
The Arrestin-like Protein palF Contributes to Growth, Sporulation, Spore Germination, Osmolarity, and Pathogenicity of Coniella vitis
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
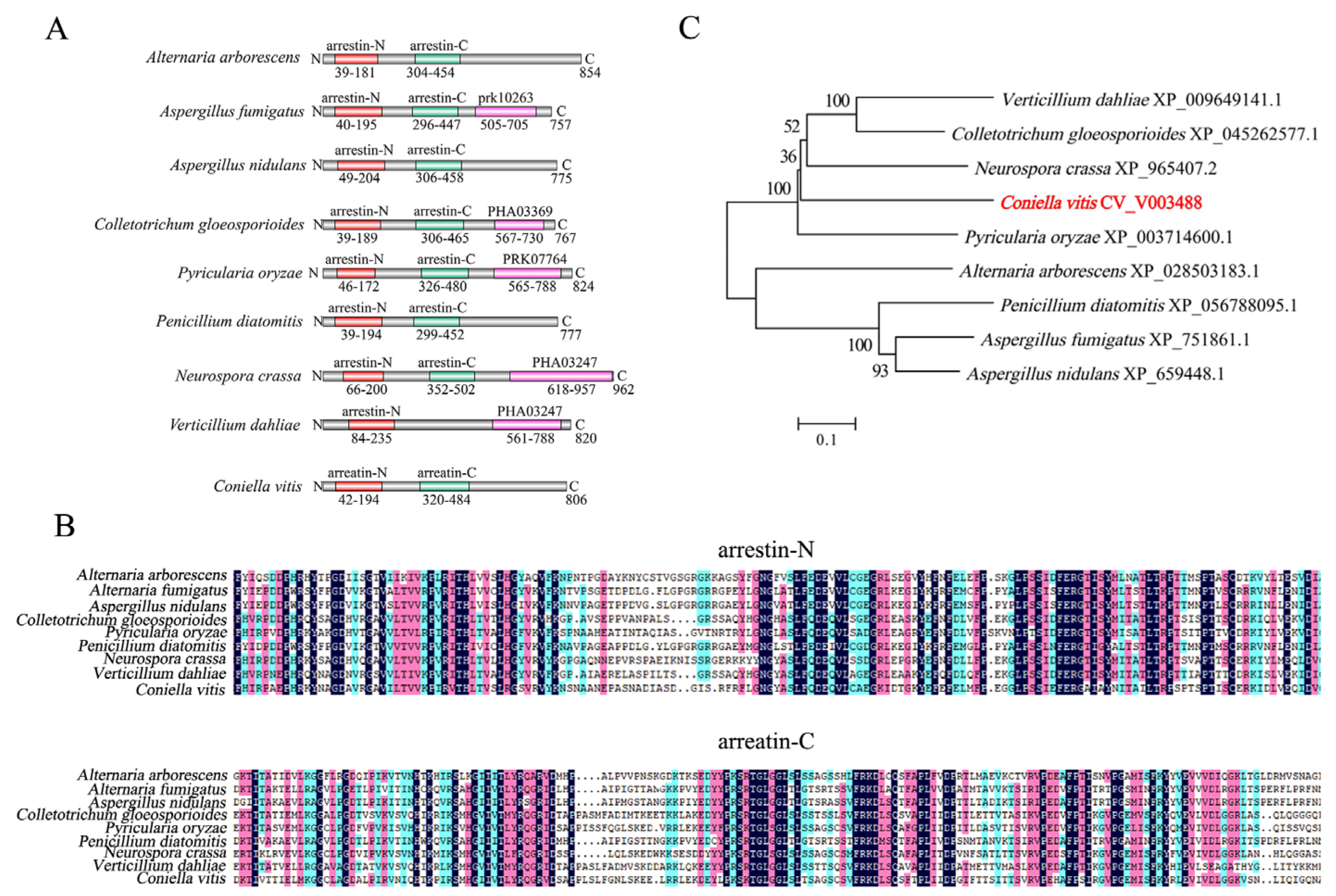
2.1. Strains, Culture Conditions, DNA Extraction, and Bioinformatics Analysis
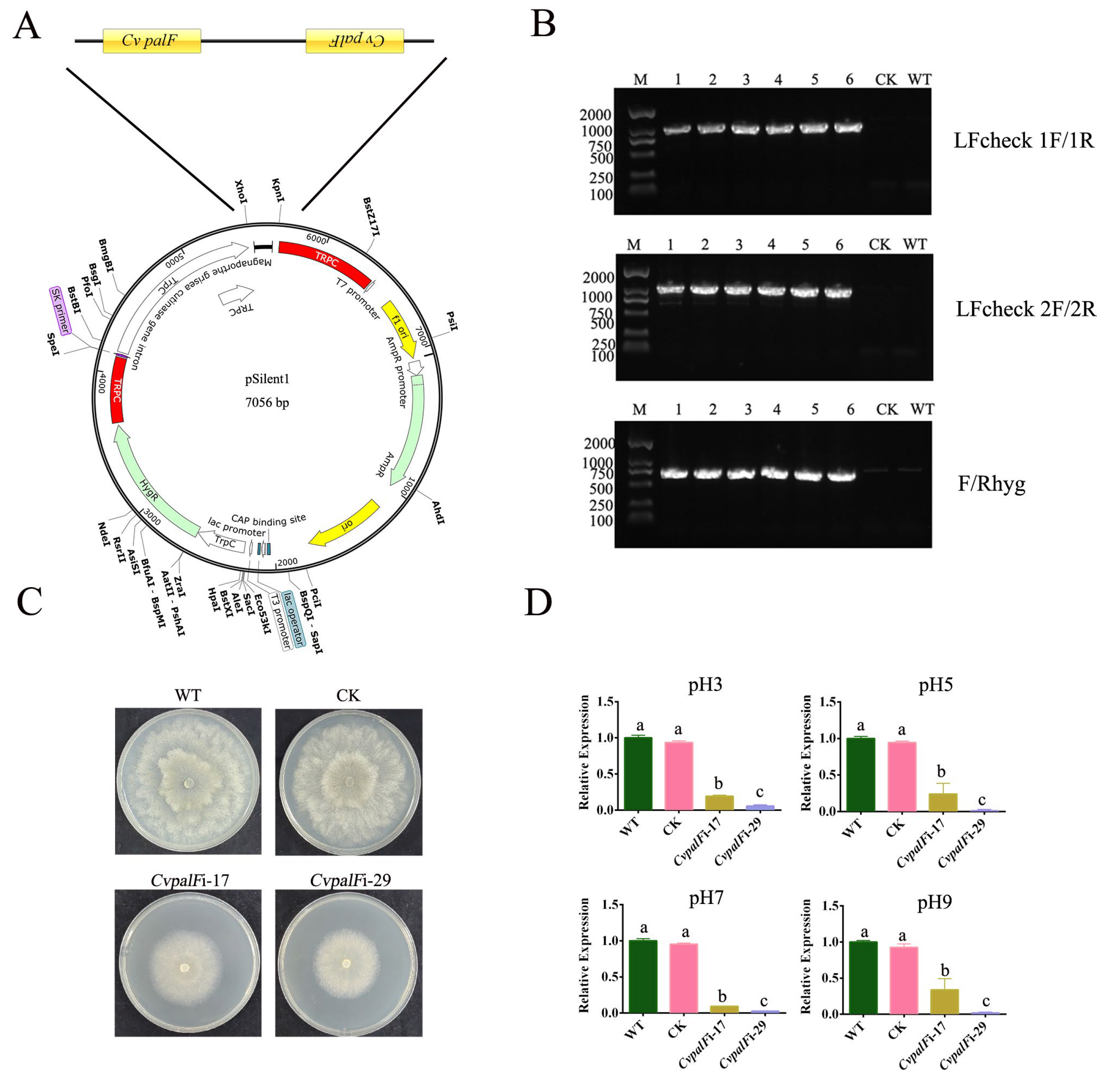
2.2. Construction of the CvpalF-Silenced Strains
2.2.1. Hygromycin Sensitivity
2.2.2. Construction of the CvpalF-Silencing Plasmid
2.2.3. Transformation Procedure
2.2.4. Gene-Expression Analysis
2.3. Silenced Strains Characterization
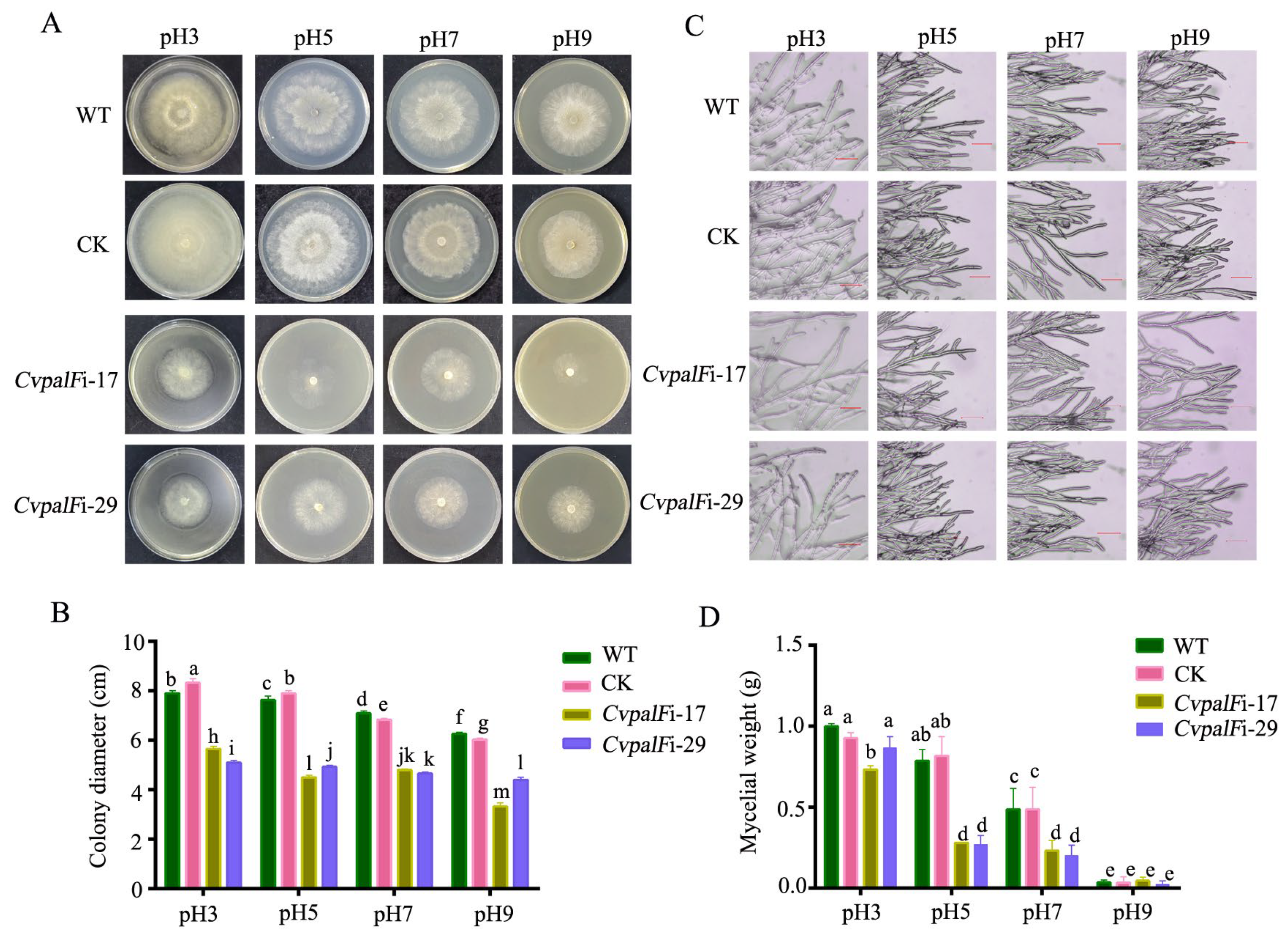
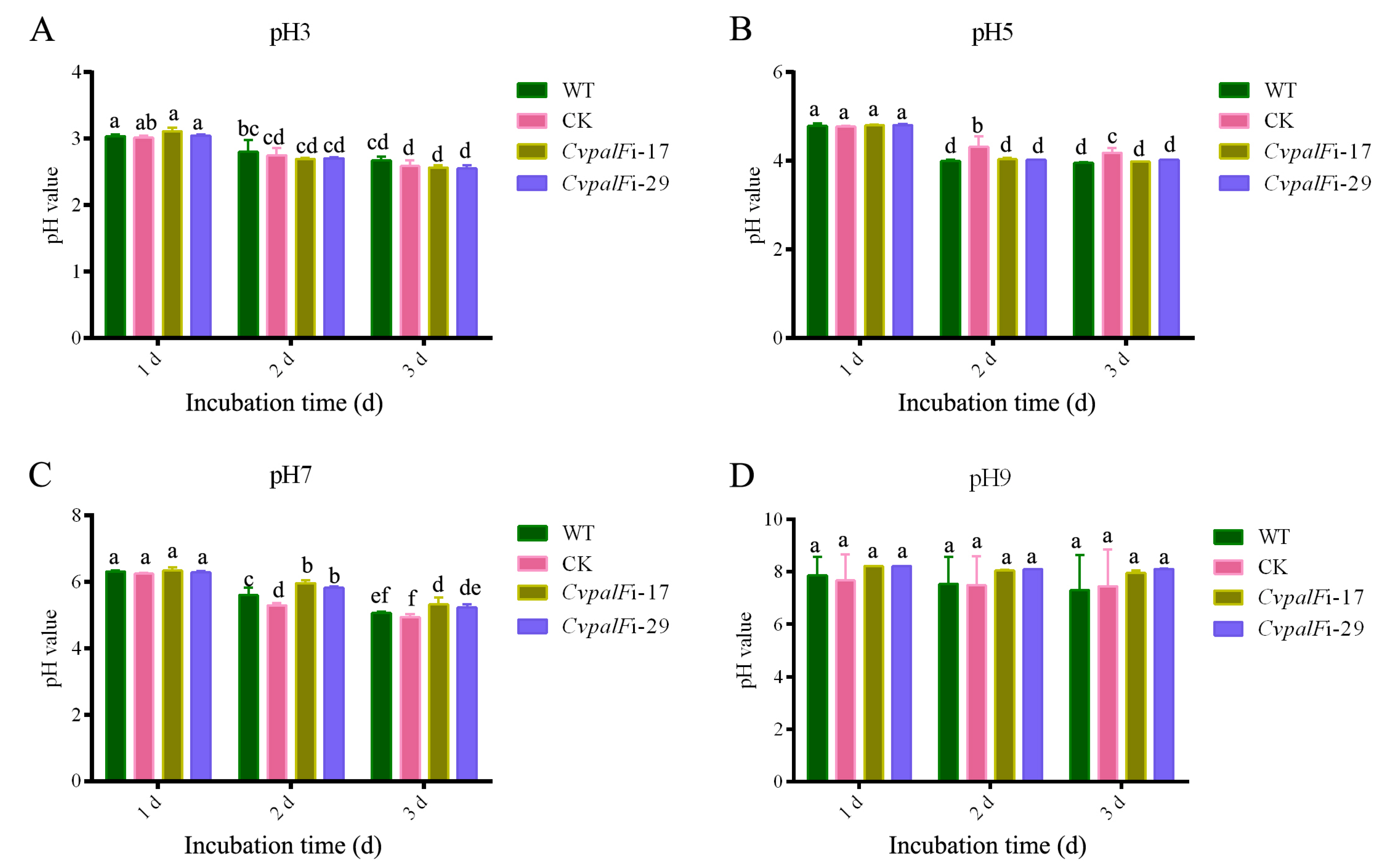
2.3.1. Growth under Different pH
On Solid Media
In Liquid Media
2.3.2. Spore Production and Vitality
2.3.3. Pathogenicity
2.3.4. Effects of Osmotic Stress
2.3.5. Virlence Gene Expression
2.4. Statistical Analysis
3. Results
3.1. Bioinformatics Analysis of CvpalF
3.2. Construction of CvpalF-Silenced Strains
3.3. Silenced Strains Characterization
3.3.1. The Growth of CvpalF-Silenced Strains Under Different pH Values
3.3.2. The pH Change of Liquid Media
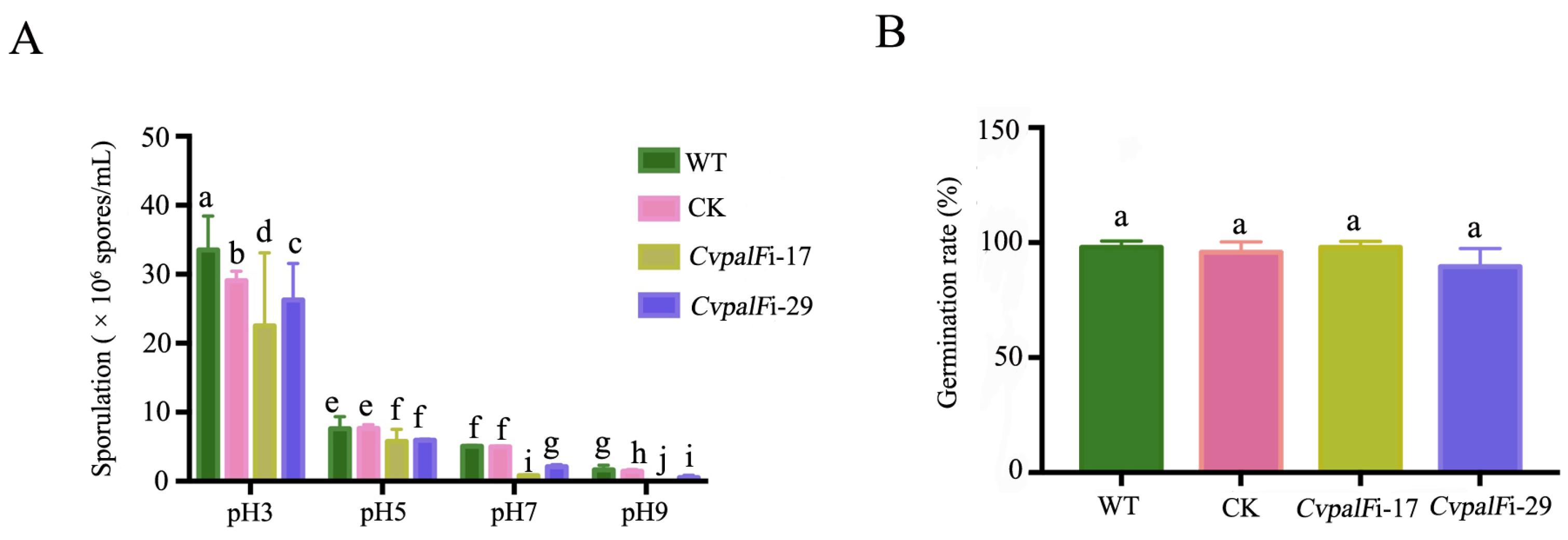
3.3.3. Sporulation and Spore Germination of CvpalF-Silenced Strains under Different pH Values
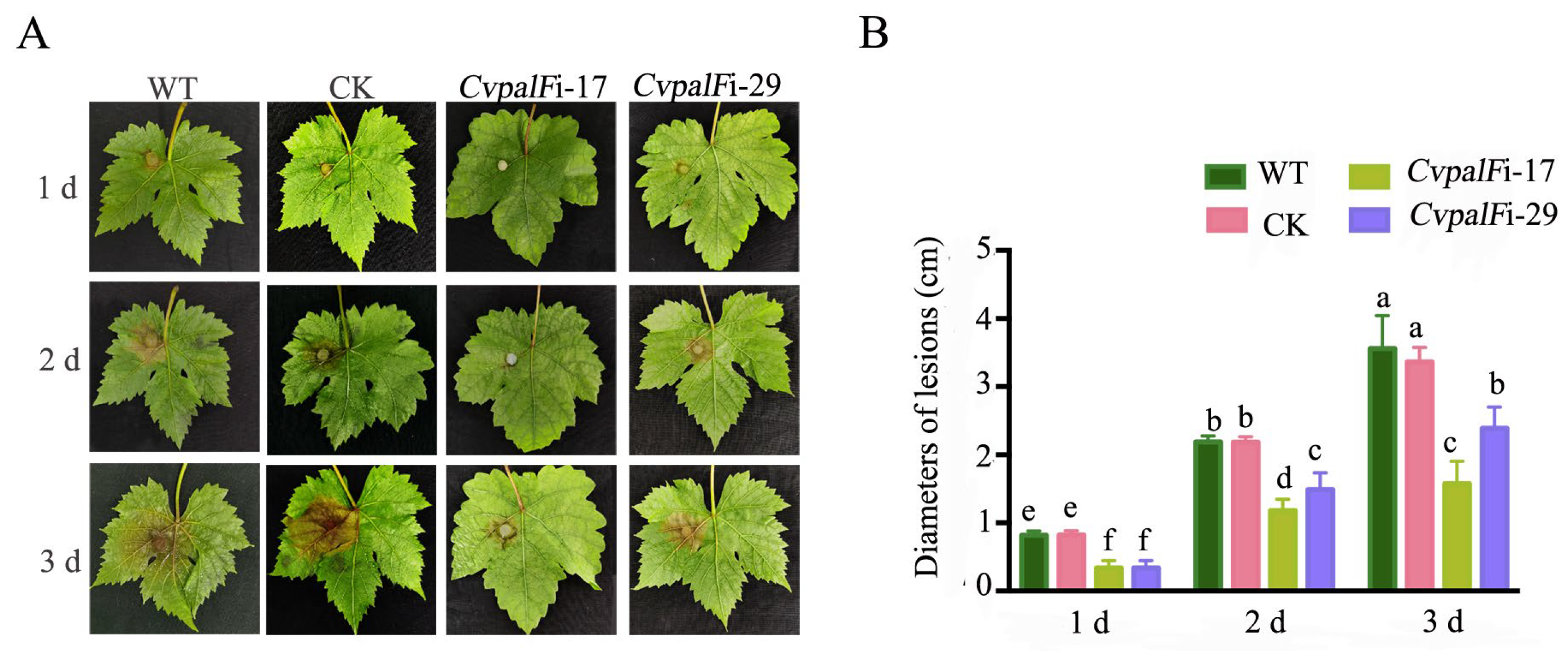
3.3.4. Pathogenicity of CvpalF-Silenced Strains
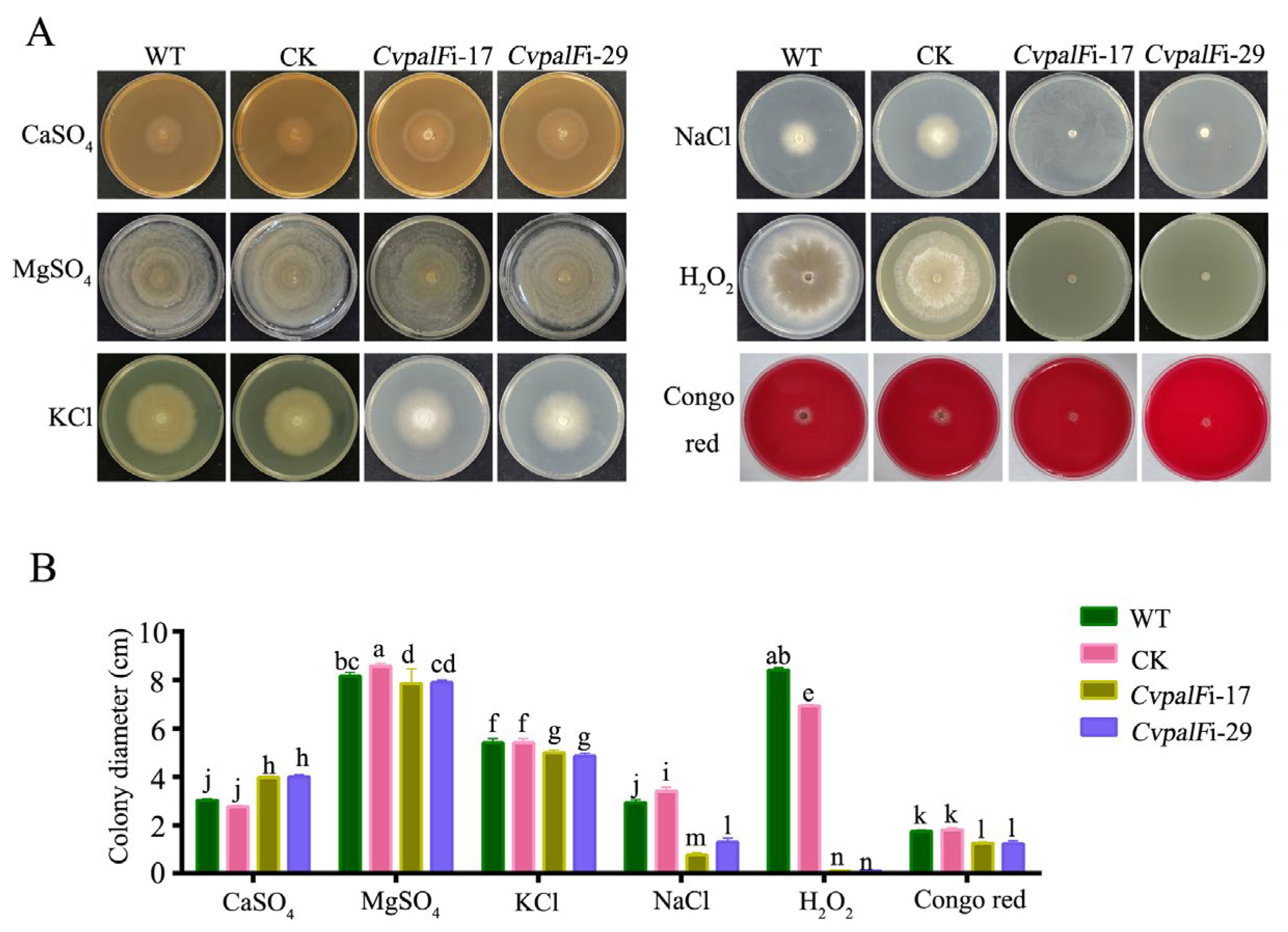
3.3.5. The Responses of CvpalF-Silenced Strains to Cell Wall Integrity, Oxidation, and Ionic Stress
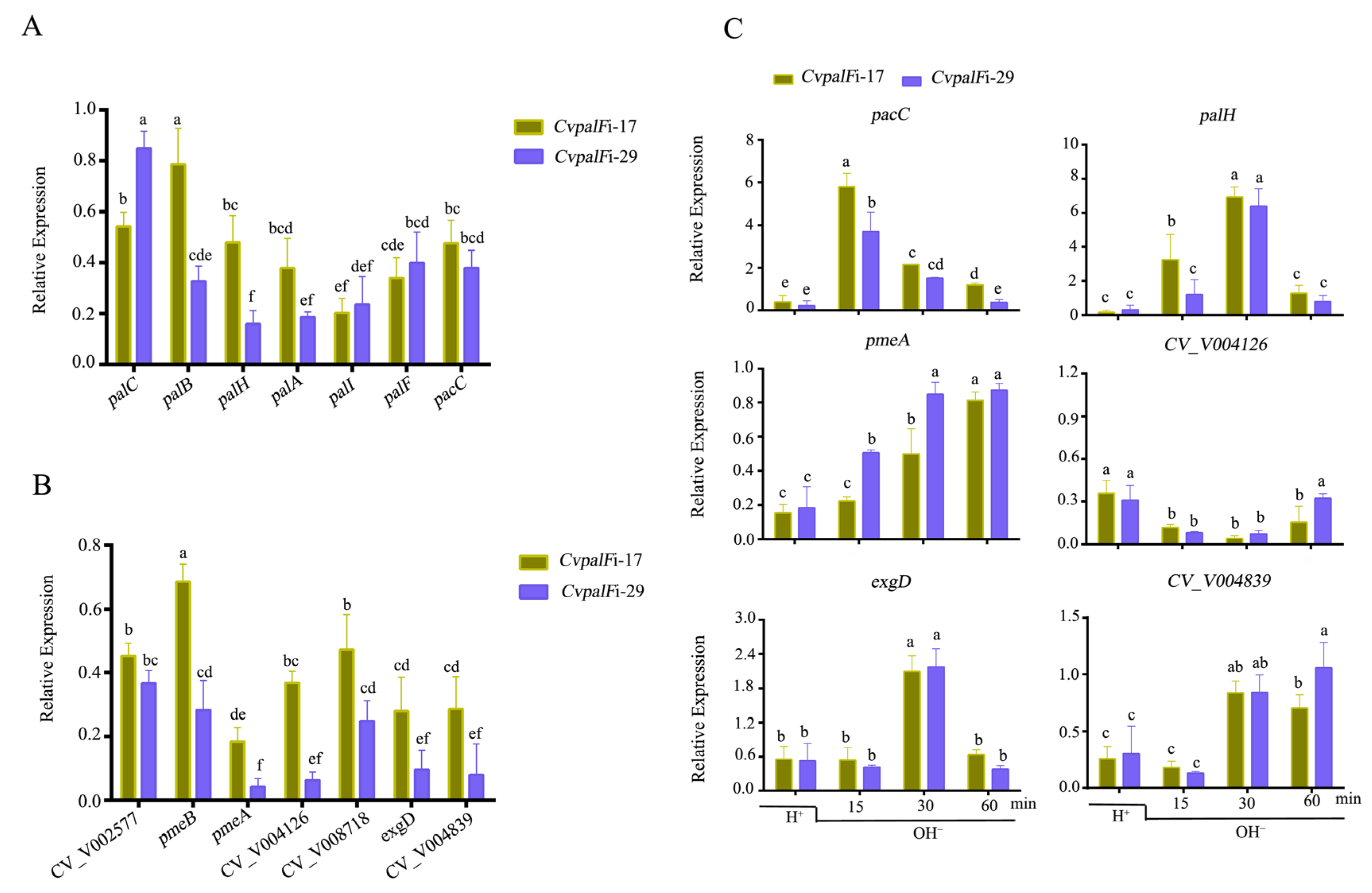
3.3.6. CvpalF Affected the Expression of Genes Related to Pal-Signaling Pathway and Plant Cell Wall-Degrading Enzyme
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, Y.; Yao, J.L.; Feng, H.; Jiang, J.; Fan, X.; Jia, Y.F.; Wang, R.; Liu, C. Identification of the defense-related gene VdWRKY53 from the wild grapevine Vitis davidii using RNA sequencing and ectopic expression analysis in Arabidopsis. Hereditas 2019, 156, 14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Liang, L.; Xie, Y.; Zhao, Z.; Su, L.; Tang, Y.; Sun, B.; Lai, Y.; Li, H. Transcriptome and metabolome analyses reveal molecular responses of two pepper (Capsicum annuum L.) cultivars to cold stress. Front. Plant Sci. 2022, 13, 819630. [Google Scholar] [CrossRef] [PubMed]
- Ji, T.; Languasco, L.; Li, M.; Rossi, V. Effects of temperature and wetness duration on infection by Coniella diplodiella, the fungus causing white rot of grape berries. Plants 2021, 10, 1696. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Li, B. Genome sequence resource of Coniella vitis, a fungal pathogen causing grape white rot disease. Mol. Plant-Microbe Interact. 2020, 33, 787–789. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Cui, C.; Jiang, J. First report of white rot of grape caused by Pilidiella castaneicola in China. Plant Dis. 2017, 101, 1673. [Google Scholar] [CrossRef]
- Qin, J.; Li, B.; Zhou, S. A novel glycoside hydrolase 74 xyloglucanase CvGH74A is a virulence factor in Coniella vitis. J. Integr. Agric. 2020, 19, 2725–2735. [Google Scholar] [CrossRef]
- Wang, Q.; Sun, J.; Wang, W.; Ren, W.; Li, B.; Zhou, S. Baseline sensitivity of Coniella vitis to fluazinam and resistance risk assessment. Plant Pathol. 2023, 72, 807–818. [Google Scholar] [CrossRef]
- Chethana, K.W.T.; Zhou, Y.; Zhang, W.; Liu, M.; Xing, Q.K.; Li, X.H.; Yan, J.Y.; Chethana, K.W.T.; Hyde, K.D. Coniella vitis sp. nov. is the common pathogen of white rot in Chinese vineyards. Plant Dis. 2017, 101, 2123–2136. [Google Scholar] [CrossRef]
- Yuan, L.; Jiang, H.; Liu, Q.; Jiang, X.; Wei, Y.; Yin, X.; Li, T. Acidic environment favors the development and pathogenicity of the grape white rot fungus Coniella vitis. J. Integr. Agric. 2024, in press. [Google Scholar] [CrossRef]
- Gordon, T.; Stueven, M.; Pastrana, A.M.; Dennehy, C.; Daugovish, O. The effect of pH on spore germination, growth, and infection of strawberry roots by Fusarium oxysporum f. sp. fragariae, cause of fusarium wilt of strawberry. Plant Dis. 2019, 103, 697–704. [Google Scholar] [CrossRef]
- Barad, S.; Horowitz, S.B.; Moskovitch, O.; Lichter, A.; Sherman, A.; Prusky, D.A. Penicillium expansum glucose oxidase-encoding gene, GOX2, is essential for gluconic acid production and acidification during colonization of deciduous fruit. Mol. Plant Microbe Interact. 2012, 25, 779–788. [Google Scholar] [CrossRef] [PubMed]
- Miyara, I.; Shafran, H.; Davidzon, M.; Sherman, A.; Prusky, D. pH regulation of ammonia secretion by Colletotrichum gloeosporioides and its effect on appressorium formation and pathogenicity. Mol. Plant Microbe Interact. 2010, 23, 304–316. [Google Scholar] [CrossRef] [PubMed]
- Manteau, S.; Abouna, S.; Lambert, B.; Legendre, L. Differential regulation by ambient pH of putative virulence factor secretion by the phytopathogenic fungus Botrytis cinerea. FEMS Microbiol. Ecol. 2003, 43, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Chen, Y.; Tian, S. Function of pH-dependent transcription factor PacC in regulating development, pathogenicity, and mycotoxin biosynthesis of phytopathogenic fungi. FEBS J. 2022, 289, 1723–1730. [Google Scholar] [CrossRef] [PubMed]
- Penalva, M.A.; Tilburn, J.; Bignell, E.; Arst, H.N. Ambient pH gene regulation in fungi: Making connections. Trends Microbiol. 2008, 16, 291–300. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, B.; Xu, X.; Zhang, Z.; Tian, S. The pH-responsive PacC transcription factor plays pivotal roles in virulence and patulin biosynthesis in Penicillium expansum. Environ. Microbiol. 2018, 20, 4063–4078. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Han, Z.; Gong, D.; Xu, X.; Li, Y.; Sionov, E.; Prusky, D.; Bi, Y.; Zong, Y. The pH signalling transcription factor PacC modulate growth, development, stress response and pathogenicity of Trichothecium roseum. Environ. Microbiol. 2022, 24, 1608–1621. [Google Scholar] [CrossRef] [PubMed]
- Herranz, S.; Rodríguez, J.M.; Bussink, H.J.; Sánchez-Ferrero, J.C.; Arst, H.N., Jr.; Peñalva, M.A.; Vincent, O. Arrestin-related proteins mediate pH signaling in fungi. Proc. Natl. Acad. Sci. USA 2005, 102, 12141–12146. [Google Scholar] [CrossRef] [PubMed]
- Hervás-Aguilar, A.; Galindo, A.; Peñalva, M.A. Receptor-independent Ambient pH signaling by ubiquitin attachment to fungal arrestin-like PalF. J. Biol. Chem. 2010, 285, 18095–18102. [Google Scholar] [CrossRef]
- Selvig, K.; Alspaugh, J.A. pH Response Pathways in Fungi: Adapting to host-derived and environmental signals. Mycobiology 2011, 39, 249–256. [Google Scholar] [CrossRef]
- Li, L.; Hao, B.; Zhang, Y.; Ji, S.; Chou, G. Metabolite profiling and distribution of militarine in rats using UPLC-Q-TOF-MS/MS. Molecules 2020, 25, 1082. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Fang, T.; Xu, H.; Jiang, L. The pH-sensing Rim101 pathway positively regulates the transcriptional expression of the calcium pump gene PMR1 to affect calcium sensitivity in budding yeast. Biochem. Biophys. Res. Commun. 2020, 532, 453–458. [Google Scholar] [CrossRef]
- Peñalva, M.A.; Lucena-Agell, D.; Arst, H.N., Jr. Liaison alcaline: Pals entice non-endosomal ESCRTs to the plasma membrane for pH signaling. Curr. Opin. Microbiol. 2014, 22, 49–59. [Google Scholar] [CrossRef]
- Gomez-Raja, J.; Davis, D.A. The β-arrestin-like protein Rim8 is hyperphosphorylated and complexes with Rim21 and Rim101 to promote adaptation to neutral-alkaline pH. Eukaryot. Cell 2012, 11, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Zou, M.; Xin, B.; Sun, X.; Lin, R.; Lu, J.; Qi, J.; Xie, B.; Cheng, X. URA3 as a Selectable Marker for Disruption and Functional Assessment of PacC Gene in the Entomopathogenic Fungus Isaria javanica. J. Fungi 2023, 9, 92. [Google Scholar] [CrossRef]
- Gu, Q.; Wang, Y.; Zhao, X.; Yuan, B.; Zhang, M.; Tan, Z.; Zhang, X.; Chen, Y.; Wu, H.; Luo, Y.; et al. Inhibition of histone acetyltransferase GCN5 by a transcription factor FgPacC controls fungal adaption to host-derived iron stress. Nucleic Acids Res. 2022, 50, 6190–6210. [Google Scholar] [CrossRef] [PubMed]
- Yun, Y.; Liu, Z.; Zhang, J.; Shim, W.B.; Chen, Y.; Ma, Z. The MAPKK FgMkk1 of Fusarium graminearum regulates vegetative differentiation, multiple stress response, and virulence via the cell wall integrity and high-osmolarity glycerol signaling pathways. Environ. Microbiol. 2014, 16, 2023–2037. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Hervás-Aguilar, A.; Rodríguez, J.M.; Tilburn, J.; Arst, H.N., Jr.; Peñalva, M.A. Evidence for the direct involvement of the proteasome in the proteolytic processing of the Aspergillus nidulans zinc finger transcription factor PacC. J. Biol. Chem. 2007, 282, 34735–34747. [Google Scholar] [CrossRef]
- Mushtaq, S.; Tayyeb, A.; Firdaus-E-Bareen. A comparison of total RNA extraction methods for RT-PCR based differential expression of genes from Trichoderma atrobrunneum. J. Microbiol. Methods. 2022, 200, 106535. [Google Scholar] [CrossRef]
- Kwak, S.K.; Kim, J. Transparency considerations for describing statistical analyses in research. Korean J. Anesthesiol. 2021, 74, 488–495. [Google Scholar] [CrossRef] [PubMed]
- Wei, K.; Zheng, Y.; Li, J.; Shen, F.; Zou, Z.; Fan, H.; Li, X.; Wu, C.; Yao, M. Microbial aerosol character istics in highly polluted and near-pristine environments featuring different climatic conditions. Sci. Bull. 2015, 60, 1439–1447. [Google Scholar] [CrossRef]
- Li, D.; Wan, Y.; Wang, Y.; He, P. Relatedness of resistance to anthracnose and to white rot in Chinese wild grapes. Vitis 2008, 47, 213–215. [Google Scholar]
- Liu, R.; Wang, Y.; Li, P.; Sun, L.; Jiang, J.; Fan, X.; Liu, C.; Zhang, Y. Genome assembly and transcriptome analysis of the fungus Coniella diplodiella during infection on grapevine (Vitis vinifera L.). Front. Microbiol. 2021, 11, 599150. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Li, T.; Wei, Y.; Liu, Q.; Jiang, X.; Yuan, L. First report of Coniella vitis causing white rot on Virginia creeper (Parthenocissus quinquefolia [L.] Planch.) in China. Plant Dis. 2023, 107, 1244. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Yin, X.; Jiang, X.; Li, T.; Wei, Y. Identification of the pathogen Coniella vitis causing grape white rot in Shandong province. Acta Phytopathol. Sin. 2023, 53, 729–733. [Google Scholar]
- Fitzgerald, A.; Van Kan, J.A.; Plummer, K.M. Simultaneous silencing of multiple genes in the apple scab fungus, Venturia inaequalis, by expression of RNA with chimeric inverted repeats. Fungal Genet. Biol. 2004, 41, 963–971. [Google Scholar] [CrossRef] [PubMed]
- Hammond, T.M.; Keller, N.P. RNA silencing in Aspergillus nidulans is independent of RNA-dependent RNA polymerases. Genetics 2005, 169, 607–617. [Google Scholar] [CrossRef] [PubMed]
- Kadotani, N.; Nakayashiki, H.; Tosa, Y.; Mayama, S. RNA silencing in the phytopathogenic fungus Magnaporthe oryzae. Mol. Plant Microbe Interact. 2003, 16, 769–776. [Google Scholar] [CrossRef]
- Prusky, D.; Barad, S.; Ment, D.; Bi, F. The pH modulation by fungal secreted molecules: A mechanism affecting pathogenicity by postharvest pathogens. Isr. J. Plant Sci. 2016, 63, 22–30. [Google Scholar] [CrossRef]
- Li, H.; Chen, Y.; Zhang, Z.; Li, B.; Qin, G.; Tian, S. Pathogenic mechanisms and control strategies of Botrytis cinerea causing post-harvest decay in fruits and vegetables. Food Qual. Saf. 2018, 2, 111–119. [Google Scholar] [CrossRef]
- Rascle, C.; Dieryckx, C.; Dupuy, J.W.; Muszkieta, L.; Souibgui, E.; Droux, M.; Bruel, C.; Girard, V.; Poussereau, N. The pH regulator PacC: A host-dependent virulence factor in Botrytis cinerea. Environ. Microbiol. Rep. 2018, 10, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Ying, S.H.; Feng, M.G. The Pal pathway required for ambient pH adaptation regulates growth, conidiation, and osmotolerance of Beauveria bassiana in a pH-dependent manner. Appl. Microbiol. Biotechnol. 2016, 100, 4423–4433. [Google Scholar] [CrossRef]
- Cervantes-Chávez, J.A.; Oetiz-Castellanos, L.; Tejeda-Sartorius, M.; Gold, S.; Ruiz-Herrera, J. Functional analysis of the pH responsive pathway Pal/Rim in the phytopathogenic basidiomycete Ustilago maydis. Fungal Genet. Biol. 2010, 47, 446–457. [Google Scholar]
- Wu, F.L.; Zhang, G.; Ren, A.; Dang, Z.H.; Shi, L.; Jiang, A.L.; Zhao, M.W. The pH-responsive transcription factor PacC regulates mycelial growth, fruiting body development, and ganoderic acid biosynthesis in Ganoderma lucidum. Mycologia 2016, 108, 1104–1113. [Google Scholar] [CrossRef] [PubMed]
- Zuriegat, Q.; Zheng, Y.; Liu, H.; Wang, Z.; Yun, Y. Current progress on pathogenicity-related transcription factors in Fusarium oxysporum. Mol. Plant Pathol. 2021, 22, 882–895. [Google Scholar] [CrossRef] [PubMed]
- O’Meara, T.R.; Norton, D.; Price, M.S.; Hay, C.; Clements, M.F.; Nichols, C.B.; Alspaugh, J.A. Interaction of Cryptococcus neoformans rim101 and protein kinase a regulates capsule. PLoS Pathog. 2010, 6, e1000776. [Google Scholar] [CrossRef]
- Torres, M.A.; Jones, J.D.; Dangl, J.L. Reactive Oxygen Species Signaling in Response to Pathogens. Plant Physiol. 2006, 141, 373–378. [Google Scholar] [CrossRef]
- Li, Z.; Xu, X.; Leng, X.; He, M.; Wang, J.; Cheng, S.; Wu, H. Roles of reactive oxygen species in cell signaling pathways and immune responses to viral infections. Arch. Virol. 2017, 162, 603–610. [Google Scholar] [CrossRef]
- Wan, J.; He, M.; Hou, Q.; Zou, L.; Yang, Y.; Wei, Y.; Chen, X. Cell wall associated immunity in plants. Stress Biol. 2021, 1, 3. [Google Scholar] [CrossRef]
- Tomassini, A.; Sella, L.; Raiola, A.; D’Ovidio, R.; Favaron, F. Characterization and expression of Fusarium graminearum endo- polygalacturonases in vitro and during wheat infection. Plant Pathol. 2009, 58, 556–564. [Google Scholar] [CrossRef]
- Fonseca-García, C.; León-Ramírez, C.G.; Ruiz-Herrera, J. The regulation of different metabolic pathways through the Pal/Rim pathway in Ustilago maydis. FEMS Yeast Res. 2012, 12, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Valette-Collet, O.; Cimerman, A.; Reignault, P.; Levis, C.; Boccara, M. Disruption of Botrytis cinerea pectin methylesterase gene Bcpme1 reduces virulence on several host plants. Mol. Plant-Microbe Interact. 2003, 16, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Sella, L.; Castiglioni, C.; Paccanaro, M.C.; Janni, M.; Schäfer, W.; D’Ovidio, R.; Favaron, F. Involvement of Fungal Pectin Methylesterase Activity in the Interaction Between Fusarium graminearum and Wheat. Mol. Plant Microbe Interact. 2016, 29, 258–267. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) | Fragments (bp) |
|---|---|---|
| palF1 | AGCATCGATACCGTCGACCACCGTTTGGAGGTTATTGTGGAC | 485 |
| palF2 | AGCAAGCTTGTACGTACGAACTGCGGAGGAGGTATGTAGT | |
| palF3 | TAAGTGGATCCGGGGCCCAGACCGTTTGGAGGTTATTGTGGAC | 485 |
| palF4 | TCGCATGCTAAGGCCTGTGAACTGCGGAGGAGGTATGTAGT | |
| LFcheck 1F | ATGAGCAAGCGGACGGAGTG | 1072 |
| LFcheck 1R | CTTGTCAGTCCCTTCCATTTATTT | |
| LFcheck 2F | ACACACAGCCAGGGAACGG | 1164 |
| LFcheck 2R | GGAGCATTCACTAGGCAACCA | |
| Fhyg | GTCCTGCGGGTAAATAGCTG | 750 |
| Rhyg | ATTTGTGTACGCCCGACAGT | |
| palF-RTF | GGGTTCCGGGAGAAATGATTAG | 102 |
| palF-RTR | GTCTTGCATTCTGTCCGATTTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, X.; Chen, Z.; Li, T.; Liu, Q.; Jiang, X.; Han, X.; Wang, C.; Wei, Y.; Yuan, L. The Arrestin-like Protein palF Contributes to Growth, Sporulation, Spore Germination, Osmolarity, and Pathogenicity of Coniella vitis. J. Fungi 2024, 10, 508. https://doi.org/10.3390/jof10070508
Yin X, Chen Z, Li T, Liu Q, Jiang X, Han X, Wang C, Wei Y, Yuan L. The Arrestin-like Protein palF Contributes to Growth, Sporulation, Spore Germination, Osmolarity, and Pathogenicity of Coniella vitis. Journal of Fungi. 2024; 10(7):508. https://doi.org/10.3390/jof10070508
Chicago/Turabian StyleYin, Xiangtian, Zihe Chen, Tinggang Li, Qibao Liu, Xilong Jiang, Xing Han, Chundong Wang, Yanfeng Wei, and Lifang Yuan. 2024. "The Arrestin-like Protein palF Contributes to Growth, Sporulation, Spore Germination, Osmolarity, and Pathogenicity of Coniella vitis" Journal of Fungi 10, no. 7: 508. https://doi.org/10.3390/jof10070508