42. https://doi.org/10.3390/data3040042
Vorster AG, Woodward BD, West AM, Young NE, Sturtevant RG, Mayer TJ, Girma RK, Evangelista PH. Tamarisk and Russian Olive Occurrence and Absence Dataset Collected in Select Tributaries of the Colorado River for 2017. Data. 2018; 3(4):42. https://doi.org/10.3390/data3040042
Chicago/Turabian StyleVorster, Anthony G., Brian D. Woodward, Amanda M. West, Nicholas E. Young, Robert G. Sturtevant, Timothy J. Mayer, Rebecca K. Girma, and Paul H. Evangelista. 2018. "Tamarisk and Russian Olive Occurrence and Absence Dataset Collected in Select Tributaries of the Colorado River for 2017" Data 3, no. 4: 42. https://doi.org/10.3390/data3040042
APA StyleVorster, A. G., Woodward, B. D., West, A. M., Young, N. E., Sturtevant, R. G., Mayer, T. J., Girma, R. K., & Evangelista, P. H. (2018). Tamarisk and Russian Olive Occurrence and Absence Dataset Collected in Select Tributaries of the Colorado River for 2017. Data, 3(4), 42. https://doi.org/10.3390/data3040042