
Arbuscular Mycorrhizal Fungi Regulate Lipid and Amino Acid Metabolic Pathways to Promote the Growth of Poncirus trifoliata (L.) Raf

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.1.1. The Used Citrus Plant
2.1.2. The Used AM Fungi Isolate
2.1.3. The Used Soil and Substrates
2.2. Experimental Design
2.3. Experimental Methods
2.3.1. Determination of Plant Growth Indicators
2.3.2. Root Infection Determination
2.3.3. Sampling for Metabolomics Analysis
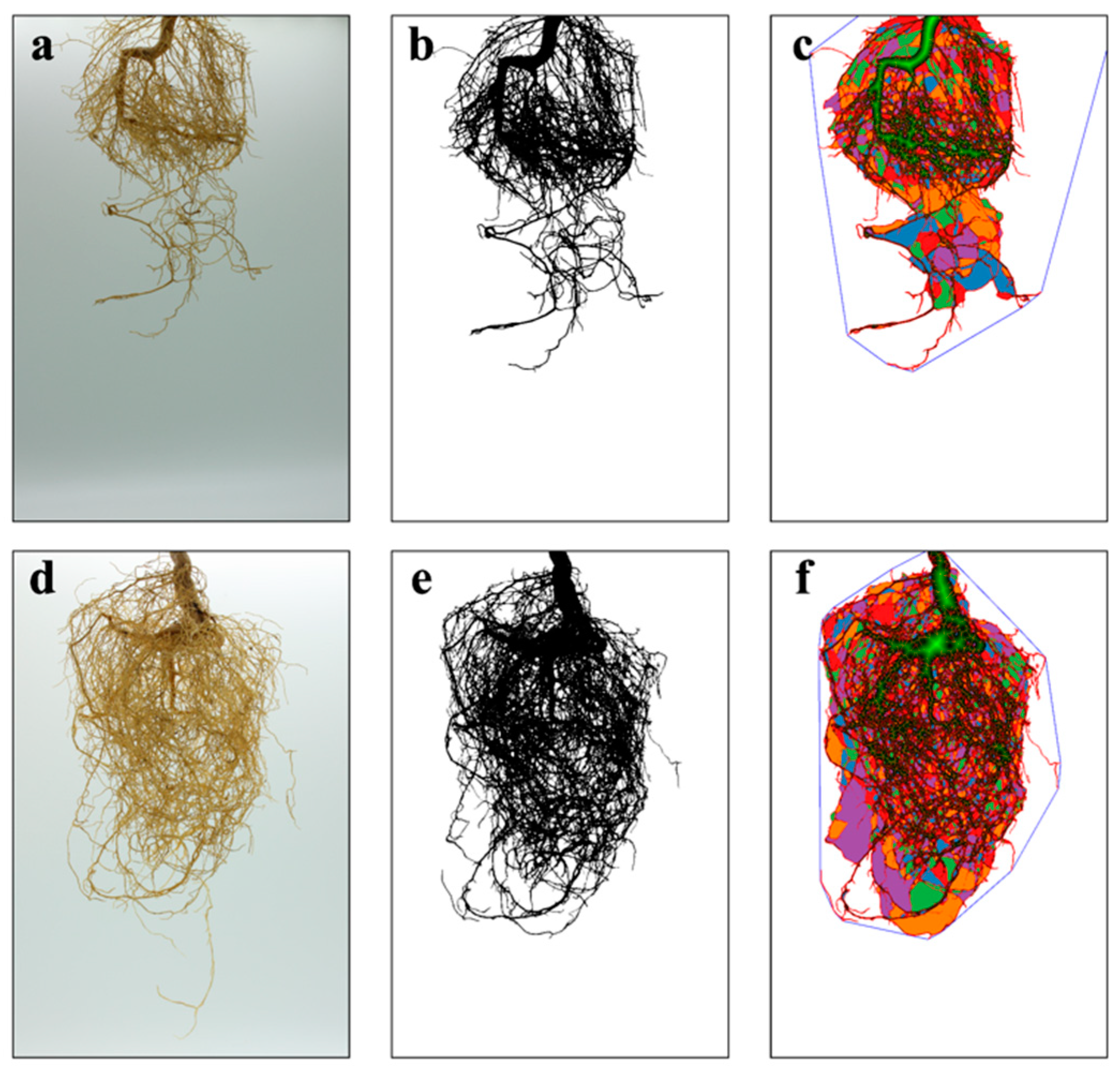
2.3.4. Root Scanning Analysis
2.3.5. Biomass Measurement
2.3.6. Data Analysis
3. Results
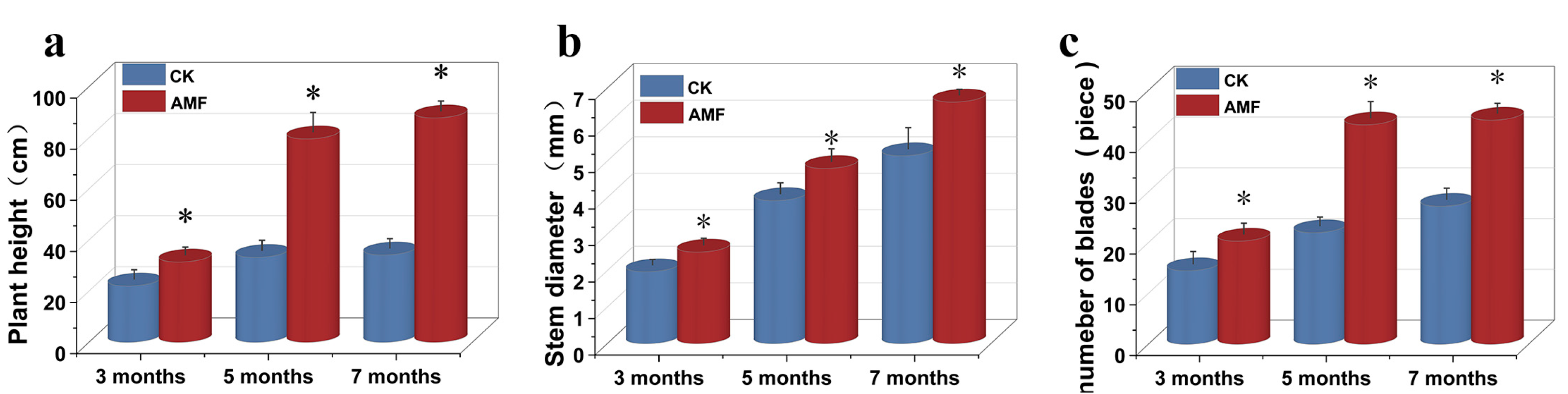
3.1. Physiological Indicators of P. trifoliata
3.2. Metabolomic Characteristics of P. trifoliata
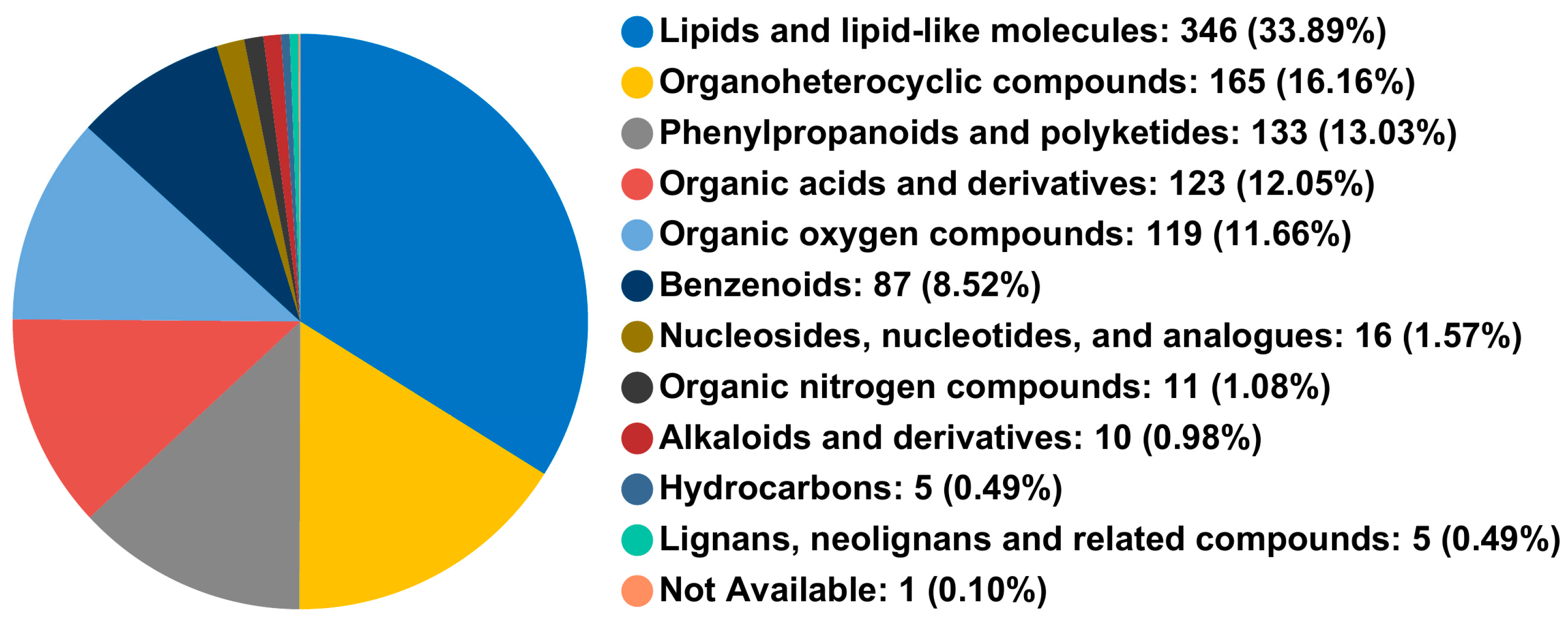
3.2.1. Metabolite Detection and Classification
3.2.2. Analysis of Differential Metabolites
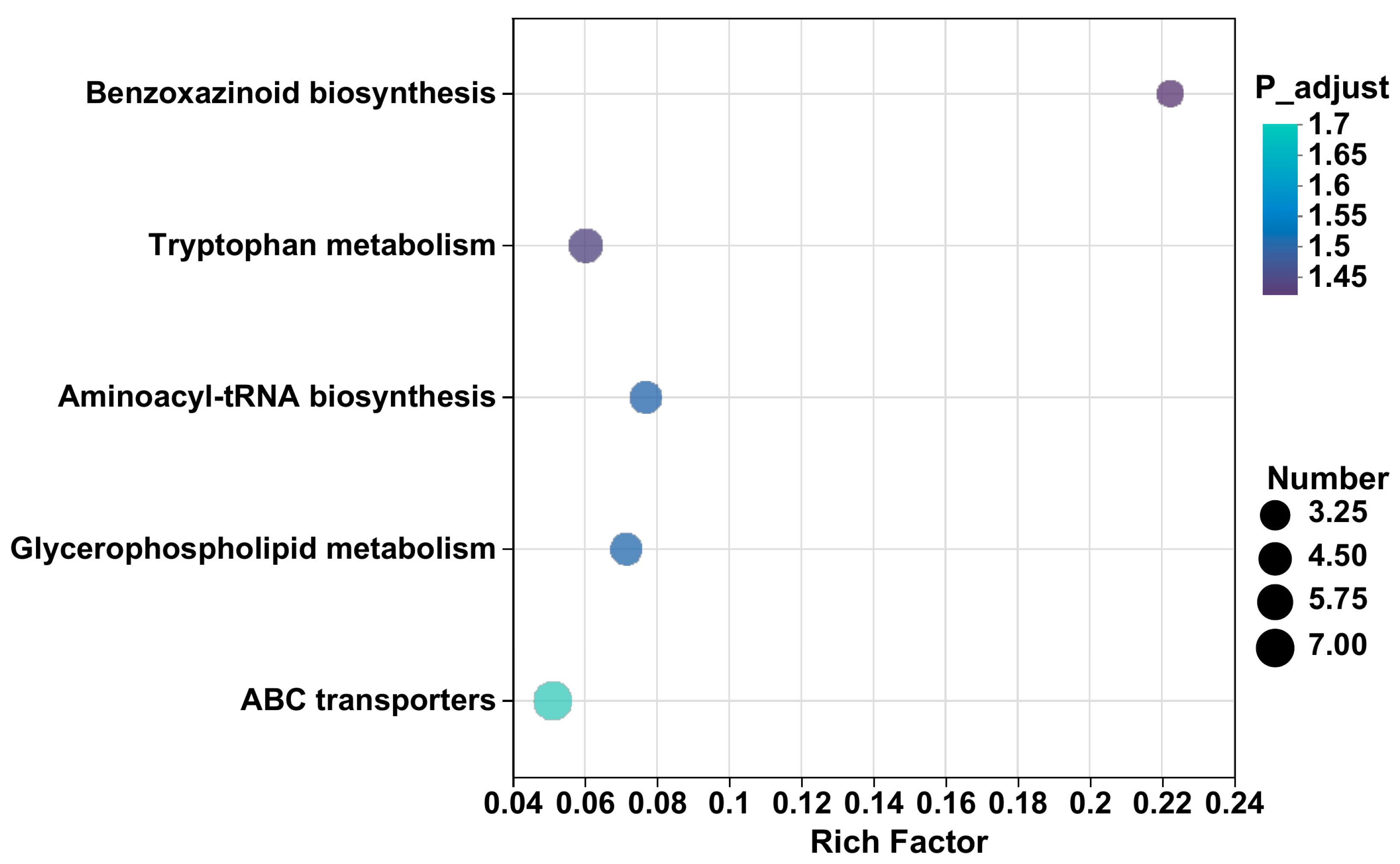
3.2.3. KEGG Enrichment Analysis
4. Discussion
4.1. Effects of AM Fungi Inoculation on Physiological Indicators of P. trifoliata
4.2. Regulation of Inoculation with AM Fungi on Metabolic Pathways in P. trifoliata
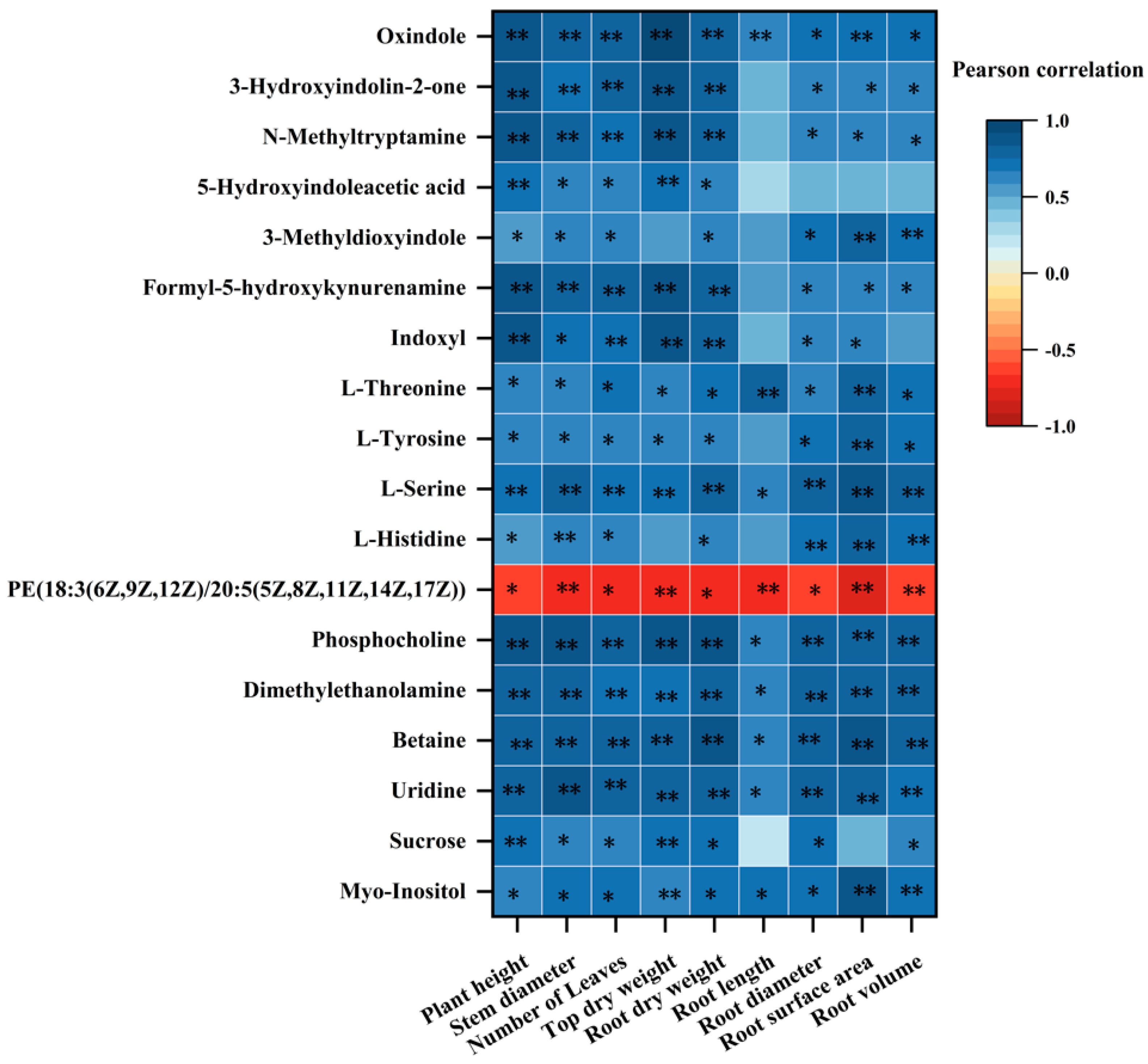
4.3. Correlation between the Differential Metabolites of P. trifoliata Roots and the Growth Indexes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, J.; Zhang, Y.; Zhang, P.; Trivedi, P.; Riera, N.; Wang, Y.; Liu, X.; Fan, G.; Tang, J.; Coletta-Filho, H.D.; et al. The structure and function of the global citrus rhizosphere microbiome. Nat. Commun. 2018, 9, 4894. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Qi, C.N. Status Quo and Development Trend of World’s Citrus Industry. Agric. Outlook 2016, 12, 46–52. [Google Scholar]
- Liu, M.Z.; Yi, X.T.; Yang, C.; Chen, X.L.; Fu, X.Z.; Li, G.G.; Peng, L.Z.; Chen, D.K.; Liao, H.H.; Huang, Q.C.; et al. Soil nutrient status analysis of citrus orchard in south-central Guangxi. J. Hunan Agric. Univ. (Nat. Sci.) 2022, 48, 460–467. [Google Scholar]
- Wu, Q.S.; Xia, R.X. Arbuscular mycorrhizal fungi influence growth, osmotic adjustment and photosynthesis of citrus under well-watered and water stress conditions. J. Plant Physiol. 2006, 163, 417–425. [Google Scholar] [CrossRef] [PubMed]
- Fang, X.; Lee, X.; Twagirayezu, G.; Cheng, H.; Lu, H.; Huang, S.; Deng, L.; Ji, B. A Critical Review of the Effectiveness of Biochar Coupled with Arbuscular Mycorrhizal Fungi in Soil Cadmium Immobilization. J. Fungi 2024, 10, 182. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Li, K.; Zhu, X.; Wang, F. The Worldwide Leaf Economic Spectrum Traits Are Closely Linked with Mycorrhizal Traits. Fungal Ecol. 2020, 43, 100877. [Google Scholar] [CrossRef]
- Gujre, N.; Soni, A.; Rangan, L.; Tsang, D.; Mitra, S. Sustainable improvement of soil health utilizing biochar and arbuscular mycorrhizal fungi: A review. Environ. Pollut. 2021, 268, 115549. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, E.; Bedini, S. Enhancing ecosystem services in sustainable agriculture. Bibfertilization Biofortification Chickpea (Cicer arietinum L.) By Arbuscular Mycorrhiza Fungi. Soil Biol. Biochem. 2014, 68, 429–439. [Google Scholar] [CrossRef]
- Zhang, H.; Cheng, H.; Twagirayezu, G.; Zhang, F.; Shi, Y.; Luo, C.; Yan, F.; Wang, Z.; Xing, D. Arbuscular Mycorrhizal Fungi Adjusts Root Architecture to Promote Leaf Nitrogen Accumulation and Reduce Leaf Carbon–Nitrogen Ratio of Mulberry Seedlings. Forests 2023, 14, 2448. [Google Scholar] [CrossRef]
- Pozo, M.J.; Azcon-Aguilar, C. Unraveling mycorrhiza-induced resistance. Curr. Opin. Plant Biol. 2007, 10, 393–398. [Google Scholar] [CrossRef]
- Riaz, M.; Kamran, M.; Fang, Y.; Wang, Q.; Cao, H.; Yang, G.; Deng, L.; Wang, Y.; Zhou, Y.; Anastopoulos, I.; et al. Arbuscular mycorrhizal fungi-induced mitigation of heavy metal phytotoxicity in metal contaminated soils: A critical review. J. Hazard. Mater. 2021, 402, 123919. [Google Scholar] [CrossRef]
- Niu, L.L.; Gao, C.; Zeng, M.; Li, W.; Lei, S.M.; Wu, J.Y. Effects of AM Fungi on Growth and Photosynthesis of Citrus Rootstock Seedlings in Soils with Different pH. South China Fruits 2012, 5, 13–16. [Google Scholar]
- Zhang, J.L.; Li, M.Y.; Kang, Y.H.; Liu, J.H.; Chen, T.S.; Li, X.L.; Song, J.; Liu, S.Q. Effect of Fourteen Arbuscular Mycorrhizal Fungi on Growth of Fructus aurantil. Chin. J. Trop. Crop. 2021, 42, 3278–3283. [Google Scholar]
- Wu, Q.S. Field Inoculation of Arbuscular mycorrhizal fungi Improves fruit quality and root physiological activity of Citrus. Agriculture 2021, 11, 1297. [Google Scholar] [CrossRef]
- Eroglu, C.G.; Cabral, C.; Ravnskov, S.; Bak, T.H.; Wollenweber, B. Arbuscular mycorrhiza influences carbon-use efficiency and grain yield of wheat grown under pre- and post-anthesis salinity stress. Plant Biol. 2020, 22, 863–871. [Google Scholar] [CrossRef] [PubMed]
- Porcel, R.; Redondo-Gomez, S.; Mateos-Naranjo, E.; Aroca, R.; Garcia, R.; Ruiz-Lozano, J.M. Arbuscular mycorrhizal symbiosis ameliorates the optimum quantum yield of photosystem II and reduces non-photochemical quenching in rice plants subjected to salt stress. J. Plant Physiol. 2015, 185, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.S.; Xie, Y.C.; Rahman, M.M.; Hashem, A.; Abd, A.E.; Wu, Q.S. Arbuscular Mycorrhizal Fungi and Endophytic Fungi Activate Leaf Antioxidant Defense System of Lane Late Navel Orange. J. Fungi 2022, 8, 282. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.R. Effects of Arbuscular Mycorrhizal Fungi Inoculation on Growth Metabolism of Rose rugosa and Resistance to Lymantria dispar. Northeast Forestry University: Harbin, China,, 2022. [Google Scholar]
- Yang, H.H.; Zhang, Q.; Dai, Y.J.; Liu, Q.; Tang, J.J.; Bian, X.M.; Chen, X. Effects of arbuscular mycorrhizal fungi on plant growth depend on root system: A meta-analysis. Plant Soil 2015, 389, 361–374. [Google Scholar] [CrossRef]
- Yang, C.; Shen, S.; Zhou, S.; Li, Y.; Mao, Y.; Zhou, J.; Shi, Y.; An, L.; Zhou, Q.; Peng, W.; et al. Rice metabolic regulatory network spanning the entire life cycle. Mol. Plant 2022, 15, 258–275. [Google Scholar] [CrossRef]
- Fàbregas, N.; Lozano-Elena, F.; Blasco-Escamez, D.; Tohge, T.; Martínez-Andújar, C.; Albacete, A.; Osorio, S.; Bustamante, M.; Riechmann, J.L.; Nomura, T.; et al. Overexpression of the vascular brassinosteroid receptor BRL3 confers drought resistance without penalizing plant growth. Nat. Commun. 2018, 9, 4680. [Google Scholar] [CrossRef]
- Li, D.P.; Song, J.; LI, M.Y.; Liu, S.Q.; Guo, D.J.; Li, X.L.; Zhang, J.L.; Chen, T.S. Comparison of the Staining Effect of Five Dyes on AM Fungi in the Roots of Trifoliate Orange Grafted with Newhall Navel Orange. South China Fruits 2019, 49, 36–40. [Google Scholar]
- Edriss, M.H.; Davis, R.M.; Burger, D.W. Influence of mycorrhizal fungi on cytokinin production in sour orange. J. Am. Soc. Hortic. Sci. 1984, 109, 587–590. [Google Scholar] [CrossRef]
- Wang, C.H.; Yang, X.H.; Li, D.Y.; Yu, G.B.; Qin, Q. Effects of the Diferrent Species of Arbuscular Mycorrhizal Fungi on the Vegetative Growth and Mineral Contents in Trifoliate Orange Seedings. Chin. Agric. Sci. Bull. 2006, 12, 199–203. [Google Scholar]
- Schmitz, J.A.k.; de Souza, P.V.D.; Koler, O.C. Vegetatve growth of Ponirus trifolata L. Raf Inoculated Mycorhizal Fung Three Grow. Medlia. Commun. Soil Sci. Plant Anal. 2001, 32, 3031–3043. [Google Scholar] [CrossRef]
- Wu, Q.S.; Zou, Y.N.; Wang, G.Y. Arbuscular mycorrhizal fungi and acclimatization of micropropagated citrus. Commun. Soil Sci. Plant Analvsis 2011, 42, 1825–1832. [Google Scholar] [CrossRef]
- Cheng, H.; Wang, J.; Tu, C.; Lin, S.; Xing, D.; Hill, P.; Chadwick, D.; Jones, D.L. Arbuscular mycorrhizal fungi and biochar influence simazine decomposition and leaching. Glob. Chang. Biol. Bioenergy 2021, 13, 708–718. [Google Scholar] [CrossRef]
- Yang, H.S.; Zhang, Q.; Koide, R.T.; Hoeksema, J.D.; Tang, J.J.; Bian, X.M.; Hu, S.J.; Chen, X. Taxonomic resolution is a determinant of biodiversity effects in arbuscular mycorrhizal fungal communities. J. Ecol. 2017, 105, 219–228. [Google Scholar] [CrossRef]
- Yao, Q.; Wang, L.R.; Zhu, H.H.; Chen, J.Z. Effect of arbuscular mycorrhizal fungal inoculation on root system architecture of trifoliate orange (Poncirus trifoliata L. Raf.) seedlings. Sci. Hort. 2009, 21, 458–461. [Google Scholar] [CrossRef]
- Li, L.L. Regulation of Root System Architecture in Trifoliate Orangeseedlings Treated by Peroxide Hydrogen and Arbuscularmycorrhizal Fungi; Yangtze University: Jingzhou, China, 2013. [Google Scholar]
- Saia, S.; Ruisi, P.; Fileccia, V.; Di Miceli, G.; Amato, G.; Martinelli, F. Metabolomics Suggests That Soil Inoculation with Arbuscular Mycorrhizal Fungi Decreased Free Amino Acid Content in Roots of Durum Wheat Grown under N-Limited, P-Rich Field Conditions. PLoS ONE 2015, 10, e0129591. [Google Scholar] [CrossRef]
- Wu, W.Y.; Wu, K.X.; Zang, M.; Mu, L.Q. Metabolic Difference Analysis of Three Gentiana Species Based on GC-MS. J. Northwest For. Univ. 2023, 6, 1–8. [Google Scholar]
- Clemente, M.; Corigliano, M.G.; Pariani, S.A.; Sanchez-Lopez, E.F.; Sander, V.A.; Ramos-Duarte, V.A. Plant Serine Protease Inhibitors: Biotechnology Application in Agriculture and Molecular Farming. Int. J. Mol. Sci. 2019, 20, 1345. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.S.; He, J.D.; Srivastava, A.K.; Zou, Y.N.; Kuca, K. Mycorrhizas enhance drought tolerance of citrus by altering root fatty acid compositions and their saturation levels. Tree Physiol. 2019, 39, 1149–1158. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Y. Effect of Exogenous Trehalose on Sugar Metabolism Response to Salt Stress in Tomato Seedlings; Tianjin University: Tianjin, China, 2019. [Google Scholar]
- Jiang, Y.; Wang, W.; Xie, Q.; Liu, N.; Liu, L.; Wang, D.; Zhang, X.; Yang, C.; Chen, X.; Tang, D.; et al. Plants transfer lipids to sustain colonization by mutualistic mycorrhizal and parasitic fungi. Science 2017, 356, 1172–1175. [Google Scholar] [CrossRef] [PubMed]
- Keymer, A.; Gutjahr, C. Cross-kingdom lipid transfer in arbuscular mycorrhiza symbiosis and beyond. Curr. Opin. Plant Biol. 2018, 44, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Seddas, P.M.; Arias, C.M.; Arnould, C.; van-Tuinen, D.; Godfroy, O.; Benhassou, H.A.; Gouzy, J.; Morandi, D.; Dessaint, F.; Gianinazzi-Pearson, V.; et al. Symbiosis-related plant genes modulate molecular responses in an arbuscular mycorrhizal fungus during early root interactions. Mol. Plant Microbe Interact. 2009, 22, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Bravo, A.; York, T.; Pumplin, N.; Mueller, L.A.; Harrison, M.J. Genes conserved for arbuscular mycorrhizal symbiosis identified through phylogenomics. Nat. Plants 2016, 2, 15208. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, T.M.; Nunes, N.A.; Araujo, W.L.; Braun, H.P. Amino Acid Catabolism in Plants. Mol. Plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef] [PubMed]
- Gachomo, E.; Allen, J.W.; Pfeffer, P.E.; Govindarajulu, M.; Douds, D.D.; Jin, H.; Nagahashi, G.; Lammers, P.J.; Shachar-Hill, Y.; Bucking, H. Germinating spores of Glomus intraradices can use internal and exogenous nitrogen sources for de novo biosynthesis of amino acids. New Phytol. 2009, 184, 399–411. [Google Scholar] [CrossRef] [PubMed]
- Fester, T.; Fetzer, I.; Buchert, S.; Lucas, R.; Rillig, M.C.; Hartig, C. Towards a systemic metabolic signature of the arbuscular mycorrhizal interaction. Oecologia 2011, 167, 913–924. [Google Scholar] [CrossRef] [PubMed]
- Souza, L.A.; Camargos, L.S.; Schiavinato, M.A.; Andrade, S.A.L. Mycorrhization alters foliar soluble amino acid composition and influences tolerance to Pb in Calopogonium mucunoides. Theor. Exp. Plant Physiol. 2014, 26, 211–216. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Soil |
|---|---|
| pH | 6.73 ± 0.02 |
| Organic matter (g/kg) | 79.99 ± 2.58 |
| Total phosphorus (g/kg) | 0.60 ± 0.01 |
| Alkali-hydrolyzed nitrogen (mg/kg) | 95.16 ± 3.12 |
| Available phosphorus (mg/kg) | 67.75 ± 2.74 |
| Available potassium (g/kg) | 708.50 ± 20.11 |
| Pathway Description | Rich Factor | p-Value | Differential Metabolites | Up/Down |
|---|---|---|---|---|
| ABC transporters | 0.05 | 0.02 | L-threonine | up |
| Myo-inositol | up | |||
| L-histidine | up | |||
| L-serine | up | |||
| Sucrose | up | |||
| Uridine | up | |||
| Betaine | up | |||
| Glycerophospholipid metabolism | 0.07 | 0.03 | Dimethylethanolamine | up |
| Phosphocholine | up | |||
| L-serine | up | |||
| PE(18:3(6Z,9Z,12Z)/20:5(5Z,8Z,11Z,14Z,17Z)) | down | |||
| Aminoacyl-tRNA biosynthesis | 0.08 | 0.03 | L-histidine | up |
| L-serine | up | |||
| L-tyrosine | up | |||
| L-threonine | up | |||
| Tryptophan metabolism | 0.06 | 0.04 | Indoxyl | up |
| Formyl-5-hydroxykynurenamine | up | |||
| 3-methyldioxyindole | up | |||
| 5-hydroxyindoleacetic acid | up | |||
| N-methyltryptamine | up | |||
| Benzoxazinoid biosynthesis | 0.22 | 0.04 | 3-hydroxyindolin-2-one | up |
| Oxindole | up |
| Parameters | CK Treatment Substrate | AMF Treatment Substrate |
|---|---|---|
| pH | 6.66 ± 0.02 | 6.70 ± 0.01 |
| Organic matter (g/kg) | 97.50 ± 5.34 | 80.51 ± 1.79 |
| Total phosphorus (g/kg) | 0.77 ± 0.01 | 0.69 ± 0.01 |
| Alkali-hydrolyzed nitrogen (mg/kg) | 103.16 ± 1.62 | 94.03 ± 1.2 |
| Available phosphorus (mg/kg) | 77.09 ± 3.66 | 91.22 ± 1.95 |
| Available potassium (g/kg) | 714.67 ± 20.56 | 660.00 ± 11.12 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, Y.; Twagirayezu, G.; Xu, J.; Wen, Y.; Shang, P.; Song, J.; Wang, Q.; Li, X.; Liu, S.; Chen, T.; et al. Arbuscular Mycorrhizal Fungi Regulate Lipid and Amino Acid Metabolic Pathways to Promote the Growth of Poncirus trifoliata (L.) Raf. J. Fungi 2024, 10, 427. https://doi.org/10.3390/jof10060427
Kang Y, Twagirayezu G, Xu J, Wen Y, Shang P, Song J, Wang Q, Li X, Liu S, Chen T, et al. Arbuscular Mycorrhizal Fungi Regulate Lipid and Amino Acid Metabolic Pathways to Promote the Growth of Poncirus trifoliata (L.) Raf. Journal of Fungi. 2024; 10(6):427. https://doi.org/10.3390/jof10060427
Chicago/Turabian StyleKang, Yihao, Gratien Twagirayezu, Jie Xu, Yunying Wen, Pengxiang Shang, Juan Song, Qian Wang, Xianliang Li, Shengqiu Liu, Tingsu Chen, and et al. 2024. "Arbuscular Mycorrhizal Fungi Regulate Lipid and Amino Acid Metabolic Pathways to Promote the Growth of Poncirus trifoliata (L.) Raf" Journal of Fungi 10, no. 6: 427. https://doi.org/10.3390/jof10060427
APA StyleKang, Y., Twagirayezu, G., Xu, J., Wen, Y., Shang, P., Song, J., Wang, Q., Li, X., Liu, S., Chen, T., Cheng, T., & Zhang, J. (2024). Arbuscular Mycorrhizal Fungi Regulate Lipid and Amino Acid Metabolic Pathways to Promote the Growth of Poncirus trifoliata (L.) Raf. Journal of Fungi, 10(6), 427. https://doi.org/10.3390/jof10060427