Pathogenicity Levels of Colombian Strains of Candida auris and Brazilian Strains of Candida haemulonii Species Complex in Both Murine and Galleria mellonella Experimental Models

 ,
,  , , ,
, , , 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms
2.2. Galleria mellonella
2.3. Galleria mellonella Infection
2.4. Survival in Galleria mellonella
2.5. Fungal Burden in Galleria mellonella
2.6. Histopathology of Galleria mellonella
2.7. Yeast Phagocytic Capacity of Galleria Mellonella hemocytes
2.8. BALB/c Mice
2.9. Survival in BALB/c Mice
2.10. Biofilm Formation
2.11. Statistical Analysis
3. Results
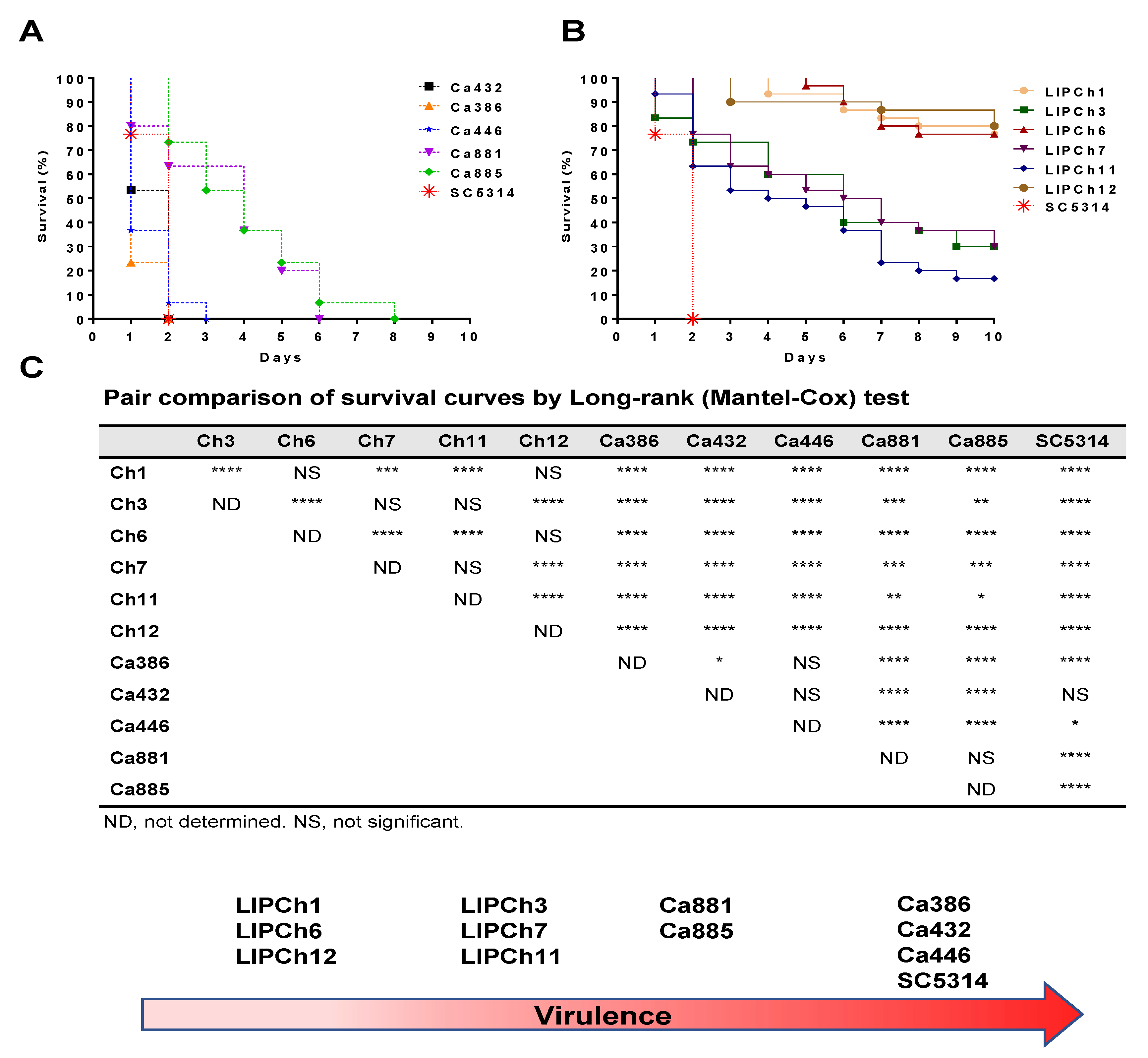
3.1. Galleria mellonella Infected with C. auris Survive Less than the Larvae Infected with the C. haemulonii Complex.
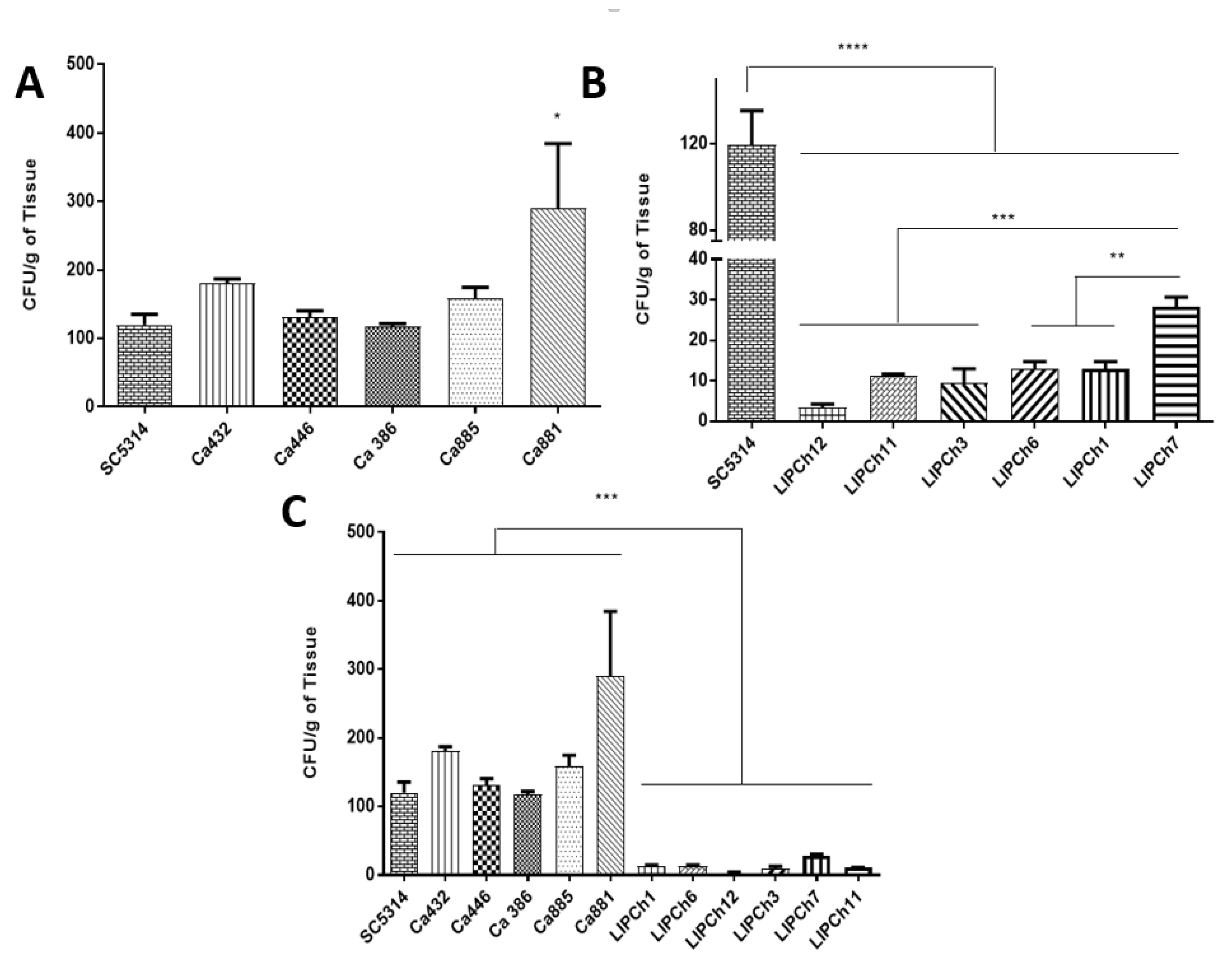
3.2. Galleria mellonella Infected with C. auris Displayed a High Fungal Burden
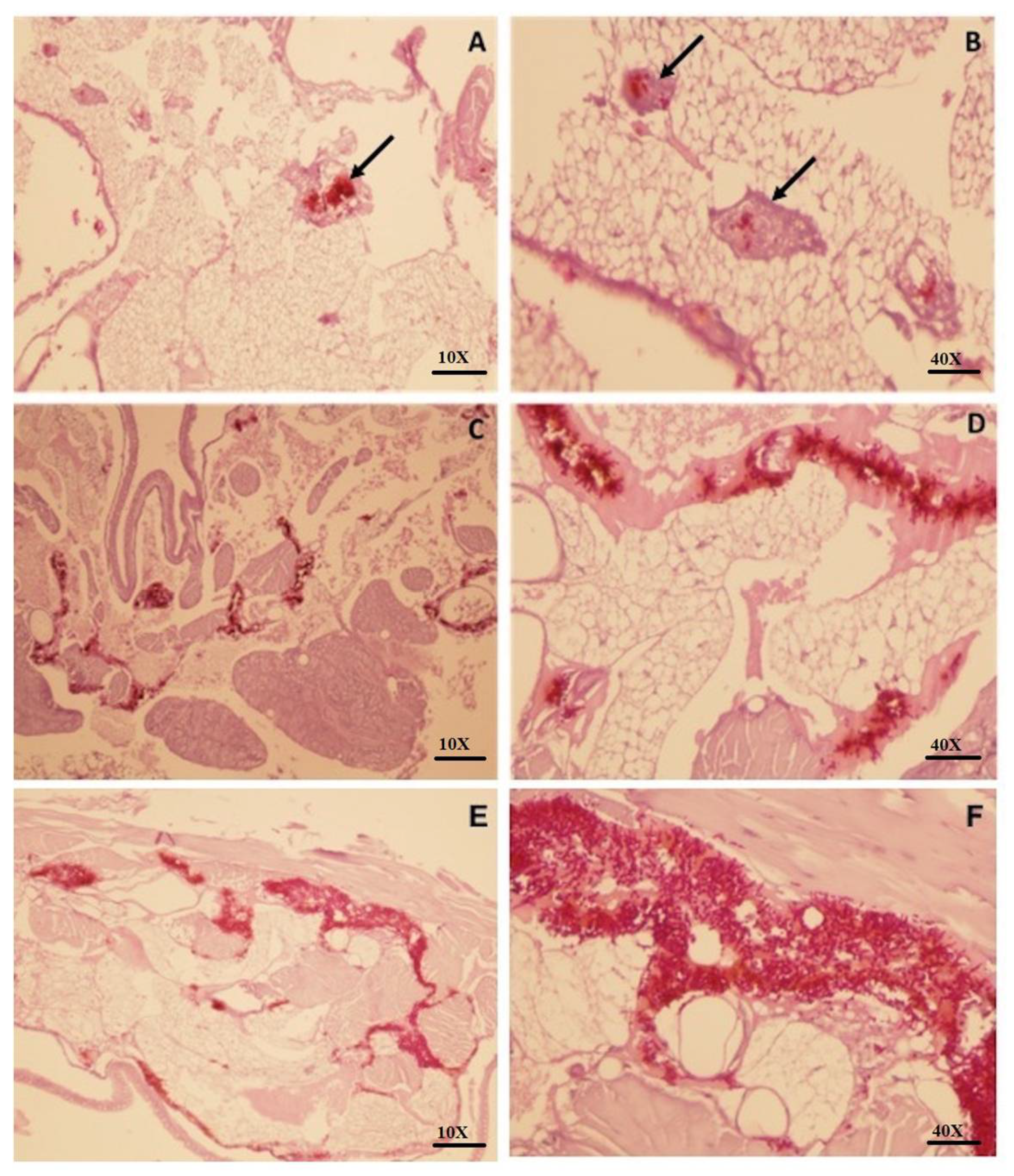
3.3. Larvae Infected with C. auris Leads to a Severe Tissue Invasion and Absence of Granuloma-Like Cell Aggregates
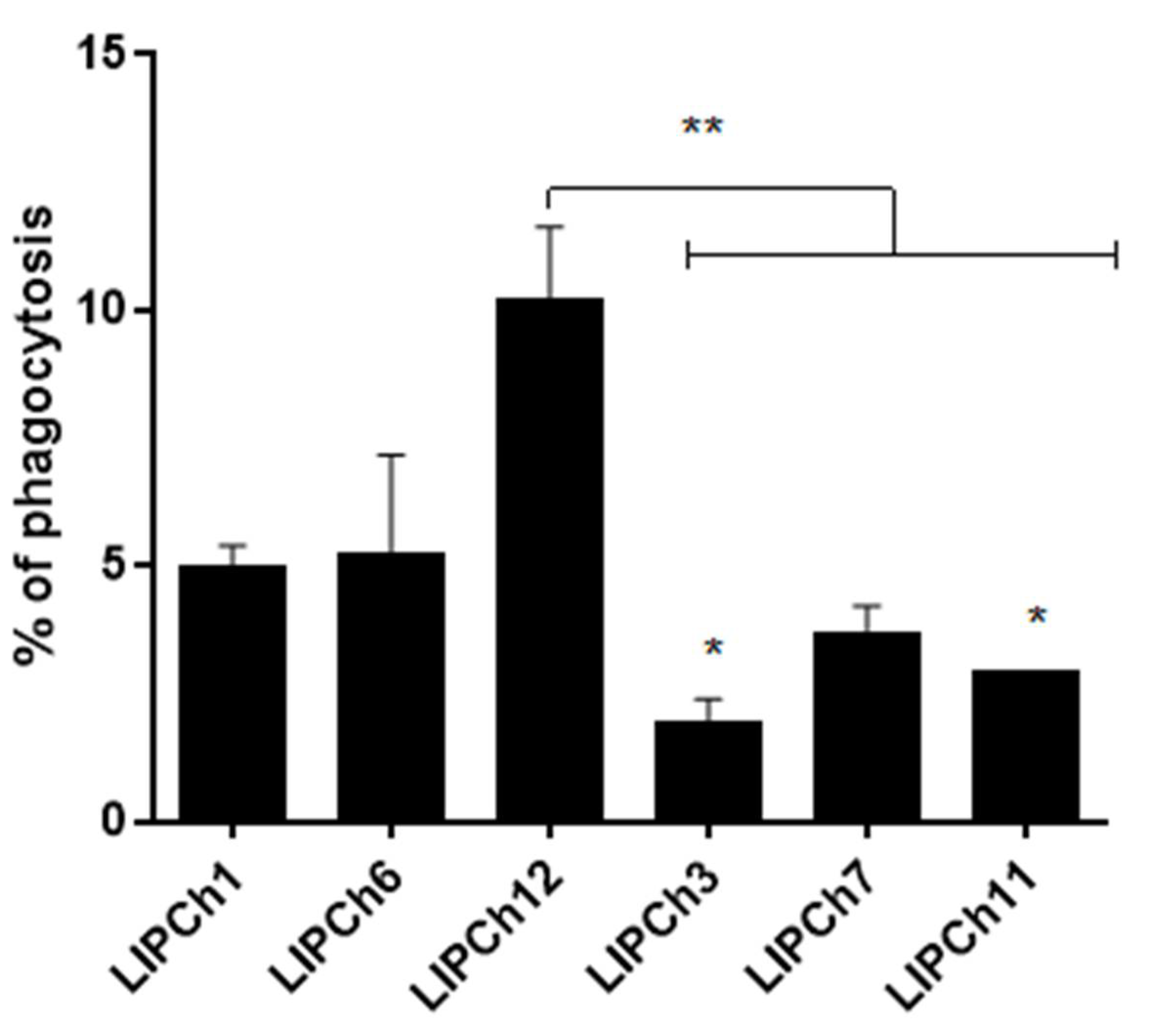
3.4. Virulence of C. auris and C. haemulonii in G. mellonella Model is Associated with Decrease in Phagocytosis
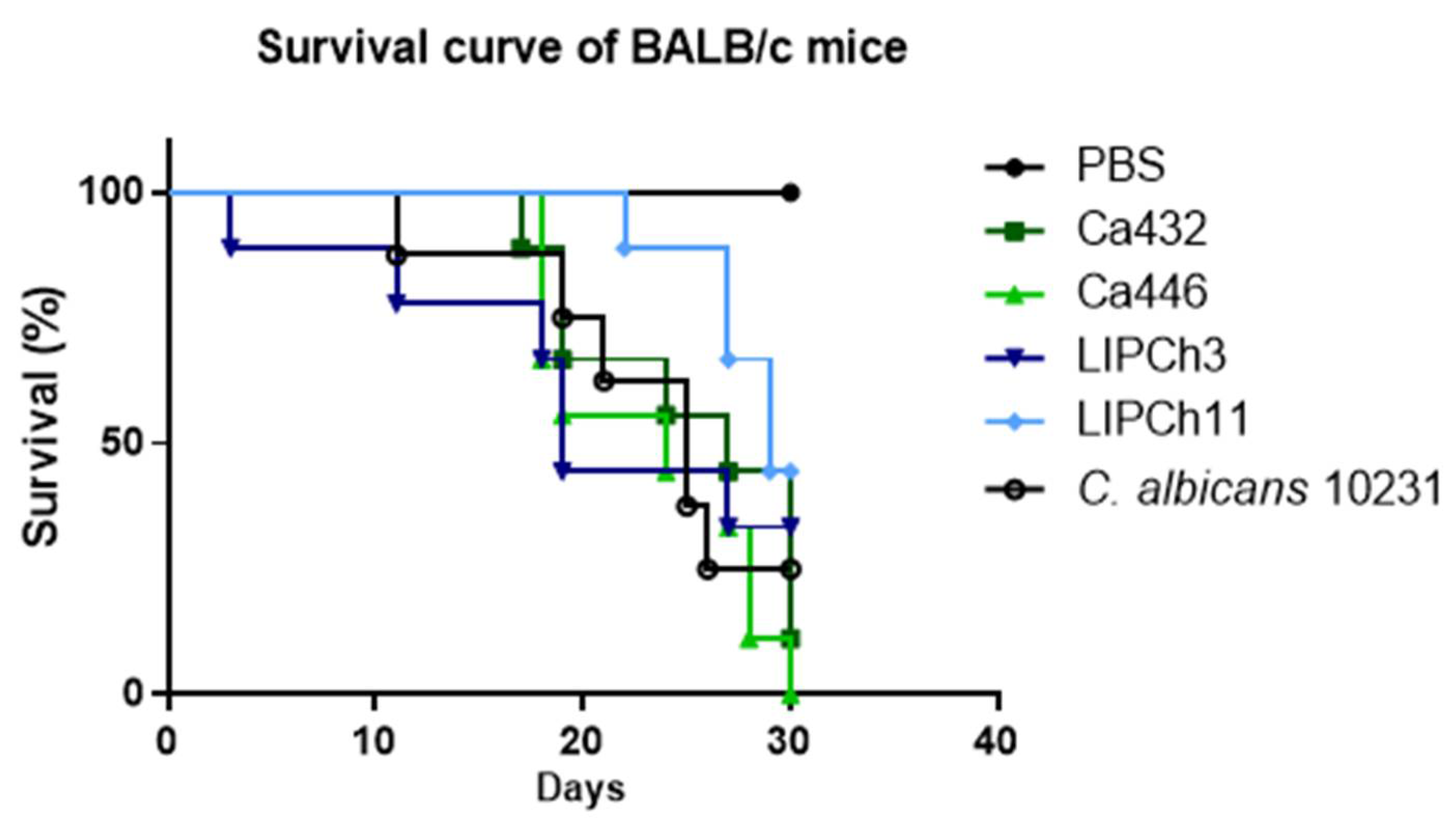
3.5. (C) auris and C. haemulonii Complex Infection have the Same Mortality Profile in an Immunosuppressed Animal Model
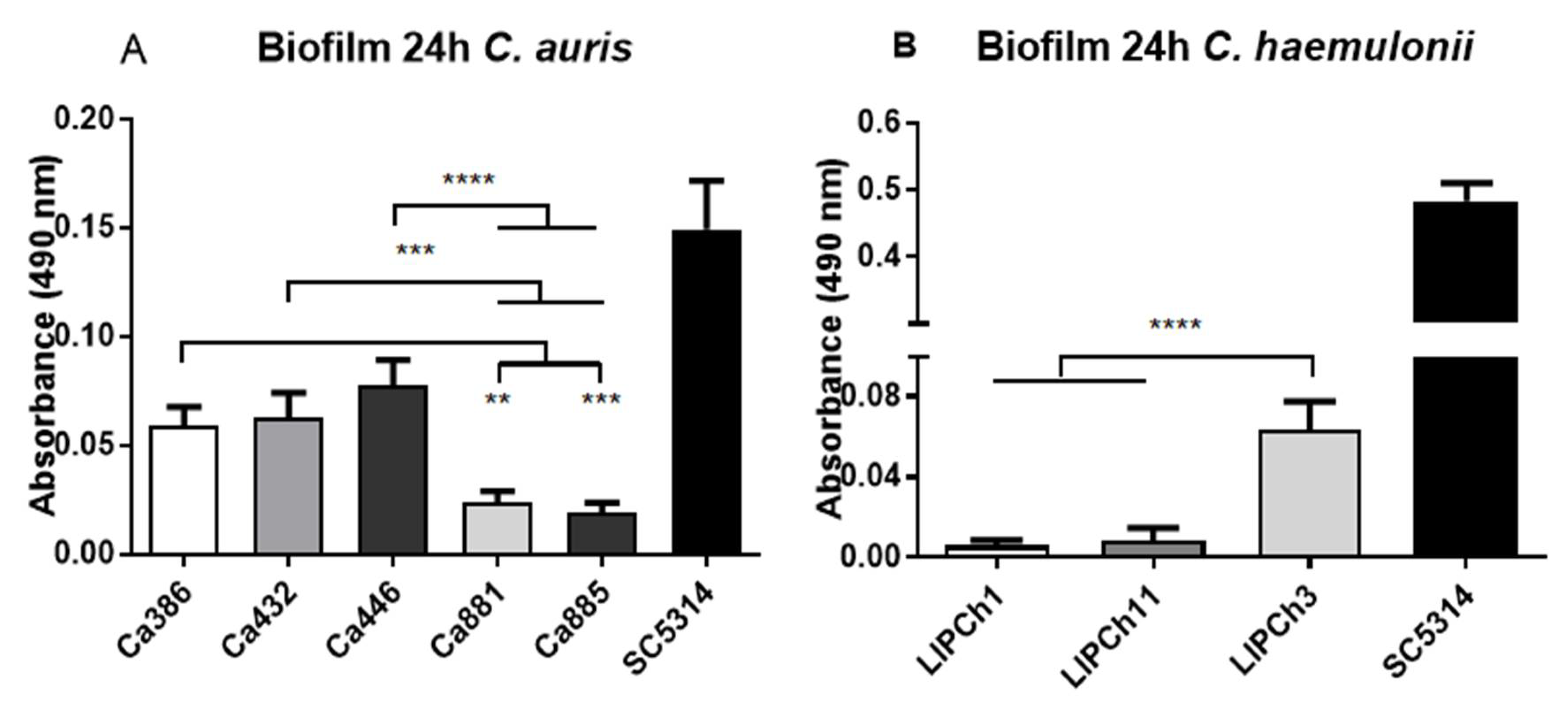
3.6. Biofilm Formation by C. auris and C. haemulonii Complex
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.L.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A.; et al. Simultaneous emergence of multidrug-resistant candida auris on 3 continents confirmed by whole-genome sequencing and epidemiological analyses. Clin. Infect. Dis. 2017, 64, 134–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.; Shin, J.H.; Sung, H.; Lee, K.; Kim, E.; Ryoo, N.; Lee, J.-S.; Jung, S.-I.; Park, K.H.; Kee, S.J.; et al. Candida haemulonii and closely related species at 5 university hospitals in Korea: Identification, antifungal susceptibility, and clinical features. Clin. Infect. Dis. 2009, 48, e57–e61. [Google Scholar] [CrossRef] [Green Version]
- Satoh, K.; Makimura, K.; Hasumi, Y.; Nishiyama, Y.; Uchida, K.; Yamaguchi, H. Candida auris sp. nov., a novel ascomycetous yeast isolated from the external ear canal of an inpatient in a Japanese hospital. Microbiol. Immunol. 2009, 53, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, A.; Sharma, C.; Duggal, S.; Agarwal, K.; Prakash, A.; Singh, P.K.; Jain, S.; Kathuria, S.; Randhawa, H.S.; Hagen, F.; et al. New clonal strain of Candida auris, Delhi, India. Emerg. Infect. Dis. 2013, 19, 1670–1673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magobo, R.E.; Corcoran, C.; Seetharam, S.; Govender, N.P. Candida auris—Associated candidemia, South Africa. Emerg. Infect. Dis. 2014, 20, 1250–1251. [Google Scholar] [CrossRef]
- Emara, M.; Ahmad, S.; Khan, Z.; Joseph, L.; Al-Obaid, I.M.; Purohit, P.; Bafna, R. Candida auris Candidemia in Kuwait, 2014. Emerg. Infect. Dis. 2015, 21, 1091–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nucci, M.; Queiroz-telles, F.; Alvarado-matute, T.; Tiraboschi, I.N.; Cortes, J.; Zurita, J.; Guzman-Blanco, M.; Santolaya, M.E.; Thompson, L.; Sifuentes-Osornio, J.; et al. Epidemiology of Candidemia in Latin America: A laboratory-based survey. PLoS ONE 2013, 8, e59373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales-López, S.E.; Parra-Giraldo, C.M.; Ceballos-Garzón, A.; Martínez, H.P.; Rodríguez, G.J.; Álvarez-Moreno, C.A.; Rodríguez, J.Y. Invasive infections with multidrug-resistant yeast Candida auris, Colombia. Emerg. Infect. Dis. 2017, 23, 162–164. [Google Scholar] [CrossRef] [Green Version]
- Parra-Giraldo, C.M.; Valderrama, S.L.; Cortes-Fraile, G.; Garzón, J.R.; Ariza, B.E.; Morio, F.; Linares-Linares, M.Y.; Ceballos-Garzón, A.; de la Hoz, A.; Hernandez, C.; et al. First report of sporadic cases of Candida auris in Colombia. Int. J. Infect. Dis. 2018, 69, 63–67. [Google Scholar] [CrossRef] [Green Version]
- Escandón, P.; Chow, N.A.; Caceres, D.H.; Gade, L.; Berkow, E.L.; Armstrong, P.; Rivera, S.; Misas, E.; Duarte, C.; Moulton-Meissner, H.; et al. Molecular epidemiology of Candida auris in Colombia reveals a highly related, countrywide colonization with regional patterns in amphotericin b resistance. Clin. Infect. Dis. 2019, 68, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive candidiasis. Nat Rev Dis Prim. 2018, 4, 18026. [Google Scholar] [CrossRef]
- Vallabhaneni, S.; Kallen, A.; Tsay, S.; Chow, N.; Welsh, R.; Kerins, J. Investigation of the first seven reported cases of Candida auris, a globally emerging invasive, multidrug-resistant fungus—United States, May 2013–August 2016. Am. J. Transpl. 2017, 17, 296–299. [Google Scholar] [CrossRef] [PubMed]
- Schelenz, S.; Hagen, F.; Rhodes, J.L.; Abdolrasouli, A.; Chowdhary, A.; Hall, A.; Ryan, L.; Shackleton, J.; Trimlett, R.; Meis, J.F.; et al. First hospital outbreak of the globally emerging Candida auris in a European hospital. Antimicrob. Resist. Infect. Control. 2016, 5, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Ramos, L.S.; Figueiredo-Carvalho, M.H.G.; Barbedo, L.S.; Ziccardi, M.; Chaves, A.L.S.; Zancopé-Oliveira, R.M.; Pinto, M.R.; Sgarbi, D.B.G.; Dornelas-Ribeiro, M.; Branquinha, M.H.; et al. Candida haemulonii complex: Species identification and antifungal susceptibility profiles of clinical isolates from Brazil. J. Antimicrob. Chemother. 2015, 70, 111–115. [Google Scholar] [CrossRef] [Green Version]
- Shackleton, J.; Schelenz, S.; Rochon, M.; Hall, A.; Ryan, L.; Cervera-Jackson, R. The impact of environmental decontamination in a Candida auris outbreak. J. Hosp. Infect. 2016, 94, S24–S134. [Google Scholar] [CrossRef]
- Sarma, S.; Kumar, N.; Sharma, S.; Govil, D.; Ali, T.; Mehta, Y.; Rattan, A. Candidemia caused by amphotericin B and Fluconazole resistant Candida auris. Indian J. Med. Microbiol. 2013, 31, 90. [Google Scholar] [CrossRef]
- Rossato, L.; Colombo, A.L. Candida auris: What have we learned about its mechanisms of pathogenicity? Front Microbiol. 2018, 9, 1–6. [Google Scholar] [CrossRef]
- Lavarde, V. Peritonite mycosique a Torulopsis haemulonii. Bul. Soc. Fr. Mycol. Med. 1984, 13, 173–176. [Google Scholar]
- Khan, Z.U.; Al-Sweih, N.A.; Ahmad, S.; Al-Kazemi, N.; Khan, S.; Joseph, L.; Chandy, R. Outbreak of fungemia among neonates caused by Candida haemulonii resistant to amphotericin B, itraconazole, and fluconazole. J. Clin. Microbiol. 2007, 45, 2025–2027. [Google Scholar] [CrossRef] [Green Version]
- Muro, M.D.; Motta, F.D.A.; Burger, M.; Melo, A.S.D.A.; Dalla-Costa, L.M. Echinocandin resistance in two Candida haemulonii isolates from pediatric patients. J. Clin. Microbiol. 2012, 50, 3783–3785. [Google Scholar] [CrossRef] [Green Version]
- Ruan, S.; Kuo, Y.; Huang, C.; Hsiue, H.; Hsueh, P. Infections due to Candida haemulonii: Species identification, antifungal susceptibility and outcomes. Int. J. Antimicrob. Agents. 2010, 35, 85–88. [Google Scholar] [CrossRef]
- Cendejas-Bueno, E.; Kolecka, A.; Alastruey-Izquierdo, A.; Theelen, B.; Groenewald, M.; Kostrzewa, M.; Boekhout, T. Reclassification of the Candida haemulonii complex as Candida. J. Clin. Microbiol. 2012, 50, 3641–3651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, D.C.; Muñoz, J.E.; Carvalho, D.D.; Belmonte, R.; Faintuch, B.; Borelli, P.; Daffre, S. Therapeutic use of a cationic antimicrobial peptide from the spider Acanthoscurria gomesiana in the control of experimental candidiasis. BMC Microbiol. 2012, 12, 28. [Google Scholar] [CrossRef] [Green Version]
- Pierce, C.G.; Uppuluri, P.; Tristan, A.R.; Jr, F.L.W.; Mowat, E.; Ramage, G.; Lopez-Ribot, J.L. A simple and reproducible 96 well plate-based method for the formation of fungal biofilms and its application to antifungal susceptibility testing. Nat. Protoc. 2008, 3, 1494–1500. [Google Scholar] [CrossRef] [PubMed]
- Gandra, R.M.; McCarron, P.; Viganor, L.; Fernandes, M.F.; Kavanagh, K.; McCann, M.; Devereux, M. In vivo activity of Copper(II), manganese(II), and silver(I) 1,10-phenanthroline chelates against Candida haemulonii using the Galleria mellonella model. Front Microbiol. 2020, 11, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Sherry, L.; Ramage, G.; Kean, R.; Borman, A.; Johnson, E.M.; Richardson, M.D. Biofilm-forming capability of highly virulent, multidrug-resistant Candida auris. Emerg. Infect. Dis. 2017, 23, 328–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borman, A.M.; Szekely, A.; Johnson, E.M. Comparative pathogenicity of United Kingdom isolates of the emerging. mSphere 2016, 1, e00189-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azar, M.M.; Turbett, S.E.; Fishman, J.A.; Pierce, V.M. Donor-derived transmission of Candida auris during lung transplantation. Clin. Infect. Dis. 2017, 65, 1040–1042. [Google Scholar] [CrossRef]
- Bravo Ruiz, G.; Ross, Z.K.; Gow, N.A.R.; Lorenz, A. Pseudohyphal growth of the emerging pathogen Candida auris is triggered by genotoxic stress through the S Phase Checkpoint. mSphere 2020, 5, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Ben-Ami, R.; Berman, J.; Novikov, A.; Bash, E.; Shachor-Meyouhas, Y.; Zakin, S.; Finn, T. Multidrug resistant Candida haemulonii and Candida auris Tel Aviv, Israel. Emerg. Infect. Dis. 2017, 23, 195–203. [Google Scholar] [CrossRef]
- Bergin, D.; Brennan, M.; Kavanagh, K. Fluctuations in haemocyte density and microbial load may be used as indicators of fungal pathogenicity in larvae of Galleria mellonella. Microbes Infect. 2003, 5, 1389–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gago, S.; García-Rodas, R.; Cuesta, I.; Mellado, E.; Alastruey-Izquierdo, A. Candida parapsilosis, Candida orthopsilosis, and Candida metapsilosis virulence in the non-conventional host Galleria mellonella. Virulence 2014, 5, 278–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, L.N.; Campos-Silva, R.; Ramos, L.S.; Trentin, D.S.; Macedo, A.J.; Branquinha, M.H.; Santos, A.L. Virulence of Candida haemulonii complex in Galleria mellonella and efficacy of classical antifungal drugs: A comparative study with other clinically relevant non-albicans Candida species. FEMS Yeast Res. 2018, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conti, H.R.; Huppler, A.R.; Whibley, N.; Gaffen, S.L. Animal models for Candidiasis. Curr. Protoc. Immunol. 2014. [Google Scholar] [CrossRef] [Green Version]
- Larkin, E.; Hager, C.; Chandra, J.; Mukherjee, P.K. The emerging pathogen Candida auris: Growth phenotype, virulence factors, activity of antifungals, and effect of SCY-078, a novel glucan synthesis inhibitor, on growth morphology and biofilm formation emily. Antimicrob. Agents Chemother. 2017, 61, e02396-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fakhim, H.; Vaezi, A.; Dannaoui, E.; Chowdhary, A.; Nasiry, D.; Faeli, L.; Meis, J.F.; Badali, H. Comparative virulence of Candida auris with Candida haemulonii, Candida glabrata and Candida albicans in a murine model. Mycoses 2018, 61, 377–382. [Google Scholar] [CrossRef] [Green Version]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Isolation Date | Source | Country | Susceptibility Profile | |
|---|---|---|---|---|---|
| AMB | FLZ | ||||
| C. auris 432 | 2014 | Craniotomy (Secretion) | Colombia | R | R |
| C. auris 446 | 2015 | Blood culture | Colombia | S | R |
| C. auris 386 | 2015 | Bone Tissue (Biopsy) | Colombia | S | R |
| C. auris 885 | 2016 | Blood culture | Colombia | S | R |
| C. auris 881 | 2016 | Cerebrospinal fluid | Colombia | S | R |
| C. duobushaemulonii LIPCh1 | 2005 | Finger nail | Brazil | R | R |
| C. duobushaemulonii LIPCh6 | 2009 | Toe nail | Brazil | R | R |
| C. haemulonii LIPCh3 | 2009 | Toe nail | Brazil | R | R |
| C. haemulonii LIPCh7 | 2009 | Toe nail | Brazil | R | R |
| C. haemulonii LIPCh12 | 2013 | Blood | Brazil | R | R |
| C. haemulonii var. vulnera LIPCh11 | 2013 | Blood | Brazil | R | R |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muñoz, J.E.; Ramirez, L.M.; Dias, L.d.S.; Rivas, L.A.; Ramos, L.S.; Santos, A.L.S.; Taborda, C.P.; Parra-Giraldo, C.M. Pathogenicity Levels of Colombian Strains of Candida auris and Brazilian Strains of Candida haemulonii Species Complex in Both Murine and Galleria mellonella Experimental Models. J. Fungi 2020, 6, 104. https://doi.org/10.3390/jof6030104
Muñoz JE, Ramirez LM, Dias LdS, Rivas LA, Ramos LS, Santos ALS, Taborda CP, Parra-Giraldo CM. Pathogenicity Levels of Colombian Strains of Candida auris and Brazilian Strains of Candida haemulonii Species Complex in Both Murine and Galleria mellonella Experimental Models. Journal of Fungi. 2020; 6(3):104. https://doi.org/10.3390/jof6030104
Chicago/Turabian StyleMuñoz, Julián E., Laura M. Ramirez, Lucas dos Santos Dias, Laura A. Rivas, Lívia S. Ramos, André L. S. Santos, Carlos P. Taborda, and Claudia M. Parra-Giraldo. 2020. "Pathogenicity Levels of Colombian Strains of Candida auris and Brazilian Strains of Candida haemulonii Species Complex in Both Murine and Galleria mellonella Experimental Models" Journal of Fungi 6, no. 3: 104. https://doi.org/10.3390/jof6030104