
Microascaceae from the Marine Environment, with Descriptions of Six New Species

 ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Fungal Isolation

2.2. Morphological Observation
2.3. DNA Extraction and Amplification
2.4. Phylogenetic Analyses
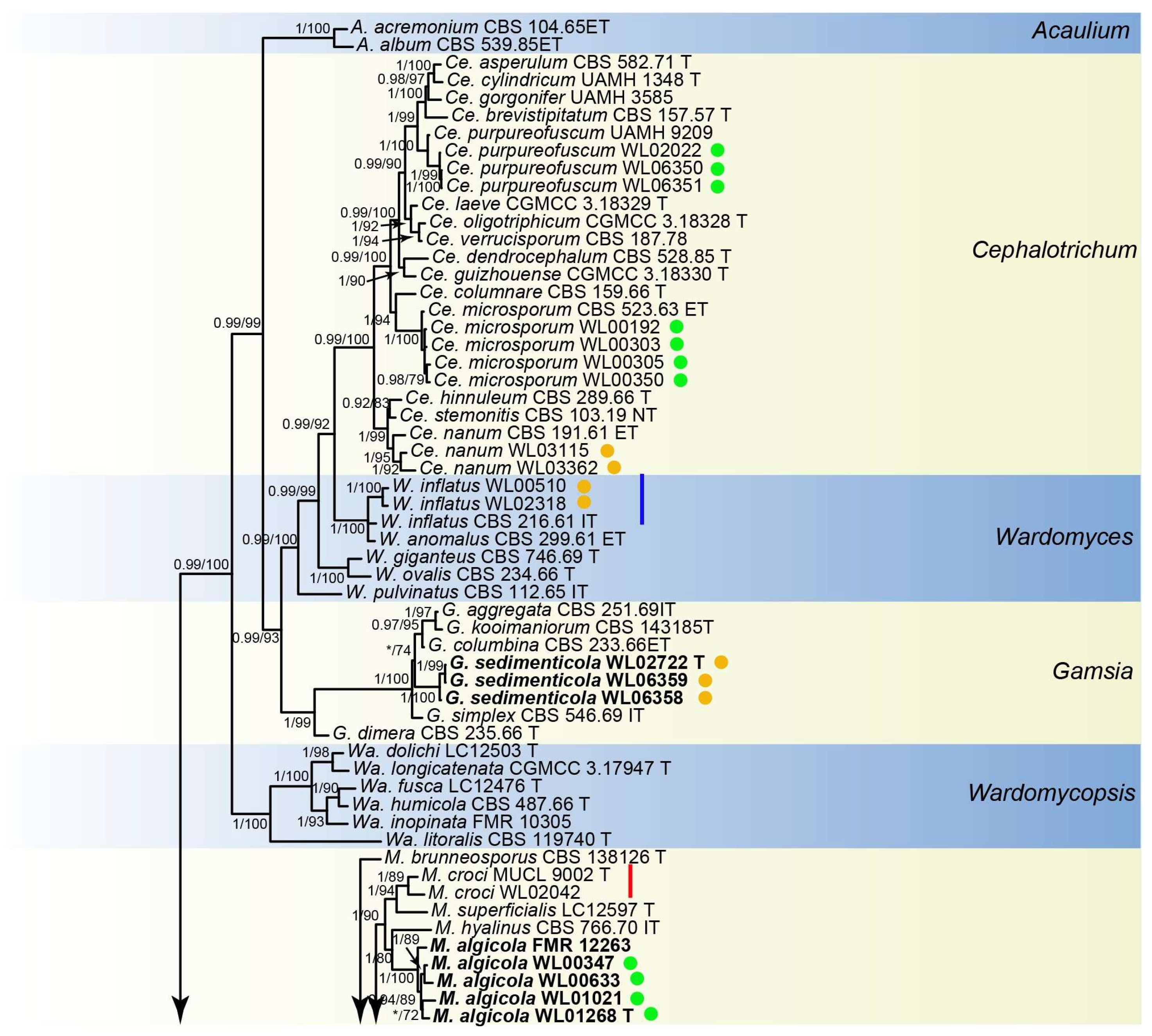
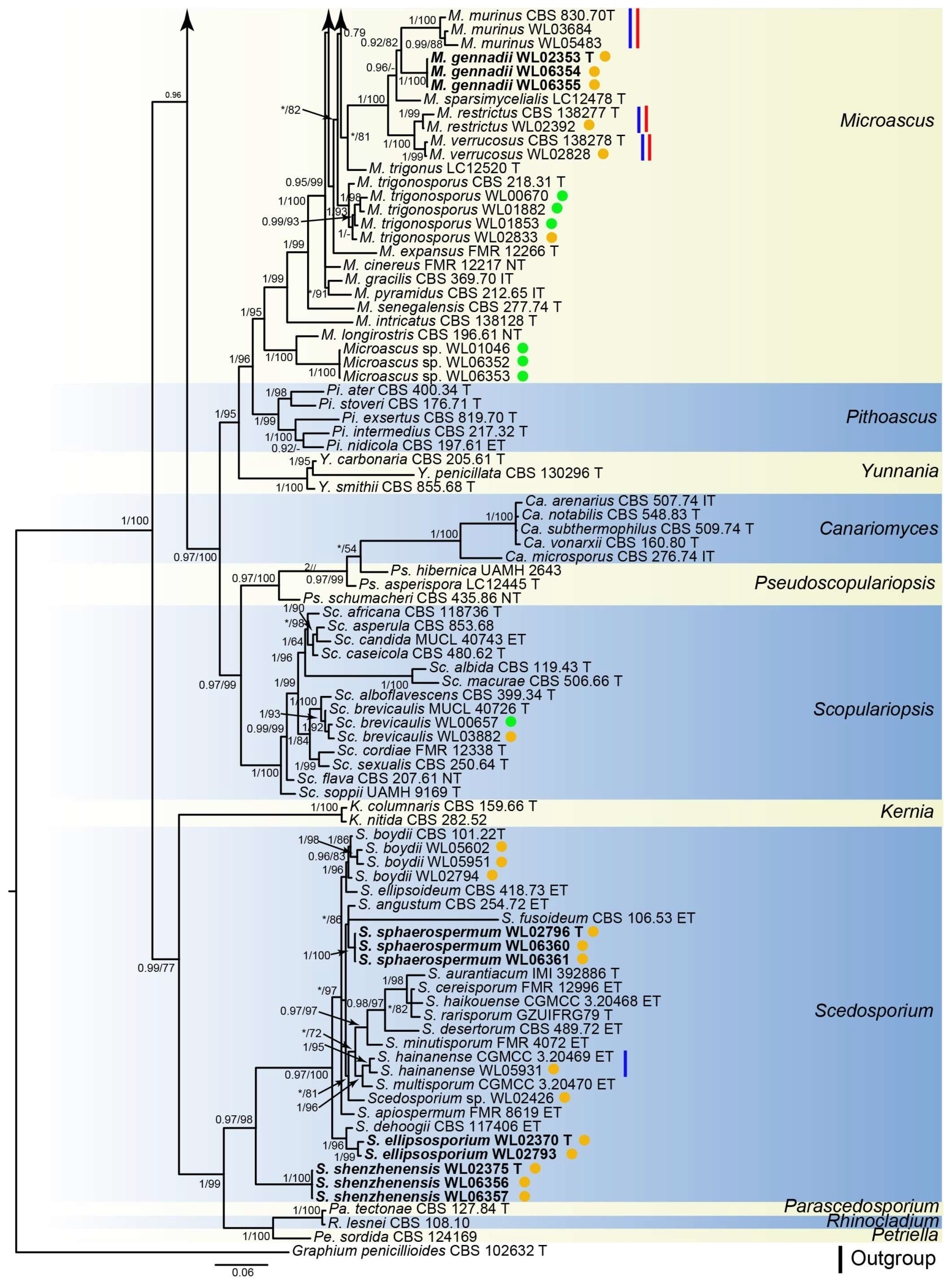
3. Results
3.1. Phylogenetic Analyses
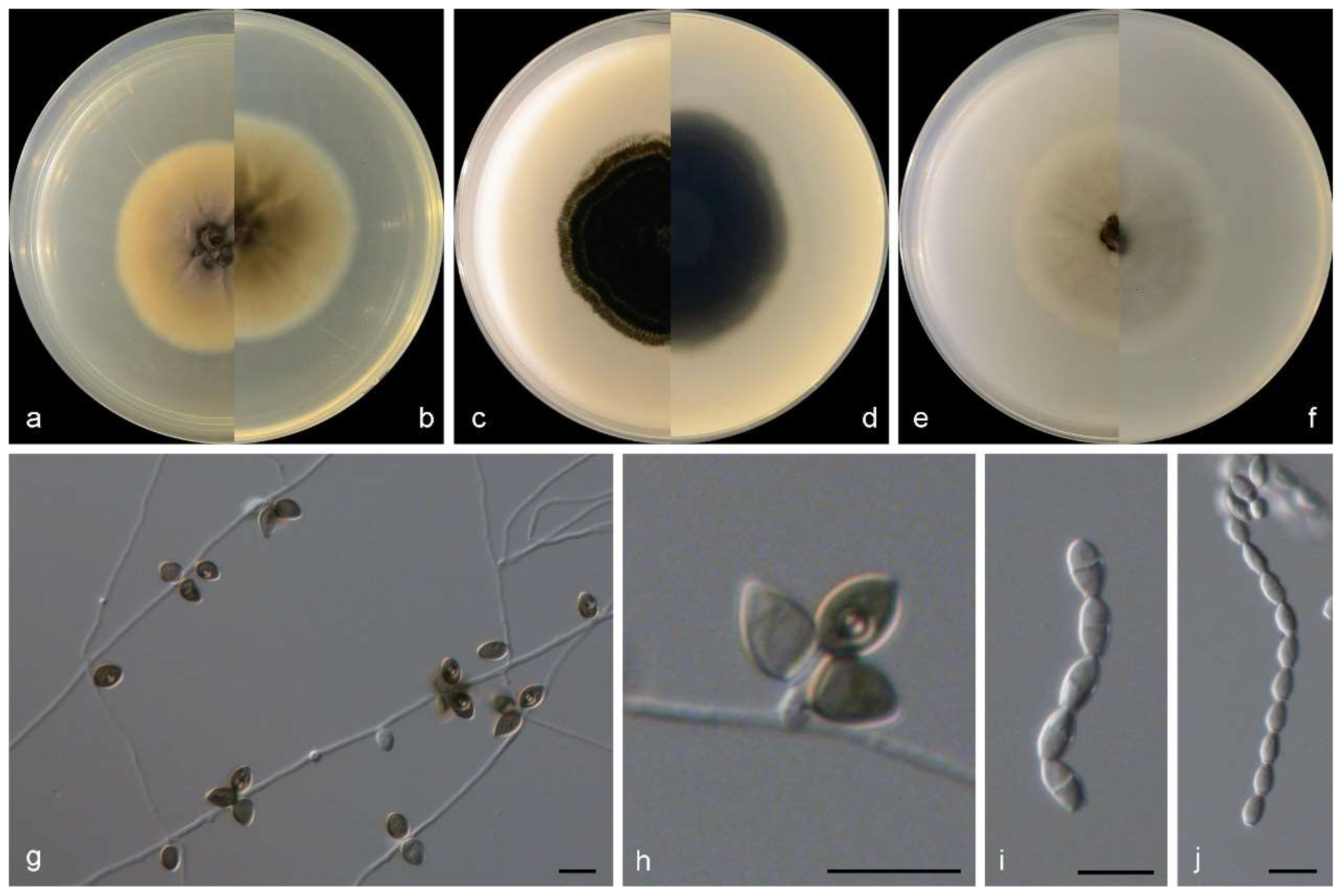
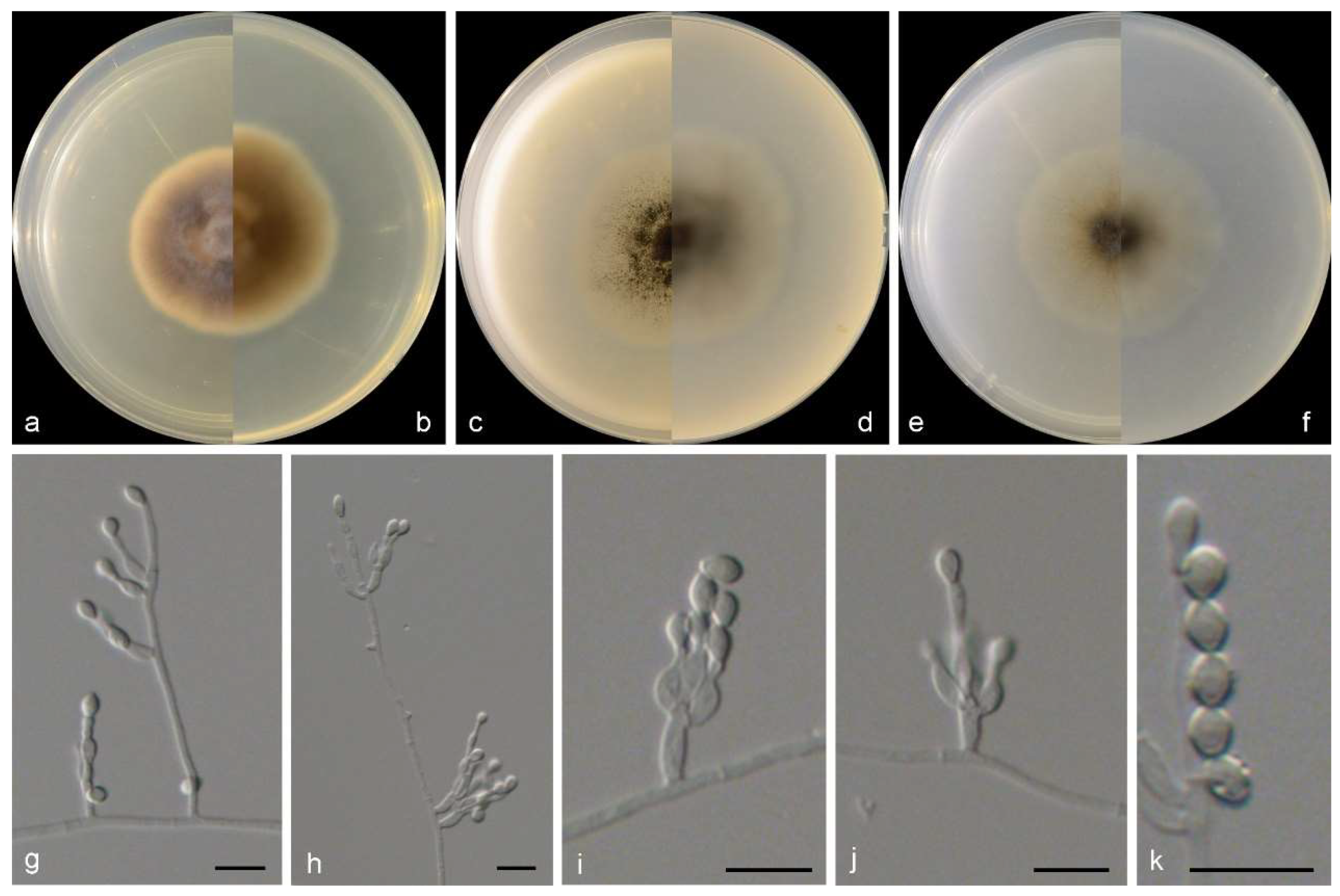
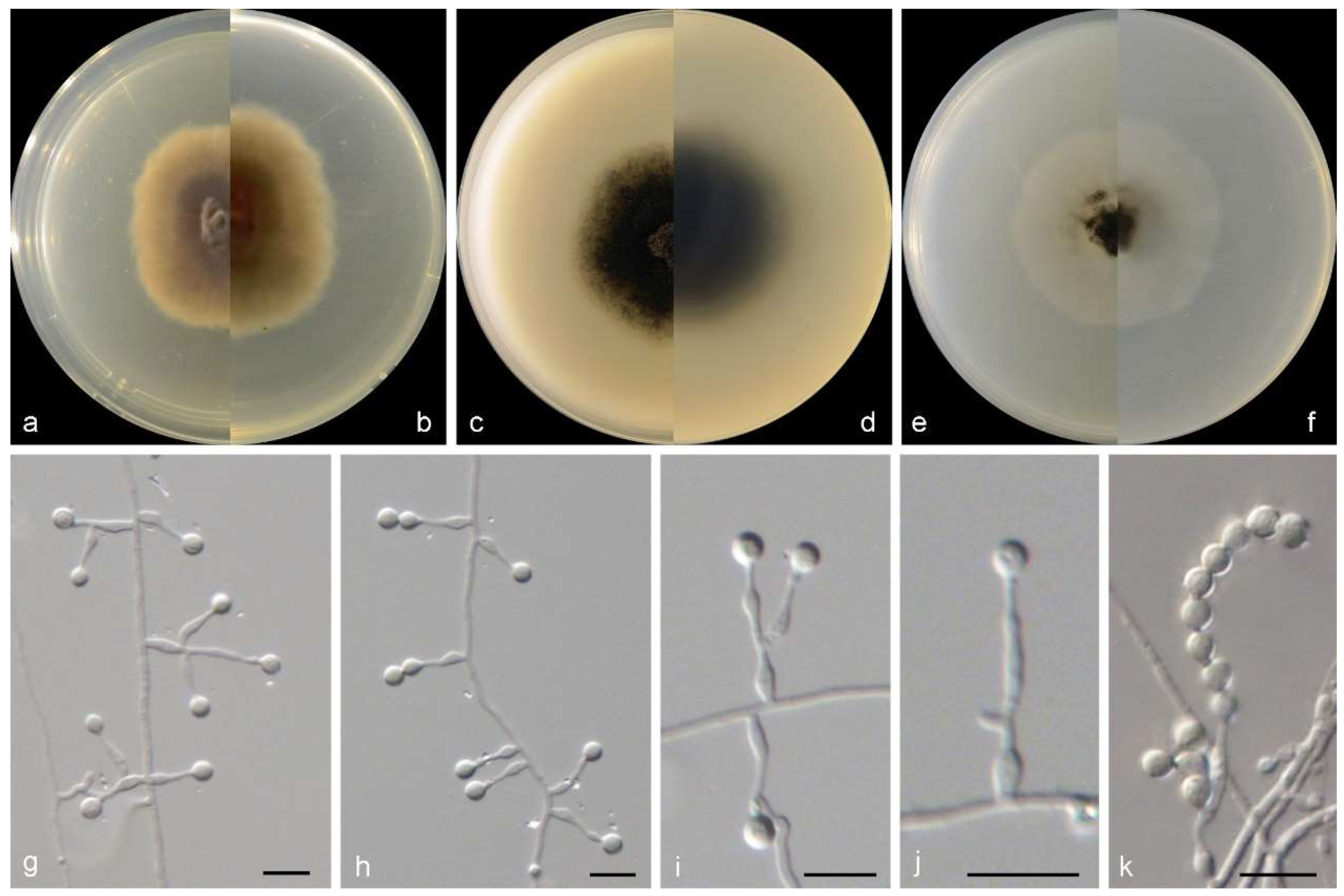
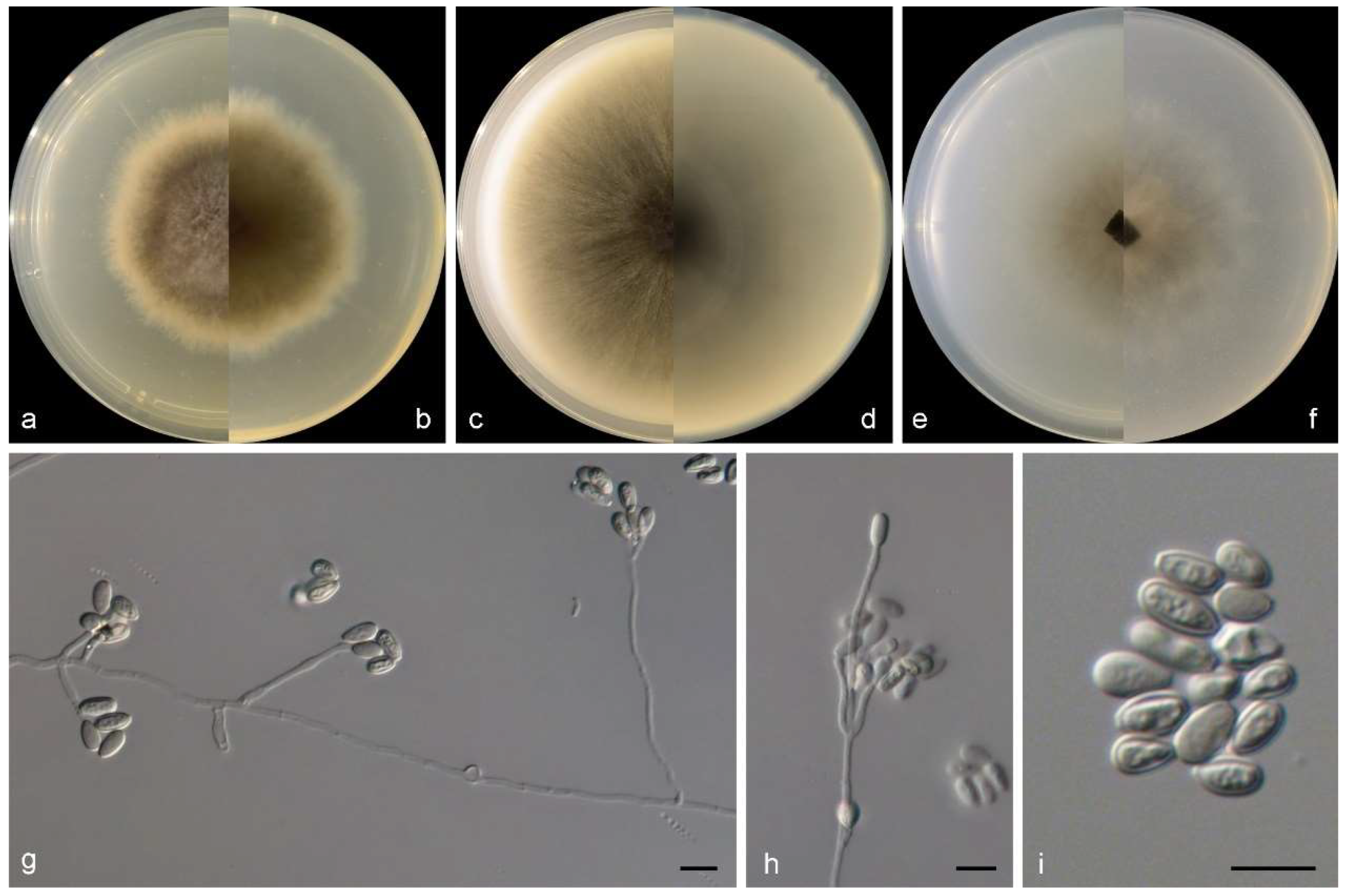
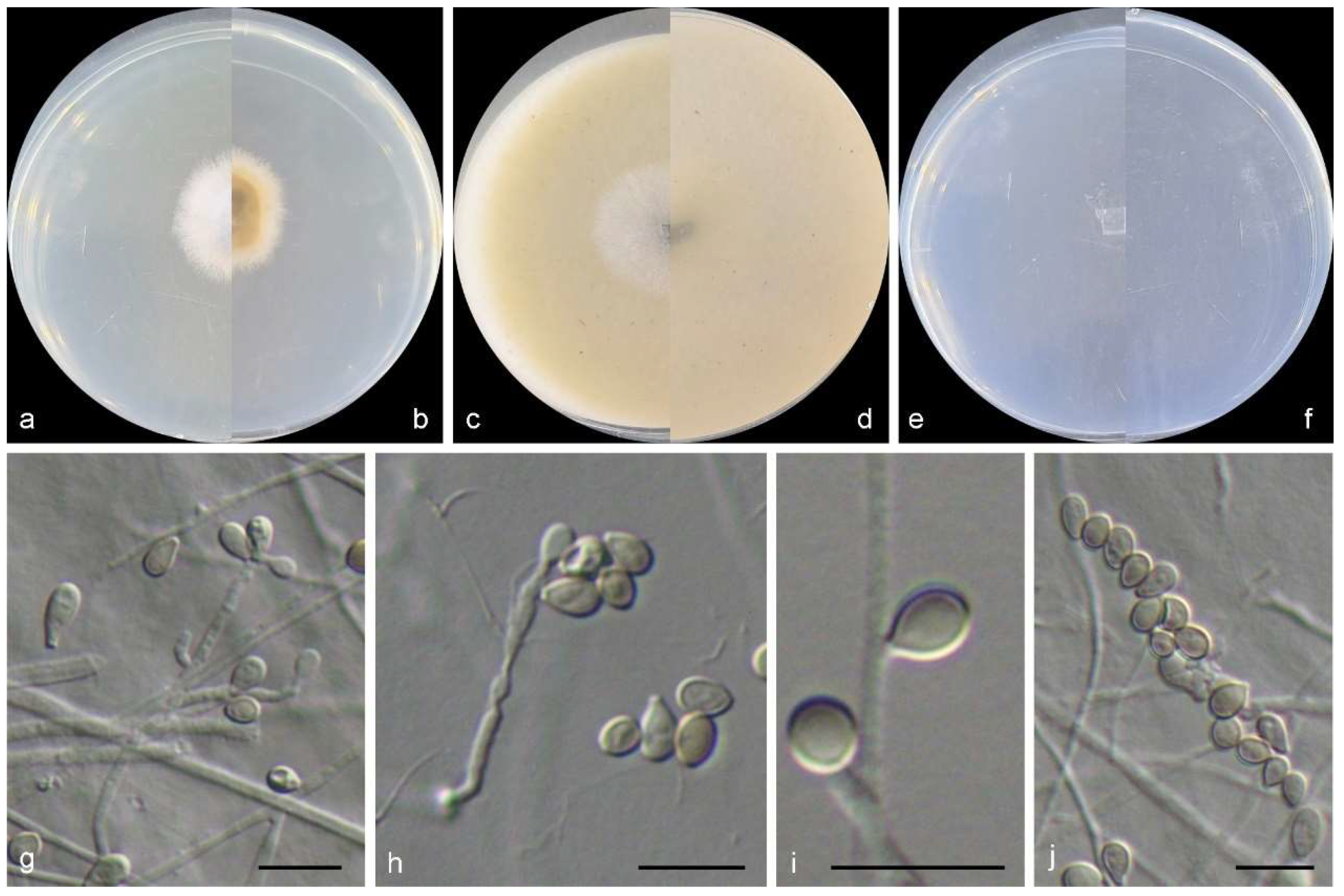
3.2. Species List and Taxonomy






4. Discussion

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Country/Region/Location | Known Habitats/Hosts | Recorded Database/Reference |
|---|---|---|---|
| Acaulium acremonium | na | na | MF |
| Cephalotrichum microsporum | China; Europe | unidentified brown algae, green algae, and red algae | WoRMS; this study |
| Ce. nanum | China; Europe | Intertidal sediment | WoRMS; this study |
| Ce. purpureofuscum | China | Marine algae Sargassum canfusum | This study |
| Ce. stemonitis | Europe | na | MF and WoRMS |
| Gamsia sedimenticola * | China | Intertidal sediment | This study |
| Microascus algicola * | China | Unidentified brown algae; marine algae Ahnfeltiopsis flabelliformis, Colpomenia sinuosa, Grateloupia filicina | This study |
| M. croci# | China; Hawaiian, Line, and Phoenix Islands | Marine algae Blidingia minima; coastal sands | [52]; This study |
| M. gennadii * | China | Intertidal sediment | This study |
| M. murinus +# | China | Intertidal sediment | This study |
| M. paisii | na | na | MF |
| M. restrictus +# | China | Intertidal sediment | This study |
| M. senegalensis | Senegal | Mangrove soil | [53] |
| M. trigonosporus | China, Europe; Hawaiian, Line, and Phoenix Islands | Marine algae Cladophora sp., Ulva linza, Gracilaria textorii; intertidal sediment; coastal sand | MF and WoRMS; [52] |
| M. verrucosus +# | China | Intertidal sediment | This study |
| Petriella sordida | na | na | MF |
| Scedosporium boydii | China | Intertidal sediment | MF; this study |
| S. dehoogii | China | Coastal sediment | [8] |
| S. ellipsosporium * | China | Intertidal sediment | This study |
| S. hainanense + | China | Intertidal sediment | This study |
| S. marinum | India | Decaying woody stem of Suaeda monoica | [54] |
| S. shenzhenensis * | China | Intertidal sediment | This study |
| S. sphaerospermum * | China | Intertidal sediment | This study |
| Scopulariopsis brevicaulis | China, Europe, Croatia; Hawaiian, Line, and Phoenix Islands | Marine sponge Tethya aurantium; marine algae Ulva pertusa; intertidal sediment; coastal sands | MF and WoRMS; [52]; This study |
| Sc. Brumptii | Hawaiian, Line, and Phoenix Islands | Coastal sands | MF; [52] |
| Sc. Candida | Egypt | Salt marshes | MF; [55] |
| Sc. Coprophila | Hawaiian, Line, and Phoenix Islands | Coastal seawater and sands | [52] |
| Sc. Fusca | Europe | na | WoRMS |
| Sc. Halophilica | na | na | MF and WoRMS |
| Sc. Hibernica | na | na | MF |
| Wardomyces anomalus | na | na | MF |
| W. inflatus+ | China | Intertidal sediment | This study |
| Yunnania carbonaria | Hawaiian | Coastal seawater | [52] |
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Luttrell, E.S. Taxonomy of the Pyrenomycetes. Univ. Mo. Stud. 1951, 24, 1–121. [Google Scholar]
- Malloch, D. New concepts in the Microascaceae illustrated by two new species. Mycologia 1970, 62, 727–740. [Google Scholar] [CrossRef]
- Zhang, Z.F.; Zhou, S.Y.; Eurwilaichitr, L.; Ingsriswang, S.; Raza, M.; Chen, Q.; Zhao, P.; Liu, F.; Cai, L. Culturable mycobiota from Karst caves in China II, with descriptions of 33 new species. Fungal Divers 2021, 106, 29–126. [Google Scholar] [CrossRef]
- Wijayawardene, N.N.; Hyde, K.D.; Dai, D.Q.; Sánchez-García, M.; Goto, B.; Saxena, R.; Erdoğdu, M.; Selçuk, F.; Rajeshkumar, K.; Aptroot, A.; et al. Outline of Fungi and fungus-like taxa—2021. Mycosphere 2022, 13, 53–453. [Google Scholar] [CrossRef]
- Sandoval-Denis, M.; Guarro, J.; Cano-Lira, J.F.; Sutton, D.; Wiederhold, N.; de Hoog, G.; Abbott, S.; Decock, C.; Sigler, L.; Gené, J. Phylogeny and taxonomic revision of Microascaceae with emphasis on synnematous fungi. Stud. Mycol 2016, 83, 193–233. [Google Scholar] [CrossRef]
- Tubaki, K. An undescribed halophilic species of Scopulariopsis. Trans Mycol Soc Jpn. 1973, 14, 367–369. [Google Scholar]
- Clipson, N.; Landy, E.; Otte, M. European register of marine species: A check-list of the marine species in Europe and a bibliography of guides to their identification. Collect. Patrim. Nat. 2001, 50, 15–19. [Google Scholar]
- Feng, C.H.; Li, C.L.; Peng, Y.T.; Sun, K.; Li, W. A preliminary report of hyphomycetes from sediment of southern Yellow Sea. Mycosystema 2013, 32, 35–41. [Google Scholar]
- Jones, E.B.G.; Suetrong, S.; Sakayaroj, J.; Bahkali, A.H.; Abdel-Wahab, M.A.; Boekhout, T.; Pang, K.-L. Classification of marine Ascomycota, Basidiomycota, Blastocladiomycota and Chytridiomycota. Fungal Divers 2015, 73, 1–72. [Google Scholar] [CrossRef]
- Jones, E.B.G.; Pang, K.L.; Abdel-Wahab, M.A.; Scholz, B.; Hyde, K.D.; Boekhout, T.; Ebel, R.; Rateb, M.E.; Henderson, L.; Sakayaroj, J.; et al. An online resource for marine fungi. Fungal Divers 2019, 96, 347–433. [Google Scholar] [CrossRef]
- Li, W.; Wang, M.M.; Wang, X.G.; Cheng, X.L.; Guo, J.J.; Bian, X.M.; Cai, L. Fungal communities in sediments of subtropical Chinese seas as estimated by DNA metabarcoding. Sci. Rep. 2016, 6, 26528. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, M.M.; Pan, H.Q.; Burgaud, G.; Liang, S.; Guo, J.; Luo, T.; Li, Z.; Zhang, S.; Cai, L. Highlighting patterns of fungal diversity and composition shaped by ocean currents using the East China Sea as a model. Mol. Ecol. 2018, 27, 564–576. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.M.; Ma, Y.Y.; Cai, L.; Tedersoo, L.; Bahram, M.; Burgaud, G.; Long, X.; Zhang, S.; Li, W. Seasonal dynamics of mycoplankton in the Yellow Sea reflect the combined effect of riverine inputs and hydrographic conditions. Mol. Ecol. 2021, 30, 3624–3637. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, M.M.; Bian, X.M.; Guo, J.; Cai, L. A high-level fungal diversity in the intertidal sediment of Chinese Seas presents the spatial variation of community composition. Front. Microbiol. 2016, 7, 2098. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, M.M.; Burgaud, G.; Yu, H.; Cai, L. Fungal community composition and potential depth-related driving factors impacting distribution pattern and trophic modes from epi- to abyssopelagic zones of the Western Pacific Ocean. Microb. Ecol. 2019, 78, 820–831. [Google Scholar] [CrossRef] [PubMed]
- Wong Chin, J.M.; Puchooa, D.; Bahorun, T.; Jeewon, R. Antimicrobial properties of marine fungi from sponges and brown algae of Mauritius. Mycology 2021, 22, 231–244. [Google Scholar] [CrossRef] [PubMed]
- Mou, X.F.; Liu, X.; Xu, R.F.; Wei, M.Y.; Fang, Y.W.; Shao, C.L. Scopuquinolone B, a new monoterpenoid dihydroquinolin-2(1H)-one isolated from the coral-derived Scopulariopsis sp. Fungus. Nat. Prod. Res. 2018, 32, 773–776. [Google Scholar] [CrossRef]
- Kramer, A.; Beck, H.C.; Kumar, A.; Kristensen, L.P.; Imhoff, J.F.; Labes, A. Proteomic analysis of anti-cancerous Scopularide production by a marine Microascus brevicaulis strain and its UV mutant. PLoS ONE 2015, 10, e0140047. [Google Scholar] [CrossRef]
- Wang, X.F.; Ji, G.X.; Cun, J.F.; Xu, P.J.; Wang, X.W.; Ren, G.W.; Li, W. Screening of insecticidal and antifungal activities of the culturable fungi isolated from the intertidal zones of Qingdao, China. J. Fungi 2022, 8, 1240. [Google Scholar] [CrossRef]
- Zeng, C.K. Seaweeds in Yellow Sea and Bohai Sea of China; Science Press: Beijing, China, 2009; 453p. [Google Scholar]
- Zhang, Z.F.; Liu, F.; Zhou, X.; Liu, X.; Liu, S.; Cai, L. Culturable mycobiota from Karst caves in China, with descriptions of 20 new species. Persoonia 2017, 39, 1–31. [Google Scholar] [CrossRef]
- Suryanarayanan, T.S.; Venkatachalam, A.; Thirunavukkarasu, N.; Ravishankar, J.P.; Doble, M.; Geetha, V. Internal mycobiota of marine macroalgae from the Tamilnadu coast: Distribution, diversity and biotechnological potential. Bot. Mar. 2010, 53, 457–468. [Google Scholar] [CrossRef]
- Zhang, K.; Su, Y.Y.; Cai, L. An optimized protocol of single spore isolation for fungi. Cryptogam. Mycol. 2013, 34, 349–356. [Google Scholar] [CrossRef]
- Nirenberg, H.I. Untersuchungen über die morphologische und biologische differenzierung in der Fusarium-Sektion Liseola. Mitteilungen Biol. Bundesanst. Land Forstwirtsch. 1976, 169, 1–117. [Google Scholar]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour, 3rd ed.; Eyre Methuen: London, UK, 1978. [Google Scholar]
- Guo, L.D.; Hyde, K.D.; Liew, E.C.Y. Identification of endophytic fungi from Livistona chinensis based on morphology and rDNA sequences. New Phytol. 2000, 147, 617–630. [Google Scholar] [CrossRef] [PubMed]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [PubMed]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to the Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Rehner, S.A.; Buckley, E.A. Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: Evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 2005, 97, 84–98. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Env. Microbiol 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef]
- Posada, D. jModelTest: Phylogenetic ModelAveraging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat Methods 2012, 9, 772. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Ronquist, F. MrBayes: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef]
- Wang, M.M.; Chen, Q.; Diao, Y.Z.; Duan, W.; Cai, L. Fusarium incarnatum-equiseti complex from China. Persoonia 2019, 43, 70–89. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Nylander, J.A.A.; Wilgenbusch, J.C.; Warren, D.L.; Swofford, D.L. AWTY (are we there yet?): A system for graphical exploration of MCMC convergence in Bayesian phylogenetics. Bioinformatics 2008, 24, 581–583. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate Maximum-Likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Morelet, M. Micromycètes du var et d’ailleurs (2me Note). Ann. Société Sci. Nat. D’archéologie Toulon Var. 1969, 21, 104–106. [Google Scholar]
- Sandoval-Denis, M.; Gené, J.; Sutton, D.A.; Cano-Lira, J.; de Hoog, G.; Decock, C.; Wiederhold, N.; Guarro, J. Redefining Microascus, Scopulariopsis and allied genera. Persoonia 2016, 36, 1–36. [Google Scholar] [CrossRef]
- von Arx, J.A. The Genera of Fungi Sporulating in Pure Culture, 3rd ed.; V.J. Cramer: Vaduz, Liechtenstein, 1981; pp. 1–424. [Google Scholar] [CrossRef]
- Crous, P.W.; Wingfield, M.J.; Burgess, T.I.; Carnegie, A.; Hardy, G.; Smith, D.; Summerell, B.; Cano-Lira, J.; Guarro, J.; Houbraken, J.; et al. Fungal Planet description sheets: 625-715. Persoonia 2017, 39, 270–467. [Google Scholar] [CrossRef]
- Malloch, D. Wardomyces aggregatus sp. nov. and its possible relationship to Gymnodochium fimicolum. Can. J. Bot. 1970, 48, 883–885. [Google Scholar] [CrossRef]
- Malloch, D.; Cain, R.F. The genus Kernia. Can. J. Bot. 1971, 49, 855–867. [Google Scholar] [CrossRef]
- Guarro, J.; Gené, J.; Stchigel, A.M.; Figueras, M.J. Atlas of soil ascomycetes. In CBS Biodiversity Series; CBS-KNAW Fungal Biodiversity Centre: Utrecht, The Netherlands, 2012. [Google Scholar]
- Gilgado, F.; Cano, J.; Gené, J.; Sutton, D.A.; Guarro, J. Molecular and phenotypic data supporting distinct species statuses for Scedosporium apiospermum and Pseudallescheria boydii and the proposed new species Scedosporium dehoogii. J. Clin. Microbiol. 2008, 46, 766–771. [Google Scholar] [CrossRef] [PubMed]
- Gilgado, F.; Cano, J.; Gené, J.; Guarro, J. Molecular phylogeny of the Pseudallescheria boydii species complex: Proposal of two new species. J. Clin. Microbiol. 2005, 43, 4930–4942. [Google Scholar] [CrossRef] [PubMed]
- Ramsperger, M.; Duan, S.Y.; Sorrell, T.C.; Meyer, W.; Chen, S.C.A. The genus Scedosporium and Pseudallescheria: Current challenges in laboratory diagnosis. Curr. Clin. Micro. Rpt. 2014, 1, 27–36. [Google Scholar] [CrossRef]
- von Arx, J.A. The genera Petriellidium and Pithoascus (Microascaceae). Persoonia 1973, 7, 367–375. [Google Scholar]
- Burgaud, G.; Edgcomb, V.P.; Hassett, B.T.; Kumar, A.; Li, W.; Mara, P.; Peng, X.; Philippe, A.; Phule, P.; Prado, S.; et al. Marine fungi. In The Marine Microbiome; Stal, L.J., Cretoiu, M.S., Eds.; Springer: Cham, Switzerland, 2022; Volume 2, pp. 243–295. [Google Scholar]
- Yu, Z.G.; Lang, G.; Kajahn, I.; Schmaljohann, R.; Imhoff, J.F. Scopularides A and B, cyclodepsipeptides from a marine sponge-derived fungus, Scopulariopsis brevicaulis. J. Nat. Prod. 2008, 71, 1052–1054. [Google Scholar] [CrossRef] [PubMed]
- Steele, C.W. Fungus populations in marine waters and coastal sands of the Hawaiian Line, and Phenix Islands. Pac. Sci. 1967, 21, 317–331. [Google Scholar]
- von Arx, J.A. Revision of Microascus with the description of a new species. Persoonia 1975, 8, 191–197. [Google Scholar]
- Jayawardena, R.S.; Hyde, K.D.; Wang, S.; Sun, Y.-R.; Suwannarach, N.; Sysouphanthong, P.; Abdel-Wahab, M.A.; Abdel-Aziz, F.A.; Abeywickrama, P.D.; Abreu, V.P.; et al. Fungal diversity notes 1512–1610: Taxonomic and phylogenetic contributions on genera and species of fungal taxa. Fungal Divers 2022, 117, 1–272. [Google Scholar] [CrossRef]
- Abdel-Fattah, H.M.; Moubasher, A.H.; Abdel-Hafez, S.I. Studies on mycoflora of salt marshes in Egypt. I. Sugar Fungi. Mycopathologia 1977, 61, 19–26. [Google Scholar] [CrossRef]


| Species | Isolate and Status | Source | Origin | GenBank Accession Number | |||
|---|---|---|---|---|---|---|---|
| LSU | ITS | TEF | TUB | ||||
| Acaulium acremonium | MUCL 8274 = CBS 104.65 ET | Wheatfield soil | Schleswig-Holstein, Germany | KY852479 | KY852468 | -- | -- |
| A. album | CBS 539.85 ET | Hair in dung in pole cat | The Netherlands | KY852484 | KY852473 | -- | -- |
| Canariomyces arenarius | CBS 507.74 IT | Desert soil | Egypt | KM655383 | KM655344 | KM655474 | MK926898 |
| Ca. microsporus | CBS 276.74 IT | Desert soil | Egypt | MH872590 | MH860852 | MN078437 | MK926899 |
| Ca. notabilis | CBS 548.83 T | Litter of Phoenix canariensis | Egypt | MH873362 | MH861648 | MN078432 | MK926902 |
| Ca. subthermophilus | CBS 509.74 T | Desert soil | Egypt | MH872612 | MH860876 | MN078436 | MK926904 |
| Ca. vonarxii | CBS 160.80 = NHL 2831 T | Dried flower of Hibiscus | Sudan | -- | -- | MN078435 | MK926905 |
| Cephalotrichum asperulum | CBS 582.71 T | Soil | Buenos Aires, Argentina | LN851007 | LN850960 | LN851061 | LN851114 |
| Ce. brevistipitatum | CBS 157.57 T | Tuber | Wageningen, The Netherlands | LN851031 | LN850984 | LN851084 | LN851138 |
| Ce. columnare | CBS 159.66 T | Dung of hare | Johannesburg, South Africa | LN851010 | LN850963 | LN851064 | LN851117 |
| Ce. cylindricum | UAMH 1348 T | Seed of sorghum | Kansas, USA | LN851012 | LN850965 | LN851066 | LN851119 |
| Ce. dendrocephalum | CBS 528.85 T | Cultivated soil | Basrah, Iraq | LN851013 | LN850966 | LN851067 | LN851120 |
| Ce. gorgonifer | UAMH 3585 | Mushroom compost | Alberta, Canada | LN851025 | LN850978 | LN851078 | LN851132 |
| Ce. guizhouense | CGMCC 3.18330 T | Air from cave | Guizhou, China | MF419758 | MF419788 | MF419728 | MF434549 |
| Ce. hinnuleum | CBS 289.66 T | Dung of deer | Tasmania, Australia | LN851032 | LN850985 | LN851085 | LN851139 |
| Ce. laeve | CGMCC 3.18329 T | Limestone from cave | Guizhou, China | MF419778 | MF419808 | MF419748 | MF434569 |
| Ce. microsporum | CBS 523.63 ET | Wheatfield soil | Schleswig-Holstein, Germany | LN851014 | LN850967 | LN851068 | LN851121 |
| WL00192 | Unidentified red algae | Qingdao, Shandong, China | OR339938 | OR339892 | OR347703 | OR338658 | |
| WL00303 | Unidentified green algae | Weihai, Shandong, China | OR339939 | OR339893 | OR347704 | OR338659 | |
| WL00305 | Unidentified green algae | Weihai, Shandong, China | OR339940 | OR339894 | OR347705 | OR338660 | |
| WL00350 | Unidentified brown algae | Weihai, Shandong, China | OR339941 | OR339895 | OR347706 | OR338661 | |
| Ce. nanum | CBS 191.61 ET | Dung of deer | Surrey, England | LN851016 | LN850969 | LN851070 | LN851123 |
| WL03115 | Intertidal sediment | Ningbo, Zhejiang, China | OR339945 | OR339899 | OR347710 | OR338662 | |
| WL03362 | Intertidal sediment | Ningbo, Zhejiang, China | OR339946 | -- | OR347711 | -- | |
| Ce. oligotriphicum | CGMCC 3.18328 T | Limestone from cave | Guizhou, China | MF419771 | MF419801 | MF419741 | MF434562 |
| Ce. purpureofuscum | UAMH 9209 | Indoor air | British Columbia, Canada | LN851018 | LN850971 | LN851072 | LN851125 |
| WL06350 | Sargassum canfusum | Shenzhen, Guangdong, China | OR339942 | OR339896 | OR347707 | OR338664 | |
| WL06351 | Sargassum canfusum | Shenzhen, Guangdong, China | OR339943 | OR339897 | OR347708 | OR338665 | |
| WL02022 | Sargassum canfusum | Shenzhen, Guangdong, China | OR339950 | OR339898 | OR347709 | OR338663 | |
| Ce. stemonitis | CBS 103.19 NT | Seed | Wageningen, Netherlands | LN850952 | LN850951 | LN850953 | LN850954 |
| Ce. verrucisporum | CBS 187.78 | Sand dune soil | Katijk, The Netherlands | LN851033 | LN850986 | LN851086 | LN851140 |
| Gamsia aggregata | CBS 251.69 ET | Dung of carnivore | USA | LM652500 | LM652378 | -- | -- |
| G. columbina | CBS 233.66 ET | Sandy soil | Giessen, Germany | LN851039 | LN850990 | LN851092 | LN851146 |
| G. dimera | CBS 235.66 = ATCC 18887 = IMI 117371 = MUCL 6388 T | Wheatfield soil | Schleswig-Holstein, Germany | -- | MH858785 | -- | -- |
| G. kooimaniorum | CBS 143185 T | Garden soil | The Netherlands | -- | NR_159824 | -- | -- |
| G. sedimenticola | WL02722 = CGMCC3.25342 T | Intertidal sediment | Qingdao, Shandong, China | OR339947 | OR339900 | OR347712 | OR338666 |
| WL06358 | Intertidal sediment | Qingdao, Shandong, China | OR339948 | OR339901 | OR347713 | OR338667 | |
| WL06359 | Intertidal sediment | Qingdao, Shandong, China | OR339949 | OR339902 | OR347714 | OR338668 | |
| G. simplex | CBS 546.69 IT | Milled Oryza sativa | Japan | LM652501 | LM652379 | LN851094 | LN851148 |
| Graphium penicillioides | CBS 102632 T | Populus nigra | Czech Republic | KY852485 | KY852474 | -- | -- |
| Kernia columnaris | CBS 159.66 = IMI 116691 T | Dung of hare | South Africa | KY852486 | KY852475 | KY852478 | KY852477 |
| K. nitida | CBS 282.52 = NBRC 8200 | Chrysolina sanguinolenta | France | KY852487 | KY852476 | -- | -- |
| Microascus algicola | WL00347 | Unidentified brown algae | Weihai, Shandong, China | OR339954 | OR339907 | OR347719 | OR338669 |
| WL00633 | Colpomenia sinuosa | Qingdao, Shandong, China | OR339955 | OR339908 | OR347720 | OR338670 | |
| WL01268 = CGMCC3.25343 T | Grateloupia filicina | Qingdao, Shandong, China | OR339962 | OR339909 | OR347721 | OR338672 | |
| WL01021 | Ahnfeltiopsis flabelliformis | Qingdao, Shandong, China | OR339957 | OR339910 | OR347722 | OR338671 | |
| M. campaniformis | CBS 138126 = UTHSC 10-565 = FMR 12343 T | Human BAL | USA | HG380495 | LM652391 | HG380418 | LM652606 |
| M. cinereus | FMR 12217 = UTHSC 10-2805 NT | Human BAL | USA | -- | LM652397 | -- | LM652611 |
| M. croci | CBS 158.44 = MUCL 9002 T | Crocus sp. | Lisse, The Netherlands | LM652508 | LM652407 | LM652560 | LM652621 |
| WL02042 | Blidingia minima | Qingdao, Shandong, China | OR339982 | OR339903 | OR347715 | OR338673 | |
| M. expansus | CBS 138127 = UTHSC 06-4472 = FMR 12266 T | Human sputum | USA | HG380492 | LM652410 | HG380415 | LM652624 |
| M. gennadii | WL02353 = CGMCC3.25344 T | Intertidal sediment | Unknown | OR339971 | OR339904 | OR347716 | OR338674 |
| WL06354 | Intertidal sediment | Unknown | OR339952 | OR339905 | OR347717 | OR338675 | |
| WL06355 | Intertidal sediment | Unknown | OR339953 | OR339906 | OR347718 | OR338676 | |
| M. gracilis | CBS 369.70 IT | Food | Japan | HG380467 | LM652412 | HG380390 | LM652625 |
| M. hyalinus | CBS 766.70 IT | Dung of cow | USA | LM652513 | LM652418 | LM652564 | LM652631 |
| M. intricatus | CBS 138128 = UTHSC 07-156 = FMR 12264 T | Human BAL | USA | HG380496 | LM652419 | HG380419 | LM652632 |
| M. longirostris | CBS 196.61 = MUCL 9058 NT | Wasp’s nest | Maine, USA | LM652515 | LM652421 | LM652566 | LM652634 |
| M. murinus | CBS 830.70 = IHEM 18567 T | Composed municipal waste | Giessen, Germany | HG380481 | LM652424 | HG380404 | LM652637 |
| WL05483 | Intertidal sediment | Qingdao, Shandong, China | OR339959 | OR339912 | OR347724 | OR338678 | |
| WL03684 | Intertidal sediment | Yantai, Shandong, China | OR339958 | OR339911 | OR347723 | OR338677 | |
| M. paisii | UTHSC 07-639 = FMR 12263 | Human BAL | USA | HG380451 | LM652425 | HG380374 | LM652638 |
| M. pyramidus | CBS 212.65 IT | Desert soil | California, USA | HG380435 | LM652439 | HG380358 | LM652652 |
| M. restrictus | CBS 138277 = UTHSC 09-2704 = FMR 12227 T | Human left hallux | USA | HG380494 | LM652440 | HG380417 | LM652653 |
| WL02392 | Intertidal sediment | Huludao, Liaoning, China | -- | -- | OR347725 | OR338679 | |
| M. senegalensis | CBS 277.74 = IHEM 18561 T | Mangrove soil | Senegal | LM652523 | LM652441 | LM652574 | LM652654 |
| M. sparsimycelialis | LC12478 T | Animal faeces | Laibin, Guangxi, China | -- | -- | MK336046 | MK336124 |
| M. superficialis | LC12597 T | Animal faeces | Laibin, Guangxi, China | -- | -- | MK336048 | MK336126 |
| M. trigonosporus | CBS 218.31 T | na | USA | HG380436 | LM652443 | HG380359 | LM652655 |
| WL00670 | Cladophora sp. | Qingdao, Shandong, China | OR339961 | OR339916 | OR347729 | OR338683 | |
| WL01853 | Ulva linza | Qingdao, Shandong, China | OR339963 | OR339917 | OR347730 | OR338684 | |
| WL01882 | Gracilaria textorii | Qingdao, Shandong, China | OR339944 | OR339918 | OR347731 | OR338685 | |
| WL02833 | Intertidal sediment | Qingdao, Shandong, China | OR339964 | OR339919 | OR347732 | OR338686 | |
| M. trigonus | LC12520 T | Soil | Guilin, Guangxi, China | -- | -- | MK336052 | MK336130 |
| M. verrucosus | CBS 138278 = UTHSC 10-2601 = FMR 12219 T | Human, BAL fluid | USA | HG380493 | LM652446 | HG380416 | LM652658 |
| WL02828 | Intertidal sediment | Dongying, Shandong, China | OR339965 | OR339920 | OR347733 | OR338687 | |
| Microascus sp. | WL01046 | Grateloupia filicina | Qingdao, Shandong, China | OR339956 | OR339913 | OR347726 | OR338680 |
| WL06352 | Grateloupia filicina | Qingdao, Shandong, China | -- | OR339914 | OR347727 | OR338681 | |
| WL06353 | Grateloupia filicina | Qingdao, Shandong, China | -- | OR339915 | OR347728 | OR338682 | |
| Parascedosporium tectonae | CBS 127.84 T | Human BAL | USA | EF151332 | -- | -- | -- |
| Petriella sordida | CBS 124169 | Corner of a bathroom | The Netherlands | AY281099 | GQ426957 | -- | -- |
| Pithoascus ater | CBS 400.34 = IHEM 18608 T | na | na | LM652526 | LM652447 | LM652576 | LM652659 |
| Pi. exsertus | CBS 819.70 T | Megachile willoughbiella | Tastrup, Denmark | LM652528 | LM652449 | LM652578 | LM652661 |
| Pi. intermedius | CBS 217.32 T | Root of Fragaria vesca | North Carolina, USA | LM652529 | LM652450 | LM652579 | LM652662 |
| Pi. nidicola | CBS 197.61 ET | Dipodomys merriami | Utah, USA | LM652530 | LM652451 | LM652580 | LM652663 |
| Pi. stoveri | CBS 176.71 T | Root of Beta vulgaris | Ohio, USA | LM652532 | LM652453 | LM652581 | LM652664 |
| Pseudoscopulariopsis asperispora | LC12445 T | Animal faeces | Guilin, Guangxi, China | -- | -- | MK336064 | MK336142 |
| Ps. hibernica | UAMH 2643 = ATCC 16690 | Soil | Ireland | LM652533 | LM652454 | LM652582 | LM652665 |
| Ps. schumacheri | CBS 435.86 NT | Soil | Puerto de la Quesera, Spain | LM652534 | LM652455 | LM652583 | LM652666 |
| Rhinocladium lesnei | CBS 108.10 | Foot of human | France | MH866122 | MH854594 | -- | -- |
| Scedosporium angustum | CBS 254.72 ET | Sewage half digestion tank | Ohio, USA | -- | AJ888414 | -- | AJ889604 |
| S. apiospermum | FMR 8619 ET | Keratitis of huma | Brazil | -- | NR_130664 | -- | AJ889584 |
| S. aurantiacum | IMI 392886 T | Ulcer on ankle of human | Spain | -- | AJ888440 | -- | AJ889597 |
| S. boydii | CBS 101.22 = IMI 015407 = JCM 7441 = NCPF 2216 = UAMH 3982 T | Mycetoma | USA | EF151320 | KT008518 | KT069589 | KT008455 |
| WL02794 | Intertidal sediment | Qingdao, Shandong, China | OR339968 | OR339923 | OR347736 | OR338688 | |
| WL05602 | Intertidal sediment | Yantai, Shandong, China | OR339969 | OR339924 | OR347737 | OR338689 | |
| WL05951 | Intertidal sediment | Tianjin, China | OR339970 | OR339925 | OR347738 | OR338690 | |
| S. cereisporum | FMR 12996 ET | Wastewater sludge | Mûrs-Erignés, France | -- | KJ599660 | -- | KJ599659 |
| S. dehoogii | CBS 117406 ET | Garden soil | Barcelona, Spain | -- | KT163400 | -- | KT163401 |
| S. desertorum | CBS 489.72 ET | Salt-marsh soil | Kuwait | -- | -- | -- | AM409106 |
| S. ellipsoideum | CBS 418.73 ET | Soil | Tayikistán | -- | AJ888426 | -- | AJ889595 |
| S. ellipsosporium | WL02370 = CGMCC3.25345 T | Intertidal sediment | Shenzhen, Guangdong, China | OR339973 | OR339926 | OR347739 | OR338692 |
| WL02793 | Intertidal sediment | Qingdao, Shandong, China | OR339972 | OR339927 | OR347740 | OR338693 | |
| S. fusoideum | CBS 106.53 ET | Goat dung Aligarh | India | -- | AJ888428 | -- | AJ889601 |
| S. haikouense | CGMCC 3.20468 = GZUIFR 21.833 T | Green-belt soil | Haikou, Hainan, China | -- | MZ469289 | -- | MZ488563 |
| S. hainanense | CGMCC 3.20469 = GZUIFR 21.829 T | Green-belt soil | Sanya, Hainan, China | -- | MZ469285 | -- | MZ488559 |
| WL05931 | Intertidal sediment | Yancheng, Jiangsu, China | OR339967 | OR339922 | OR347735 | OR338691 | |
| S. minutisporum | FMR 4072 ET | River sediment Tordera river | Spain | -- | AJ888384 | -- | AJ889592 |
| S. multisporum | CGMCC 3.20470 = GZUIFR 21.830 T | Green-belt soil | Huaihua, Hunan, China | -- | MZ469286 | -- | MZ488560 |
| S. rarisporum | GZUIFR-G79 T | Root soil of Wodyetia bifurcata | Guigang, Guangxi, China | -- | KX790702 | -- | -- |
| S. shenzhenensis | WL02375 = CGMCC3.25346 T | Intertidal sediment | Shenzhen, Guangdong, China | OR339960 | OR339928 | OR347741 | OR338694 |
| WL06356 | Intertidal sediment | Shenzhen, Guangdong, China | OR339974 | OR339929 | OR347742 | OR338695 | |
| WL06357 | Intertidal sediment | Shenzhen, Guangdong, China | OR339975 | OR339930 | OR347743 | OR338696 | |
| S. sphaerospermum | WL02796 = CGMCC3.25347 T | Intertidal sediment | Qingdao, Shandong, China | OR339976 | OR339931 | OR347744 | OR338698 |
| WL06360 | Intertidal sediment | Qingdao, Shandong, China | OR339977 | OR339932 | OR347745 | OR338699 | |
| WL06361 | Intertidal sediment | Qingdao, Shandong, China | OR339978 | OR339933 | OR347746 | OR338700 | |
| Scedosporium sp. | WL02426 | Intertidal sediment | na | OR339966 | OR339921 | OR347734 | OR338697 |
| Scopulariopsis africana | CBS 118736 T | Mud, salt pan | South Africa | KX924040 | KX923954 | KX924176 | KX924388 |
| Sc. albida | CBS 119.43 T | Soil | The Netherlands | LN850849 | LN850800 | LM652592 | LN850897 |
| Sc. alboflavescens | CBS 399.34 T | Diseased skin | Austria | LM652539 | KX923956 | KX924179 | JQ434537 |
| Sc. asperula | CBS 853.68 | Compost soil | Germany | JQ434669 | LM652461 | JQ434621 | JQ434558 |
| Sc. brevicaulis | MUCL 40726 T | Indoor air | Alberta, Canada | HG380440 | LM652465 | HG380363 | LM652672 |
| WL00657 | Ulva pertusa | Qingdao, Shandong, China | OR339979 | OR339934 | OR347747 | OR338701 | |
| WL03882 | Intertidal sediment | Ningde, Fujian, China | OR339980 | OR339935 | OR347748 | OR338702 | |
| Sc. candida | MUCL 40743 ET | Indoor air | Canada | HG380458 | LM652484 | HG380381 | LM652690 |
| Sc. caseicola | CBS 480.62 T | Cheese coating | The Netherlands | KX924041 | KX924020 | KX924247 | KX924454 |
| Sc. cordiae | FMR 12338 T | Human finger | USA | HG380499 | LM652491 | HG380422 | LM652673 |
| Sc. flava | CBS 207.61 = MUCL 9031 NT | Cheese | UK | HG380464 | LM652493 | HG380387 | LM652697 |
| Sc. macurae | CBS 506.66 T | Chicken litter | Canada | LN850854 | LN850805 | KX924250 | LN850902 |
| Sc. sexualis | CBS 250.64 T | Oryza sativa | Burma | KX924042 | KX924024 | KX924251 | KX924458 |
| Sc. soppii | UAMH 9169 T | Wood of Populus tremuloides | Alberta, Canada | LM652552 | LM652495 | LM652595 | LM652698 |
| Wardomyces anomalus | CBS 299.61 ET | Air cell of egg | Ontario, Canada | MH869626 | MH858058 | LN851095 | LN851149 |
| W. giganteus | CBS 746.69 T | Insect frass in dead log | Ontario, Canada | MH871180 | MH859408 | LN851096 | LN851150 |
| W. inflatus | CBS 216.61 IT | Wood, Acer sp. | Quebec, Canada | LM652553 | LM652496 | LN851098 | -- |
| WL02318 | Intertidal sediment | Huludao, Liaoning, China | OR339951 | OR339937 | OR347750 | OR338704 | |
| WL00510 | Intertidal sediment | Weihai, Shandong, China | OR339981 | OR339936 | OR347749 | OR338703 | |
| W. ovalis | CBS 234.66 = IMI 117372 = MUCL 6031 T | Wheatfield soil | Schleswig-Holstein, Germany | LN851050 | MH858784 | LN851101 | LN851155 |
| W. pulvinatus | CBS 112.65 IT | Salt marsh | Cheshire, England | MH870142 | MH858508 | LN851102 | LN851156 |
| Wardomycopsis dolichi | LC12503 T | Soil | Guilin, Guangxi, China | MK329043 | MK329138 | MK336073 | -- |
| Wa. fusca | LC12476 T | Soil | Guilin, Guangxi, China | MK329047 | MK329142 | MK336077 | MK336148 |
| Wa. humicola | CBS 487.66 T | Soil | Ontario, Canada | LM652554 | LM652497 | -- | -- |
| Wa. inopinata | FMR 10305 | Soil | Myanmar | LM652555 | LM652498 | -- | -- |
| Wa. litoralis | CBS 119740 T | Beach soil | Castellon, Spain | LN851055 | LN851000 | LN851107 | LN851161 |
| Wa. longicatenata | CGMCC 3.17947 T | Air | Guizhou, China | KU746756 | KU746710 | KX855255 | KU746801 |
| Yunnania carbonaria | CBS 205.61 T | Soil | Panama | HG380462 | KX923820 | KX924044 | KX924254 |
| Y. penicillata | CBS 130296 T | Moulded pork sample | China | KY659809 | JN831361 | KY659808 | KY659807 |
| Y. smithii | CBS 855.68 T | Garden soil | Germany | KX924028 | KX923822 | KX924046 | KX924256 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.-M.; Yang, S.-Y.; Li, Q.; Zheng, Y.; Ma, H.-H.; Tu, Y.-H.; Li, W.; Cai, L. Microascaceae from the Marine Environment, with Descriptions of Six New Species. J. Fungi 2024, 10, 45. https://doi.org/10.3390/jof10010045
Wang M-M, Yang S-Y, Li Q, Zheng Y, Ma H-H, Tu Y-H, Li W, Cai L. Microascaceae from the Marine Environment, with Descriptions of Six New Species. Journal of Fungi. 2024; 10(1):45. https://doi.org/10.3390/jof10010045
Chicago/Turabian StyleWang, Meng-Meng, Shi-Yu Yang, Qi Li, Yao Zheng, He-He Ma, Ye-Hui Tu, Wei Li, and Lei Cai. 2024. "Microascaceae from the Marine Environment, with Descriptions of Six New Species" Journal of Fungi 10, no. 1: 45. https://doi.org/10.3390/jof10010045
APA StyleWang, M.-M., Yang, S.-Y., Li, Q., Zheng, Y., Ma, H.-H., Tu, Y.-H., Li, W., & Cai, L. (2024). Microascaceae from the Marine Environment, with Descriptions of Six New Species. Journal of Fungi, 10(1), 45. https://doi.org/10.3390/jof10010045