Abstract
Helvella bachu, an ectomycorrhizal fungus, forms a symbiotic relationship with Populus euphratica, a rare and endangered species crucial to desert riparian ecosystems. In this study, endofungal bacteria (EFBs) within the fruiting bodies of H. bachu were confirmed by a polyphasic approach, including genomic sequencing, real-time quantitative PCR targeting the 16S rRNA gene, full-length and next-generation sequencing (NGS) of the 16S rRNA gene, and culture methods. The genera Stenotrophomonas, Variovorax, Acidovorax, and Pedobacter were abundant in the EFBs of fruiting bodies associated with three Populus hosts and were consistently present across different developmental stages. Notably, S. maltophilia and V. paradoxus were detected in high abundance, as revealed by full-length 16S rRNA sequencing, with S. maltophilia also isolated by culture methods. KO-pathway analysis indicated that pathways related to primary, secondary, and energy metabolism were predominantly enriched, suggesting these bacteria may promote H. bachu growth by producing essential compounds, including sugars, proteins, and vitamins, and secondary metabolites. This study confirmed the presence of EFBs in H. bachu and provided the first comprehensive overview of their structure, functional potential, and dynamic changes throughout fruiting body maturation, offering valuable insights for advancing the artificial domestication of this species.
1. Introduction
Helvella bachu, commonly known as the Bachu mushroom, is a delectable edible fungus found in the Xinjiang Uygur Autonomous Region of China [1], as well as in Pakistan [2] and Iraq [3]. Research indicates that this mushroom can effectively enhance leukocyte phagocytosis, lymphocyte transformation rates, and antibody titers [4,5], while also exhibiting antioxidant activity and a strong inhibitory effect on HepG2 cells [6]. In recent years, it has attracted considerable attention due to its notable economic, nutritional, and medicinal values. However, the annual yield of this mushroom is limited, and fruiting bodies cannot be cultivated, as pure cultures have yet to be successfully established despite years of efforts.
Our previous studies have demonstrated that H. bachu is an ectomycorrhizal fungus. To date, H. bachu has been shown to establish a symbiotic relationship with various poplar species, including Populus euphratica, Populus pruinosa, and Populus alba var. pyramidalis [7] in desert riparian ecosystems. This association plays a crucial role in desert afforestation, but complicates efforts to obtain pure cultures of H. bachu mycelium. Additionally, during the isolation process, bacterial growth is consistently observed around tissue blocks, while mycelial germination remains absent. This suggests the possible presence of endofungal bacteria (EFBs) within the fruiting bodies of H. bachu.
EFBs are bacterial symbionts that reside within fungal hyphae and spores [8]. The presence of EFBs was initially reported by Mosse (1970) through electron microscopy [9], revealing their presence in the cytoplasm of Endogone spores. These bacteria have been identified in a variety of fungal species, including both mycorrhizal and saprophytic fungi, and have had a significant influence on the evolution of life, continuing to shape the ecology of a wide range of species [8,10].
In recent decades, the EFBs associated with mushrooms have been extensively researched. These EFBs exhibited remarkable diversity, with a wide range of taxa identified across various mushroom species. A total of sixty-six EFB isolates were obtained from wild-growing mushrooms [11]. Furthermore, a greater abundance of EFBs has been reported in ectomycorrhizal mushrooms. For instance, Proteobacteria was found to be the most abundant phylum of EFB in the fruiting body of Tylopilus neofelleus, accounting for 94.2%, with Rhizobiales and Burkholderiales also being predominant in this fruiting body [12]. In the dissected fruiting body of Tricholoma matsutake, Micrococcales, Bacillales, Caulobacter, and Sphingomonas emerged as the principal bacterial taxa across its various compartments [13]. Additionally, members of the Allorhizobium–Neorhizobium–Pararhizobium–Rhizobium complex were found to be most abundant in the fruiting bodies of Cantharellus cibarius [14]. Twelve potential endophytic nitrogen-fixing bacteria, including Paenarthrobacter, Exiguobacterium, and Paenibacillus, have been isolated from Floccularia luteovirens [15].
The ecological roles of EFBs in mushrooms have been recognized as both diverse and significant, contributing to various aspects of fungal health and productivity. EFBs could serve as source of bioenergy for fungal hosts, producing essential compounds such as sugars, proteins, vitamins, and auxins that promote hyphal growth and spore germination [16,17,18]. They facilitated nutrient transport, particularly of phosphates, to mycorrhizal structures, thereby enhancing respiration and adaptability to varying environmental conditions [19,20,21]. Certain EFB species, especially those from the Pseudomonas genus, have been identified as potential biological control agents against diseases such as brown blotch and internal stipe necrosis in Agaricus bisporus [11]. Moreover, EFBs associated with Chanterelles have been shown to positively influence fruiting body formation [22], while EFBs in ascocarp-associated bacteria may play a role in the development, maturation, and even the aroma of the Black truffle, Tuber melanosporum [23]. Overall, the intricate interactions between mushrooms and EFBs have provided valuable insights into fungal biology and presented opportunities for improving mushroom cultivation practices.
Understanding the microbiota is essential for the successful domestication of H. bachu, a valuable ectomycorrhizal fungus associated with P. euphratica, an important species in desert riparian ecosystems [24]. In the present study, EFBs were identified in the fruiting bodies of H. bachu during genome sequencing analysis. Their presence was confirmed through real-time quantitative PCR targeting the 16S rRNA gene. The bacterial abundance and community composition of EFBs associated with three types of Populus plants were then characterized using full-length 16S rRNA gene sequencing, along with predictions of their functional roles. Biomarkers specific to EFBs linked to different host plants were also identified. Additionally, EFBs from H. bachu at different developmental stages were analyzed through amplicon sequencing. Finally, EFBs isolated from H. bachu fruiting bodies using culture methods showed partial consistency with the high-throughput sequencing results. This study confirmed the presence of EFBs in H. bachu and provided the first comprehensive overview of their structure, functional potential, and dynamic changes throughout the maturation of H. bachu fruiting bodies.
2. Materials and Methods
2.1. Genome Sequencing and Discovery of EFBs in the Fruiting Body of Helvella bachu
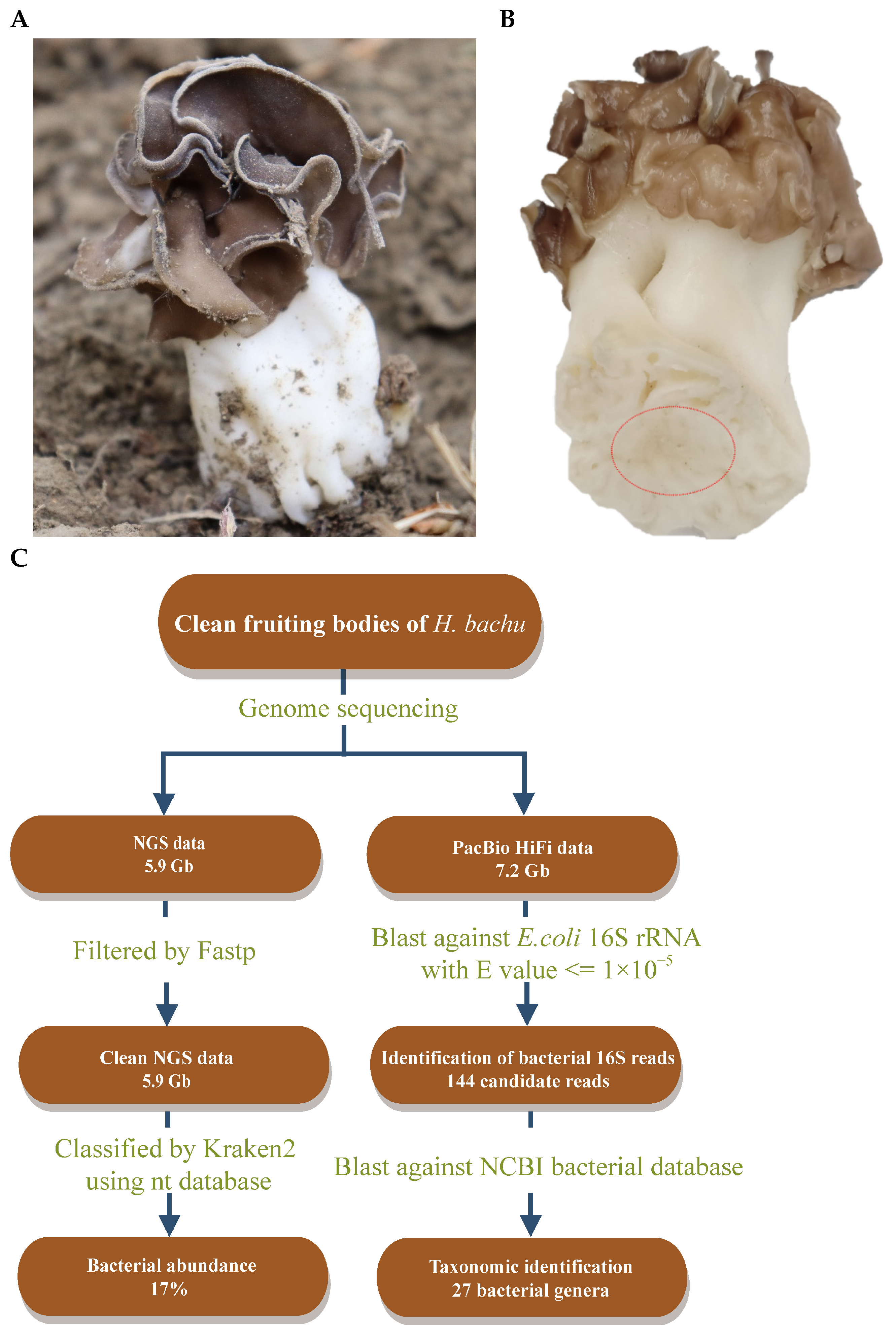
The fruiting bodies of H. bachu (Figure 1A) were collected at Tarim University in Alar City, Xinjiang Uygur Autonomous Region, China. Following the removal of debris and rinsing with sterile water, tissue blocks were carefully excised from the inner base of the H. bachu fruiting body (Figure 1B).
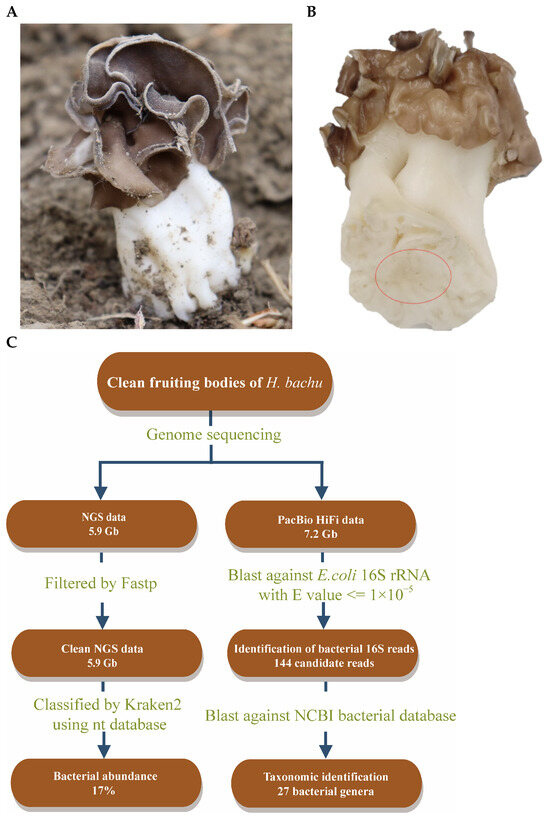
Figure 1.
Discovery of EFBs in fruiting body of Helvella bachu during genome sequencing analysis. (A) Fruiting body of H. bachu in nature; (B) cleaned fruiting body of H. bachu. The sample location was marked with a red circle; (C) flowchart illustrating discovery of EFBs in fruiting body of H. bachu.
High-quality genomic DNA was extracted following the manufacturer’s protocol, using the QIAGEN Genomic Kit (Qiagen, Düsseldorf, Germany). Genome sequencing was performed using the MGISEQ-T7 and PacBio Sequel II platforms, provided by Nextomics Biosciences Co., Ltd. (Wuhan, China). A next-generation sequencing (NGS) library with an average insert size of 350 base pairs (bp) was first prepared following the MGISEQ standard protocol and sequenced on the MGISEQ-T7 platform. Additionally, a 20 kb library was constructed according to PacBio’s standard methodology, with its quality assessed on the Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA) prior to sequencing on the PacBio Sequel II platform (Pacific Biosciences, Menlo Park, CA, USA).
NGS reads were initially filtered using Fastp v0.20.0 [25] with default parameters. The filtered NGS reads were subsequently analyzed with Kraken v2.1.3 [26] to assign taxonomic labels to short DNA sequences (Figure 1C). Following this, a BLAST search (v2.7.1) [27] was conducted using the 16S ribosomal RNA gene sequence of Escherichia coli (MN900682.1) against the original sequenced data, with an e-value threshold set at 10−5. These reads were then subjected to an online BLAST analysis on the NCBI bacterial database (https://blast.ncbi.nlm.nih.gov/Blast.cgi, accessed on 10 November 2023) using the same e-value threshold to identify the taxonomic classification of these bacteria (Figure 1C).
2.2. Analysis of EFBs in Helvella bachu Fruiting Bodies Using Absolute Quantitative PCR
The EFBs of H. bachu was quantified by absolute quantitative real-time PCR, following previously reported methods [13]. DNA was extracted from the clean tissues of the H. bachu fruiting body using a FastDNA SPIN Kit for Soil (MP Biomedicals, Inc., Santa Ana, CA, USA) in accordance with the manufacturer’s instructions. Primers 338F and 518R (338F: ACTCCTACGGGAGGCAGCAG; 518R: ATTACCGCGCGGCTGCTGG) were selected to amplify the V3–V4 region of the 16S rRNA gene sequence [28].
Quantitative PCR (qPCR) was performed in a 10 μL reaction volume using a Bio-Rad CFX Connect Real-Time PCR Detection System (Bio-Rad Laboratories Inc., Hercules, CA, USA). The reaction mixture consisted of 5 μL of Vazyme SYBR Green Master Mix (Vazyme International LLC., Nanjing, China), 0.2 μL of each forward and reverse primer, and 1 μL of DNA template. A plasmid containing a 16S rRNA gene fragment compatible with the bacterial primer pair 338F/518R served as the quantification standard.
The amplified products from H. bachu tissue DNA, generated using the universal 16S rDNA primers (338F/518R), were purified via gel electrophoresis, cloned into the pLB vector, and positive clones were selected for plasmid extraction and concentration measurement. The initial plasmid DNA concentration was 4.7 × 1012 copies/μL, which was serially diluted tenfold to quantify bacterial 16S rRNA gene copies. Utilizing the standard curve construction method, qPCR was performed on DNA extracted from H. bachu tissue samples, with three parallel sets of reactions established to detect Cq values.
The reaction conditions included an initial incubation at 95 °C for 15 s, followed by 40 cycles at 60 °C for 34 s, during which fluorescence signal detection occurred at 60 °C. Melting curves were evaluated for each reaction to confirm the specificity of amplification. All reactions were conducted in technical triplicates, with nuclease-free water samples serving as negative controls across all plates.
2.3. Full-Length 16S rRNA Gene Sequencing by PacBio
The mature fruiting bodies of H. bachu associated with three different host plants, including P. euphratica, P. pruinosa, and P. alba var. pyramidalis, were harvested separately (Table 1). Clean tissue blocks weighing about 0.5 g were taken from the inner base of each sample, with three replicates per sample, and used for the full-length 16S rRNA gene sequencing by PacBio platform at Wuhan Grandomics Biosciences Co., Ltd. (Wuhan, Hubei, China).

Table 1.
Helvella bachu samples used for full-length 16S rRNA gene sequencing.
Genomic DNA was extracted using the CTAB method with a Grandomics Genomic Kit (Grandomic Biosciences Co., Ltd., Wuhan, China), in accordance with the manufacturer’s standard operating procedure. DNA purity was assessed using a NanoDrop™ One UV-Vis spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA), yielding an OD260/280 ratio between 1.8 and 2.0 and an OD260/230 ratio between 2.0 and 2.2. The DNA concentration was further quantified using a Qubit® 4.0 Fluorometer (Invitrogen, Carlsbad, CA, USA).
For 16S rRNA sequencing, the mass-seq method [29] was utilized with a Kinnex 16S rRNA kit (Pacific Biosciences, Menlo Park, CA, USA). A total of 1–2 ng of DNA per sample was used for the amplification of full-length 16S genes (V1–V9 regions) with barcoded forward and reverse 16S primers from a Kinnex PCR 12-fold kit (Pacific Biosciences, Menlo Park, CA, USA). Following amplification, the pooled 16S PCR amplicons were purified using SMRTbell cleanup beads (Pacific Biosciences, Menlo Park, CA, USA). Kinnex adapters were then ligated to the ends of the barcoded full-length amplicons, and the PCR-amplified 16S fragments were treated with Kinnex enzyme and ligase to assemble the 16S segments into a linear array of approximately 19 kb, utilizing a Kinnex concatenation kit (Pacific Biosciences, Menlo Park, CA, USA). The final SMRTbell library was analyzed using an Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA) to determine the size of the library fragments. Sequencing was conducted on a PacBio Revio instrument by Nextomics Biosciences Co., Ltd. (Wuhan, China).
2.4. Next-Generation Sequencing (NGS) of 16S rRNA Gene Amplicons
To investigate the changes of EFBs throughout the development of the H. bachu fruiting body (host P. euphratica), clean tissue samples were collected from the inner base of the fruiting body at three distinct developmental stages, including nascent, developing, and mature fruiting bodies, with three biological replicates for each stage. DNA was extracted using the TGuide S96 Magnetic Soil/Stool DNA Kit (Tiangen Biotech Co., Ltd., Beijing, China) following the manufacturer’s protocol. DNA concentration was measured with the Qubit dsDNA HS Assay Kit and Qubit 4.0 Fluorometer (Invitrogen, Thermo Fisher Scientific Inc., Waltham, MA, USA). To amplify the V3–V4 region of the 16S rRNA gene, a universal primer set (338F: 5′–ACTCCTAC–GGGAGGCAGCA–3′ and 806R: 5′–GGACTACHVGGGTWTCTAAT–3′) was employed. Sequencing was conducted on the Illumina NovaSeq 6000 platform (Illumina, Santiago, CA, USA) by Biomarker Technologies Co., Ltd. (Beijing, China).
2.5. Data Processing for Full-Length Sequencing and NGS of the16S rRNA Gene
Data processing was carried out with the support of BMK Cloud (Biomarker Technologies Co., Ltd., Beijing, China). The analytical pipeline included data quality control, species annotation, and analyses of alpha and beta diversity, along with assessments of differential species and functional characteristics. The QIIME2 feature classifier was employed to compare reads after quality control measures, which involved noise removal and the elimination of chimeras and mitochondrial sequences, using the Silva database (https://qiime2.org/). This alignment produced abundance tables of species composition across various taxonomic levels for each sample.
For all samples, alpha diversity indices—including observed species, Shannon, Simpson, Goods coverage, Chao1, ACE, and Pielou—along with beta diversity indices, were calculated using the R packages vegan, picante, and combinat. Differential analysis was conducted using LEfSe (https://huttenhower.sph.harvard.edu/galaxy/, accessed on 12 September 2024) [30] software to identify biomarkers across two or more groups. PICRUSt2 (https://github.com/picrust/picrust2, accessed on 16 September 2024) [31] was employed to predict the functions of representative sequences. Functional KO-pathway analysis was performed to forecast the KO results for different samples, and the corresponding functional pathway table was utilized to calculate abundance across various pathways. All boxplots were generated using the RandomcoloR (https://www.npmjs.com/package/randomcolor, accessed on 19 September 2024) and ggplot2 packages (https://ggplot2.tidyverse.org/, accessed on 10 September 2024).
2.6. Isolation and Identification EFBs from Helvella bachu Fruiting Bodies Using Culture Methods
The EFBs from H. bachu fruiting bodies associated with P. euphratica were isolated using four different media: Luria–Bertani (tryptone 10 g/L, Yeast extract 5 g/L, NaCl 10 g/L, pH 7); R2A (yeast extract powder 0.5 g, peptone 0.5 g, casein hydrolysate 0.5 g, glucose 0.5 g, soluble starch 0.5 g, dipotassium hydrogen phosphate 0.3 g, anhydrous magnesium sulfate 0.024 g, pyruvate 0.3 g, agar 15.0 g, pH 7.2 ± 0.2 [25]; TSA (cheese peptone 15.0 g, soybean papain hydrolysate 5.0 g, sodium chloride 5.0 g, agar 15.0 g, distilled water 1000 mL, pH 7.3 ± 0.2) [32], and YMG (glucose 4.0 g, yeast extract 4.0 g, malt extract 10.0 g, agar 15.0 g, distilled water 1000.0 mL, pH 7.2). The bacteria were purified and identified using universal primers 27F and 1492R.
3. Results
3.1. Discovery of EFBs in the Fruiting Body of Helvella bachu During Genome Sequencing Analysis
Clean tissue blocks were collected from the inner base of the H. bachu fruiting body (Figure 1A,B) to eliminate any risk of contamination. Genome sequencing was conducted using the MGISEQ-T7 and PacBio Sequel II platforms, yielding a total of 5.9 Gb of NGS reads and 7.2 Gb of PacBio HiFi reads, respectively. The generated reads were analyzed using Kraken 2.0, a taxonomic sequence classifier that assigns taxonomic labels to short DNA sequences [26].
The results indicated that 73% of the reads were fungal, while bacteria constituted 17% (Figure 1C). Subsequently, a BLAST search was performed using the 16S ribosomal RNA gene sequence of E. coli (MN900682.1) against the original sequenced data, with an e-value threshold set at 10−5, yielding 144 reads (Figure 1C). These reads were then queried against the NCBI bacterial database, also using an e-value threshold of 10−5. This analysis identified 27 bacterial genera, including Acidovorax, Pedobacter, Agrobacterium, Stenotrophomonas, Aminobacter, etc. (Table S1). Consequently, it was suspected that EFBs may be present within the fruiting body of H. bachu.
3.2. QPCR Analyses Confirmed the Presence of EFBs in the Fruiting Body of Helvella bachu
Real-time quantitative PCR targeting the 16S rRNA gene was employed to detect bacteria in the clean tissues of the H. bachu fruiting body. The standard curve generated from the plasmid standard (Figure S1A) and the melting curve (Figure S1B) demonstrated the reliability of the method. The copy number of 16S rRNA in H. bachu tissue was quantified at 3.52 × 1014 copies/g, thereby confirming the presence of EFBs within the internal tissues of H. bachu.
3.3. Bacterial Abundance and Community Composition of Helvella bachu EFBs Associated with Three Types of Host Revealed by Full-Length 16S rRNA Gene Sequencing
3.3.1. Bacterial Abundance and Community Composition
Clean tissues of H. bachu fruiting body associated with three hosts, P. euphratica (FPE), P. pruinosa (FPP), and P. alba var. pyramidalis (FPA) (Figure 2A, Table 1), were collected for full-length 16S rRNA gene sequencing using PacBio technology. After denoising and removing chimeric sequences, a total of 430,853 reads were obtained, yielding an average of 47,872 reads per sample (Table S2).
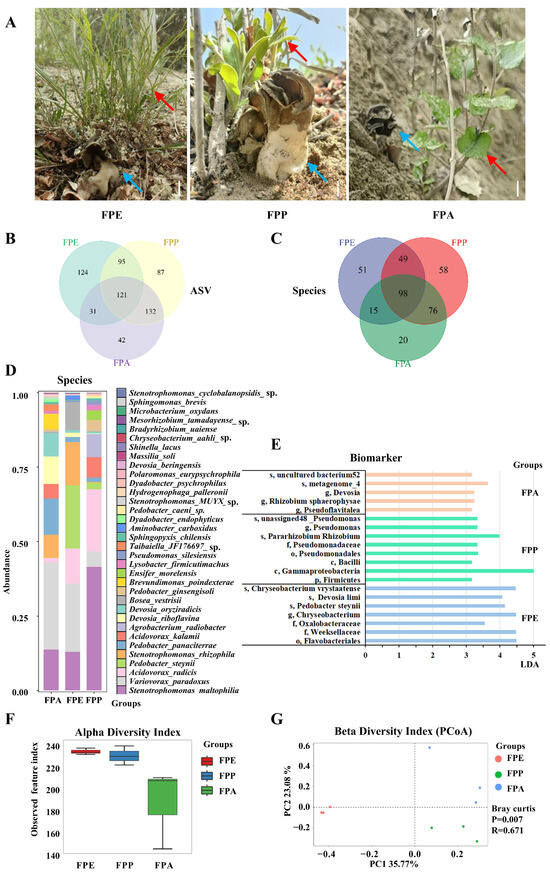
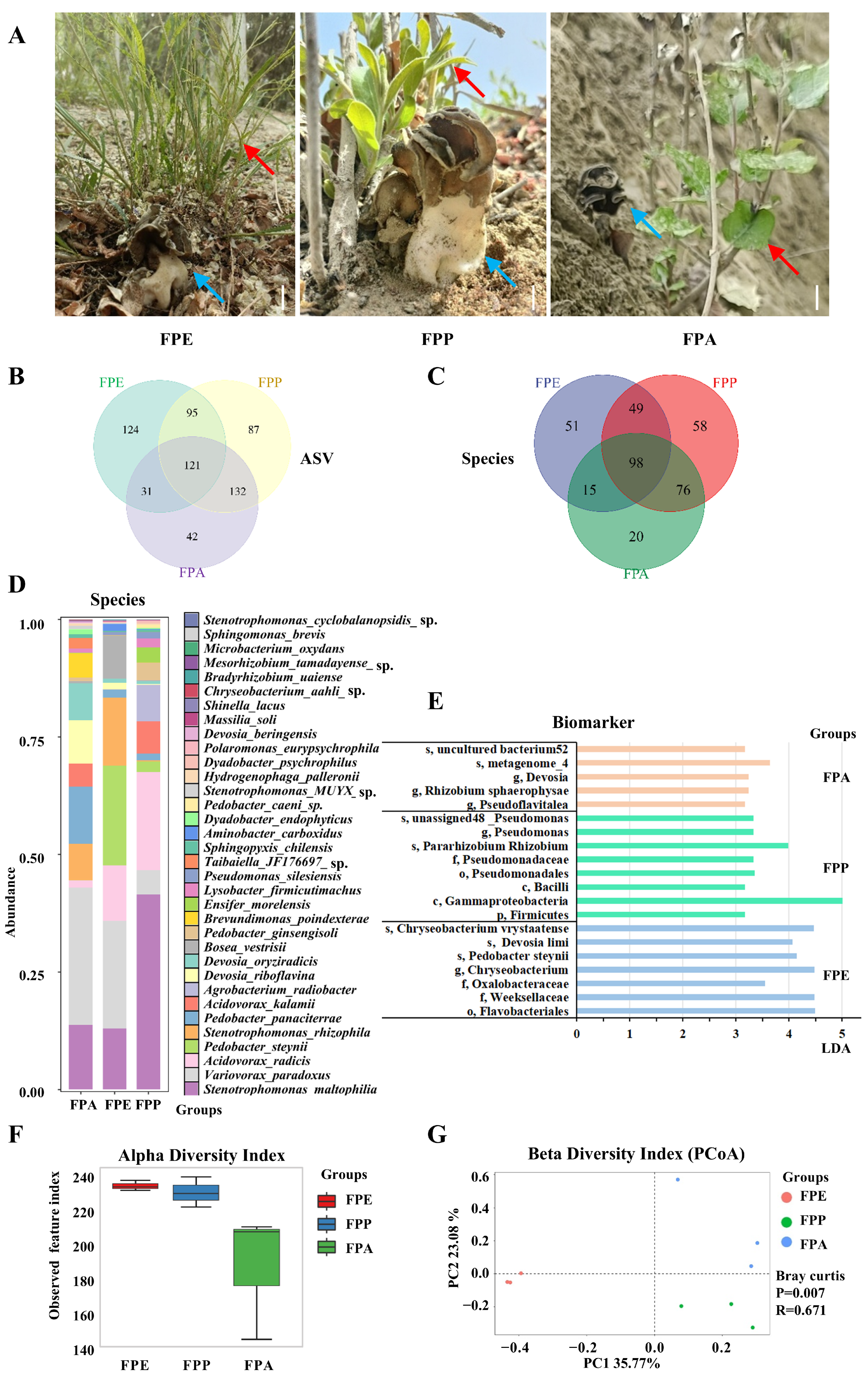
Figure 2.
Abundance and community composition of Helvella bachu EFBs associated with different hosts revealed by full-length 16S rRNA gene sequencing. (A) Fruiting bodies of H. bachu associated with different hosts. Blue arrow is H. bachu, and red arrow is host plant. Bar: 1.5 cm; (B) Venn diagram of amplicon sequence variant (ASV) of H. bachu EFBs associated with three host plants; (C) Venn diagram of species of H. bachu EFBs associated with three host plants; (D) common species of H. bachu EFBs associated with three host plants; (E) biomarkers of H. bachu EFBs associated with three host plants; (F) alpha diversity index; (G) beta diversity index (PCoA). FPE: H. bachu fruiting body associated with P. euphratica; FPP: H. bachu fruiting body associated with P. pruinosa; FPA: H. bachu fruiting body associated with P. alba var. Pyramidalis.
The dilution curve (Figure S2) indicated that the sequencing data for all samples stabilized and reached saturation, suggesting that the sample data were both abundant and uniform, with FPA being particularly distinctive. Furthermore, the curve showed that saturation was achieved when the sequencing data reached 40,000 CCS (Figure S2), confirming the adequacy of the sequencing data.
At the amplicon sequence variant (ASV) level, a total of 632 features were identified across the three kinds of sample associated with different hosts. Among these, 121 ASVs were shared among all three samples, while 124 were unique to FPE, 87 to FPP, and 42 to FPA. Additionally, 95 ASVs were shared between FPE and FPP, 31 between FPE and FPA, and 132 between FPP and FPA (Figure 2B). In terms of abundance, the distribution of ASVs among the samples follows this order: FPE > FPP > FPA.
Species composition and abundance at various taxonomic levels were assessed for each sample. In total, 17 phyla, 32 classes, 88 orders, 135 families, 259 genera, and 367 species of EFBs associated with fruiting bodies of H. bachu from the three host plants were identified (Table S3). Among these, Proteobacteria was the most abundant phylum found in the EFBs of all three host plants (Table S4), followed by Bacteroidota. Together, these two phyla accounted for over 98% of the total reads across all samples. Additionally, Planctomycetota, Verrucomicrobiota, Patescibacteria, Acidobacteriota, Myxococcota, Actinobacteriota, and Firmicutes were also detected in the EFBs of the three host plants, albeit in lower proportions.
A total of 24 genera were identified with an abundance greater than 0.01% of the total (Table S5). Notably, the genus Stenotrophomonas was abundant in the EFBs of all three host plants, accounting for over 20%. Other notable genera included Variovorax, Acidovorax, and Pedobacter, each contributing over 5% across all samples. There were some differences in abundance among the three host plants. The top three genera for FPE and FPP were Stenotrophomonas, Variovorax, and Pedobacter; however, the top three genera for FPA were Variovorax (29.26%), Stenotrophomonas (22.09%), and Devosia (17.49%).
A total of 367 species of EFBs were identified across three types of samples associated with different host plants. Among these, 98 species were shared among all three samples, while 51 species were unique to FPE, 58 to FPP, and 20 to FPA (Figure 2C). Additionally, 147 species were shared between FPE and FPP, 113 species between FPE and FPA, and 174 species between FPP and FPA. In terms of abundance, the distribution of species among the samples followed this order: FPP > FPE > FPA.
The common species shared among the three hosts included 34 species with an abundance greater than 0.01% of the total (Figure 2D and Table S6). However, there were great differences among the three hosts. For FPE, six species had an abundance over 5%, listed in the following order: Variovorax paradoxus (29.26%), Stenotrophomonas maltophilia (13.72%), Pedobacter panaciterrae (12.25%), Devosia riboflavina (9.26%), Devosia oryziradicis (7.87%), Stenotrophomonas rhizophila (7.65%), and Brevundimonas poindexterae (5.21%). For FPP, six species also exceeded 5% abundance, listed as follows: V. paradoxus (22.91%), Pedobacter steynii (21.15%), S. rhizophila (14.47%), S. maltophilia (12.98%), Acidovorax radicis (11.81%), and Bosea vestrisii (9.19%). In the case of FPA, the leading species was S. maltophilia (41.48%), followed by A. radicis (20.89%) and Agrobacterium radiobacter (7.77%). Other notable species included Acidovorax kalamii (6.85%) and V. paradoxus (5.14%). In general, the species S. maltophilia and V. paradoxus were abundant in the fruiting bodies of H. bachu associated with all three hosts.
3.3.2. Biomarkers for EFBs Associated with Various Host Plants
Biomarkers for various host plants were identified based on a linear discriminant analysis (LDA) value greater than 3 (Table S7, Figure 2E). For FPE, the identified biomarker included Flavobacteriales, Weeksellaceae, and Oxalobacteraceae, along with specific species such as P. steynii, Devosia limi, and Chryseobacterium vrystaatense. In the case of FPP, key biomarker included Firmicutes, Gammaproteobacteria, and various members of Pseudomonas, such as Pararhizobium, Rhizobium, and several unclassified Pseudomonas species. FPA was characterized by the presence of Devosia, Pseudoflavitalea, and Rhizobium sphaerophysae, along with various uncultured bacteria.
3.3.3. Diversity for EFBs in Helvella bachu Fruiting Bodies Associated with Various Host Plants
Various diversity indices, including the observed species index, Shannon index, Simpson index, Goods coverage index, Chao1 index, ACE index, and Pielou index, were calculated based on the abundance of ASVs (Table S8). The alpha diversity index for FPE was the highest, followed by FPP, with no significant difference observed between the two groups (Figure 2F). In contrast, the α diversity index for FPA was the lowest, significantly lower than that of both FPE and FPP (p < 0.05).
The beta diversity of EFBs associated with three different host plants was analyzed using Principal Coordinates Analysis (PCoA). The percentage of variability explained by each principal component was as follows: PC1 accounted for 35.77%, while PC2 accounted for 23.08%. Ordinations based on the Bray–Curtis metric showed a clear separation among the EFBs associated with the three host plants (Figure 2G). Additionally, the bacterial communities in the FPA samples were more similar to those found in the FPP samples.
Analysis of similarities (ANOSIM) also revealed differences among the groups (Table S9). However, the p-values for pairwise comparisons between the three host plants were greater than 0.05, indicating that these differences were not statistically significant. The R values for FPE, FPP, and FPA were 0.8519, 0.7407, and 0.4074, respectively. All R values fell within the range of 0 to 1, suggesting that the diversity of EFBs in different host plant groups was greater than that found within each individual host plant group.
3.3.4. Functional Prediction and Functional Diversity Analysis
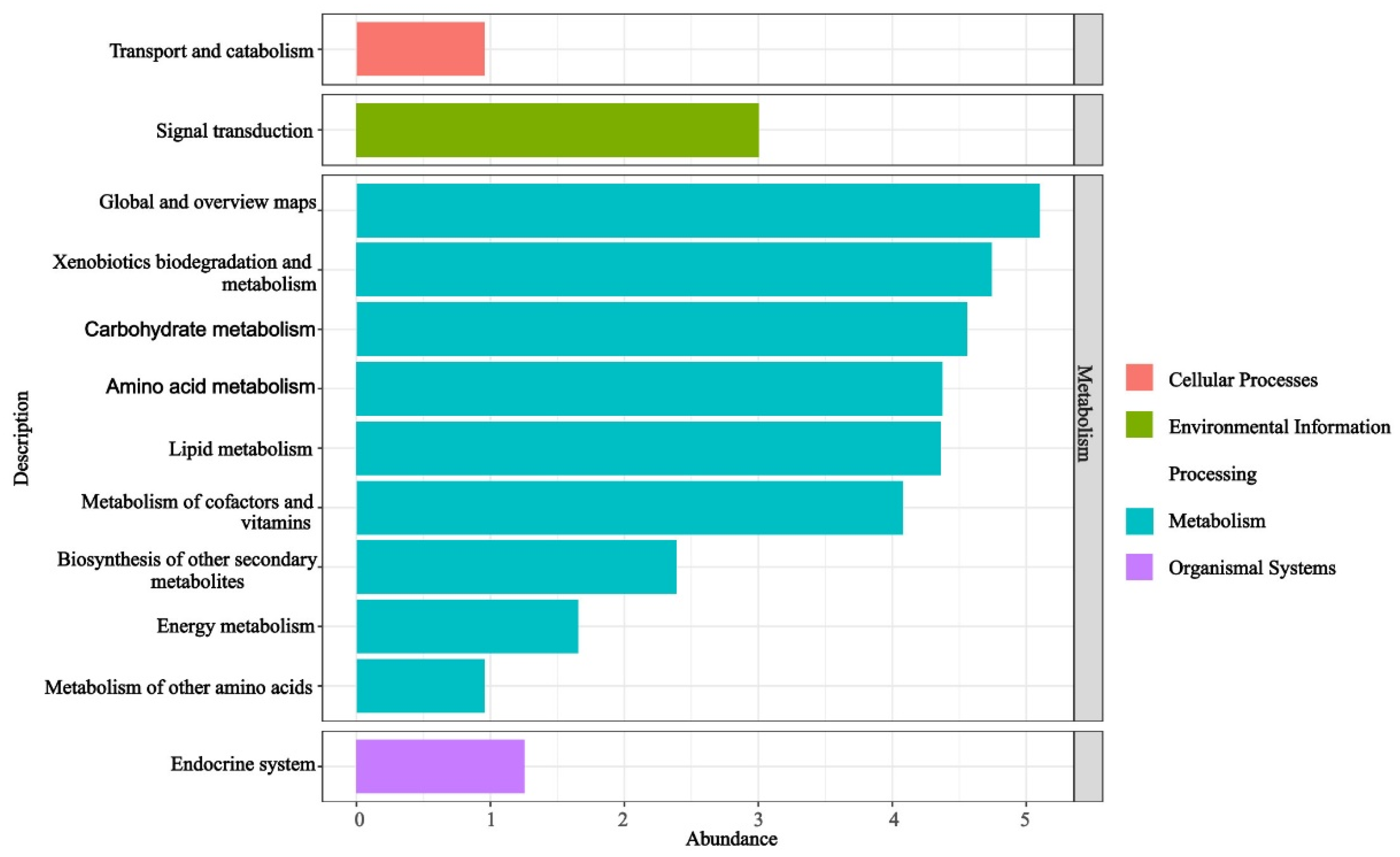
The functional KO-pathway analysis of the predicted KO results for the EFBs of H. bachu across all samples revealed that the enriched pathways were primarily associated with metabolism (Figure 3). Ranked by abundance from largest to smallest, these pathways included global and overview maps, xenobiotics biodegradation and metabolism, carbohydrate metabolism, amino acid metabolism, lipid metabolism, metabolism of cofactors and vitamins, biosynthesis of other secondary metabolites, energy metabolism, and metabolism of other amino acids. Additionally, pathways related to signal transduction within environmental information processing, endocrine systems of organismal systems, and transport and catabolism within cellular processes were also found to be enriched (Figure 3).
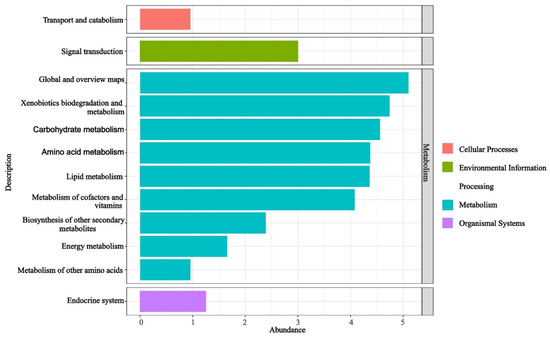
Figure 3.
Functional pathways based on KO abundance of EFBs associated with three hosts.
3.4. EFBs of Helvella bachu Fruiting Bodies at Different Developmental Stages
To investigate the dynamic changes in EFBs during the development of H. bachu fruiting bodies, samples from various developmental stages, including nascent (less than 3 cm in height, NF), developing (3–5 cm in height, DF), and mature (containing mature ascospores, MF) fruiting bodies (Figure 4A) were collected. These samples were analyzed using next-generation high-throughput sequencing of the 16S rRNA gene.
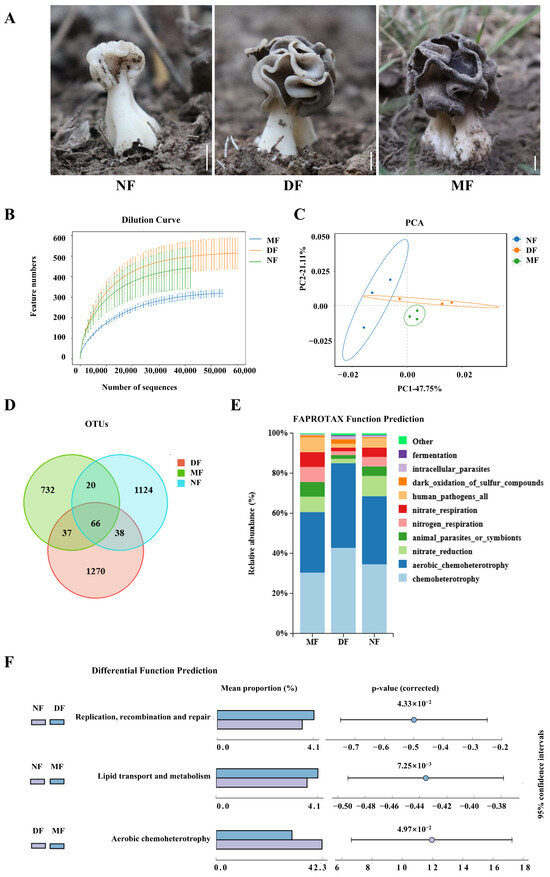
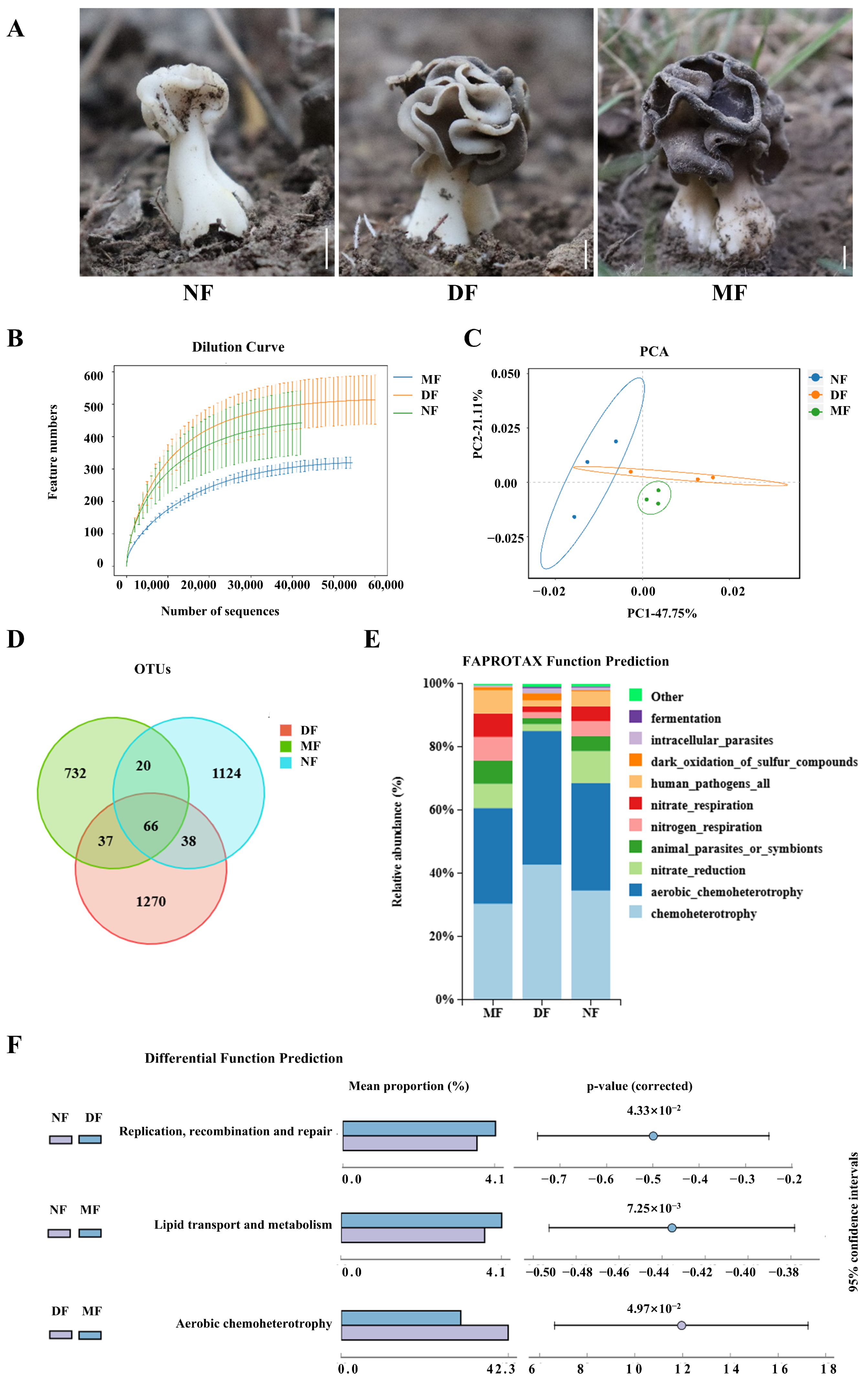
Figure 4.
EFBs of Helvella bachu fruiting bodies at different developmental stages revealed by next-generation sequencing of 16S rRNA gene. (A) H. bachu fruiting body samples at different stages. Bar: 1cm; (B) dilution curve; (C) beta diversity index (PCA); (D) Venn diagram of OTUs of H. bachu EFBs at different stages; (E) ecological functions of EFBs of H. bachu at different stages; (F) differential analyses of functions of EFBs of H. bachu through pairwise comparisons at different stages. NF: nascent fruiting bodies (less than 3 cm in height); DF: developing fruiting bodies (3–5 cm in height); MF: mature (containing mature ascospores) fruiting bodies.
The dilution curves showed that all final profiles flattened, signifying that the sequencing data quantity was adequate (Figure 4B). The EFBs of H. bachu at various stages can be distinguished through PCA analysis following dimensionality reduction, accounting for 47.75% and 21.11% of the variance, respectively (Figure 4C). The bacterial operational taxonomic units (OTUs) were determined to be 1351, 1542, and 961 in the NF, DF, and MF, respectively (Table S10). Only 66 OTUs were shared among all three stages (Figure 4D). Additionally, 104 OTUs were shared between the NF and DF samples, while 86 were shared between the NF and MF stages, and 103 were shared between the DF and MF. The number of unique OTUs at each stage was significantly greater than the number of common OTUs.
The EFBs across the three developmental stages included 30 phyla, 73 classes, 187 orders, 336 families, and 649 genera (Table S11). At the phylum level, EFBs showed consistency across different stages, predominantly composed of Proteobacteria and Bacteroidetes, which together accounted for over 95% of the total. There was also a minor presence of Firmicutes and Actinobacteriota (Table S12).
At the genus level, Pedobacter and Acidovorax were the most abundant genera shared among the three stages (Table S13). At the NF stage, the abundances were as follows: Stenotrophomonas (15.36%), Pedobacter (15.04%), Devosia (14.32%), and Acidovorax (11.04%). At the DF stage, Chryseobacterium was the most abundant at 37.58%, followed by Pedobacter (18.93%), Acidovorax (6.00%), and an unclassified member of Xanthobacteraceae at 5.06%. At the MF stage, Chryseobacterium represented 27.91%, Pedobacter 22.49%, Stenotrophomonas 21.10%, Acidovorax 6.50%, and Variovorax 4.98%.
The ecological functions of the EFBs at three different developmental stages are illustrated for the top ten in functional abundance (Figure 4E). In general, consistency was observed across all three stages. Six ecological functions of the EFBs were highlighted across the samples, including chemoheterotrophy, aerobic chemoheterotrophy, nitrate reduction, and functions associated with animal parasitism or symbionts, nitrogen respiration, nitrate respiration, as well as those related to human pathogenic bacteria.
Differential analyses of functions of EFBs of H. bachu through pairwise comparisons at different developmental stages revealed that DNA replication, recombination, and repair at the NF stage were significantly more pronounced than at the DF stage (Figure 4F). Additionally, lipid transcription and metabolism in the NF stage were significantly higher than in the MF stage. In terms of aerobic chemoheterotrophy, the DF stage exhibited significantly greater levels than the MF.
3.5. EFBs from Helvella bachu Fruiting Bodies Using Culture Methods
The absence of bacterial colonies on the control plates indicated that surface contaminants were effectively removed, and the isolates were obtained from within the fungal tissue. A total of 46 bacterial strains were isolated and identified (Table S14), belonging to four genera: Pseudomonas, Stenotrophomonas, Chryseobacterium, and Variovorax. Among these forty-six strains, thirty-four were classified under the genus Pseudomonas (73.91%), including seven strains of Pseudomonas fluorescens, four strains of Pseudomonas gessardii, three strains of Pseudomonas azotoformans, three strains of Pseudomonas poae, one strain of Pseudomonas putida, and one strain of Pseudomonas simiae. There were five strains of Stenotrophomonas (10.87%), comprising three strains of S. maltophilia, one strain of S. rhizophila, and one strain of Stenotrophomonas sp. Additionally, three strains of Chryseobacterium (6.52%), two strains of Microbacterium, one strain of Serratia sp., and one strain of Variovorax boronicumulans were identified.
4. Discussion
EFBs have emerged as a significant area of research within mycology, particularly for their roles in ecology, metabolism, and interactions with host fungi. H. bachu is a valuable ectomycorrhizal fungus associated with P. euphratica, a rare, ancient, and endangered species in desert riparian ecosystems. Despite numerous efforts, pure mycelial cultures have remained elusive, and attempts to cultivate its fruiting bodies have been unsuccessful for many years. This study confirmed the presence of EFBs within the fruiting bodies of H. bachu by a polyphasic approach, including genomic sequencing analysis, real-time quantitative PCR targeting the 16S rRNA gene, full-length sequencing and NGS of the 16S rRNA gene, as well as the culture methods. The genera Stenotrophomonas, Variovorax, Acidovorax, and Pedobacter were found to be abundant in H. bachu fruiting bodies associated with all three host plants, and these genera were also prevalent across different developmental stages. Notably, S. maltophilia and V. paradoxus were consistently detected by full-length 16S rRNA sequencing, and S. maltophilia was also isolated through culture methods. Functional KO-pathway analysis of the predicted KO results for EFBs in H. bachu revealed that metabolism-related pathways were predominantly enriched. This study not only confirmed the presence of EFBs in H. bachu, but also provided a comprehensive overview of their structure, functional potential, and dynamic changes throughout the maturation of H. bachu fruiting bodies, offering valuable insights that may facilitate the artificial domestication of this species.
4.1. EFBs Are Ubiquitously Present in Mushrooms, Especially in Ectomycorrhizal Mushrooms
EFBs resided within the vegetative or reproductive structures of fungi, with their presence documented in various species. Notable examples included saprophytic mushrooms such as A. bisporus, Lentinus edodes, Pleurotus cornucopiae [33], and Sang Huang [34], among others. However, EFBs are more commonly found in ectomycorrhizal fungi. These included Ascomycota species such as some species of Morchella [35,36] and Tuber [37,38,39], as well as Basidiomycota species, including Tylopilus felleus, T. areolatus, Boletus queletii, Phlebopus portentosus [19,40] Tricholoma bicolor [41], Russula griseocarnosa [42], Amanita pantherina, Suillus placidus, T. felleus, Agaricus flocculosipes, Chlorophyllum molybdites [43], and F. luteovirens [15]. In the present study, EFBs were confirmed in the fruiting bodies of H. bachu by a combination of methods, including genomic sequencing analysis, real-time quantitative PCR targeting the 16S rRNA gene, full-length sequencing and NGS of the 16S rRNA gene, as well as cultural methods. EFBs are ubiquitously present in mushrooms, particularly in ectomycorrhizal mushrooms.
4.2. Species Composition and Abundance of EFBs in Helvella bachu
Proteobacteria and Bacteroidetes were consistently identified as the predominant phyla in the EFBs of fruiting bodies associated with all three host plants (Table S4) and across the three different stages (Table S12). Together, these two phyla accounted for more than 95% of the total reads across all samples. It was reported that the relative abundance of Proteobacteria, Actinobacteria and Bacteroidetes were dominant in EFBs of Cantharellales and Hydnum [44].
At the genus level, some differences in abundance were observed among the EFBs in fruiting bodies associated with the three host plants (Table S5). However, the genus Stenotrophomonas remained consistently abundant across all three host plants, accounting for over 20% of the total. Other notable genera, including Variovorax, Acidovorax, and Pedobacter, each contributed over 5% across all samples. These genera were also detected in the fruiting bodies at different stages, with Pedobacter and Acidovorax being the most abundant genera shared across all three stages. Different fungal groups harbor specific EFB communities. In ascomycetous mushrooms, these included Bacillus, Mesorhizobium, Variovorax, Burkholderia, Pseudoma, Sphingobacteriaceae, Flavobacteriaceae, Ensifer, Rhizobium, and Rhodococcus [38,45]. In ectomycorrhizal fungi, Pedobacter, Pseudomonas, Variovorax, Chitinophaga, Ewingella, and Stenotrophomonas were the dominant genera [22,46].
Full-length 16S rRNA gene sequencing, utilizing third-generation sequencing platforms such as PacBio SMRT, increases read lengths and confers species-level resolution [47,48]. In the present study, EFBs in the fruiting bodies associated with three hosts were identified by full-length 16S rRNA gene sequencing. Despite great differences among the three hosts, the species S. maltophilia and V. paradoxus were consistently abundant in the fruiting bodies of H. bachu associated with all three hosts. Other species, including A. kalamii, A. radiobacter, B. poindexterae, B. vestrisii, D. oryziradicis, D. riboflavina, P. panaciterrae, P. steynii, and S. rhizophila, were also identified, with varying abundances across the three hosts (Table S6). Especially, S. maltophilia and S. rhizophila were also isolated through culture methods (Table S14).
4.3. Potential Functions of EFBs in Helvella bachu
Endosymbioses have profoundly influenced the evolution of life and continue to shape the ecology of numerous species [10]. The widespread presence of EFBs in ectomycorrhizal mushrooms highlights their diverse and significant ecological roles. EFBs contribute to fungal health by serving as a bioenergy source, producing essential compounds like sugars, proteins, vitamins, and auxins [16,17,18], facilitating nutrient transport, enhancing respiration and environmental adaptability [19,20,21], acting as biological control agents against fungal diseases [11], and impacting the development, maturation, and aroma of mushrooms [22,23].
In this study, the functional KO-pathway analysis of the EFBs in the fruiting bodies associated with three host plants revealed that the pathways were predominantly enriched in metabolic processes, including primary, secondary, and energy metabolism (Figure 3). Primary metabolic pathways encompassed carbohydrate metabolism, amino acid metabolism, lipid metabolism, metabolism of cofactors and vitamins, and metabolism of other amino acids (Figure 3). These findings suggested that the EFBs may contribute to the growth of H. bachu by producing essential compounds such as sugars, proteins, and vitamins. Additionally, secondary metabolism and energy metabolism pathways were also identified as being significant.
The ecological function analysis of EFBs in H. bachu fruiting bodies at different developmental stages by APROTAX revealed that nitrogen respiration, nitrate reduction, and nitrate respiration were prominent across all developmental stages. This suggested that EFBs play a significant role in nitrogen cycling, providing nutrients for the host H. bachu. Other studies also have demonstrated that EFBs are crucial for carbon cycling [49], lipid metabolism [50], and nitrogen complex [51].
The species S. maltophilia and V. paradoxus were consistently the most abundant in the fruiting bodies of H. bachu associated with all three host plants. S. maltophilia has been reported to have several roles, including: (1) biological nitrogen fixation and secretion of plant hormones to promote plant growth [52]; (2) production of antibiotics and various enzymes to control pathogenic microorganisms and support plant growth [53,54]; and (3) enhancement of saline–alkaline resistance [55]. A. radicis has been shown to stimulate root growth and induce systemic resistance in plants [56], thereby benefiting agriculture by promoting plant growth and providing pest suppression under changing climatic conditions [57]. Future studies will further explore the functions of S. maltophilia and V. paradoxus in these aspects.
The dynamic changes of EFBs across the three developmental stages revealed higher nitrogen fixation in the NF and DF stages compared to the MF stage. In contrast, aerobic chemoheterotrophs were more prevalent in the MF stage than in the earlier two stages. Similarly, the Allorhizobium–Neorhizobium–Pararhizobium–Rhizobium complex plays a key role in nitrogen fixation, with this function diminishing as the fruiting bodies mature in C. cibarius [14]. These intricate interactions offer valuable insights for improving mushroom cultivation practices.
Lastly, species-specific symbiotic associations, such as those between Lactarius deliciosus and pine trees, or Suillus luteus and pine trees [58], are rare, typically occurring within certain taxa, including Boletales, Russulales, and Agaricales. Actually, some species of Helvella form associations with a broader range of host types, including conifers, Castanopsis, Fagus, Populus, and several species of Quercus [59]. While H. bachu may also establish symbiotic relationships with tree species other than Populus in the wild, such associations remain undocumented due to insufficient research and data. Future studies will be valuable in exploring both the host species and EFBs associated with H. bachu.
5. Conclusions
A polyphasic approach confirmed the presence of EFBs in the fruiting bodies of H. bachu and provided the first comprehensive overview of their structure, functional potential, and dynamic changes throughout fruiting body maturation. Future experiments could further validate the functions of these EFBs, which could be utilized to obtain pure cultured mycelia and incorporated into mycorrhizal synthesis for artificial propagation.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof10120889/s1, Figure S1: Real-time quantitative PCR based on 16S rRNA gene; Figure S2: Dilution curve for data of full-length 16S rRNA gene sequencing; Table S1: Possible EFBs identified during genome sequencing analysis; Table S2: Data statistics of EFBs in Helvella bachu fruiting body associated with three hosts by full-length 16S rRNA gene sequencing; Table S3: Statistical data of EFBs of Helvella bachu under different hosts; Table S4: Reads composition of the EFBs of Helvella bachu under different hosts at phylum level; Table S5: Common genus of EFBs in Helvella bachu associated with different hosts; Table S6: Common species of EFBs in Helvella bachu associated with different hosts; Table S7: Biomarkers of EFBs of Helvella bachu under three host plants; Table S8: Alpha diversity index of EFBs in Helvella bachu fruiting bodies associated with various host plants; Table S9: ANOSIM intergroup difference analysis of EFBs in Helvella bachu fruiting bodies associated with various host plants; Table S10: OTUs of EFBs in Helvella bachu at different developmental stages; Table S11: EFBs of Helvella bachu at different taxonomic levels at different developmental stages; Table S12: EFBs of Helvella bachu at different developmental stages at the phylum level; Table S13: Shared genus of EFBs in Helvella bachu at different developmental stages; Table S14: EFBs of Helvella bachu obtained by culture methods.
Author Contributions
Conceptualization, C.W., L.Z. and C.D.; methodology, C.W., X.Z. and J.X.; software, M.W. and G.M.; validation, C.W. and C.D.; formal analysis, C.W.; investigation, C.W. and J.H.; resources, C.W. and J.H.; data curation, C.W., M.L., G.M. and M.W.; writing—original draft preparation, C.W.; writing—review and editing, C.W., L.Z. and C.D.; visualization, C.W. and M.L.; supervision, L.Z. and C.D.; project administration, C.W., L.Z. and C.D.; funding acquisition, C.W., L.Z. and C.D. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Third Xinjiang Scientific Expedition Program under the project “Survey of the Components of Desert Ecosystems and Genetic Resources of Unique Organisms in Extreme Environments” (grant number 2022xjkk0200), and by the Scientific Research and Innovation Project for Postgraduates at Tarim University under the project “Microbiomic Study on the Fruiting Body Development of Bachu Mushroom Based on Metagenomics” (grant number TDBSCX202110).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The original contributions presented in this study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Acknowledgments
Special gratitude is extended to Zhanfeng Xia and Xiaoxia Luo from Tarim University for providing the experimental platform; to Ying Yang, a researcher at the Chinese Academy of Sciences, for revising this paper; and to Jiaxin Kong and Bingbing Feng, students from Tarim University, for their support in conducting part of the experiments.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Zhao, Q.; Sulayman, M.; Zhu, X.T.; Zhao, Y.C.; Yang, Z.L.; Hyde, K.D. Species clarification of the culinary Helvella bachu in western China. Mycologia 2016, 108, 828–836. [Google Scholar] [CrossRef] [PubMed]
- Badshah, H.; Ali, B.; Khan, S.M.; Chaudhari, M.I.; Nisa, S.U.; Badshah, L.; Mumtaz, A.S. The in vitro antibacterial and antioxidant potentials of Helvetia bachu (Ascomycetes): A newly recorded species from Pakistan. Int. J. Med. Mushrooms 2020, 22, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Al-Rawi, J.M.; Abdul-Hadi, S.Y. First new record and molecular identification of H. bachu (Ascomycetes) isolated from Iraq/Mosul. Int. J. Res. Appl. Sci. Biotechnol. 2022, 9, 143–148. [Google Scholar]
- Xu, J.Y.; Hu, G.H.; Gao, X.H. Resources and research of wild Helvella leucopus in Xinjiang. Chin. Wild Pl. Resour. 2004, 22, 25–26. (In Chinese) [Google Scholar]
- Meng, Q.L.; Zhang, P.P.; Hu, J.W. Effects of immunoregulatory activity of polysaccharide from Bachu-mushroom. China Poult. 2005, 9, 118–120. (In Chinese) [Google Scholar]
- Zeng, D.; Zhu, S.M. Purification, characterization, antioxidant and anticancer activities of novel polysaccharides extracted from Helvella bachu. Int. J. Biol. Macromol. 2018, 107, 1086–1092. [Google Scholar] [CrossRef]
- Wei, C.H.; Liu, M.Q.; Hu, J.W.; Zhang, L.L.; Dong, C.H. Mycorrhizal associations between Helvella bachu and its host plants. Forests 2024, 15, 721. [Google Scholar] [CrossRef]
- Alabid, I.; Glaeser, S.P.; Kogel, K.H. Endofungal bacteria increase fitness of their host fungi and impact their association with crop plants. Curr. Issues Mol. Biol. 2019, 30, 59–74. [Google Scholar] [CrossRef]
- Mosse, B. Honey-coloured, sessile Endogone spores: II. Changes in fine structure during spore development. Archiv. Mikrobiol. 1970, 74, 129–145. [Google Scholar] [CrossRef]
- Giger, G.H.; Ernst, C.; Richter, I.; Gassler, T.; Field, C.M.; Sintsova, A.; Kiefer, P.; Gäbelein, C.G.; Guillaume-Gentil, O.; Scherlach, K.; et al. Inducing novel endosymbioses by implanting bacteria in fungi. Nature 2024, 635, 415–422. [Google Scholar] [CrossRef]
- Aslani, M.A.; Harighi, B.; Abdollahzadeh, J. Screening of endofungal bacteria isolated from wild growing mushrooms as potential biological control agents against brown blotch and internal stipe necrosis diseases of Agaricus bisporus. Biol. Control 2018, 119, 20–26. [Google Scholar] [CrossRef]
- Zhang, A.Y.; Zhang, M.L.; Zhu, J.L.; Yan, M.; Xu, F.J.; Bai, H.Y.; Sun, K.; Zhang, W.; Dai, C.C.; Jia, Y. Endofungal bacterial microbiota promotes the absorption of chelated inorganic phosphorus by host pine through the ectomycorrhizal system. Microbiol. Spectr. 2023, 11, e00162-23. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Perez-Moreno, J.; Zhang, P.; Wang, R.; Chater, C.C.C.; Yu, F.Q. Distinct compartmentalization of microbial community and potential metabolic function in the fruiting body of Tricholoma matsutake. J. Fungi 2021, 7, 586. [Google Scholar] [CrossRef] [PubMed]
- Ge, W.; Zhang, Z.Y.; Dong, C.B.; Shao, Q.Y.; Liu, Y.X.; Han, Y.F.; Liang, Z.Q. Diversity and functional analysis of the culturable microbes isolated from the fruiting bodies of wild Cantharellus cibarius. Mycosystema 2021, 40, 1054–1073. [Google Scholar]
- Feng, Z.L.; Li, N.; Deng, Y.F.; Yu, Y.; Gao, Q.B.; Wang, J.L.; Chen, S.L.; Xing, R. Biogeography and assembly processes of abundant and rare soil microbial taxa in the southern part of the Qilian Mountain National Park, China. Ecol. Evol. 2024, 14, e11001. [Google Scholar] [CrossRef]
- Schüßler, A. 5 The Geosiphon-Nostoc endosymbiosis and its role as a model for arbuscular mycorrhiza research. In Fungal Associations, 2nd ed.; Hock, B., Ed.; The Mycota; Springer: Berlin, Germany, 2012; Volume 9, pp. 77–91. [Google Scholar]
- Salvioli, A.; Ghignone, S.; Novero, M.; Navazio, L.; Venice, F.; Bagnaresi, P.; Bonfante, P. Symbiosis with an endobacterium increases the fitness of a mycorrhizal fungus, raising its bioenergetic potential. ISME J. 2016, 10, 130–144. [Google Scholar] [CrossRef]
- Bonfante, P.; Desirò, A. Who lives in a fungus? The diversity, origins and functions of fungal endobacteria living in Mucoromycota. ISME J. 2017, 11, 1727–1735. [Google Scholar] [CrossRef]
- Bai, H.Y. Diversity and Ecological Function of Endophytic Bacteria in Boletus Fruiting Bodie. Master’s Thesis, Nanjing Normal University, Nanjing, China, 2021. [Google Scholar]
- Cruz, A.F.; Horii, S.; Ochiai, S.; Yasuda, A.; Ishii, T. Isolation and analysis of bacteria associated with spores of Gigaspora margarita. J. Appl. Microbiol. 2008, 104, 1711–1717. [Google Scholar] [CrossRef]
- Guo, H.; Glaeser, S.P.; Alabid, I.; Imani, J.; Haghighi, H.; Kämpfer, P.; Kogel, K.H. The abundance of endofungal bacterium Rhizobium radiobacter (syn. Agrobacterium tumefaciens) increases in its fungal host Piriformospora indica during the tripartite sebacinalean symbiosis with higher plants. Front. Microbiol. 2017, 8, 629. [Google Scholar] [CrossRef]
- Kumari, D.; Reddy, M.S.; Upadhyay, R.C. Diversity of cultivable bacteria associated with fruiting bodies of wild Himalayan Cantharellus spp. Ann. Microbiol. 2013, 63, 845–853. [Google Scholar] [CrossRef]
- Antony-Babu, S.; Deveau, A.; Van Nostrand, J.D.; Zhou, J.; Le Tacon, F.; Robin, C.; Frey-Klett, P.; Uroz, S. Black truffle-associated bacterial communities during the development and maturation of Tuber. melanosporum ascocarps and putative functional roles. Environ. Microbiol. 2014, 16, 2831–2847. [Google Scholar] [CrossRef] [PubMed]
- Ling, H.; Zhang, P.; Xu, H.; Zhao, X.F. How to regenerate and protect desert riparian Populus euphratica forest in arid areas. Sci. Rep. 2015, 5, 15418. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Nomura, K.; Wang, X.; Sohrabi, R.; Xu, J.; Yao, L.; Paasch, B.C.; Ma, L.; Kremer, J.; Cheng, Y.; et al. A plant genetic network for preventing dysbiosis in the phyllosphere. Nature 2020, 580, 653–657. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Salzberg, S.L. Ultrafast and accurate 16S rRNA microbial community analysis using Kraken 2. Microbiome 2020, 8, 124. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Ding, C.; Adrian, L.; Peng, Y.Z.; He, J. 16S rRNA gene-based primer pair showed high specificity and quantification accuracy in detecting freshwater Brocadiales anammox bacteria. FEMS Microb. Ecol. 2020, 96, fiaa013. [Google Scholar] [CrossRef]
- Al’Khafaji, A.M.; Smith, J.T.; Garimella, K.V.; Babadi, M.; Popic, V.; Sade-Feldman, M.; Gatzen, M.; Sarkizova, S.; Schwartz, M.A.; Blaum, E.M.; et al. High-throughput RNA isoform sequencing using programmed cDNA concatenation. Nat. Biotechnol. 2024, 42, 582–586. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.R.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2 for prediction of metagenome functions. Nat. Biotechnol. 2020, 38, 685–688. [Google Scholar] [CrossRef]
- Pent, M.; Põldmaa, K.; Bahram, M. Bacterial communities in boreal forest mushrooms are shaped both by soil parameters and host identity. Front. Microbiol. 2017, 8, 836. [Google Scholar] [CrossRef]
- Zhang, J.L. Isolation and Identification of Mushroom Endophyte and the Preliminary of Their Metabolites. Master’s Thesis, Fujian Agriculture and Forestry University, Fujian, China, 2010. [Google Scholar]
- Ma, Y.J.; Gao, W.Q.; Zhu, X.T.; Kong, W.B.; Zhang, F.; Yang, H.Q. Identification and profiling of the community structure and potential function of bacteria from the fruiting bodies of Sanghuangporus vaninii. Arch. Microbiol. 2022, 204, 564. [Google Scholar] [CrossRef] [PubMed]
- Buscot, F. Ectomycorrhizal types and endobacteria associated with ectomycorrhizas of Morchella elata (Fr.) Boudier with Picea abies (L.) Karst. Mycorrhiza 1994, 4, 223–232. [Google Scholar] [CrossRef]
- Shen, H.; Chen, M.J.; Zhao, Y.C. PCR-DGGE identification of endophytes in Morels. Acta Agric. Shanghai 2008, 24, 58–60. [Google Scholar]
- Gazzanelli, G.; Malatesta, M.; Pianetti, A.; Baffone, W.; Stocchi, V.; Citterio, B. Bacteria associated to fruit bodies of the ecto-mycorrhizal fungus Tuber. borchii Vittad. Symbiosis 1999, 26, 211–222. [Google Scholar]
- Giorgio, M.; Niccolò, B.G.M.; Benedetta, T.; Luisa, M.; Leonardo, B.F.; Gregory, B.; Pietro, B.; Alberto, A.; Domizia, D.; Emidio, A. Fungal and bacterial diversity in the Tuber. magnatum ecosystem and microbiome. Microb. Ecol. 2023, 85, 508–521. [Google Scholar] [CrossRef]
- Martin, F.; Kohler, A.; Murat, C.; Balestrini, R.; Coutinho, P.M.; Jaillon, O.; Montanini, B.; Morin, E.; Noel, B.; Percudani, R.; et al. Pe’rigord black truffle genome uncovers evolutionary origins and mechanisms of symbiosis. Nature 2010, 464, 1033–1038. [Google Scholar] [CrossRef]
- Li, C.H.; Hou, D.; Li, Y.; Guo, T.; Ji, G.Y. Bacterial community structure in the fruiting–bodies of cultivated Phlebopus portentosus. J. Microbiol. 2024, 44, 68–76. [Google Scholar]
- Bertaux, J.; Schmid, M.; Prevost-Boure, N.C.; Churin, J.L.; Hartmann, A.; Garbaye, J.; Frey-Klett, P. In situ identification of intracellular bacteria related to Paenibacillus spp. in the mycelium of the ectomycorrhizal fungus Laccaria bicolor S238N. Appl. Environ. Microbiol. 2003, 69, 4243–4248. [Google Scholar] [CrossRef]
- Wu, X.X.; Liu, Y.C.; Wang, A.; Zhuo, L.J.; Hu, H.P. Isolation, identification and its antibacterial activity of an endophytic fungus of Russula griseocarnosa. Biotic Resources 2020, 42, 585–592. [Google Scholar]
- Liu, Y.P.; Sun, Q.B.; Li, J.; Lian, B. Bacterial diversity among the fruit bodies of ectomycorrhizal and saprophytic fungi and their corresponding hyphosphere soils. Sci. Rep. 2018, 8, 11672. [Google Scholar] [CrossRef]
- Gohar, D.; Pent, M.; Põldmaa, K.; Bahram, M. Bacterial community dynamics across developmental stages of fungal fruiting bodies. FEMS Microbiol. Ecol. 2020, 96, fiaa175. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Sun, X.; Liu, X.L.; Jia, B.S.; Liu, X.Y. Research progress on endofungal bacteria. Mycosystema 2019, 38, 1581–1599. [Google Scholar]
- Brabcová, V.; Nováková, M.; Davidová, A.; Baldrian, P. Dead fungal mycelium in forest soil represents a decomposition hotspot and a habitat for a specific microbial community. New Phytol. 2016, 210, 1369–1381. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y.; Komiya, S.; Yasumizu, Y.; Yasuoka, Y.; Mizushima, K.; Takagi, T.; Kryukov, K.; Fukuda, A.; Morimoto, Y.; Naito, Y.; et al. Full-length 16S rRNA gene amplicon analysis of human gut microbiota using MinION™ nanopore sequencing confers species-level resolution. BMC Microbiol. 2021, 21, 35. [Google Scholar] [CrossRef] [PubMed]
- Matsuo, Y. Full-length 16S rRNA gene analysis using long-read Nanopore sequencing for rapid identification of bacteria from clinical specimens. Methods Mol. Biol. 2023, 2632, 193–213. [Google Scholar]
- Rangel-Castro, J.I.; Danell, E.; Pfeffer, P.E. A 13C-NMR study of exudation and storage of carbohydrates and amino acids in the ectomycorrhizal edible mushroom Cantharellus cibarius. Mycologia 2002, 94, 190–199. [Google Scholar]
- Lazzaroni, S.; Bassarello, C.; Bifulco, G.; Lo Cantore, P.; Evidente, A.; Iacobellis, N.S.; Riccio, R.; Gomezpaloma, L. Chemical and biological characterization of Tolaasins A–E: New lipodepsipeptides produced by Pseudomona stolaasii. In Pseudomonas Syringae and Related Pathogens, 1st ed.; Iacobellis, N.S., Steven, A.C., Hutcheson, J.W., Mansfield, J.W., Morris, C.E., Murillo, J., Schaad, N.W., Stead, D.E., Surico, G., Ullrich, M.S., Eds.; Springer: Potenza, Italy, 2003; pp. 245–254. [Google Scholar]
- Riedlinger, J.; Schrey, S.D.; Tarkka, M.T.; Hampp, R.; Kapur, M.; Fiedler, H.P. Auxofuran, a novel metabolite that stimulates the growth of fly agaric, is produced by the mycorrhiza helper bacterium Streptomyces strain AcH 505. Appl. Environ. Microbiol. 2006, 72, 3550–3557. [Google Scholar] [CrossRef]
- Yang, N.; Røder, H.L.; Wicaksono, W.A.; Wassermann, B.; Russel, J.; Li, X.; Nesme, J.; Berg, G.; Sørensen, S.J.; Burmølle, M. Interspecific interactions facilitate keystone species in a multispecies biofilm that promotes plant growth. ISME J. 2024, 18, wrae012. [Google Scholar] [CrossRef]
- Liu, H.; Li, J.; Carvalhais, L.C.; Percy, C.D.; Prakash Verma, J.; Schenk, P.M.; Singh, B.K. Evidence for the plant recruitment of beneficial microbes to suppress soil-borne pathogens. New Phytol. 2021, 229, 2873–2885. [Google Scholar] [CrossRef]
- Raio, A.; Brilli, F.; Neri, L.; Baraldi, R.; Orlando, F.; Pugliesi, C.; Chen, X.; Baccelli, I. Stenotrophomonas rhizophila Ep2.2 inhibits growth of Botrytis cinerea through the emission of volatile organic compounds, restricts leaf infection and primes defense genes. Front. Plant Sci. 2023, 14, 1235669. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, J.; Wang, N.; Li, X.L.; Fang, N.; Zhuang, X.L. Diffusible signal factor enhances the saline-alkaline resistance and rhizosphere colonization of Stenotrophomonas rhizophila by coordinating optimal metabolism. Sci. Total Environ. 2022, 834, 155403. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Li, D.; Trost, E.; Mayer, K.F.; Vlot, A.C.; Heller, W.; Schmid, M.; Hartmann, A.; Rothballer, M. Systemic responses of barley to the 3-hydroxy-decanoyl-homoserine lactone producing plant beneficial endophyte Acidovorax radicis N35. Front. Plant Sci. 2016, 7, 1868. [Google Scholar] [CrossRef]
- Zytynska, S.E.; Eicher, M.; Rothballer, M.; Weisser, W.W. Microbial-mediated plant growth promotion and pest suppression varies underclimate change. Front. Plant Sci. 2020, 11, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Wang, Y.L.; Guerin-Laguette, A.; Zhang, P.; Colinas, C.; Yu, F.Q. Factors influencing successful establishment of exotic Pinus radiata seedlings with co-introduced Lactarius deliciosus or local ectomycorrhizal fungal communities. Front. Microbiol. 2022, 13, 973483. [Google Scholar] [CrossRef]
- Hwang, J.; Zhao, Q.; Yang, Z.L.; Wang, Z.; Townsend, J.P. Solving the ecological puzzle of mycorrhizal associations using data from annotated collections and environmental samples–an example of saddle fungi. Environ. Microbiol. Rep. 2015, 7, 658–667. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).