EFE-Mediated Ethylene Synthesis Is the Major Pathway in the Citrus Postharvest Pathogen Penicillium digitatum during Fruit Infection



Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Culture Conditions
2.2. Gene Expression Analysis
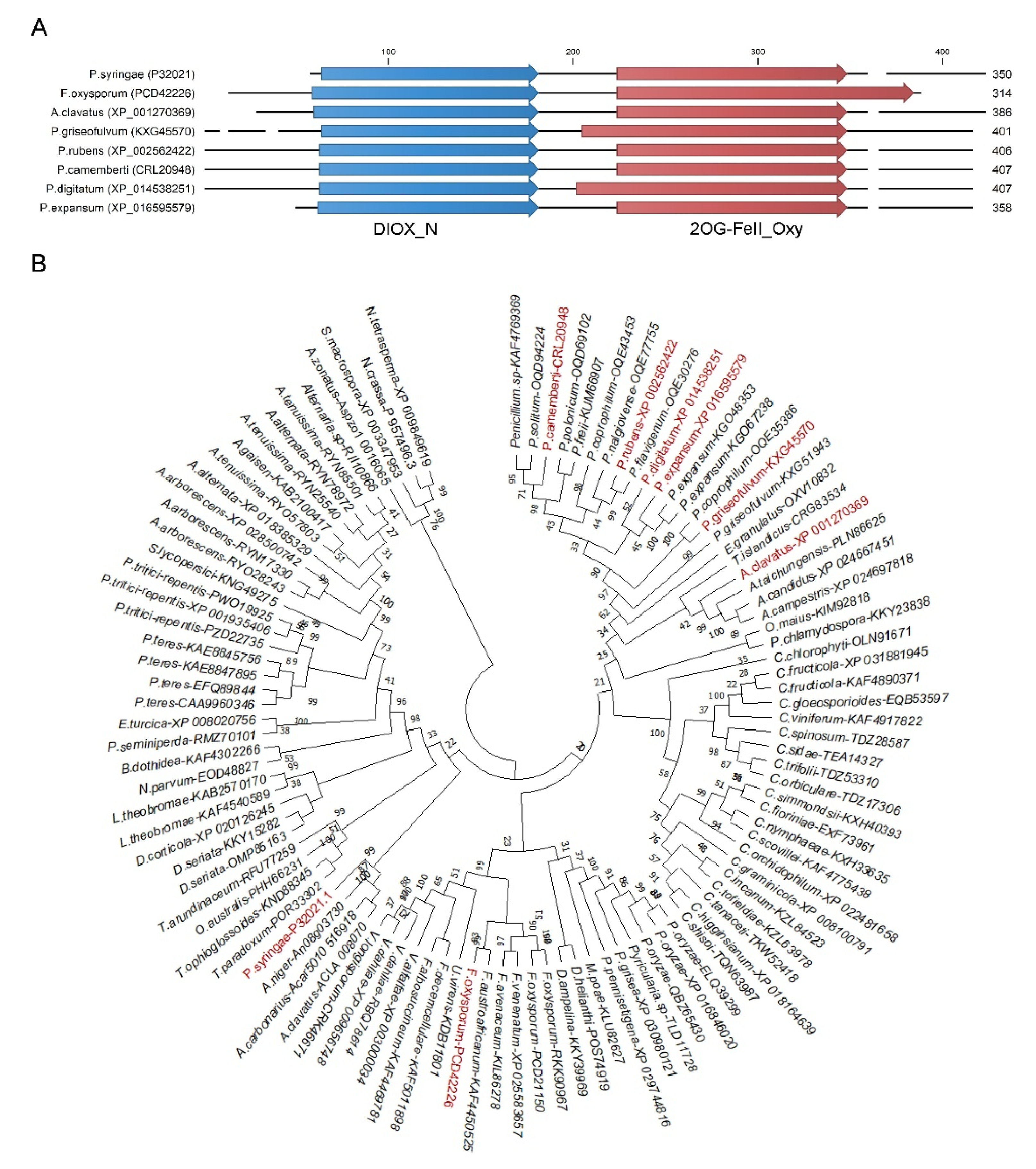
2.3. Sequence and Phylogenetic Analysis
2.4. Construction and Verification of the P. digitatum ΔefeA Knockout Mutants
2.5. Characterization of the ΔefeA Knockout Mutants: Mycelial Growth, Sporulation, and In Vitro Ethylene Production
2.6. Ethylene Production during P. digitatum Growth on Orange Discs
2.7. Citrus Fruit Pathogenicity Assays
2.8. Ethylene Production during Orange Infection by P. digitatum
3. Results
3.1. Identification and Phylogenetic Analysis of Fungal efe Genes
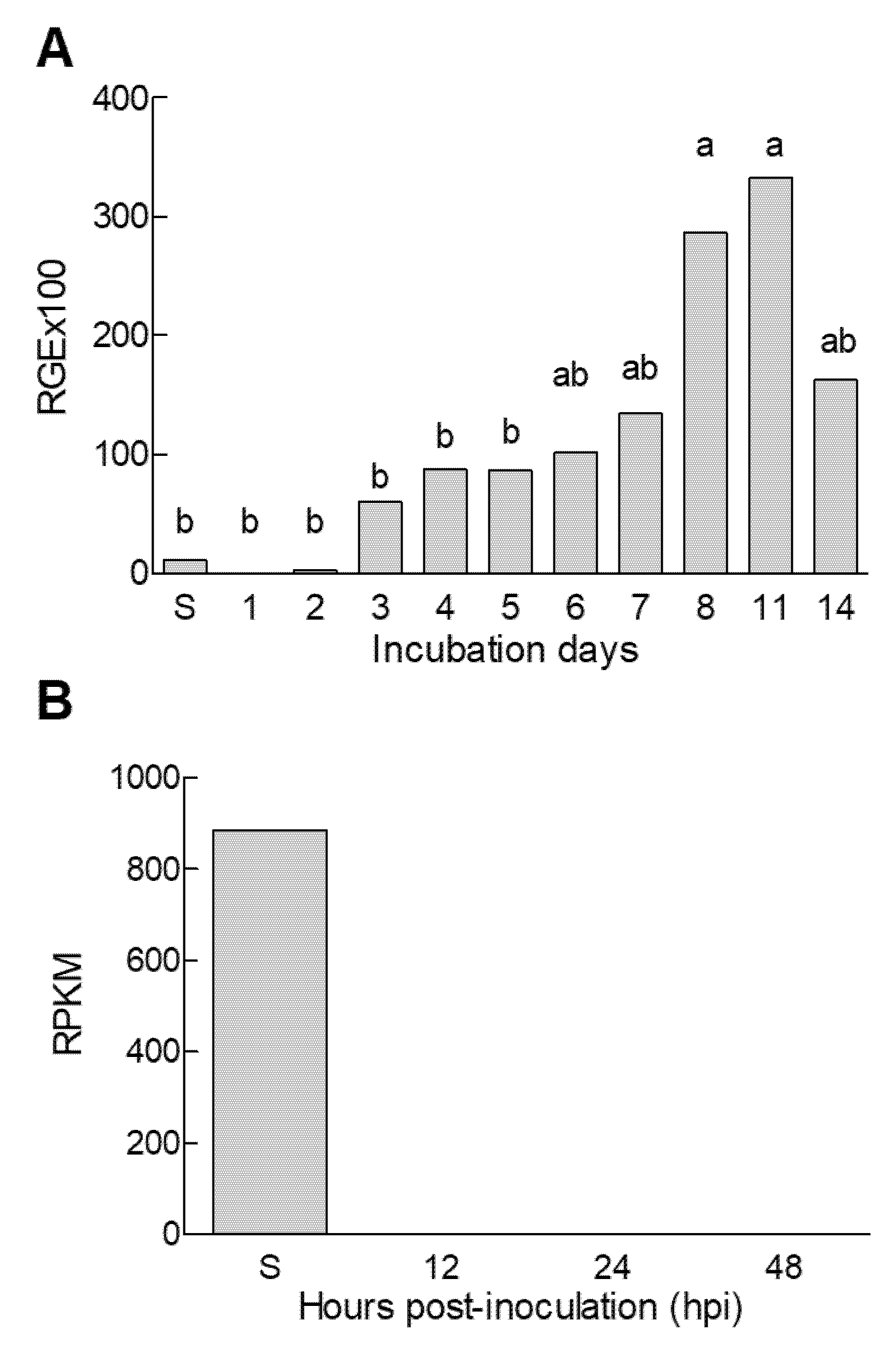
3.2. Characterization of P. digitatum efeA Gene Expression
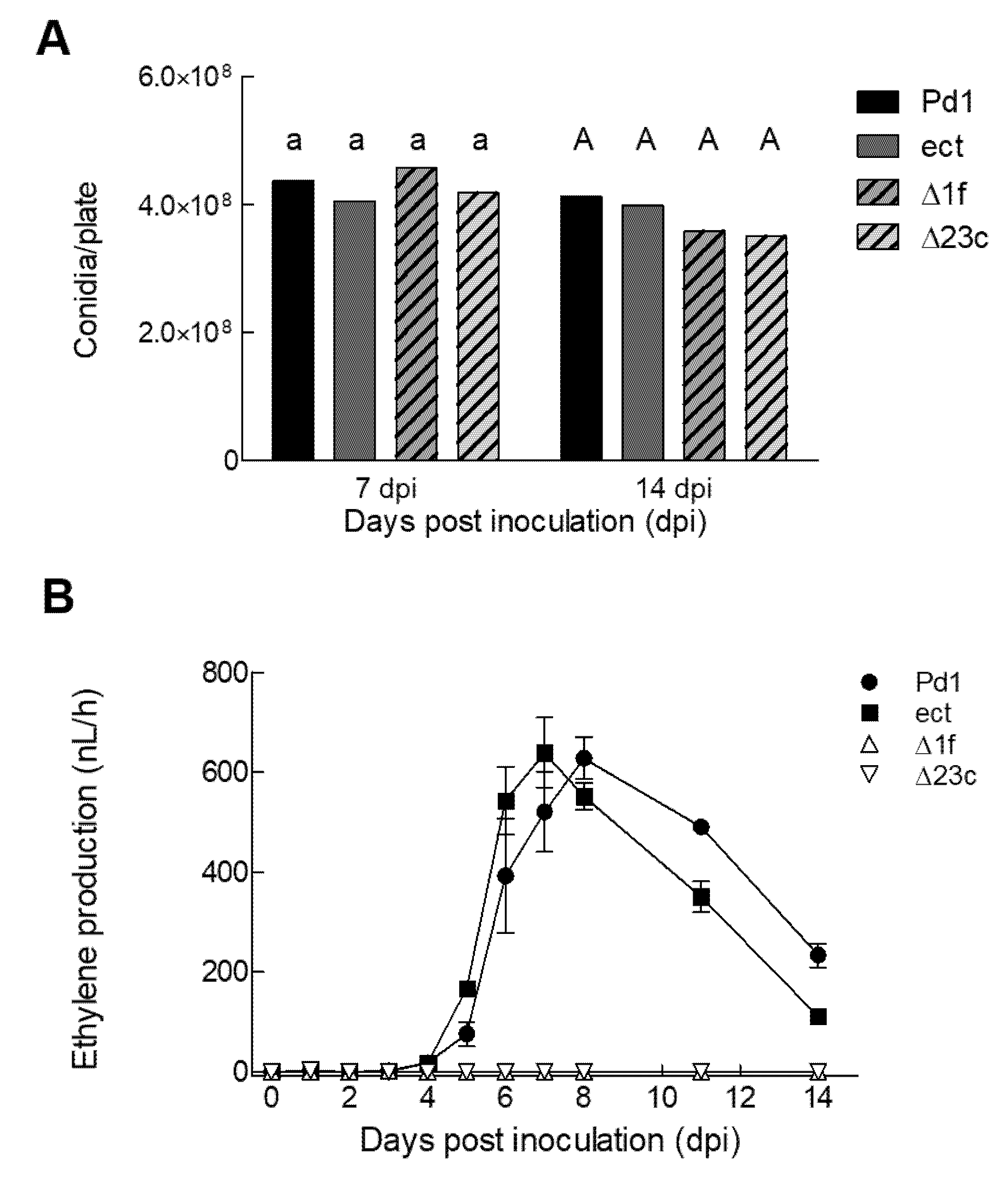
3.3. Phenotypic Characterization of the Mutants
3.4. P. digitatum ΔefeA Knockout Mutant does not Produce Ethylene during Growth on Orange Peel Discs
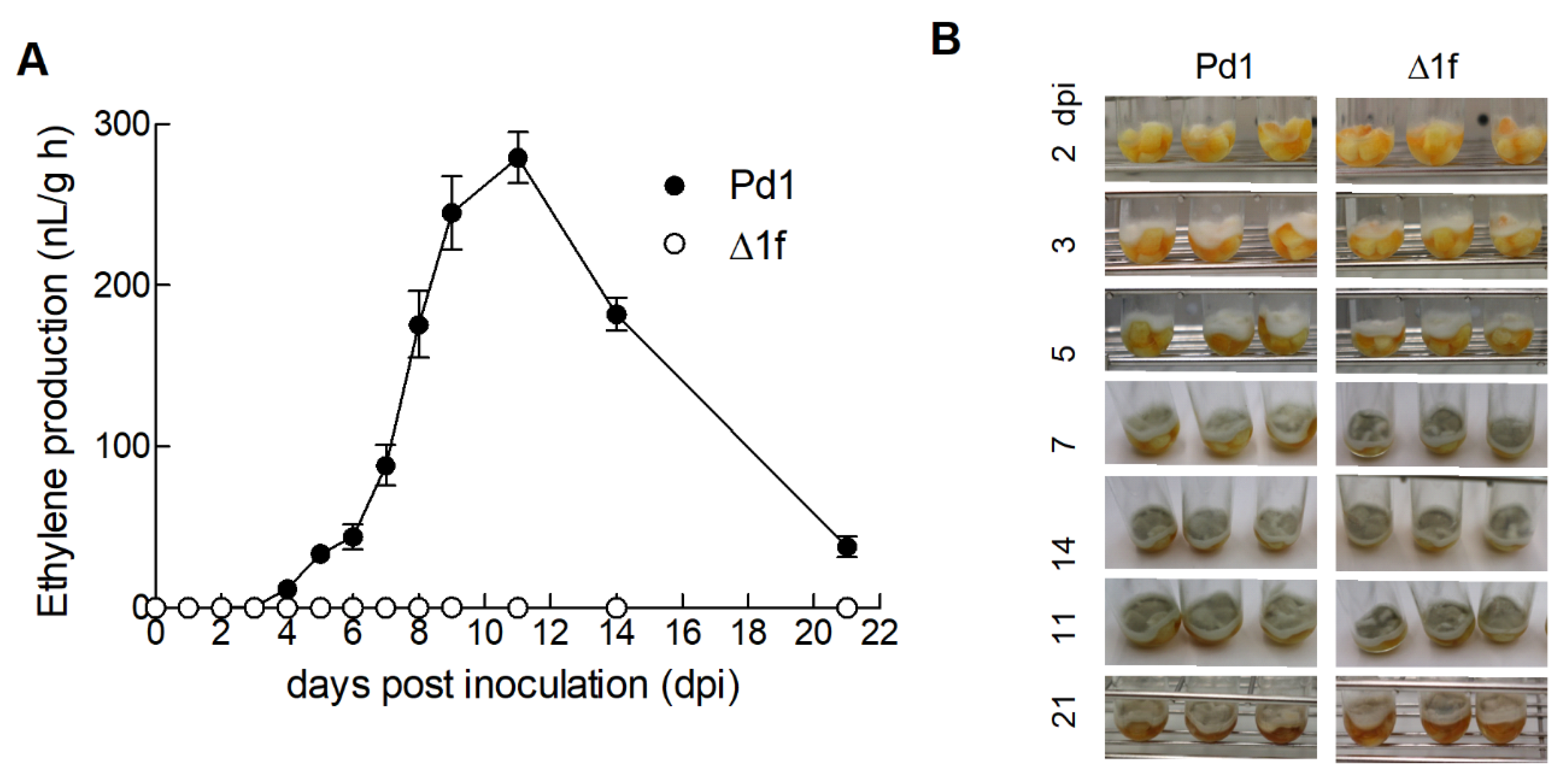
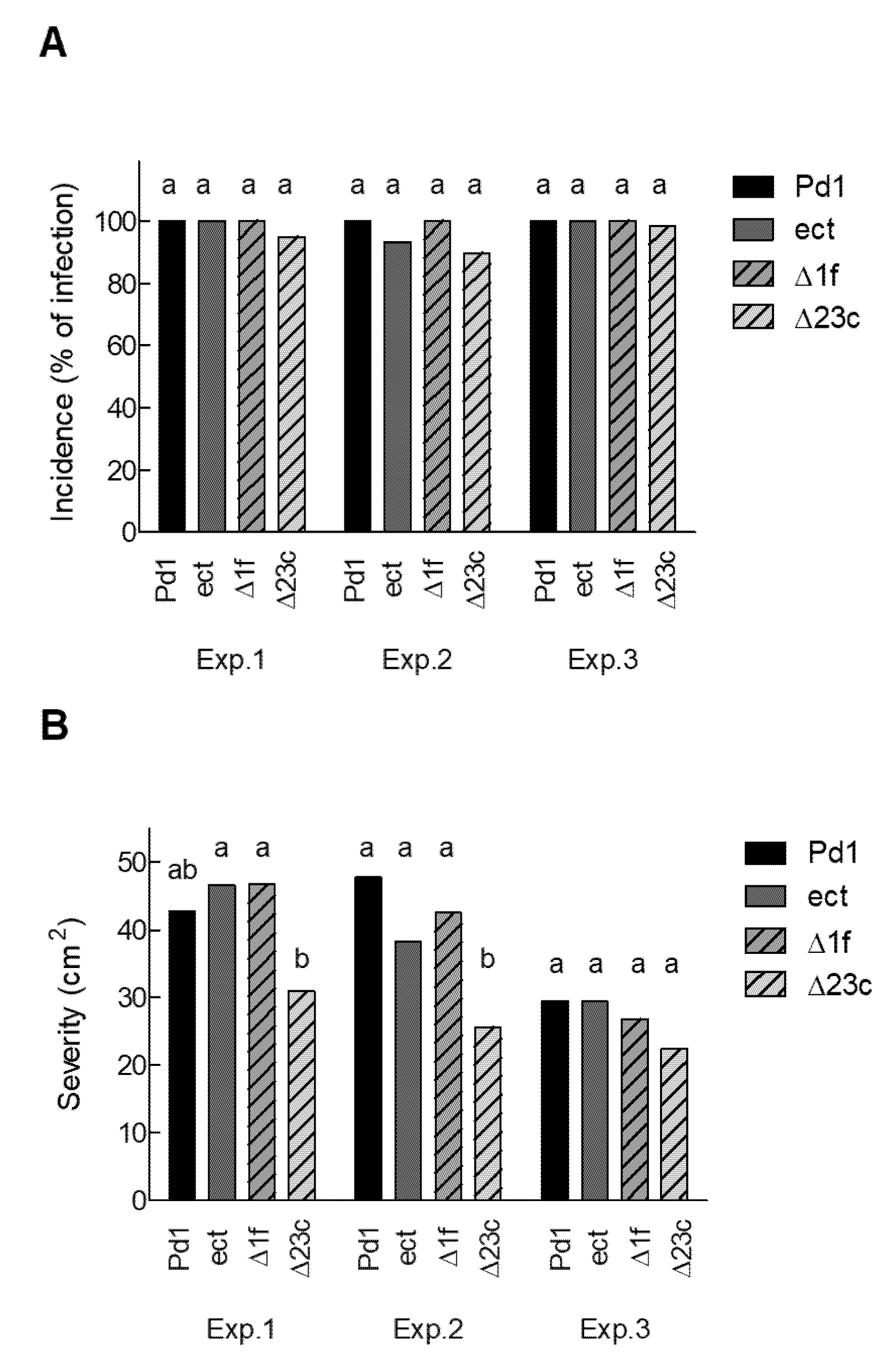
3.5. Ethylene Production by P. digitatum Is Dispensable for Orange Fruit Infection
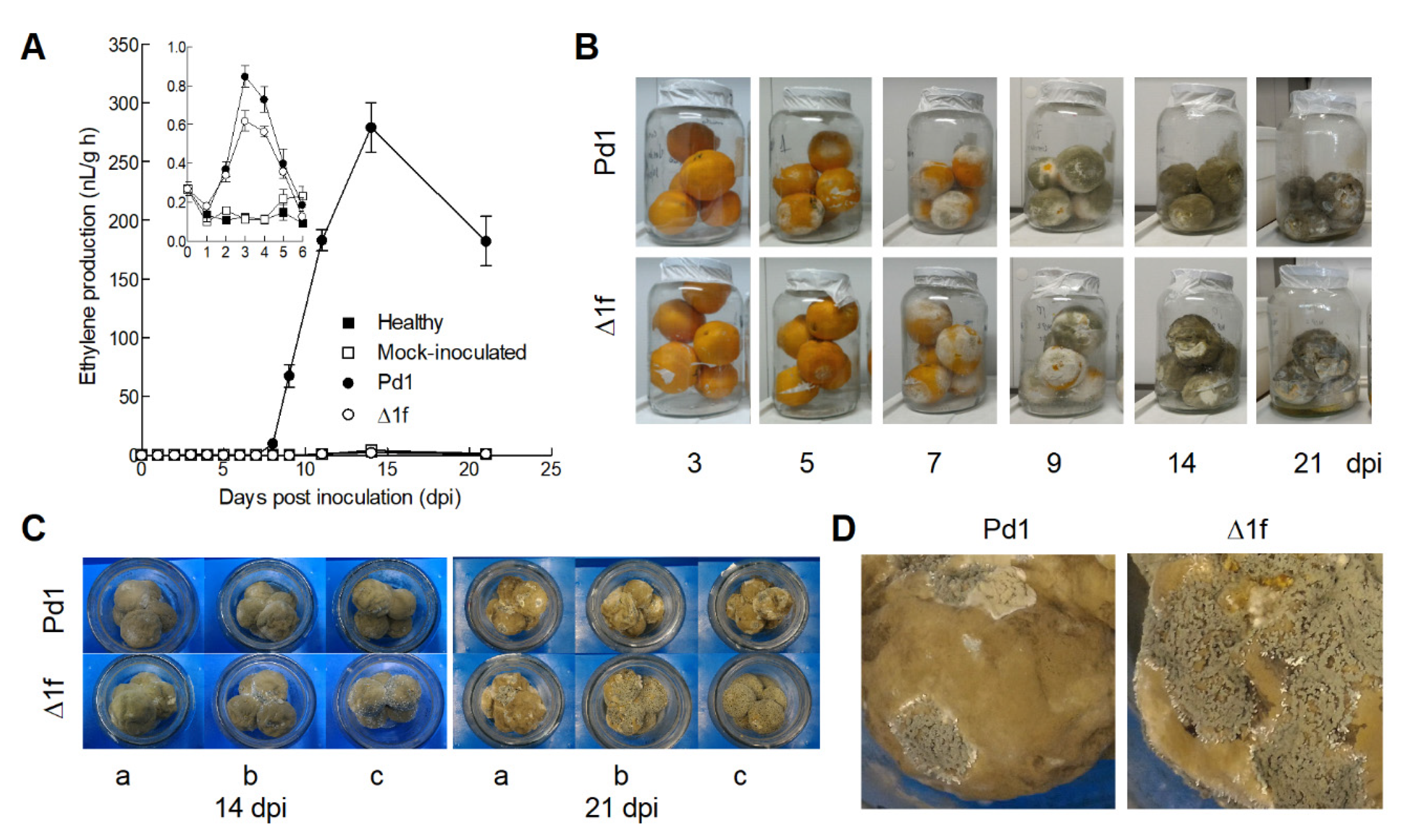
3.6. Changes in Ethylene Production during the Orange Fruit Infection by P. digitatum
4. Discussion
4.1. The 2-Oxoglutarate EFE-Mediated Pathway Is the Major Ethylene Biosynthetic Pathway Used by P. digitatum during in Vitro Growth on PDA
4.2. Ethylene Is Synthesized by EFE in P. digitatum during Citrus Infection, but Is Dispensable for Establishing Infection
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Klee, H.J.; Giovannoni, J.J. Genetics and control of tomato fruit ripening and quality attributes. Annu. Rev. Genet. 2011, 45, 41–59. [Google Scholar] [CrossRef]
- Chagué, V.; Elad, Y.; Barakat, R.; Tudzynski, P.; Sharon, A. Ethylene biosynthesis in Botrytis cinerea. FEMS Microbiol. Ecol. 2002, 40, 143–149. [Google Scholar] [CrossRef]
- De Paepe, A.; Van Der Straeten, D. Ethylene biosynthesis and signaling: An overview. In Vitamins & Hormones; Litwack, G., Ed.; Plant Hormones; Academic Press: New York, NY, USA, 2005; Volume 72, pp. 399–430. [Google Scholar]
- Fukuda, H.; Ogawa, T.; Tanase, S. Ethylene production by micro-organisms. In Advances in Microbial Physiology; Rose, A., Ed.; Academic Press Inc.: London, UK, 1993; Volume 35, pp. 275–306. ISBN 0-12-027735-2. [Google Scholar]
- Ladygina, N.; Dedyukhina, E.G.; Vainshtein, M.B. A review on microbial synthesis of hydrocarbons. Process Biochem. 2006, 41, 1001–1014. [Google Scholar] [CrossRef]
- Wang, K.L.C.; Li, H.; Ecker, J.R. Ethylene biosynthesis and signaling networks. Plant Cell 2002, 14, S131–S151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Billington, D.C.; Golding, B.T.; Primrose, S.B. Biosynthesis of ethylene from methionine. Isolation of the putative intermediate 4-methylthio-2-oxobutanoate from culture fluids of bacteria and fungi. Biochem. J. 1979, 182, 827–836. [Google Scholar] [CrossRef] [PubMed]
- Cristescu, S.M.; De Martinis, D.; te Lintel Hekkert, S.; Parker, D.H.; Harren, F.J.M. Ethylene production by Botrytis cinerea in vitro and in tomatoes. Appl. Environ. Microbiol. 2002, 68, 5342–5350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chalutz, E.; Lieberman, M.; Sisler, H.D. Methionine-induced ethylene production by Penicillium digitatum. Plant Physiol. 1977, 60, 402–406. [Google Scholar] [CrossRef] [Green Version]
- Chou, T.W.; Yang, S.F. The biogenesis of ethylene in Penicillium digitatum. Arch. Biochem. Biophys. 1973, 157, 73–82. [Google Scholar] [CrossRef]
- Eckert, C.; Xu, W.; Xiong, W.; Lynch, S.; Ungerer, J.; Tao, L.; Gill, R.; Maness, P.-C.; Yu, J. Ethylene-forming enzyme and bioethylene production. Biotechnol. Biofuels 2014, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- North, J.A.; Miller, A.R.; Wildenthal, J.A.; Young, S.J.; Tabita, F.R. Microbial pathway for anaerobic 5′-methylthioadenosine metabolism coupled to ethylene formation. Proc. Natl. Acad. Sci. USA 2017, 114, E10455–E10464. [Google Scholar] [CrossRef] [Green Version]
- Biale, J.B. Effect of emanations from several species of fungi on respiration and color development of citrus fruits. Sci. Wash. 1940, 91, 458–459. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.V.; Winston, J.R.; Fisher, D.F. Production of epinasty by emanations from normal and decaying citrus fruits and from Penicillium digitatum. J. Agric. Res. 1940, 60, 269–277. [Google Scholar]
- Chagué, V. Ethylene production by fungi: Biological questions and future developments towards a sustainable polymers industry. In Handbook of Hydrocarbon and Lipid Microbiology; Timmis, K.N., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 3011–3020. ISBN 978-3-540-77587-4. [Google Scholar]
- van Loon, L.C.; Geraats, B.P.J.; Linthorst, H.J.M. Ethylene as a modulator of disease resistance in plants. Trends Plant Sci. 2006, 11, 184–191. [Google Scholar] [CrossRef] [PubMed]
- van der Ent, S.; Pieterse, C.M.J. Ethylene: Multi-tasker in plant–attacker interactions. In Annual Plant Reviews; American Cancer Society: Atlanta, GA, USA, 2012; Volume 44, pp. 343–377. ISBN 978-1-119-31299-4. [Google Scholar]
- Achilea, O.; Fuchs, Y.; Chalutz, E.; Rot, I. The contribution of host and pathogen to ethylene biosynthesis in Penicillium digitatum-infected citrus fruit. Physiol. Plant Pathol. 1985, 27, 55–63. [Google Scholar] [CrossRef]
- Achilea, O.; Chalutz, E.; Fuchs, Y.; Rot, I. Ethylene biosynthesis and related physiological changes in Penicillium digitatum infected grapefruit (Citrus paradisi). Physiol. Plant Pathol. 1985, 26, 125–134. [Google Scholar] [CrossRef]
- Bockhaven, J.V.; Spíchal, L.; Novák, O.; Strnad, M.; Asano, T.; Kikuchi, S.; Höfte, M.; Vleesschauwer, D.D. Silicon induces resistance to the brown spot fungus Cochliobolus miyabeanus by preventing the pathogen from hijacking the rice ethylene pathway. New Phytol. 2015, 206, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Chanclud, E.; Morel, J.-B. Plant hormones: A fungal point of view. Mol. Plant Pathol. 2016, 17, 1289–1297. [Google Scholar] [CrossRef]
- Goyal, A.; Kalia, A. Fungal phytohormones: Plant growth-regulating substances and their applications in crop productivity. In Agriculturally Important Fungi for Sustainable Agriculture: Volume 1: Perspective for Diversity and Crop Productivity; Yadav, A.N., Mishra, S., Kour, D., Yadav, N., Kumar, A., Eds.; Fungal Biology; Springer International Publishing: Cham, Switzerland, 2020; pp. 143–169. ISBN 978-3-030-45971-0. [Google Scholar]
- Shen, Q.; Liu, Y.; Naqvi, N.I. Fungal effectors at the crossroads of phytohormone signaling. Curr. Opin. Microbiol. 2018, 46, 1–6. [Google Scholar] [CrossRef]
- Chalutz, E. No role for ethylene in the pathogenicity of Penicillium digitatum. Physiol. Plant Pathol. 1979, 14, 259–262. [Google Scholar] [CrossRef]
- Fukuda, H.; Fujii, T.; Ogawa, T. Preparation of a cell-free ethylene-forming system from Penicillium digitatum. Agric. Biol. Chem. 1986, 50, 977–981. [Google Scholar] [CrossRef]
- Yang, J.; Giné-Bordonaba, J.; Vilanova, L.; Teixidó, N.; Usall, J.; Larrigaudière, C.; Torres, R. An insight on the ethylene biosynthetic pathway of two major fruit postharvest pathogens with different host specificity: Penicillium digitatum and Penicillium expansum. Eur. J. Plant Pathol. 2017, 149, 575–585. [Google Scholar] [CrossRef]
- Chalutz, E.; Lieberman, M. Inhibition of ethylene production in Penicillium digitatum. Plant Physiol. 1978, 61, 111–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcet-Houben, M.; Ballester, A.-R.; De la Fuente, B.; Harries, E.; Marcos, J.F.; González-Candelas, L.; Gabaldón, T. Genome sequence of the necrotrophic fungus Penicillium digitatum, the main postharvest pathogen of citrus. BMC Genom. 2012, 13, 646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johansson, N.; Persson, K.O.; Larsson, C.; Norbeck, J. Comparative sequence analysis and mutagenesis of ethylene forming enzyme (EFE) 2-oxoglutarate/Fe(II)-dependent dioxygenase homologs. BMC Biochem. 2014, 15, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballester, A.R.; Lafuente, M.T.; González-Candelas, L. Spatial study of antioxidant enzymes, peroxidase and phenylalanine ammonia-lyase in the citrus fruit-Penicillium digitatum interaction. Postharvest Biol. Technol. 2006, 39, 115–124. [Google Scholar] [CrossRef]
- Ballester, A.-R.; López-Pérez, M.; de la Fuente, B.; González-Candelas, L. Functional and pharmacological analyses of the role of Penicillium digitatum proteases on virulence. Microorganisms 2019, 7, 198. [Google Scholar] [CrossRef] [Green Version]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: A multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 2004, 5, 113. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Frandsen, R.J.; Andersson, J.A.; Kristensen, M.B.; Giese, H. Efficient four fragment cloning for the construction of vectors for targeted gene replacement in filamentous fungi. BMC Mol. Biol. 2008, 9, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Pérez, M.; Ballester, A.-R.; González-Candelas, L. Identification and functional analysis of Penicillium digitatum genes putatively involved in virulence towards citrus fruit. Mol. Plant Pathol. 2015, 16, 262–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lafuente, M.T.; Ballester, A.-R.; González-Candelas, L. Involvement of abscisic acid in the resistance of citrus fruit to Penicillium digitatum infection. Postharvest Biol. Technol. 2019, 154, 31–40. [Google Scholar] [CrossRef]
- Marcos, J.F.; González-Candelas, L.; Zacarías, L. Involvement of ethylene biosynthesis and perception in the susceptibility of citrus fruit to Penicillium digitatum infection and the accumulation of defense-related mRNAs. J. Exp. Bot. 2005, 56, 2183–2193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCollum, G.; Maul, P. 1-Methylcyclopropene inhibits degreening but stimulates respiration and ethylene biosynthesis in grapefruit. Hortscience 2007, 42, 120–124. [Google Scholar] [CrossRef] [Green Version]
- El-Kazzaz, M.K.; Sommer, N.F.; Kader, A.A. Ethylene effects on in vitro and in vivo growth of certain post-harvest fruit-infecting fungi. Phytopathology 1983, 73, 998–1001. [Google Scholar] [CrossRef]
- Flaishman, M.A.; Kolattukudy, P.E. Timing of fungal invasion using host’s ripening hormone as a signal. Proc. Natl. Acad. Sci. USA 1994, 91, 6579–6583. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.-J.; Kakuta, Y.; Sugawara, M.; Igarashi, T.; Oki, N.; Kisaki, M.; Shoji, T.; Kanetuna, Y.; Horita, T.; Matsui, H.; et al. Synthesis and degradation of 1-aminocyclopropane-1-carboxylic acid by Penicillium citrinum. Biosci. Biotechnol. Biochem. 1999, 63, 542–549. [Google Scholar] [CrossRef]
- Kakuta, Y.; Igarashi, T.; Murakami, T.; Ito, H.; Matsui, H.; Honma, M. 1-aminocyclopropane-1-carboxylate synthase of Penicillium citrinum: Primary structure and expression in Escherichia coli and Saccharomyces cerevisiae. Biosci. Biotechnol. Biochem. 2001, 65, 1511–1518. [Google Scholar] [CrossRef]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.B.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Varga, J.; Yaguchi, T.; Samson, R.A. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.J.; Ito, H.; Matsui, H.; Honma, M. 1-aminocyclopropane-1-carboxylate (ACC) deaminase induced by ACC synthesized and accumulated in Penicillium citrinum intracellular spaces. Biosci. Biotechnol. Biochem. 2000, 64, 299–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chagué, V.; Danit, L.-V.; Siewers, V.; Gronover, C.S.; Tudzynski, P.; Tudzynski, B.; Sharon, A. Ethylene sensing and gene activation in Botrytis cinerea: A missing link in ethylene regulation of fungus-plant interactions? Mol. Plant. Microbe Interact. 2006, 19, 33–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, P.; Xu, Z.; Cui, Z.; Zhang, Z.; Xu, L. Ethylene production by Alternaria alternata and its association with virulence on inoculated grape berries. Phytoparasitica 2017, 45, 273–279. [Google Scholar] [CrossRef]
- Zhu, P.; Xu, L.; Zhang, C.; Toyoda, H.; Gan, S.-S. Ethylene produced by Botrytis cinerea can affect early fungal development and can be used as a marker for infection during storage of grapes. Postharvest Biol. Technol. 2012, 66, 23–29. [Google Scholar] [CrossRef]
- Robison, M.M.; Griffith, M.; Pauls, K.P.; Glick, B.R. Dual role for ethylene in susceptibility of tomato to Verticillium wilt. J. Phytopathol. 2001, 149, 385–388. [Google Scholar] [CrossRef]
- Yang, Q.; Qian, X.; Dhanasekaran, S.; Boateng, N.A.S.; Yan, X.; Zhu, H.; He, F.; Zhang, H. Study on the infection mechanism of Penicillium digitatum on postharvest citrus (Citrus reticulata Blanco) based on transcriptomics. Microorganisms 2019, 7, 672. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | 2 dpi | 3 dpi | 4 dpi | 6 dpi |
|---|---|---|---|---|
| Pd1 | 12.3 ab | 23.0 b | 31.4 ab | 53.0 a |
| ect | 11.4 b | 21.1 c | 30.9 b | 53.0 a |
| Δ1f | 11.6 b | 22.8 b | 32.1 ab | 53.0 a |
| Δ23c | 13.3 a | 24.3 a | 33.8 a | 53.0 a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballester, A.-R.; González-Candelas, L. EFE-Mediated Ethylene Synthesis Is the Major Pathway in the Citrus Postharvest Pathogen Penicillium digitatum during Fruit Infection. J. Fungi 2020, 6, 175. https://doi.org/10.3390/jof6030175
Ballester A-R, González-Candelas L. EFE-Mediated Ethylene Synthesis Is the Major Pathway in the Citrus Postharvest Pathogen Penicillium digitatum during Fruit Infection. Journal of Fungi. 2020; 6(3):175. https://doi.org/10.3390/jof6030175
Chicago/Turabian StyleBallester, Ana-Rosa, and Luis González-Candelas. 2020. "EFE-Mediated Ethylene Synthesis Is the Major Pathway in the Citrus Postharvest Pathogen Penicillium digitatum during Fruit Infection" Journal of Fungi 6, no. 3: 175. https://doi.org/10.3390/jof6030175
APA StyleBallester, A.-R., & González-Candelas, L. (2020). EFE-Mediated Ethylene Synthesis Is the Major Pathway in the Citrus Postharvest Pathogen Penicillium digitatum during Fruit Infection. Journal of Fungi, 6(3), 175. https://doi.org/10.3390/jof6030175