Differences in Sirtuin Regulation in Response to Calorie Restriction in Cryptococcus neoformans

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Strains
2.3. Growth Curves
2.4. Replicative and Chronological Lifespan
2.5. Disruption and Complementation of the SIR2 Gene
2.6. Phenotypic Characterization
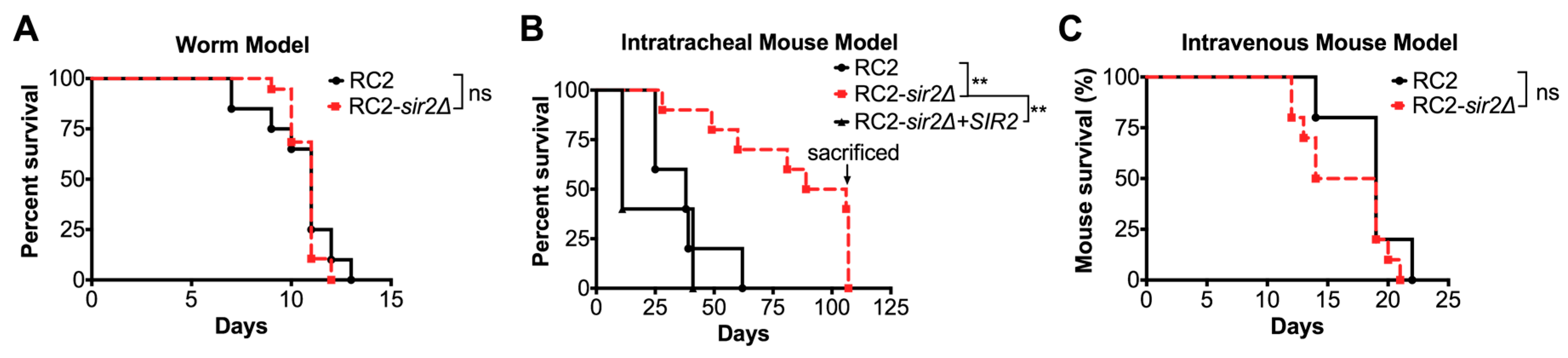
2.7. Infection Studies
2.8. Transcriptome Analysis
2.9. Statistics
3. Results
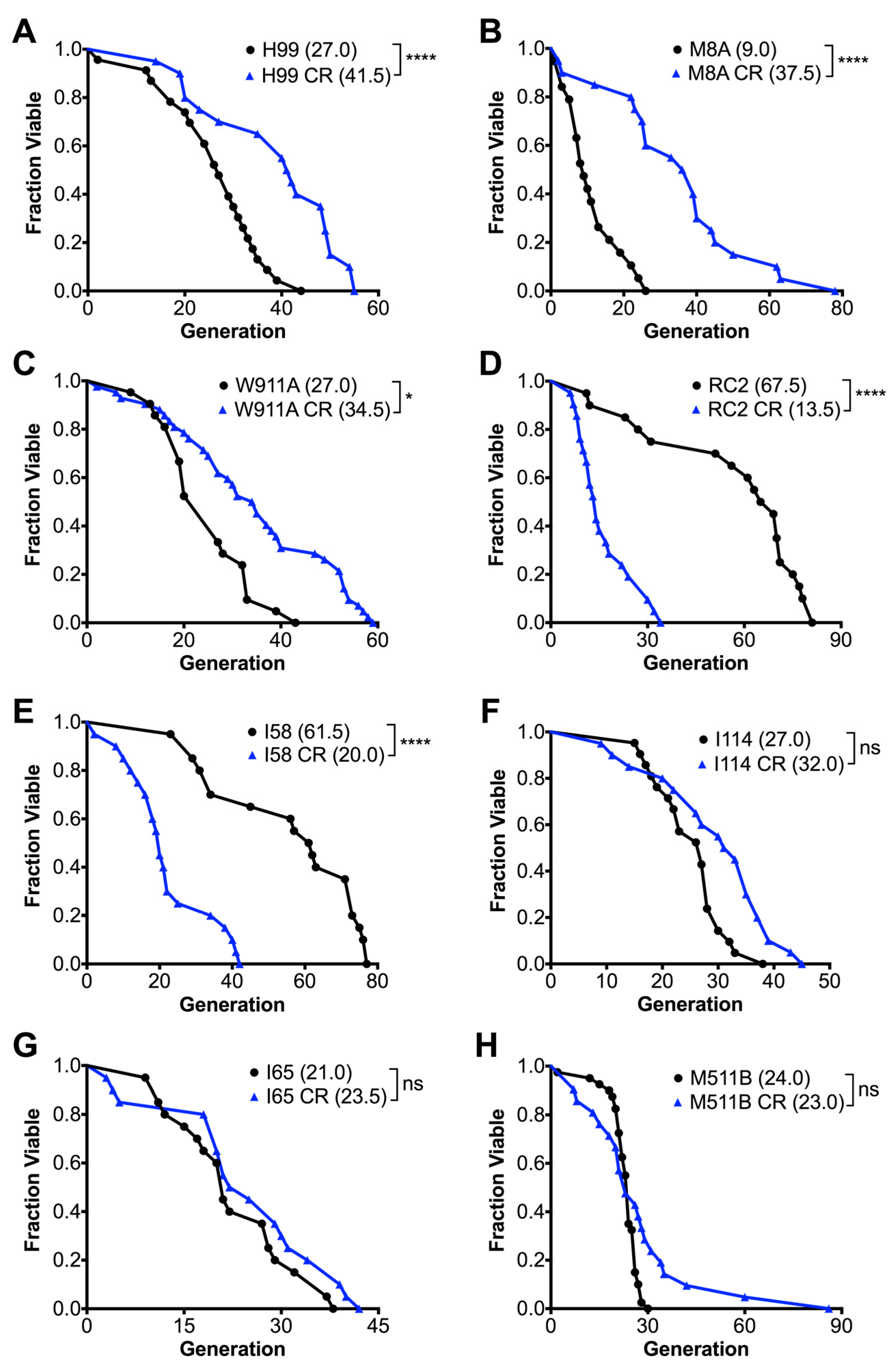
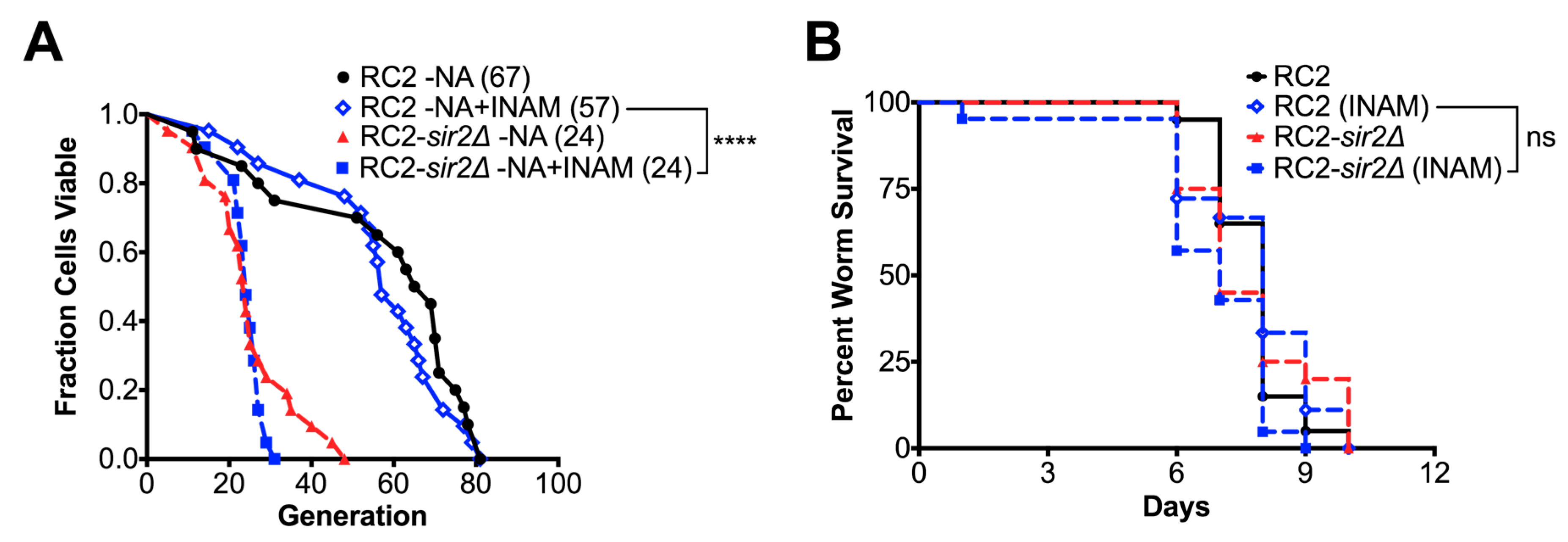
3.1. Calorie Restriction Variably Affects the Replicative Lifespan of C. neoformans
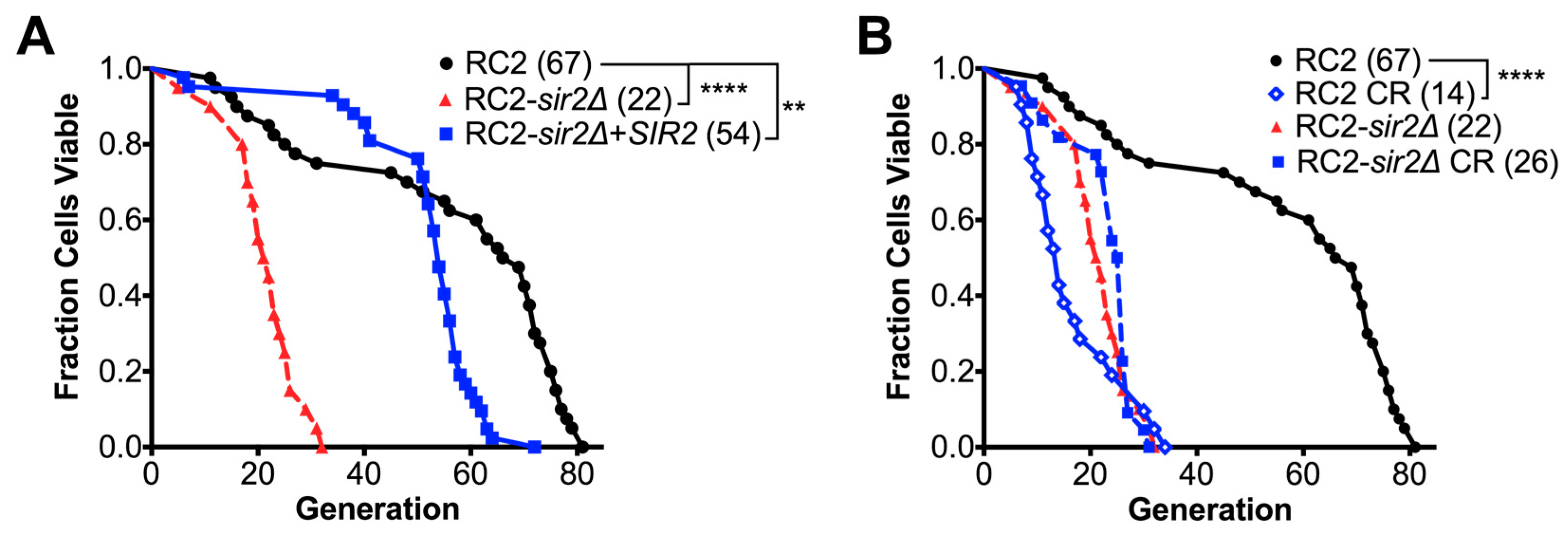
3.2. Loss of SIR2 Has a Significant Effect on the Phenotype of Strain RC2
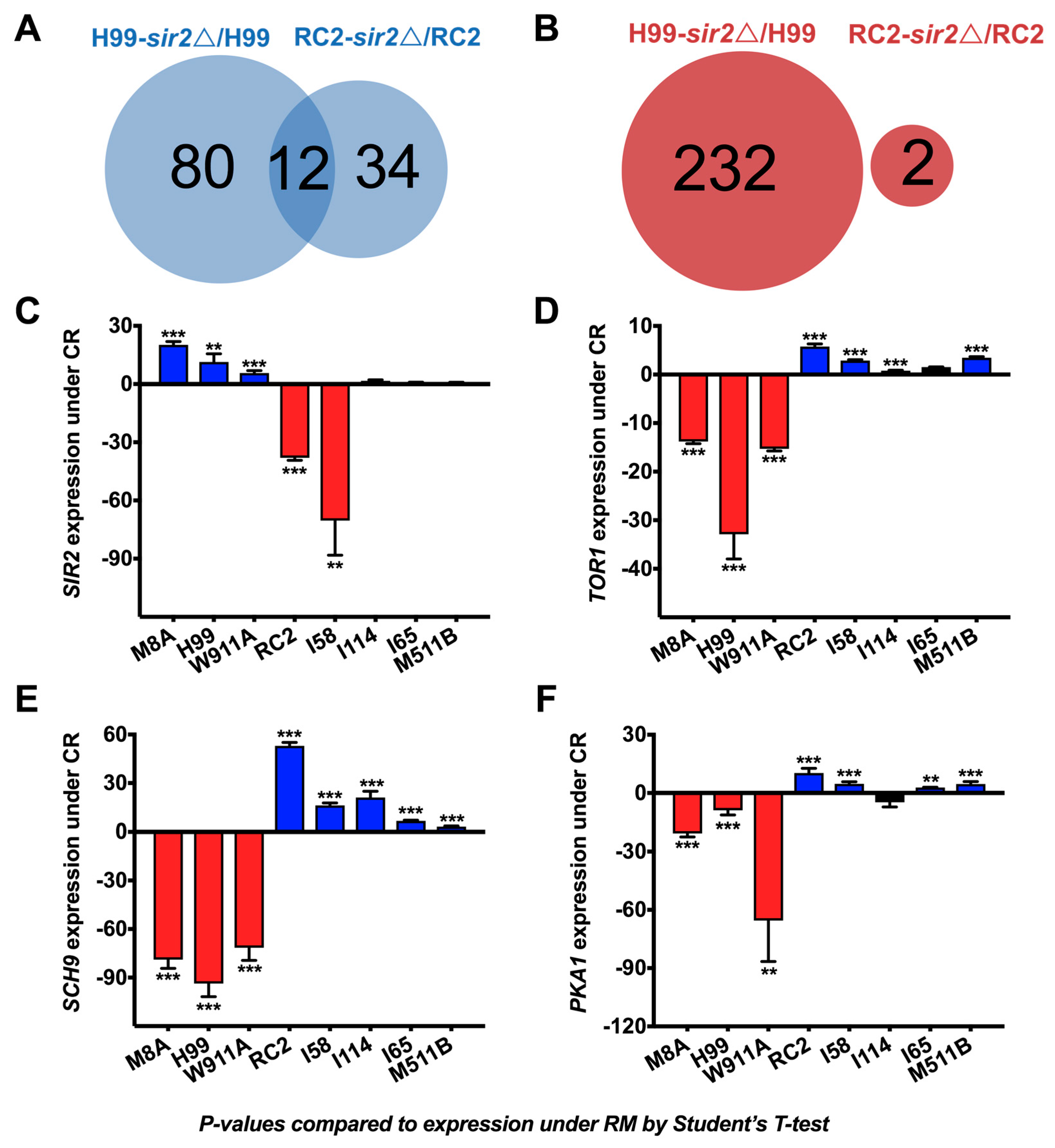
3.3. Gene Regulation of C. neoformans under Calorie Restriction
3.4. SIR2 Is Differentially Regulated in RC2
4. Discussion
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Perfect, J.R.; Casadevall, A. Cryptococcosis. Infect. Dis. Clin. North Am. 2002, 16, 837–874. [Google Scholar] [CrossRef]
- Rajasingham, R.; Smith, R.M.; Park, B.J.; Jarvis, J.N.; Govender, N.P.; Chiller, T.M.; Denning, D.W.; Loyse, A.; Boulware, D.R. Global burden of disease of HIV-associated cryptococcal meningitis: An updated analysis. Lancet Infect. Dis. 2017, 17, 873–881. [Google Scholar] [CrossRef]
- Alanio, A.; Desnos-Ollivier, M.; Dromer, F. Dynamics of Cryptococcus neoformans-macrophage interactions reveal that fungal background influences outcome during cryptococcal meningoencephalitis in humans. MBio 2011, 2, 41–43. [Google Scholar] [CrossRef] [PubMed]
- Steinkraus, K.A.; Kaeberlein, M.; Kennedy, B.K. Replicative aging in yeast: The means to the end. Annu. Rev. Cell Dev. Bi. 2008, 24, 29–54. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Dungrawala, H.; Hua, H.; Manukyan, A.; Abraham, L.; Lane, W.; Mead, H.; Wright, J.; Schneider, B.L. Cell size and growth rate are major determinants of replicative lifespan. Cell Cycle 2011, 10, 144–155. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.H.; Meng, F.L.; Hu, Y.; Zhou, J.Q. Candida albicans, a distinctive fungal model for cellular aging study. Aging Cell 2008, 7, 746–757. [Google Scholar] [CrossRef] [PubMed]
- Roux, A.E.; Chartrand, P.; Ferbeyre, G.; Rokeach, L.A. Fission yeast and other yeasts as emergent models to unravel cellular aging in eukaryotes. J. Gerontol. A Biol. Sci. Med. Sci. 2010, 65, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bouklas, T.; Pechuan, X.; Goldman, D.L.; Edelman, B.; Bergman, A.; Fries, B.C. Old Cryptococcus neoformans cells contribute to virulence in chronic cryptococcosis. MBio 2013, 4, e00455-13. [Google Scholar] [CrossRef] [PubMed]
- Jain, N.; Cook, E.; Xess, I.; Hasan, F.; Fries, D.; Fries, B.C. Isolation and characterization of senescent Cryptococcus neoformans and implications for phenotypic switching and pathogenesis in chronic cryptococcosis. Eukaryot. Cell 2009, 8, 858–866. [Google Scholar] [CrossRef] [PubMed]
- Greiss, S.; Gartner, A. Sirtuin/Sir2 phylogeny, evolutionary considerations and structural conservation. Mol. Cells 2009, 28, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Kaeberlein, M.; McVey, M.; Guarente, L. The Sir2/3/4 complex and Sir2 alone promote longevity in saccharomyces cerevisiae by two different mechanisms. Genes Dev. 1999, 13, 2570–2580. [Google Scholar] [CrossRef] [PubMed]
- Guarente, L. Calorie restriction and sirtuins revisited. Genes. Dev. 2013, 27, 2072–2085. [Google Scholar] [CrossRef] [PubMed]
- Kaeberlein, M.; Kirkland, K.T.; Fields, S.; Kennedy, B.K. Sir2-independent life span extension by calorie restriction in yeast. PLoS Biol. 2004, 2, E296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouklas, T.; Jain, N.; Fries, B.C. Modulation of replicative lifespan in Cryptococcus neoformans: Implications for virulence. Front Microbiol. 2017, 8, 98. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.J.; Defossez, P.A.; Guarente, L. Requirement of nad and Sir2 for life-span extension by calorie restriction in saccharomyces cerevisiae. Science 2000, 289, 2126–2128. [Google Scholar] [CrossRef] [PubMed]
- Fabrizio, P.; Pletcher, S.D.; Minois, N.; Vaupel, J.W.; Longo, V.D. Chronological aging-independent replicative life span regulation by Msn2/Msn4 and Sod2 in saccharomyces cerevisiae. FEBS Lett. 2004, 557, 136–142. [Google Scholar] [CrossRef]
- Schmelzle, T.; Hall, M.N. Tor, a central controller of cell growth. Cell 2000, 103, 253–262. [Google Scholar] [CrossRef]
- McClure, J.M.; Wierman, M.B.; Maqani, N.; Smith, J.S. Isonicotinamide enhances Sir2 protein-mediated silencing and longevity in yeast by raising intracellular NAD+ concentration. J. Biol. Chem. 2012, 287, 20957–20966. [Google Scholar] [CrossRef] [PubMed]
- Jain, N.; Li, L.; Hsueh, Y.P.; Guerrero, A.; Heitman, J.; Goldman, D.L.; Fries, B.C. Loss of allergen 1 confers a hypervirulent phenotype that resembles mucoid switch variants of Cryptococcus neoformans. Infect. Immun. 2009, 77, 128–140. [Google Scholar] [CrossRef] [PubMed]
- Park, P.U.; McVey, M.; Guarente, L. Separation of mother and daughter cells. Method. Enzymol. 2002, 351, 468–477. [Google Scholar]
- Murakami, C.; Kaeberlein, M. Quantifying yeast chronological life span by outgrowth of aged cells. JOVE-J. Vis. Exp. 2009, 27, 3044–3054. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A.; Cleare, W.; Feldmesser, M.; Glatman-Freedman, A.; Goldman, D.L.; Kozel, T.R.; Lendvai, N.; Mukherjee, J.; Pirofski, L.A.; Rivera, J.; et al. Characterization of a murine monoclonal antibody to Cryptococcus neoformans polysaccharide that is a candidate for human therapeutic studies. Antimicrob. Agents Ch. 1998, 42, 1437–1446. [Google Scholar]
- Arras, S.D.M.; Chitty, J.L.; Wizrah, M.S.I.; Erpf, P.E.; Schulz, B.L.; Tanurdzic, M.; Fraser, J.A. Sirtuins in the phylum basidiomycota: A role in virulence in Cryptococcus neoformans. Sci. Rep. 2017, 7, 46567. [Google Scholar] [CrossRef] [PubMed]
- Cotter, G.; Doyle, S.; Kavanagh, K. Development of an insect model for the in vivo pathogenicity testing of yeasts. FEMS Immunol. Med. Microbiol. 2000, 27, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Skinner, C.; Lin, S.J. Effects of calorie restriction on life span of microorganisms. Appl. Microbiol. Biotechnol. 2010, 88, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Ene, I.V.; Brunke, S.; Brown, A.J.; Hube, B. Metabolism in fungal pathogenesis. Cold Spring Harb. Perspect. Med. 2014, 4, a019695. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.J.; Kaeberlein, M.; Andalis, A.A.; Sturtz, L.A.; Defossez, P.A.; Culotta, V.C.; Fink, G.R.; Guarente, L. Calorie restriction extends saccharomyces cerevisiae lifespan by increasing respiration. Nature 2002, 418, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Kaeberlein, M.; Powers, R.W., 3rd; Steffen, K.K.; Westman, E.A.; Hu, D.; Dang, N.; Kerr, E.O.; Kirkland, K.T.; Fields, S.; Kennedy, B.K. Regulation of yeast replicative life span by TOR and SCH9 in response to nutrients. Science 2005, 310, 1193–1196. [Google Scholar] [CrossRef] [PubMed]
- Franzot, S.P.; Mukherjee, J.; Cherniak, R.; Chen, L.C.; Hamdan, J.S.; Casadevall, A. Microevolution of a standard strain of Cryptococcus neoformans resulting in differences in virulence and other phenotypes. Infect. Immun. 1998, 66, 89–97. [Google Scholar] [PubMed]
- Fries, B.C.; Taborda, C.P.; Serfass, E.; Casadevall, A. Phenotypic switching of Cryptococcus neoformans occurs in vivo and influences the outcome of infection. J. Clin. Invest. 2001, 108, 1639–1648. [Google Scholar] [CrossRef] [PubMed]
- Goldman, D.L.; Fries, B.C.; Franzot, S.P.; Montella, L.; Casadevall, A. Phenotypic switching in the human pathogenic fungus Cryptococcus neoformans is associated with changes in virulence and pulmonary inflammatory response in rodents. Proc. Natl. Acad. Sci. U S A 1998, 95, 14967–14972. [Google Scholar] [CrossRef] [PubMed]
- Fries, B.C.; Goldman, D.L.; Cherniak, R.; Ju, R.; Casadevall, A. Phenotypic switching in Cryptococcus neoformans results in changes in cellular morphology and glucuronoxylomannan structure. Infect. Immun. 1999, 67, 6076–6083. [Google Scholar] [PubMed]
- Bouklas, T.; Alonso-Crisostomo, L.; Szekely, T., Jr.; Diago-Navarro, E.; Orner, E.P.; Smith, K.; Munshi, M.A.; Del Poeta, M.; Balazsi, G.; Fries, B.C. Generational distribution of a candida glabrata population: Resilient old cells prevail, while younger cells dominate in the vulnerable host. PLoS Pathog. 2017, 13, e1006355. [Google Scholar] [CrossRef] [PubMed]
- Zurita-Martinez, S.A.; Cardenas, M.E. Tor and cyclic amp-protein kinase a: Two parallel pathways regulating expression of genes required for cell growth. Eukaryot. Cell 2005, 4, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Martin, D.E.; Soulard, A.; Hall, M.N. Tor regulates ribosomal protein gene expression via PKA and the forkhead transcription factor FHL1. Cell 2004, 119, 969–979. [Google Scholar] [CrossRef] [PubMed]
- Pedruzzi, I.; Dubouloz, F.; Cameroni, E.; Wanke, V.; Roosen, J.; Winderickx, J.; De Virgilio, C. TOR and PKA signaling pathways converge on the protein kinase Rim15 to control entry into G0. Mol. Cell 2003, 12, 1607–1613. [Google Scholar] [CrossRef]
- Helliwell, S.B.; Howald, I.; Barbet, N.; Hall, M.N. TOR2 is part of two related signaling pathways coordinating cell growth in saccharomyces cerevisiae. Genetics 1998, 148, 99–112. [Google Scholar] [PubMed]
- Beck, T.; Hall, M.N. The tor signalling pathway controls nuclear localization of nutrient-regulated transcription factors. Nature 1999, 402, 689–692. [Google Scholar] [CrossRef] [PubMed]
- Davidson, R.C.; Blankenship, J.R.; Kraus, P.R.; de Jesus Berrios, M.; Hull, C.M.; D'Souza, C.; Wang, P.; Heitman, J. A PCR-based strategy to generate integrative targeting alleles with large regions of homology. Microbiology 2002, 148, 2607–2615. [Google Scholar] [CrossRef] [PubMed]
- Jorgensen, P.; Rupes, I.; Sharom, J.R.; Schneper, L.; Broach, J.R.; Tyers, M. A dynamic transcriptional network communicates growth potential to ribosome synthesis and critical cell size. Genes Dev. 2004, 18, 2491–2505. [Google Scholar] [CrossRef] [PubMed]
- Shamji, A.F.; Kuruvilla, F.G.; Schreiber, S.L. Partitioning the transcriptional program induced by rapamycin among the effectors of the tor proteins. Curr. Biol. 2000, 10, 1574–1581. [Google Scholar] [CrossRef]
- Fuchs, B.B.; Tang, R.J.; Mylonakis, E. The temperature-sensitive role of Cryptococcus neoformans ROM2 in cell morphogenesis. PLoS One 2007, 2, e368. [Google Scholar] [CrossRef] [PubMed]
- Varma, A.; Wu, S.; Guo, N.; Liao, W.; Lu, G.; Li, A.; Hu, Y.; Bulmer, G.; Kwon-Chung, K.J. Identification of a novel gene, URE2, that functionally complements a urease-negative clinical strain of Cryptococcus neoformans. Microbiology 2006, 152, 3723–3731. [Google Scholar] [CrossRef] [PubMed]
- Stajich, J.E.; Dietrich, F.S. Evidence of mRNA-mediated intron loss in the human-pathogenic fungus Cryptococcus neoformans. Eukaryot. Cell 2006, 5, 789–793. [Google Scholar] [CrossRef] [PubMed]
- Williams, V.; Del Poeta, M. Role of glucose in the expression of Cryptococcus neoformans antiphagocytic protein 1, app1. Eukaryot. Cell 2011, 10, 293–301. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Cheng, P.Y.; Sham, A.; Perfect, J.R.; Kronstad, J.W. Metabolic adaptation in Cryptococcus neoformans during early murine pulmonary infection. Mol. Microbiol. 2008, 69, 1456–1475. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.; Price, M.S.; Toffaletti, D.L.; Tenor, J.; Betancourt-Quiroz, M.; Price, J.L.; Pan, W.H.; Liao, W.Q.; Perfect, J.R. Pleiotropic effects of deubiquitinating enzyme ubp5 on growth and pathogenesis of Cryptococcus neoformans. PLoS One 2012, 7, e38326. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.C.; Bien, C.M.; Lee, H.; Espenshade, P.J.; Kwon-Chung, K.J. Sre1p, a regulator of oxygen sensing and sterol homeostasis, is required for virulence in Cryptococcus neoformans. Mol. Microbiol. 2007, 64, 614–629. [Google Scholar] [CrossRef] [PubMed]
- Cramer, K.L.; Gerrald, Q.D.; Nichols, C.B.; Price, M.S.; Alspaugh, J.A. Transcription factor nrg1 mediates capsule formation, stress response, and pathogenesis in Cryptococcus neoformans. Eukaryot. Cell 2006, 5, 1147–1156. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Doubling Time in RM (h) | Doubling Time in CR (h) | P-Value |
|---|---|---|---|
| H99 | 2.1 ± 0.3 | 2.8 ± 0.2 | 0.01 |
| M511B | 3.1 ± 0.2 | 3.6 ± 0.3 | 0.01 |
| M8A | 2.4 ± 0.3 | 2.2 ± 0.2 | ns |
| I58 | 2.5 ± 0.3 | 2.9 ± 0.4 | ns |
| I65 | 2.6 ± 0.2 | 3.0 ± 0.3 | 0.05 |
| I114 | 2.3 ± 0.1 | 4.0 ± 0.2 | 0.01 |
| RC2 | 2.4 ± 0.2 | 3.0 ± 0.2 | 0.01 |
| W911A | 3.4 ± 0.3 | 3.6 ± 0.3 | ns |
| Phenotype | WT | Mutant | p-Value |
|---|---|---|---|
| Doubling time in YPD | 2.0 ± 0.2 h | 2.8 ± 0.3 h | < 0.01 |
| Doubling time in 0.05% YPD | 3.0 ± 0.2 h | 3.4 ± 0.4 h | ns |
| Mating in V8 agar | No with Kn99MATa | No with Kn99MATa | ns |
| Chronological lifespan | 19 d | 21 d | ns |
| Phenotypic switching rate | 0.5 × 10−4 | 0.5 × 10−4 | ns |
| Uninduced capsule size | 1.48 ± 0.34 µm | 2.19 ± 0.28 µm | < 0.01 |
| Induced capsule size | 5.89 ± 1.29 µm | 5.65 ± 0.92 µm | ns |
| Total cell size | 7.25 ± 1.09 µm | 9.16 ± 0.83 µm | < 0.01 |
| Phagocytosis index | 20.32% | 31.67% | ns |
| Killing in macrophages | 59.06% | 69.36% | ns |
| Colony sectoring | No sectoring | No sectoring | ns |
| Melanization | Same degree | Same degree | ns |
| GXM stain (18B7 mAb) | Same pattern | Same pattern | ns |
| MIC to amphotericin B | 0.125 µg/mL | 0.125 µg/mL | ns |
| H2O2 resistance | 3.1 cm | 3.0 cm | ns |
| 39 °C growth | Grows | Grows | ns |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bouklas, T.; Masone, L.; Fries, B.C. Differences in Sirtuin Regulation in Response to Calorie Restriction in Cryptococcus neoformans. J. Fungi 2018, 4, 26. https://doi.org/10.3390/jof4010026
Bouklas T, Masone L, Fries BC. Differences in Sirtuin Regulation in Response to Calorie Restriction in Cryptococcus neoformans. Journal of Fungi. 2018; 4(1):26. https://doi.org/10.3390/jof4010026
Chicago/Turabian StyleBouklas, Tejas, Lindsey Masone, and Bettina C. Fries. 2018. "Differences in Sirtuin Regulation in Response to Calorie Restriction in Cryptococcus neoformans" Journal of Fungi 4, no. 1: 26. https://doi.org/10.3390/jof4010026
APA StyleBouklas, T., Masone, L., & Fries, B. C. (2018). Differences in Sirtuin Regulation in Response to Calorie Restriction in Cryptococcus neoformans. Journal of Fungi, 4(1), 26. https://doi.org/10.3390/jof4010026