
Differential Development of the Chordae Tendineae and Anterior Leaflet of the Bovine Mitral Valve


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue Harvest and Anatomical Dimensions
2.2. Collagen Biochemical Analysis
2.3. Histology
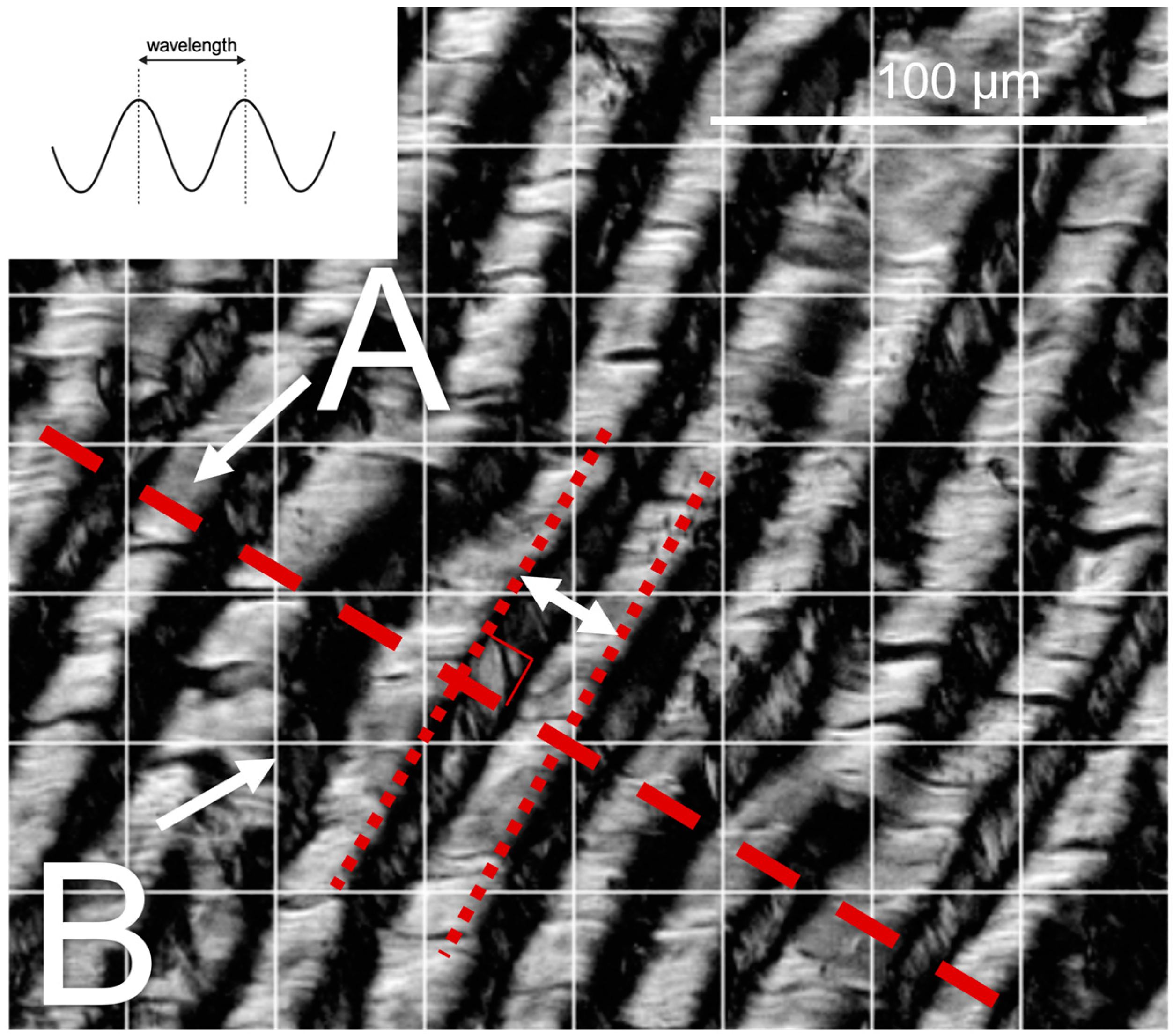
2.4. Collagen Crimp Analysis
2.5. Statistical Analysis
3. Results
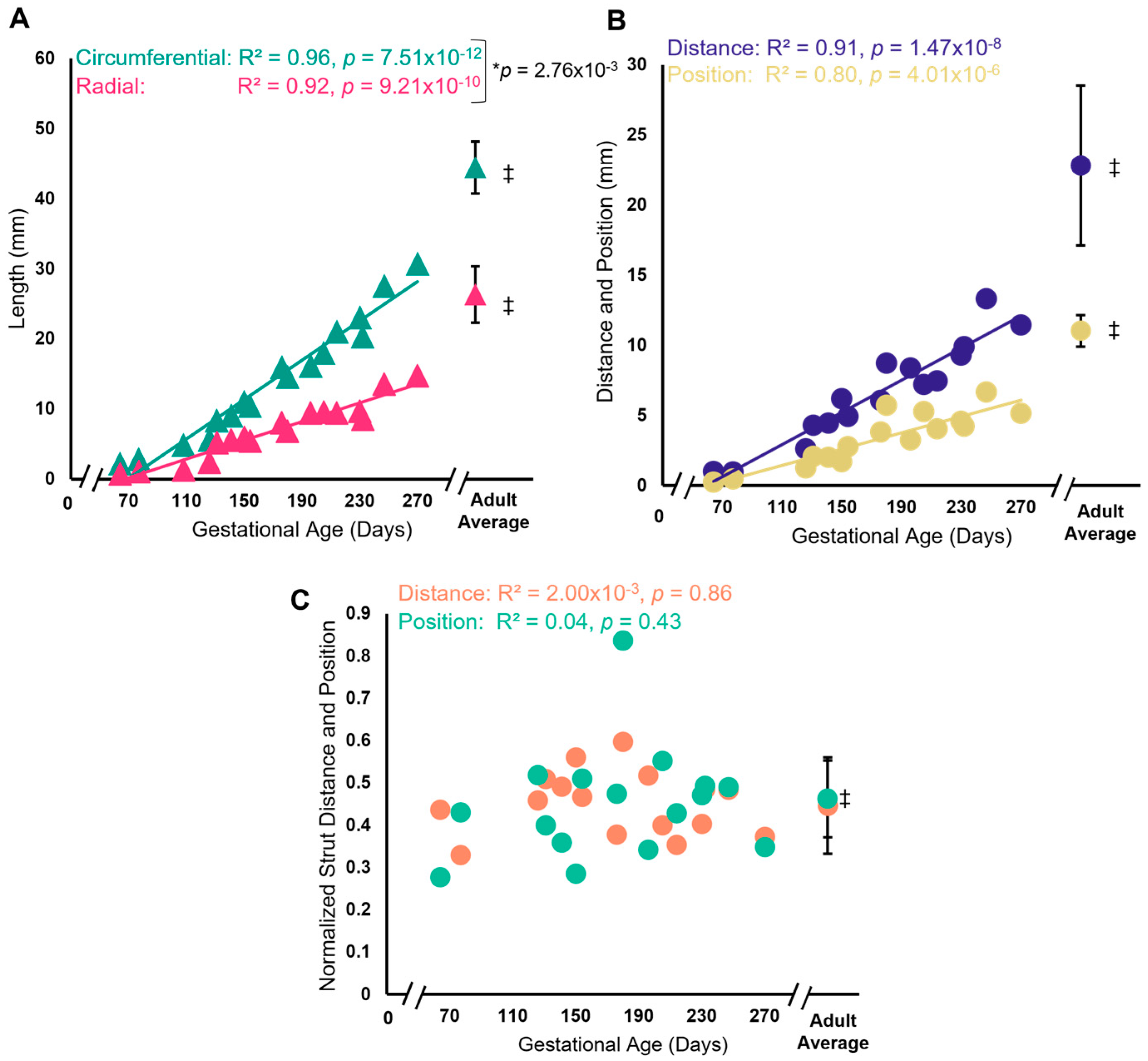
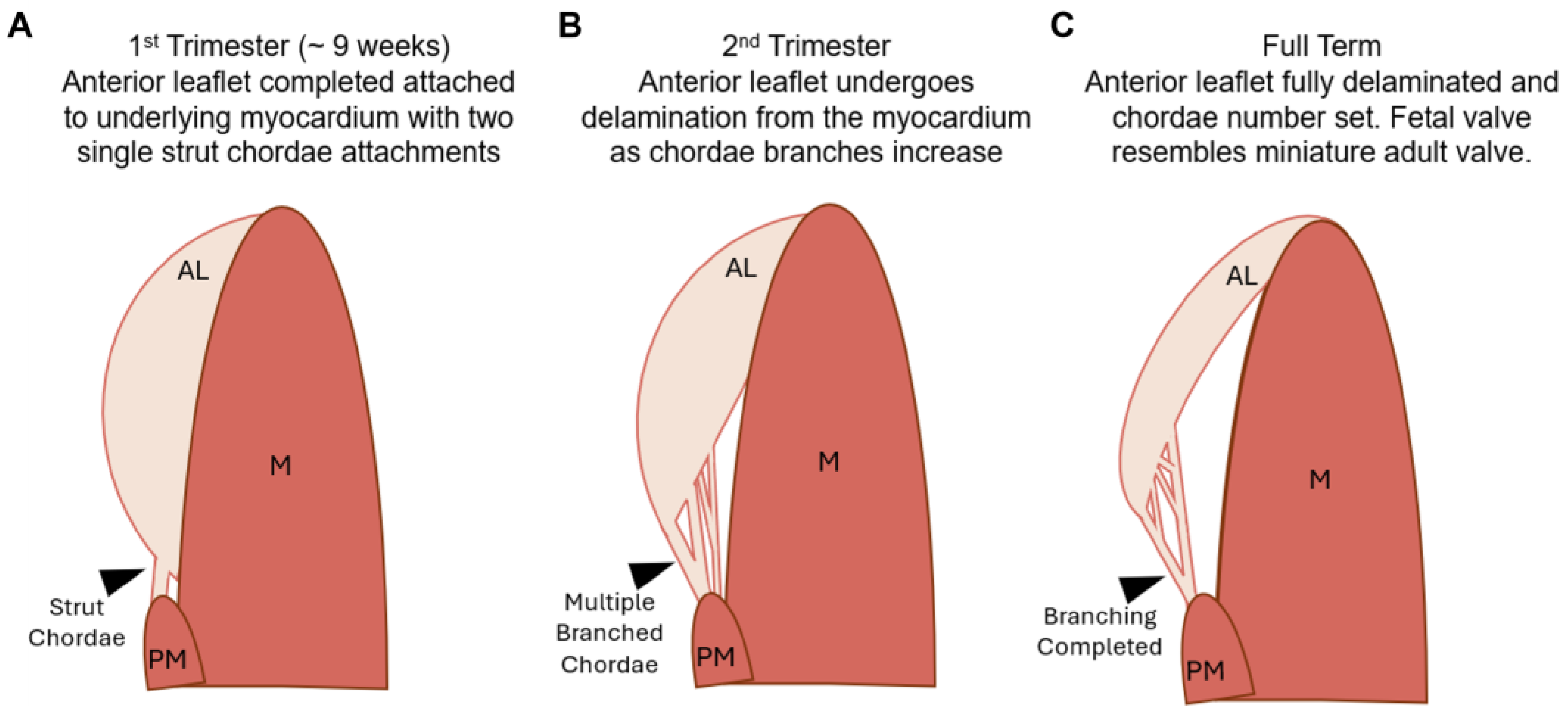
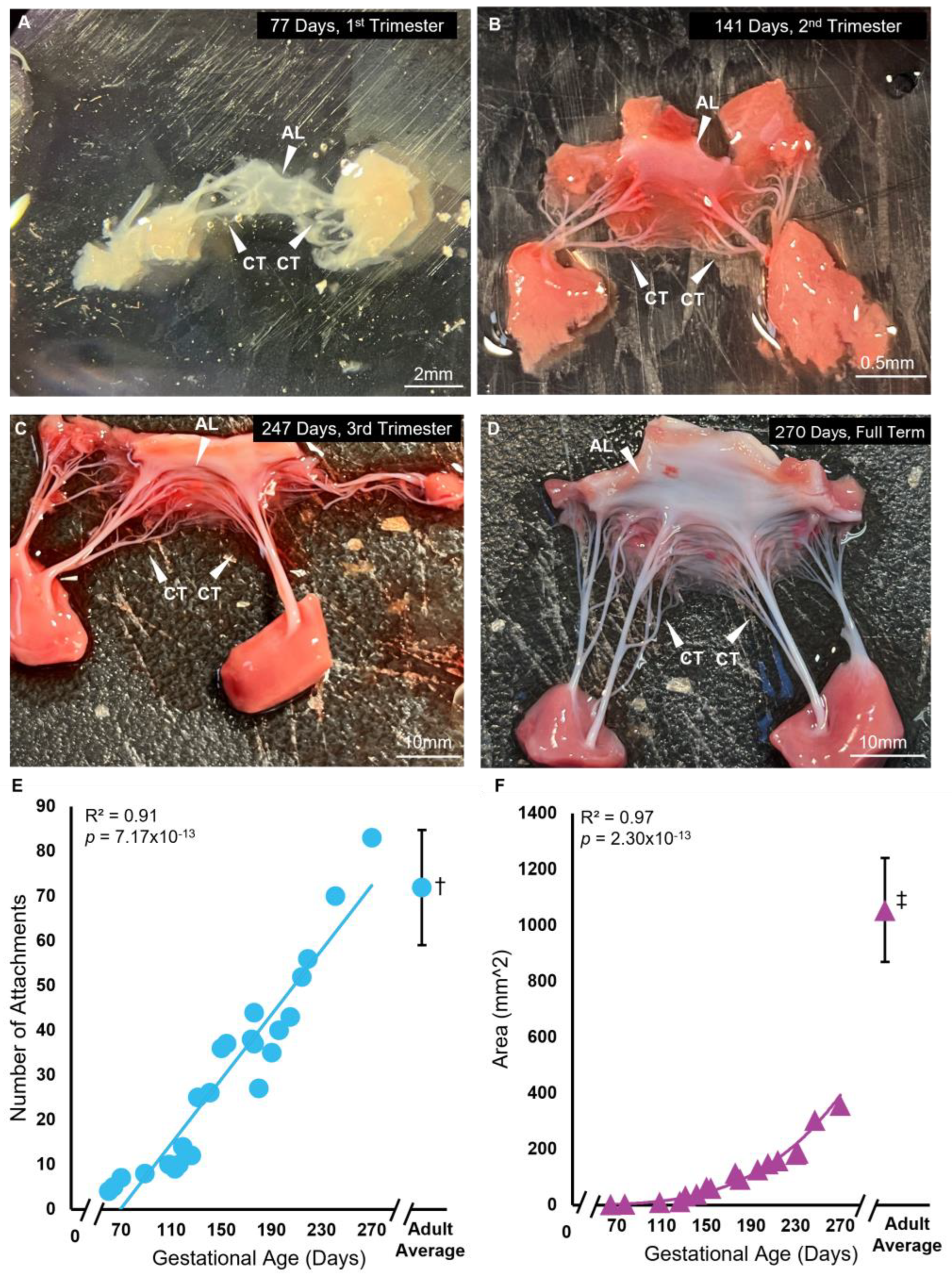
3.1. Chordae Division Occurs Only during Gestation Whereas Leaflet Area Continues to Increase Postnatally
3.2. In Leaflet and Chordae, Mature Collagen Content Increases over Gestation despite Unchanged Levels of Newly Synthesized Collagen
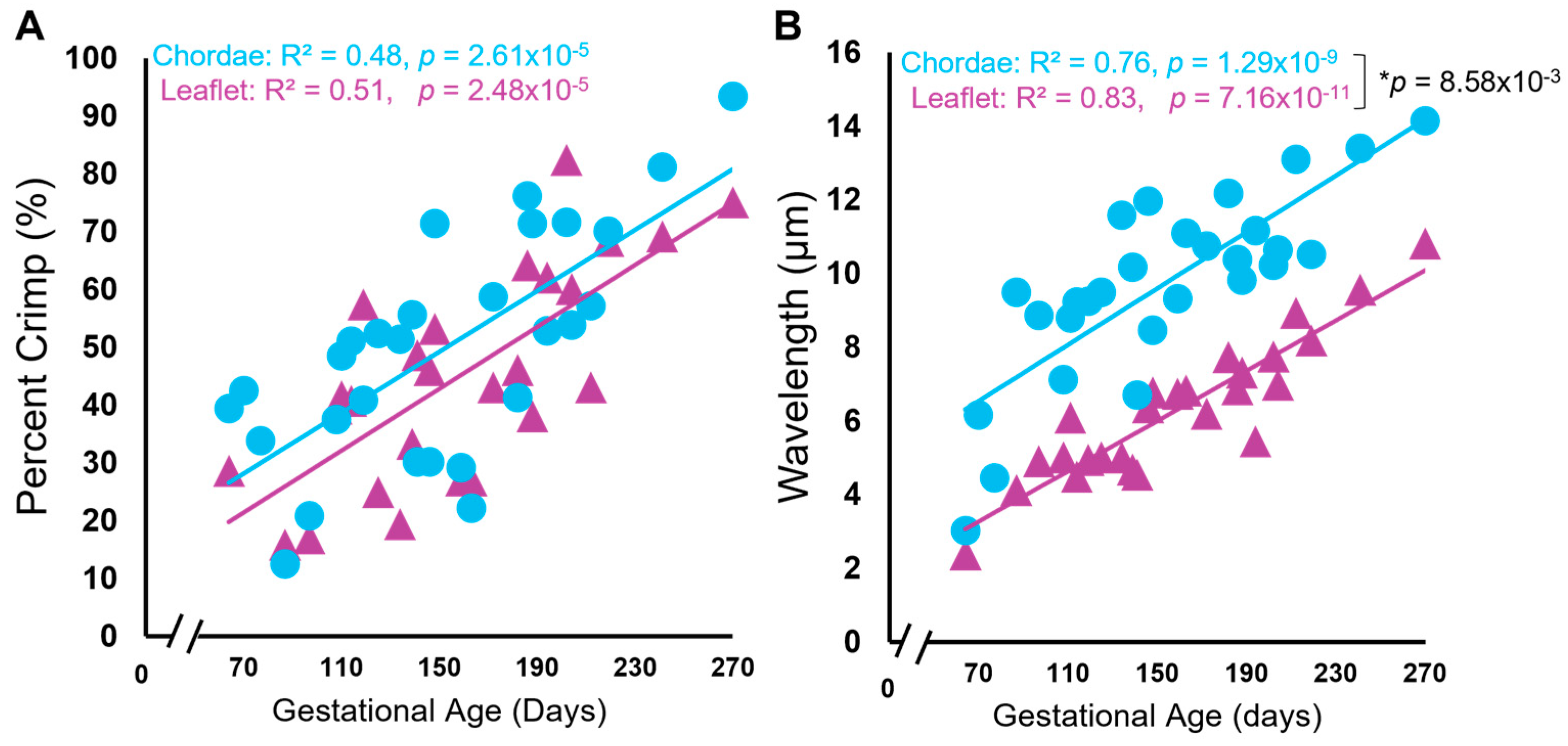
3.3. Collagen Crimp Develops Earlier in the Chordae versus the Leaflet
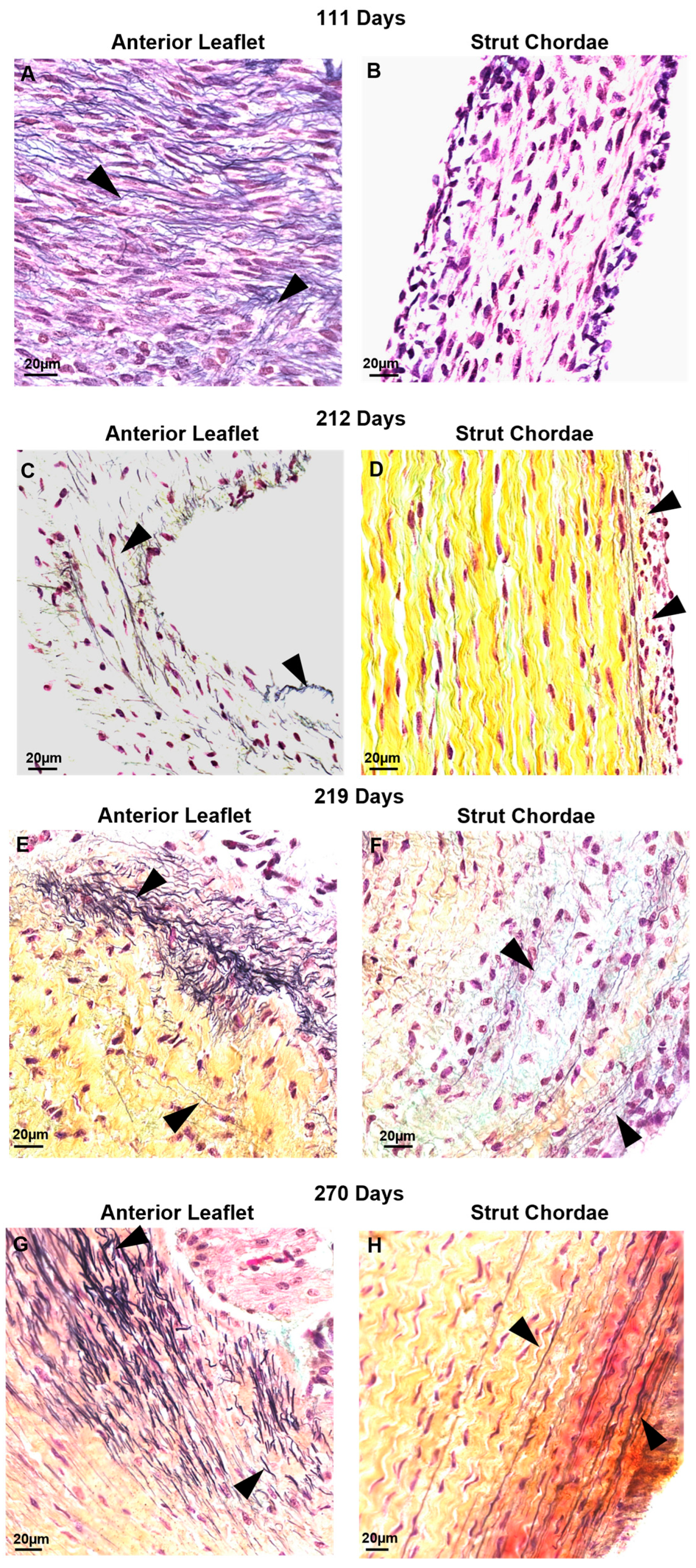
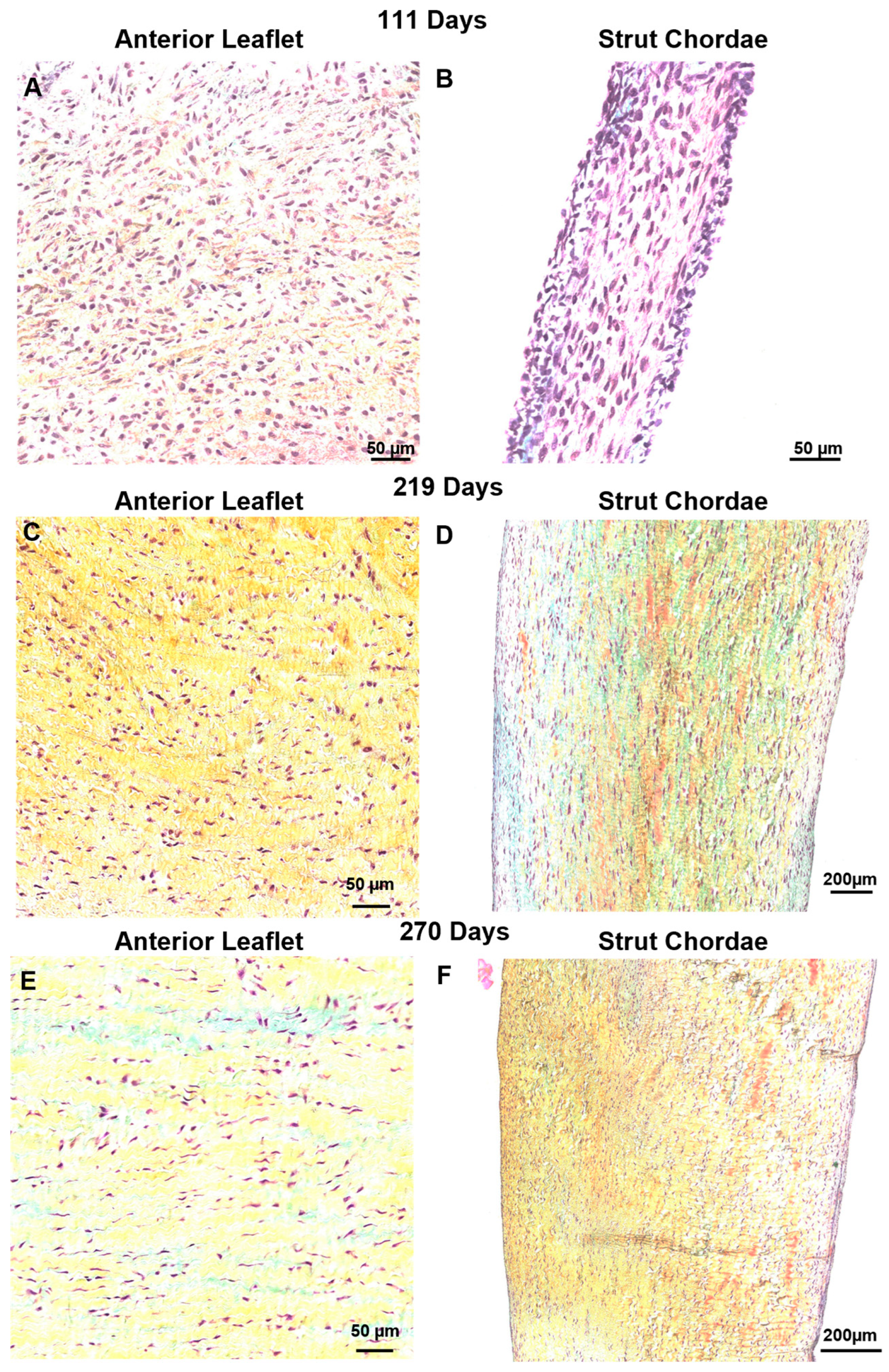
3.4. Bovine Mitral Chordae and Leaflet Collagen Fibers Are Laid Down in Their Adult-like Orientations
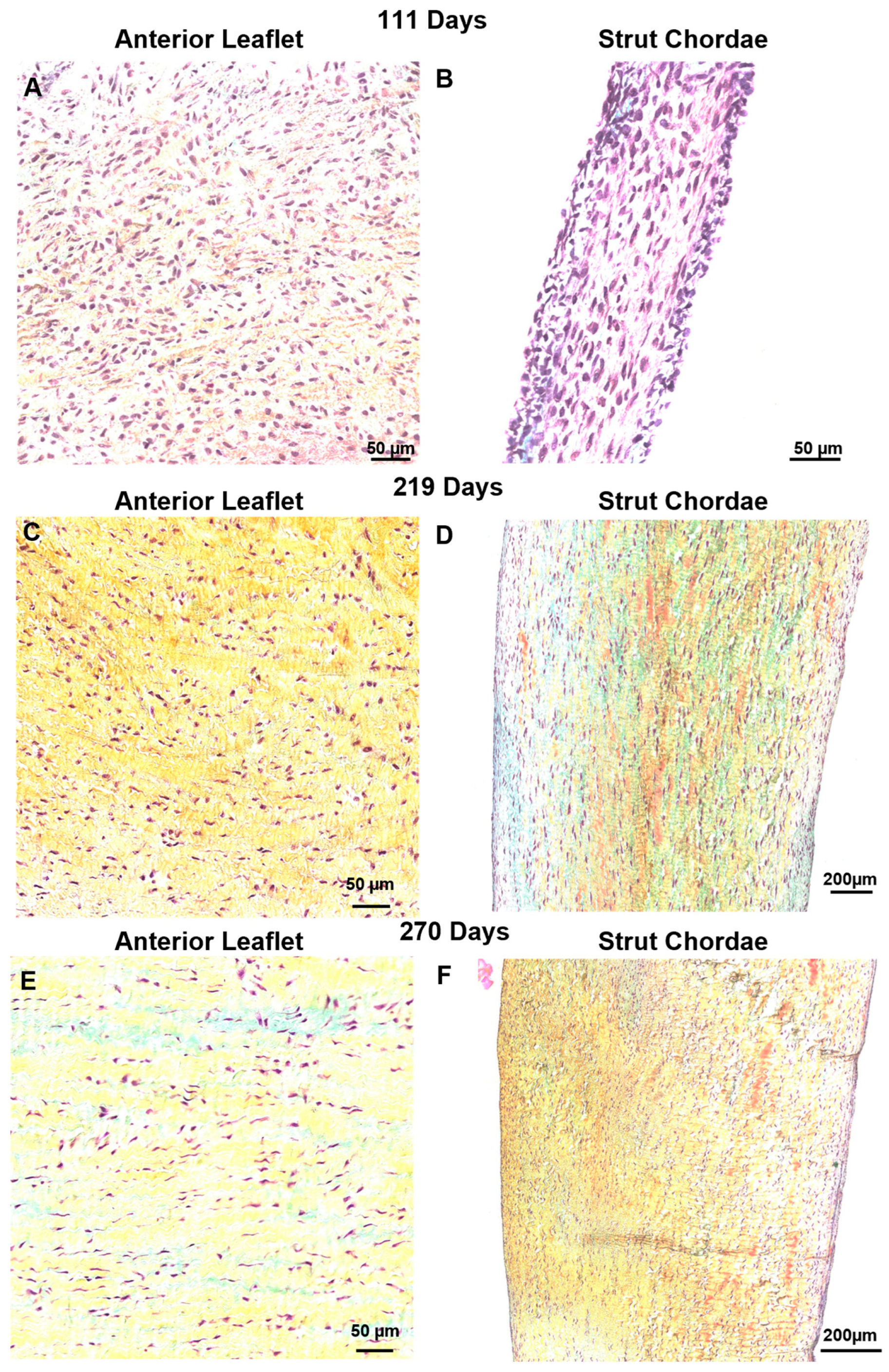
3.5. Contrary to Collagen Patterns, Elastic Fibers Appear in the Leaflet Ahead of the Chordae
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coffey, S.; Roberts-Thomson, R.; Brown, A.; Carapetis, J.; Chen, M.; Enriquez-Sarano, M.; Zühlke, L.; Prendergast, B.D. Global Epidemiology of Valvular Heart Disease. Nat. Rev. Cardiol. 2021, 18, 853–864. [Google Scholar] [CrossRef] [PubMed]
- Delling, F.N.; Noseworthy, P.A.; Adams, D.H.; Basso, C.; Borger, M.; Bouatia-Naji, N.; Elmariah, S.; Evans, F.; Gerstenfeld, E.; Hung, J.; et al. Research Opportunities in the Treatment of Mitral Valve Prolapse. J. Am. Coll. Cardiol. 2022, 80, 2331–2347. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.S.; Moore, R.; Fulmer, D.B.; Guo, L.; Gensemer, C.; Stairley, R.; Glover, J.; Beck, T.C.; Morningstar, J.E.; Biggs, R.; et al. DCHS1, Lix1L, and the Septin Cytoskeleton: Molecular and Developmental Etiology of Mitral Valve Prolapse. J. Cardiovasc. Dev. Dis. 2022, 9, 62. [Google Scholar] [CrossRef] [PubMed]
- Durst, R.; Sauls, K.; Peal, D.S.; de Vlaming, A.; Toomer, K.; Leyne, M.; Salani, M.; Talkowski, M.E.; Brand, H.; Perrocheau, M.; et al. Mutations in DCHS1 Cause Mitral Valve Prolapse. Nature 2015, 525, 109–113. [Google Scholar] [CrossRef] [PubMed]
- Sauls, K.; de Vlaming, A.; Harris, B.S.; Williams, K.; Wessels, A.; Levine, R.A.; Slaugenhaupt, S.A.; Goodwin, R.L.; Pavone, L.M.; Merot, J.; et al. Developmental Basis for Filamin-A-Associated Myxomatous Mitral Valve Disease. Cardiovasc. Res. 2012, 96, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Delwarde, C.; Capoulade, R.; Mérot, J.; Le Scouarnec, S.; Bouatia-Naji, N.; Yu, M.; Huttin, O.; Selton-Suty, C.; Sellal, J.-M.; Piriou, N.; et al. Genetics and Pathophysiology of Mitral Valve Prolapse. Front. Cardiovasc. Med. 2023, 10, 1077788. [Google Scholar] [CrossRef]
- Clemenceau, A.; Bérubé, J.; Bélanger, P.; Gaudreault, N.; Lamontagne, M.; Toubal, O.; Clavel, M.; Capoulade, R.; Mathieu, P.; Pibarot, P.; et al. Deleterious Variants in DCHS 1 Are Prevalent in Sporadic Cases of Mitral Valve Prolapse. Mol. Genet. Genom. Med. 2018, 6, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Monaghan, M.G.; Linneweh, M.; Liebscher, S.; Van Handel, B.; Layland, S.L.; Schenke-Layland, K. Endocardial-to-Mesenchymal Transformation and Mesenchymal Cell Colonization at the Onset of Human Cardiac Valve Development. Development 2016, 143, 473–482. [Google Scholar] [CrossRef]
- Camenisch, T.D.; Schroeder, J.A.; Bradley, J.; Klewer, S.E.; McDonald, J.A. Heart-Valve Mesenchyme Formation Is Dependent on Hyaluronan-Augmented Activation of ErbB2–ErbB3 Receptors. Nat. Med. 2002, 8, 850–855. [Google Scholar] [CrossRef]
- Enciso, J.M.; Gratzinger, D.; Camenisch, T.D.; Canosa, S.; Pinter, E.; Madri, J.A. Elevated Glucose Inhibits VEGF-A–Mediated Endocardial Cushion Formation. J. Cell Biol. 2003, 160, 605–615. [Google Scholar] [CrossRef]
- Kim, K.; Lee, D. ERBB3-Dependent AKT and ERK Pathways Are Essential for Atrioventricular Cushion Development in Mouse Embryos. PLoS ONE 2021, 16, e0259426. [Google Scholar] [CrossRef] [PubMed]
- Butcher, J.T.; McQuinn, T.C.; Sedmera, D.; Turner, D.; Markwald, R.R. Transitions in Early Embryonic Atrioventricular Valvular Function Correspond With Changes in Cushion Biomechanics That Are Predictable by Tissue Composition. Circ. Res. 2007, 100, 1503–1511. [Google Scholar] [CrossRef] [PubMed]
- Buskohl, P.R.; Gould, R.A.; Butcher, J.T. Quantification of Embryonic Atrioventricular Valve Biomechanics during Morphogenesis. J. Biomech. 2012, 45, 895–902. [Google Scholar] [CrossRef] [PubMed]
- Heckel, E.; Boselli, F.; Roth, S.; Krudewig, A.; Belting, H.-G.; Charvin, G.; Vermot, J. Oscillatory Flow Modulates Mechanosensitive Klf2a Expression through Trpv4 and Trpp2 during Heart Valve Development. Curr. Biol. 2015, 25, 1354–1361. [Google Scholar] [CrossRef] [PubMed]
- Donat, S.; Lourenço, M.; Paolini, A.; Otten, C.; Renz, M.; Abdelilah-Seyfried, S. Heg1 and Ccm1/2 Proteins Control Endocardial Mechanosensitivity during Zebrafish Valvulogenesis. eLife 2018, 7, e28939. [Google Scholar] [CrossRef] [PubMed]
- Gunawan, F.; Gentile, A.; Gauvrit, S.; Stainier, D.Y.R.; Bensimon-Brito, A. Nfatc1 Promotes Interstitial Cell Formation During Cardiac Valve Development in Zebrafish. Circ. Res. 2020, 126, 968–984. [Google Scholar] [CrossRef] [PubMed]
- Gittenberger-de Groot, A.C.; Vrancken Peeters, M.-P.F.M.; Mentink, M.M.T.; Gourdie, R.G.; Poelmann, R.E. Epicardium-Derived Cells Contribute a Novel Population to the Myocardial Wall and the Atrioventricular Cushions. Circ. Res. 1998, 82, 1043–1052. [Google Scholar] [CrossRef]
- Pérez-Pomares, J.M.; Phelps, A.; Sedmerova, M.; Carmona, R.; González-Iriarte, M.; Muñoz-Chápuli, R.; Wessels, A. Experimental Studies on the Spatiotemporal Expression of WT1 and RALDH2 in the Embryonic Avian Heart: A Model for the Regulation of Myocardial and Valvuloseptal Development by Epicardially Derived Cells (EPDCs). Dev. Biol. 2002, 247, 307–326. [Google Scholar] [CrossRef]
- Wessels, A.; Van Den Hoff, M.J.B.; Adamo, R.F.; Phelps, A.L.; Lockhart, M.M.; Sauls, K.; Briggs, L.E.; Norris, R.A.; Van Wijk, B.; Perez-Pomares, J.M.; et al. Epicardially Derived Fibroblasts Preferentially Contribute to the Parietal Leaflets of the Atrioventricular Valves in the Murine Heart. Dev. Biol. 2012, 366, 111–124. [Google Scholar] [CrossRef]
- Bassen, D.; Wang, M.; Pham, D.; Sun, S.; Rao, R.; Singh, R.; Butcher, J. Hydrostatic Mechanical Stress Regulates Growth and Maturation of the Atrioventricular Valve. Development 2021, 148, dev196519. [Google Scholar] [CrossRef]
- Wang, M.; Lin, B.Y.; Sun, S.; Dai, C.; Long, F.; Butcher, J.T. Shear and Hydrostatic Stress Regulate Fetal Heart Valve Remodeling through YAP-Mediated Mechanotransduction. eLife 2023, 12, e83209. [Google Scholar] [CrossRef] [PubMed]
- Gould, R.A.; Yalcin, H.C.; MacKay, J.L.; Sauls, K.; Norris, R.; Kumar, S.; Butcher, J.T. Cyclic Mechanical Loading Is Essential for Rac1-Mediated Elongation and Remodeling of the Embryonic Mitral Valve. Curr. Biol. 2016, 26, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Vermot, J.; Forouhar, A.S.; Liebling, M.; Wu, D.; Plummer, D.; Gharib, M.; Fraser, S.E. Reversing Blood Flows Act through Klf2a to Ensure Normal Valvulogenesis in the Developing Heart. PLoS Biol. 2009, 7, e1000246. [Google Scholar] [CrossRef]
- Goddard, L.M.; Duchemin, A.-L.; Ramalingan, H.; Wu, B.; Chen, M.; Bamezai, S.; Yang, J.; Li, L.; Morley, M.P.; Wang, T.; et al. Hemodynamic Forces Sculpt Developing Heart Valves through a KLF2-WNT9B Paracrine Signaling Axis. Dev. Cell 2017, 43, 274–289.e5. [Google Scholar] [CrossRef]
- Vignes, H.; Vagena-Pantoula, C.; Prakash, M.; Fukui, H.; Norden, C.; Mochizuki, N.; Jug, F.; Vermot, J. Extracellular Mechanical Forces Drive Endocardial Cell Volume Decrease during Zebrafish Cardiac Valve Morphogenesis. Dev. Cell 2022, 57, 598–609.e5. [Google Scholar] [CrossRef] [PubMed]
- Oosthoek, P.W.; Wenink, A.C.G.; Vrolijk, B.C.M.; Wisse, L.J.; DeRuiter, M.C.; Poelmann, R.E.; Gittenberger-de Groot, A.C. Development of the Atrioventricular Valve Tension Apparatus in the Human Heart. Anat. Embryol. 1998, 198, 317–329. [Google Scholar] [CrossRef]
- Kruithof, B.P.T.; Krawitz, S.A.; Gaussin, V. Atrioventricular Valve Development during Late Embryonic and Postnatal Stages Involves Condensation and Extracellular Matrix Remodeling. Dev. Biol. 2007, 302, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Peacock, J.D.; Lu, Y.; Koch, M.; Kadler, K.E.; Lincoln, J. Temporal and Spatial Expression of Collagens during Murine Atrioventricular Heart Valve Development and Maintenance. Dev. Dyn. 2008, 237, 3051–3058. [Google Scholar] [CrossRef] [PubMed]
- de Lange, F.J.; Moorman, A.F.M.; Anderson, R.H.; Männer, J.; Soufan, A.T.; de Vries, C.G.; Schneider, M.D.; Webb, S.; van den Hoff, M.J.B.; Christoffels, V.M. Lineage and Morphogenetic Analysis of the Cardiac Valves. Circ. Res. 2004, 95, 645–654. [Google Scholar] [CrossRef]
- Lincoln, J.; Alfieri, C.M.; Yutzey, K.E. Development of Heart Valve Leaflets and Supporting Apparatus in Chicken and Mouse Embryos. Dev. Dyn. 2004, 230, 239–250. [Google Scholar] [CrossRef]
- Norris, R.A.; Kern, C.B.; Wessels, A.; Moralez, E.I.; Markwald, R.R.; Mjaatvedt, C.H. Identification and Detection of the Periostin Gene in Cardiac Development. Anat. Rec. 2004, 281A, 1227–1233. [Google Scholar] [CrossRef] [PubMed]
- Morse, D.E.; Hamlett, W.C.; Noble, C.W. Morphogenesis of Chordae Tendineae. I: Scanning Electron Microscopy. Anat. Rec. 1984, 210, 629–638. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.; Junor, L.; Price, R.L.; Norris, R.A.; Potts, J.D.; Goodwin, R.L. Expression and Deposition of Fibrous Extracellular Matrix Proteins in Cardiac Valves during Chick Development. Microsc. Microanal. 2011, 17, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Stephens, E.H.; Grande-Allen, K.J. Age-Related Changes in Collagen Synthesis and Turnover in Porcine Heart Valves. J. Heart Valve Dis. 2007, 16, 11. [Google Scholar]
- Kasyanov, V.; Moreno-Rodriguez, R.A.; Kalejs, M.; Ozolanta, I.; Stradins, P.; Wen, X.; Yao, H.; Mironov, V. Age-Related Analysis of Structural, Biochemical and Mechanical Properties of the Porcine Mitral Heart Valve Leaflets. Connect. Tissue Res. 2013, 54, 394–402. [Google Scholar] [CrossRef] [PubMed]
- Stephens, E.H.; Post, A.D.; Laucirica, D.R.; Grande-Allen, K.J. Perinatal Changes in Mitral and Aortic Valve Structure and Composition. Pediatr. Dev. Pathol. 2010, 13, 447–458. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Wirrig, E.E.; Hinton, R.B.; Merrill, W.H.; Spicer, D.B.; Yutzey, K.E. Twist1 Promotes Heart Valve Cell Proliferation and Extracellular Matrix Gene Expression during Development in Vivo and Is Expressed in Human Diseased Aortic Valves. Dev. Biol. 2010, 347, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Norris, R.A.; Moreno-Rodriguez, R.A.; Sugi, Y.; Hoffman, S.; Amos, J.; Hart, M.M.; Potts, J.D.; Goodwin, R.L.; Markwald, R.R. Periostin Regulates Atrioventricular Valve Maturation. Dev. Biol. 2008, 316, 200–213. [Google Scholar] [CrossRef] [PubMed]
- Aikawa, E.; Whittaker, P.; Farber, M.; Mendelson, K.; Padera, R.F.; Aikawa, M.; Schoen, F.J. Human Semilunar Cardiac Valve Remodeling by Activated Cells From Fetus to Adult: Implications for Postnatal Adaptation, Pathology, and Tissue Engineering. Circulation 2006, 113, 1344–1352. [Google Scholar] [CrossRef]
- Votteler, M.; Berrio, D.A.C.; Horke, A.; Sabatier, L.; Reinhardt, D.P.; Nsair, A.; Aikawa, E.; Schenke-Layland, K. Elastogenesis at the Onset of Human Cardiac Valve Development. Development 2013, 140, 2345–2353. [Google Scholar] [CrossRef]
- Gaussin, V.; Morley, G.E.; Cox, L.; Zwijsen, A.; Vance, K.M.; Emile, L.; Tian, Y.; Liu, J.; Hong, C.; Myers, D.; et al. Alk3/Bmpr1a Receptor Is Required for Development of the Atrioventricular Canal Into Valves and Annulus Fibrosus. Circ. Res. 2005, 97, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, M.G.; Vesely, I. Structural Changes of Rat Mitral Valve Chordae Tendineae During Postnatal Development. J. Heart Valve Dis. 2012, 21, 7. [Google Scholar]
- Aldous, I.G.; Lee, J.M.; Wells, S.M. Differential Changes in the Molecular Stability of Collagen from the Pulmonary and Aortic Valves During the Fetal-to-Neonatal Transition. Ann. Biomed. Eng. 2010, 38, 3000–3009. [Google Scholar] [CrossRef] [PubMed]
- Merryman, W.D.; Youn, I.; Lukoff, H.D.; Krueger, P.M.; Guilak, F.; Hopkins, R.A.; Sacks, M.S. Correlation between Heart Valve Interstitial Cell Stiffness and Transvalvular Pressure: Implications for Collagen Biosynthesis. Am. J. Physiol.-Heart C 2006, 290, H224–H231. [Google Scholar] [CrossRef] [PubMed]
- Hinton, R.B.; Alfieri, C.M.; Witt, S.A.; Glascock, B.J.; Khoury, P.R.; Benson, D.W.; Yutzey, K.E. Mouse Heart Valve Structure and Function: Echocardiographic and Morphometric Analyses from the Fetus through the Aged Adult. Am. J. Physiol.-Heart C 2008, 294, H2480–H2488. [Google Scholar] [CrossRef]
- Filipoiu, F.M. Chapter 2: The Specific Development Period. In Atlas of Heart Anatomy and Development, 1st ed.; Springer: London, UK, 2014; ISBN 978-1-4471-5381-8. [Google Scholar]
- Aldous, I.G.; Veres, S.P.; Jahangir, A.; Lee, J.M. Differences in Collagen Cross-Linking between the Four Valves of the Bovine Heart: A Possible Role in Adaptation to Mechanical Fatigue. Am. J. Physiol.-Heart C 2009, 296, H1898–H1906. [Google Scholar] [CrossRef] [PubMed]
- Evans, H.E.; Sack, W.O. Prenatal Development of Domestic and Laboratory Mammals: Growth Curves, External Features and Selected References. Anatom. Histol. Embryol. 1973, 2, 11–45. [Google Scholar] [CrossRef] [PubMed]
- Fratzl, P.; Misof, K.; Zizak, I.; Rapp, G.; Amenitsch, H.; Bernstorff, S. Fibrillar Structure and Mechanical Properties of Collagen. J. Struct. Biol. 1998, 122, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Pierlot, C.M.; Moeller, A.D.; Lee, J.M.; Wells, S.M. Biaxial Creep Resistance and Structural Remodeling of the Aortic and Mitral Valves in Pregnancy. Ann. Biomed. Eng. 2015, 43, 1772–1785. [Google Scholar] [CrossRef]
- Pierlot, C.M.; Lee, J.M.; Amini, R.; Sacks, M.S.; Wells, S.M. Pregnancy-Induced Remodeling of Collagen Architecture and Content in the Mitral Valve. Ann. Biomed. Eng. 2014, 42, 2058–2071. [Google Scholar] [CrossRef]
- Pierlot, C.M.; Moeller, A.D.; Lee, J.M.; Wells, S.M. Pregnancy-Induced Remodeling of Heart Valves. Am. J. Physiol.-Heart C 2015, 309, H1565–H1578. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; Hamilton, ON, Canada, 2019. Available online: https://r-project.org/ (accessed on 3 March 2024).
- Jarek, S. Mvnormtest: Normality Test for Multivariate Variables; Kraków, Poland, 2012. Available online: https://cran.r-project.org/web/packages/car/index.html (accessed on 3 March 2024).
- Signorell, A. DescTools: Tools for Descriptive Statistics; Dübendorf, Switzerland, 2023. Available online: https://cran.r-project.org/web/packages/mvnormtest/index.html (accessed on 3 March 2024).
- Wickham, H.; Hester, J.; Chang, W.; Bryan, J. Devtools: Tools to Make Developing R Packages Easier; Colorado, USA, 2022. Available online: https://cran.r-project.org/web/packages/DescTools/index.html (accessed on 3 March 2024).
- Scott, B.P. Physiological Remodelling of Mitral Valve Chordae Tendineae in the Maternal Bovine Heart. Master’s Thesis, Dalhousie University, Halifax, NS, USA, 2016. [Google Scholar]
- Wells, S.M.; Pierlot, C.M.; Moeller, A.D. Physiological Remodeling of the Mitral Valve during Pregnancy. Am. J. Physiol.-Heart C 2012, 303, H878–H892. [Google Scholar] [CrossRef]
- Kodigepalli, K.M.; Thatcher, K.; West, T.; Howsmon, D.P.; Schoen, F.J.; Sacks, M.S.; Breuer, C.K.; Lincoln, J. Biology and Biomechanics of the Heart Valve Extracellular Matrix. J. Cardiovasc. Dev. Dis. 2020, 7, 57. [Google Scholar] [CrossRef]
- Millington-Sanders, C.; Meir, A.; Lawrence, L.; Stolinski, C. Structure of Chordae Tendineae in the Left Ventricle of the Human Heart. J. Anat. 1998, 192, 573–581. [Google Scholar] [CrossRef] [PubMed]
- Stephens, E.H.; de Jonge, N.; McNeill, M.P.; Durst, C.A.; Grande-Allen, K.J. Age-Related Changes in Material Behavior of Porcine Mitral and Aortic Valves and Correlation to Matrix Composition. Tissue Eng. Pt. A 2010, 16, 867–878. [Google Scholar] [CrossRef] [PubMed]
- Norris, R.A.; Potts, J.D.; Yost, M.J.; Junor, L.; Brooks, T.; Tan, H.; Hoffman, S.; Hart, M.M.; Kern, M.J.; Damon, B.; et al. Periostin Promotes a Fibroblastic Lineage Pathway in Atrioventricular Valve Progenitor Cells. Dev. Dyn. 2009, 238, 1052–1063. [Google Scholar] [CrossRef]
- Chakraborty, S.; Cheek, J.; Sakthivel, B.; Aronow, B.J.; Yutzey, K.E. Shared Gene Expression Profiles in Developing Heart Valves and Osteoblast Progenitor Cells. Physiol. Genom. 2008, 35, 75–85. [Google Scholar] [CrossRef]
- Fratzl, P. Chapter 2: Collagen Diversity, Synthesis and Assembly. In Collagen: Structure and Mechanics; Springer: New York, NY, USA, 2008; ISBN 978-0-387-73905-2. [Google Scholar]
- Kalson, N.S.; Lu, Y.; Taylor, S.H.; Starborg, T.; Holmes, D.F.; Kadler, K.E. A Structure-Based Extracellular Matrix Expansion Mechanism of Fibrous Tissue Growth. eLife 2015, 4, e05958. [Google Scholar] [CrossRef]
- Herchenhan, A.; Kalson, N.S.; Holmes, D.F.; Hill, P.; Kadler, K.E.; Margetts, L. Tenocyte Contraction Induces Crimp Formation in Tendon-like Tissue. Biomech. Model. Mechanobiol. 2012, 11, 449–459. [Google Scholar] [CrossRef]
- Legerlotz, K.; Dorn, J.; Richter, J.; Rausch, M.; Leupin, O. Age-Dependent Regulation of Tendon Crimp Structure, Cell Length and Gap Width with Strain. Acta Biomater. 2014, 10, 4447–4455. [Google Scholar] [CrossRef]
- Oryan, A.; Shoushtari, A.H. Histology and Ultrastructure of the Developing Superficial Digital Flexor Tendon in Rabbits. Anat. Histol. Embryol. 2008, 37, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Ritz-Timme, S.; Laumeier, I.; Collins, M.J. Aspartic Acid Racemization: Evidence for Marked Longevity of Elastin in Human Skin. Br. J. Dermatol. 2003, 149, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.J.; Robson, P.; Hew, Y.; Keeley, F.W. Decreased Elastin Synthesis in Normal Development and in Long-Term Aortic Organ and Cell Cultures Is Related to Rapid and Selective Destabilization of mRNA for Elastin. Circ. Res. 1995, 77, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Lincoln, J.; Alfieri, C.M.; Yutzey, K.E. BMP and FGF Regulatory Pathways Control Cell Lineage Diversification of Heart Valve Precursor Cells. Dev. Biol. 2006, 292, 290–302. [Google Scholar] [CrossRef] [PubMed]
- Hinton, R.B.; Lincoln, J.; Deutsch, G.H.; Osinska, H.; Manning, P.B.; Benson, D.W.; Yutzey, K.E. Extracellular Matrix Remodeling and Organization in Developing and Diseased Aortic Valves. Circ. Res. 2006, 98, 1431–1438. [Google Scholar] [CrossRef] [PubMed]
- Brettell, L.M.; McGowan, S.E. Basic Fibroblast Growth Factor Decreases Elastin Production by Neonatal Rat Lung Fibroblasts. Am. J. Respir. Cell Mol. Biol. 1994, 10, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Rich, C.B.; Fontanilla, M.R.; Nugent, M.; Foster, J.A. Basic Fibroblast Growth Factor Decreases Elastin Gene Transcription through an AP1/cAMP-Response Element Hybrid Site in the Distal Promoter. J. Biol. Chem. 1999, 274, 33433–33439. [Google Scholar] [CrossRef] [PubMed]
- Fowden, A.L.; Li, J.; Forhead, A.J. Glucocorticoids and the Preparation for Life after Birth: Are There Long-Term Consequences of the Life Insurance? Proc. Nutr. Soc. 1998, 57, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Rich, C.B.; Goud, H.D.; Bashir, M.; Rosenbloom, J.; Foster, J.A. Developmental Regulation of Aortic Elastin Gene Expression Involves Disruption of an IGF-I Sensitive Repressor Complex. Biochem. Biophys. Res. Commun. 1993, 196, 1316–1322. [Google Scholar] [CrossRef]
- Pierce, R.A.; Mariencheck, W.I.; Sandefur, S.; Crouch, E.C.; Parks, W.C. Glucocorticoids Upregulate Tropoelastin Expression during Late Stages of Fetal Lung Development. Am. J. Physiol.-Lung C 1995, 268, L491–L500. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martin, M.; Chen, C.-Y.; McCowan, T.; Wells, S. Differential Development of the Chordae Tendineae and Anterior Leaflet of the Bovine Mitral Valve. J. Cardiovasc. Dev. Dis. 2024, 11, 106. https://doi.org/10.3390/jcdd11040106
Martin M, Chen C-Y, McCowan T, Wells S. Differential Development of the Chordae Tendineae and Anterior Leaflet of the Bovine Mitral Valve. Journal of Cardiovascular Development and Disease. 2024; 11(4):106. https://doi.org/10.3390/jcdd11040106
Chicago/Turabian StyleMartin, Meghan, Chih-Ying Chen, Timothy McCowan, and Sarah Wells. 2024. "Differential Development of the Chordae Tendineae and Anterior Leaflet of the Bovine Mitral Valve" Journal of Cardiovascular Development and Disease 11, no. 4: 106. https://doi.org/10.3390/jcdd11040106
APA StyleMartin, M., Chen, C.-Y., McCowan, T., & Wells, S. (2024). Differential Development of the Chordae Tendineae and Anterior Leaflet of the Bovine Mitral Valve. Journal of Cardiovascular Development and Disease, 11(4), 106. https://doi.org/10.3390/jcdd11040106