
Development of an RNA Nanostructure for Effective Botrytis cinerea Control through Spray-Induced Gene Silencing without an Extra Nanocarrier

Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of Plant Materials and Inoculation of Botrytis cinerea
2.2. Design of a B. cinerea-Targeted RNA Nanoparticle
2.3. Construction of RNA Squares and dsRNA Expression Vectors
2.4. In Vivo Synthesis of the RNA Square and dsRNA
2.5. Conidiospore Germination Assays
2.6. Inhibitory Effects of the RNA Square on B. cinerea Mycelial Growth
2.7. Bioassay of the RNA Square against B. cinerea
2.8. Total RNA Extraction and Quantitative RT–PCR (RT-qPCR)
2.9. Stem-Loop RT-qPCR
2.10. Statistical Analysis
3. Results
3.1. Design of a B. cinerea-Targeted Square RNA Nanostructure
3.2. Production of RNA Nanoparticles
3.3. The Anti-B. cinerea Activity of the RNA Square
3.4. Bioassay of the RNA Square in Tobacco Leaves against B. cinerea
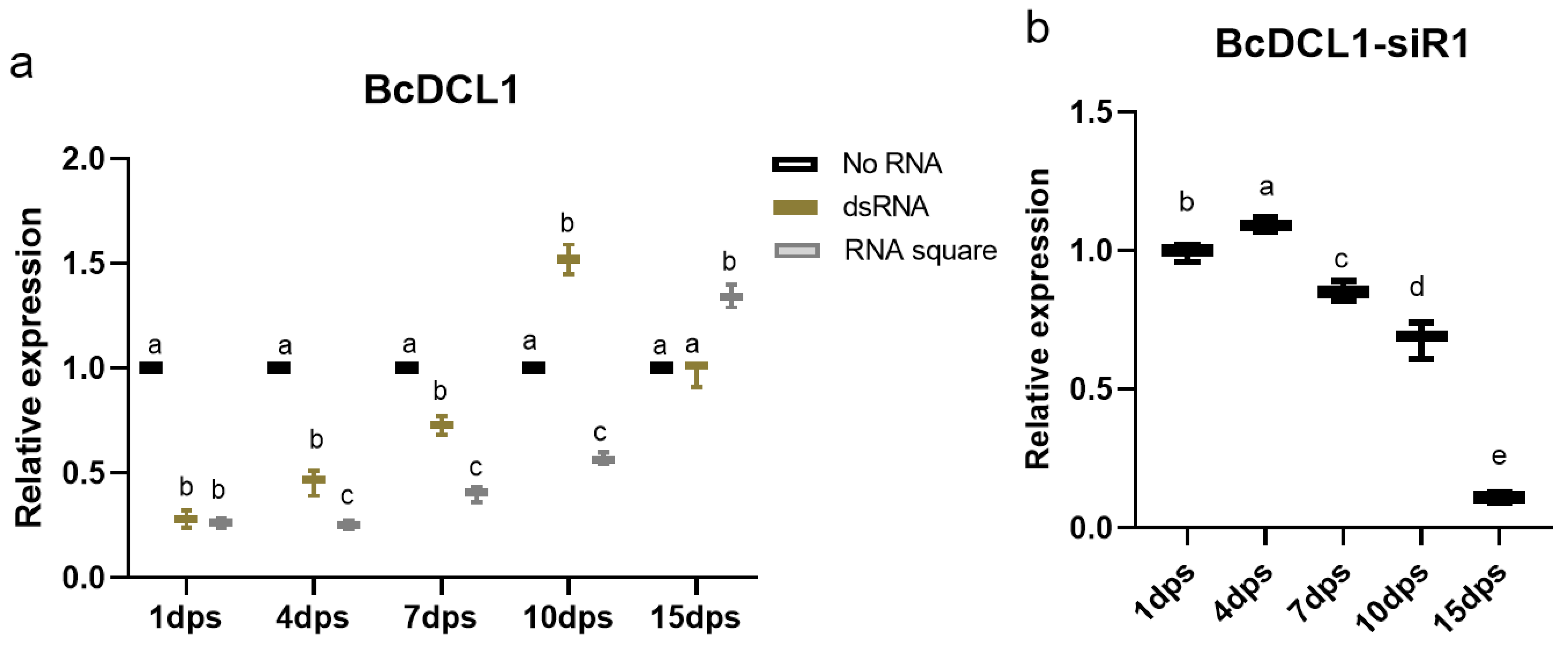
3.5. siRNA Generation and RNAi Efficiency of the RNA Squares
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qiao, L.; Lan, C.; Capriotti, L.; Ah-Fong, A.; Nino Sanchez, J.; Hamby, R.; Heller, J.; Zhao, H.; Glass, N.L.; Judelson, H.S.; et al. Spray-induced gene silencing for disease control is dependent on the efficiency of pathogen RNA uptake. Plant Biotechnol. J. 2021, 19, 1756–1768. [Google Scholar] [CrossRef]
- Hernández-Soto, A.; Chacón-Cerdas, R. RNAi Crop Protection Advances. Int. J. Mol. Sci. 2021, 22, 12148. [Google Scholar] [CrossRef] [PubMed]
- Worrall, E.A.; Hamid, A.; Mody, K.T.; Mitter, N.; Pappu, H.R. Nanotechnology for Plant Disease Management. Agronomy 2018, 8, 285. [Google Scholar] [CrossRef]
- Wang, M.; Jin, H. Spray-Induced Gene Silencing: A Powerful Innovative Strategy for Crop Protection. Trends Microbiol. 2017, 25, 4–6. [Google Scholar] [CrossRef] [PubMed]
- Niu, D.; Hamby, R.; Sanchez, J.N.; Cai, Q.; Yan, Q.; Jin, H. RNAs—A new frontier in crop protection. Curr. Opin. Biotechnol. 2021, 70, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Mitter, N.; Worrall, E.A.; Robinson, K.E.; Xu, Z.P.; Carroll, B.J. Induction of virus resistance by exogenous application of double-stranded RNA. Curr. Opin. Virol. 2017, 26, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Christiaens, O.; Tardajos, M.G.; Martinez Reyna, Z.L.; Dash, M.; Dubruel, P.; Smagghe, G. Increased RNAi Efficacy in Spodoptera exigua via the Formulation of dsRNA with Guanylated Polymers. Front. Physiol. 2018, 9, 316. [Google Scholar] [CrossRef] [PubMed]
- Taning, C.N.T.; Christiaens, O.; Berkvens, N.; Casteels, H.; Maes, M.; Smagghe, G. Oral RNAi to control Drosophila suzukii: Laboratory testing against larval and adult stages. J. Pest Sci. 2016, 89, 803–814. [Google Scholar] [CrossRef]
- Deng, R.; Lin, D.; Zhu, L.; Majumdar, S.; White, J.C.; Gardea-Torresdey, J.L.; Xing, B. Nanoparticle interactions with co-existing contaminants: Joint toxicity, bioaccumulation and risk. Nanotoxicology 2017, 11, 591–612. [Google Scholar] [CrossRef]
- Li, L.; Xu, Z.; Kah, M.; Lin, D.; Filser, J. Nanopesticides: A comprehensive assessment of environmental risk is needed before widespread agricultural application. Environ. Sci. Technol. 2019, 53, 7923–7924. [Google Scholar] [CrossRef]
- Kannan, M.; Bojan, N.; Swaminathan, J.; Zicarelli, G.; Hemalatha, D.; Zhang, Y.; Ramesh, M.; Faggio, C. Nanopesticides in agricultural pest management and their environmental risks: A review. Int. J. Environ. Sci. Technol. 2023, 20, 10507–10532. [Google Scholar] [CrossRef]
- Shu, D.; Shu, Y.; Haque, F.; Abdelmawla, S.; Guo, P.X. Thermodynamically stable RNA three-way junction for constructing multifunctional nanoparticles for delivery of therapeutics. Nat. Nanotechnol. 2011, 6, 658–667. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.R.; Wang, H.Z.; Li, Z.F.; Shu, D.; Guo, P.X. RNA Micelles for the Systemic Delivery of Anti-miRNA for Cancer Targeting and Inhibition without Ligand. ACS Nano 2019, 13, 706–717. [Google Scholar] [CrossRef] [PubMed]
- Haque, F.; Shu, D.; Shu, Y.; Shlyakhtenko, L.S.; Rychahou, P.G.; Evers, B.M.; Guo, P.X. Ultrastable synergistic tetravalent RNA nanoparticles for targeting to cancers. Nano Today 2012, 7, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Afonin, K.A.; Kireeva, M.; Grabow, W.W.; Kashlev, M.; Jaeger, L.; Shapiro, B.A. Co-transcriptional assembly of chemically modified RNA nanoparticles functionalized with siRNAs. Nano Lett. 2012, 12, 5192–5195. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.B.; Hong, J.; Bonner, D.K.; Poon, Z.; Hammond, P.T. Self-assembled RNA interference microsponges for efficient siRNA delivery. Nat. Mater. 2012, 11, 316–322. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.M.; Viard, M.; Subramanian, H.K.; Roark, B.K.; Afonin, K.A.; Franco, E. Programmable RNA microstructures for coordinated delivery of siRNAs. Nanoscale 2016, 8, 17542–17550. [Google Scholar] [CrossRef] [PubMed]
- Rackley, L.; Stewart, J.M.; Salotti, J.; Krokhotin, A.; Shah, A.; Halman, J.R.; Juneja, R.; Smollett, J.; Lee, L.; Roark, K.; et al. RNA Fibers as Optimized Nanoscaffolds for siRNA Coordination and Reduced Immunological Recognition. Adv. Funct. Mater. 2018, 28, 1805959. [Google Scholar] [CrossRef]
- Jedrzejczyk, D.; Chworos, A. Self-Assembling RNA Nanoparticle for Gene Expression Regulation in a Model System. ACS Synth. Biol. 2019, 8, 491–497. [Google Scholar] [CrossRef]
- Zakrevsky, P.; Kasprzak, W.K.; Heinz, W.F.; Wu, W.; Khant, H.; Bindewald, E.; Dorjsuren, N.; Fields, E.A.; de Val, N.; Jaeger, L.; et al. Truncated tetrahedral RNA nanostructures exhibit enhanced features for delivery of RNAi substrates. Nanoscale 2020, 12, 2555–2568. [Google Scholar] [CrossRef]
- Li, M.; Zheng, M.; Wu, S.; Tian, C.; Liu, D.; Weizmann, Y.; Jiang, W.; Wang, G.; Mao, C. In vivo production of RNA nanostructures via programmed folding of single-stranded RNAs. Nat. Commun. 2018, 9, 2196. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Weiberg, A.; Lin, F.M.; Thomma, B.P.; Huang, H.D.; Jin, H. Bidirectional cross-kingdom RNAi and fungal uptake of external RNAs confer plant protection. Nat. Plants 2016, 2, 16151. [Google Scholar] [CrossRef] [PubMed]
- Vert, J.P.; Foveau, N.; Lajaunie, C.; Vandenbrouck, Y. An accurate and interpretable model for siRNA efficacy prediction. BMC Bioinformatics 2006, 7, 520. [Google Scholar] [CrossRef] [PubMed]
- BLAST. Available online: https://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 23 April 2023).
- RNAfold Web Serve. Available online: http://rna.tbi.univie.ac.at/cgi-bin/RNAWebSuite/RNAfold.cgi (accessed on 23 April 2023).
- Bilir, Ö.; Telli, O.; Norman, C.; Budak, H.; Hong, Y.; Tör, M. Small RNA inhibits infection by downy mildew pathogen Hyaloperonospora arabidopsidis. Mol. Plant Pathol. 2019, 20, 1523–1534. [Google Scholar] [CrossRef]
- Meng, X.; Jin, W.; Wu, F. Novel tomato miRNA miR1001 initiates cross-species regulation to suppress the conidiospore germination and infection virulence of Botrytis cinerea in vitro. Gene 2020, 759, 145002. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Long, L.; Zhang, F.; Chen, Q.; Chen, C.; Yu, X.; Liu, Q.; Bao, J.; Long, Z. Antifungal activity, main active components and mechanism of Curcuma longa extract against Fusarium graminearum. PLoS ONE 2018, 13, e0194284. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Varkonyi-Gasic, E. Stem-loop qRT-PCR for the detection of plant microRNAs. Methods Mol. Biol. 2016, 1456, 163–175. [Google Scholar]
- Guo, S.; Vieweger, M.; Zhang, K.; Yin, H.; Wang, H.; Li, X.; Li, S.; Hu, S.; Sparreboom, A.; Evers, B.M.; et al. Ultra-thermostable RNA nanoparticles for solubilizing and high-yield loading of paclitaxel for breast cancer therapy. Nat. Commun. 2020, 11, 972. [Google Scholar] [CrossRef]
- Nakashima, Y.; Abe, H.; Abe, N.; Aikawa, K.; Ito, Y. Branched RNA nanostructures for RNA interference. Chem. Commun. 2011, 47, 8367–8369. [Google Scholar] [CrossRef]
- Afonin, K.A.; Viard, M.; Koyfman, A.Y.; Martins, A.N.; Kasprzak, W.K.; Panigaj, M.; Desai, R.; Santhanam, A.; Grabow, W.W.; Jaeger, L.; et al. Multifunctional RNA nanoparticles. Nano Lett. 2014, 14, 5662–5671. [Google Scholar] [CrossRef] [PubMed]
- Jang, B.; Kim, B.; Kim, H.; Kwon, H.; Kim, M.; Seo, Y.; Colas, M.; Jeong, H.; Jeong, E.H.; Lee, K.; et al. Enzymatic Synthesis of Self-assembled Dicer Substrate RNA Nanostructures for Programmable Gene Silencing. Nano Lett. 2018, 18, 4279–4284. [Google Scholar] [CrossRef] [PubMed]
- Bilir, Ö.; Göl, D.; Hong, Y.; McDowell, J.M.; Tör, M. Small RNA-based plant protection against diseases. Front. Plant Sci. 2022, 13, 951097. [Google Scholar] [CrossRef] [PubMed]
- Das, P.R.; Sherif, S.M. Application of Exogenous dsRNAs-induced RNAi in Agriculture: Challenges and Triumphs. Front. Plant Sci. 2020, 11, 946. [Google Scholar] [CrossRef] [PubMed]
- Sang, H.; Kim, J.I. Advanced strategies to control plant pathogenic fungi by host-induced gene silencing (HIGS) and spray-induced gene silencing (SIGS). Plant Biotechnol. Rep. 2020, 14, 1–8. [Google Scholar] [CrossRef]
- Chen, A.; Halilovic, L.; Shay, J.H.; Koch, A.; Mitter, N.; Jin, H. Improving RNA-based crop protection through nanotechnology and insights from cross-kingdom RNA trafficking. Curr. Opin. Plant Biol. 2023, 76, 102441. [Google Scholar] [CrossRef]
- Niño-Sánchez, J.; Sambasivam, P.T.; Sawyer, A.; Hamby, R.; Chen, A.; Czislowski, E.; Li, P.; Manzie, N.; Gardiner, D.M.; Ford, R.; et al. BioClay™ prolongs RNA interference-mediated crop protection against Botrytis cinerea. J. Integr. Plant Biol. 2022, 64, 2187–2198. [Google Scholar] [CrossRef] [PubMed]
- Qiao, L.; Niño-Sánchez, J.; Hamby, R.; Capriotti, L.; Chen, A.; Mezzetti, B.; Jin, H. Artificial nanovesicles for dsRNA delivery in spray-induced gene silencing for crop protection. Plant Biotechnol. J. 2023, 21, 854–865. [Google Scholar] [CrossRef]
- Ahn, S.J.; Donahue, K.; Koh, Y.; Martin, R.R.; Choi, M.Y. Microbial-based double-stranded RNA production to develop cost-effective RNA interference application for insect pest management. Int. J. Insect Sci. 2019, 11, 1179543319840323. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| siRNA No. | Target Site | 21nt Sense Sequence | 21nt Antisense Sequence | Score |
|---|---|---|---|---|
| BcDCL1-siR1 | 2514–2534 | GGUAGAUGCUAGAGAUAAUGU | AUUAUCUCUAGCAUCUACCGG | 105.9 |
| BcDCL1-siR2 | 5522–2542 | CGGCAUACUUGUUCAUCUAUG | UAGAUGAACAAGUAUGCCGGA | 101.9 |
| BcPPI10-siR1 | 491–519 | GCAUCUCGAUGGUCAGAAUAC | AUUCUGACCAUCGAGAUGCGG | 96.1 |
| BcPPI10-siR2 | 619–639 | GGGUGACGGUGCAUGCUAAUC | UUAGCAUGCACCGUCACCCCC | 90.3 |
| BcNmt1-siR1 | 599–619 | CCCUCGUUCUGAUGUUCAAAU | UUGAACAUCAGAACGAGGGCG | 100.2 |
| BcNmt1-siR2 | 648–668 | GGAUGUUGUUGCUUCUGUUCC | AACAGAAGCAACAACAUCCAA | 102.3 |
| BcAC-siR1 | 650–670 | GGUAGAAAUUGAUGCUCAAGU | UUGAGCAUCAAUUUCUACCCU | 104.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, F.; Yan, L.; Zhao, X.; Lv, C.; Jin, W. Development of an RNA Nanostructure for Effective Botrytis cinerea Control through Spray-Induced Gene Silencing without an Extra Nanocarrier. J. Fungi 2024, 10, 483. https://doi.org/10.3390/jof10070483
Wu F, Yan L, Zhao X, Lv C, Jin W. Development of an RNA Nanostructure for Effective Botrytis cinerea Control through Spray-Induced Gene Silencing without an Extra Nanocarrier. Journal of Fungi. 2024; 10(7):483. https://doi.org/10.3390/jof10070483
Chicago/Turabian StyleWu, Fangli, Ling Yan, Xiayang Zhao, Chongrun Lv, and Weibo Jin. 2024. "Development of an RNA Nanostructure for Effective Botrytis cinerea Control through Spray-Induced Gene Silencing without an Extra Nanocarrier" Journal of Fungi 10, no. 7: 483. https://doi.org/10.3390/jof10070483