Susceptibility of the Candida haemulonii Complex to Echinocandins: Focus on Both Planktonic and Biofilm Life Styles and a Literature Review





Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Growth Conditions
2.2. Determination of Minimal Inhibitory Concentration (MIC)
2.3. Echinocandins’ Breakpoints
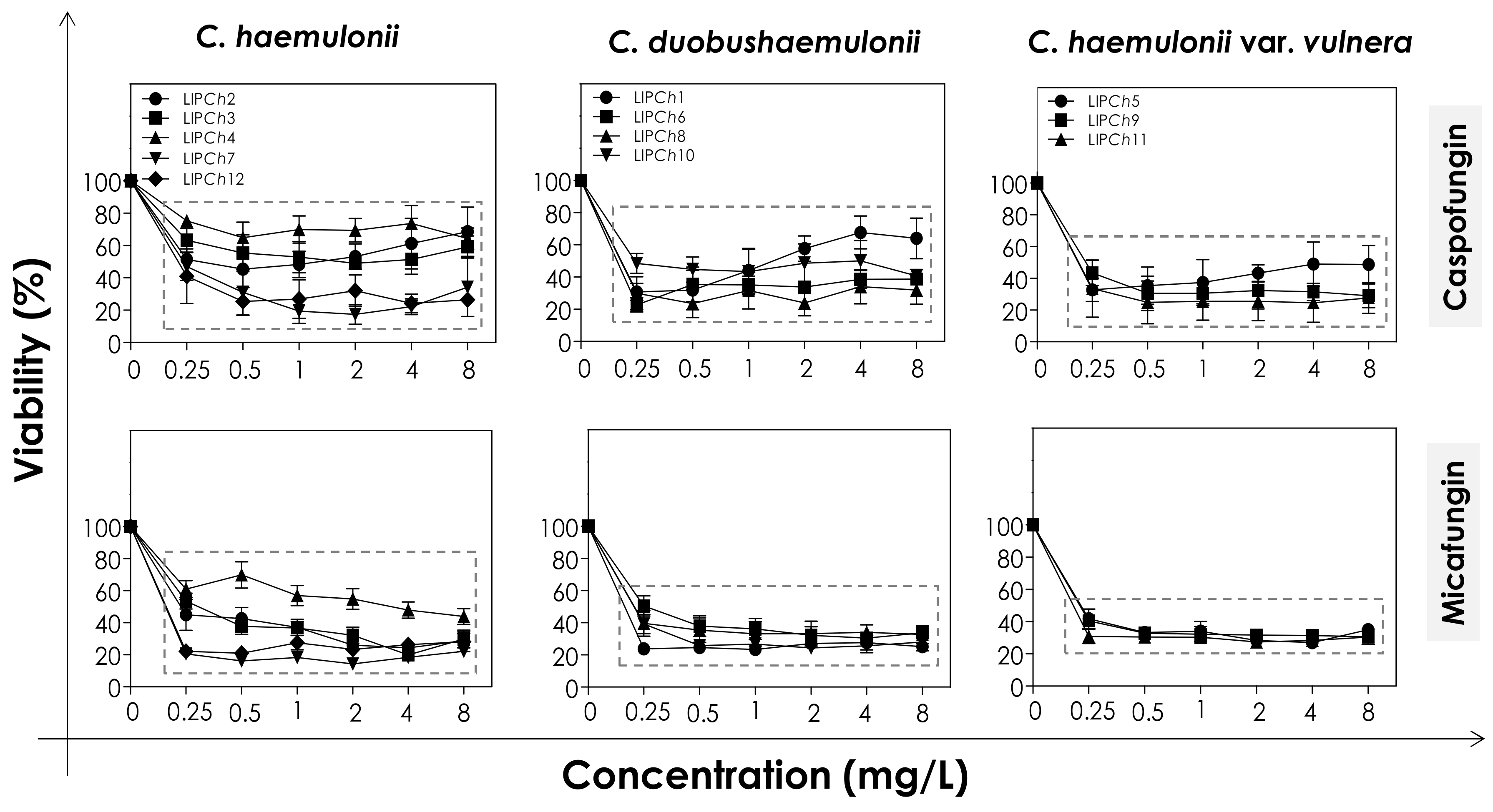
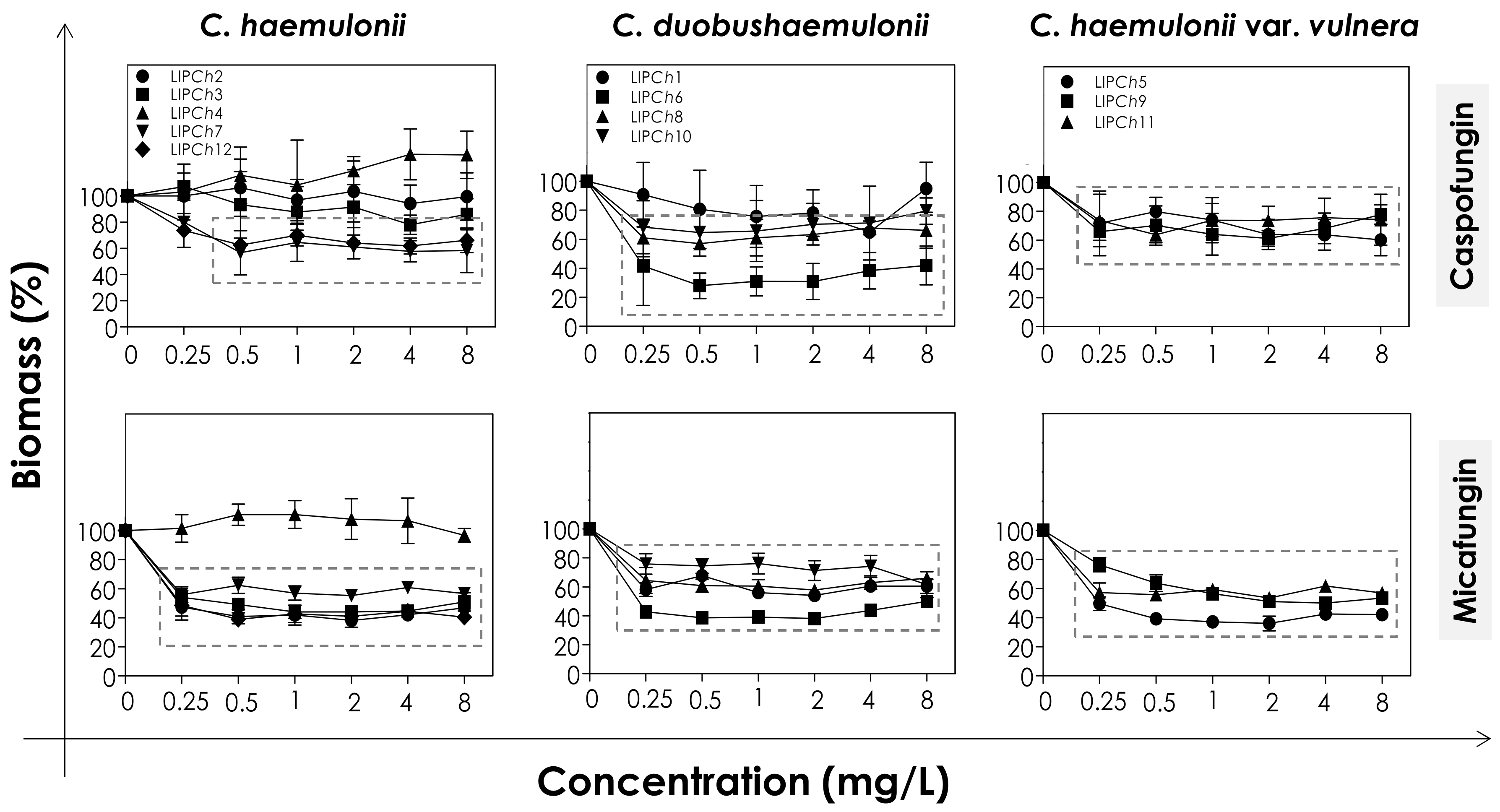
2.4. Effects of Echinocandins on the Biofilm Formed by the C. haemulonii Species Complex
2.4.1. Viability Assay
2.4.2. Biomass Measurement
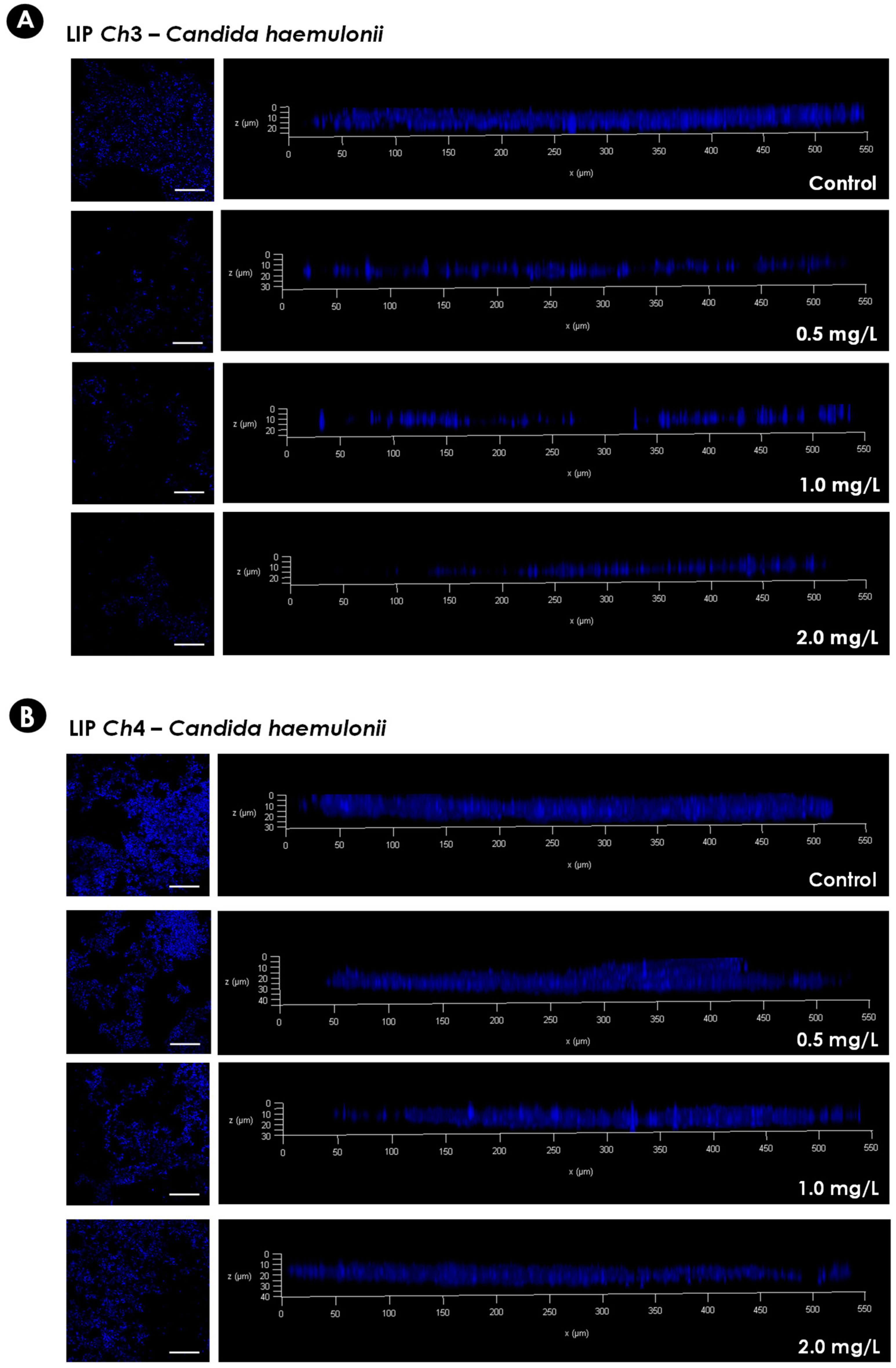
2.5. Biofilm Architecture: Confocal Laser Scanning Microscopy (CLSM) Assay
2.6. Literature Review
2.7. Statistics
3. Results and Discussion
3.1. Susceptibility of Planktonic Cells of the C. haemulonii Species Complex to Echinocandins
3.2. Effects of Echinocandins on the Biofilm Formed by C. haemulonii Species Complex
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Khan, Z.U.; Al-Sweih, N.A.; Ahmad, S.; Al-Kazemi, N.; Khan, S.; Joseph, L.; Chandy, R. Outbreak of fungemia among neonates caused by Candida haemulonii resistant to amphotericin B, itraconazole, and fluconazole. J. Clin. Microbiol. 2007, 45, 2025–2027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaller, M.A.; Diekema, D.J.; Gibbs, D.L.; Newell, V.A.; Ellis, D.; Tullio, V.; Rodloff, A.; Fu, W.; Ling, T.A. Results from the ARTEMIS DISK Global Antifungal Surveillance Study, 1997 to 2007: A 10.5-year analysis of susceptibilities of Candida Species to fluconazole and voriconazole as determined by CLSI standardized disk diffusion. J. Clin. Microbiol. 2010, 48, 1366–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Lopez, A.; Buitrago, M.J.; Rodriguez-Tudela, J.L.; Cuenca-Estrella, M. in vitro antifungal susceptibility pattern and ergosterol content in clinical yeast strains. Rev. Iberoam. Micol. 2011, 28, 100–103. [Google Scholar] [CrossRef]
- Kim, S.; Ko, K.S.; Moon, S.Y.; Lee, M.S.; Lee, M.Y.; Son, J.S. Catheter-related candidemia caused by Candida haemulonii in a patient in long-term hospital care. J. Korean Med. Sci. 2011, 26, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Cendejas-Bueno, E.; Kolecka, A.; Alastruey-Izquierdo, A.; Theelen, B.; Groenewald, M.; Kostrzewa, M.; Cuenca-Estrella, M.; Gomez-Lopez, A.; Boekhout, T. Reclassification of the Candida haemulonii complex as Candida haemulonii (C. haemulonii group I), C. duobushaemulonii sp. nov. (C. haemulonii group II), and C. haemulonii var. vulnera var. nov.: Three multiresistant human pathogenic yeasts. J. Clin. Microbiol. 2012, 50, 3641–3651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos, L.S.; Figueiredo-Carvalho, M.H.; Barbedo, L.S.; Ziccardi, M.; Chaves, A.L.; Zancope-Oliveira, R.M.; Pinto, M.R.; Sgarbi, D.B.; Dornelas-Ribeiro, M.; Branquinha, M.H.; et al. Candida haemulonii complex: Species identification and antifungal susceptibility profiles of clinical isolates from Brazil. J. Antimicrob. Chemother. 2015, 70, 111–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, X.; Xiao, M.; Chen, S.C.; Wang, H.; Cheng, J.W.; Chen, X.X.; Xu, Z.P.; Fan, X.; Kong, F.; Xu, Y.C. Identification and antifungal susceptibility profiles of Candida haemulonii species complex clinical isolates from a Multicenter Study in China. J. Clin. Microbiol. 2016, 54, 2676–2680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shapiro, R.S.; Robbins, N.; Cowen, L.E. Regulatory circuitry governing fungal development, drug resistance, and disease. Microbiol. Mol. Biol. Rev. 2011, 75, 213–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical Practice Guideline for the management of candidiasis: 2016 update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2015, 62, e1–e50. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.C.; Slavin, M.A.; Sorrell, T.C. Echinocandin antifungal drugs in fungal infections: A comparison. Drugs 2011, 71, 11–41. [Google Scholar] [CrossRef]
- Kuhn, D.M.; George, T.; Chandra, J.; Mukherjee, P.K.; Ghannoum, M.A. Antifungal susceptibility of Candida biofilms: Unique efficacy of amphotericin B lipid formulations and echinocandins. Antimicrob. Agents Chemother. 2002, 46, 1773–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.W.; Shin, J.H.; Jung, S.I.; Park, K.H.; Cho, D.; Kee, S.J.; Shin, M.G.; Suh, S.P.; Ryang, D.W. Species-specific differences in the susceptibilities of biofilms formed by Candida bloodstream isolates to echinocandin antifungals. Antimicrob. Agents Chemother. 2007, 51, 1520–1523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazzell, A.L.; Chaturvedi, A.K.; Pierce, C.G.; Prasad, D.; Uppuluri, P.; Lopez-Ribot, J.L. Treatment and prevention of Candida albicans biofilms with caspofungin in a novel central venous catheter murine model of candidiasis. J. Antimicrob. Chemother. 2009, 64, 567–570. [Google Scholar] [CrossRef] [PubMed]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts, CLSI Standard M27, 4th ed.; CLSI: Wayne, PA, USA, 2017. [Google Scholar]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts. Third Informational Supplement, CLSI Document M27-S3; CLSI: Wayne, PA, USA, 2018; Volume 28. [Google Scholar]
- CLSI. Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts. Fourth Informational Supplement, CLSI Document M27S4; CLSI: Wayne, PA, USA, 2012. [Google Scholar]
- CLSI. CLSI Document M60. In Performance Standards for Antifungal Susceptibility Testing of Yeasts, 2nd ed.; CLSI: Wayne, PA, USA, 2020; Volume 40. [Google Scholar]
- Ramos, L.S.; Oliveira, S.S.C.; Souto, X.M.; Branquinha, M.H.; Santos, A.L.S. Planktonic growth and biofilm formation profiles in Candida haemulonii species complex. Med. Mycol. 2017, 55, 785–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romera, D.; Aguilera-Correa, J.J.; Gadea, I.; Vinuela-Sandoval, L.; Garcia-Rodriguez, J.; Esteban, J. Candida auris: A comparison between planktonic and biofilm susceptibility to antifungal drugs. J. Med. Microbiol. 2019, 68, 1353–1358. [Google Scholar] [CrossRef] [PubMed]
- Peeters, E.; Nelis, H.J.; Coenye, T. Comparison of multiple methods for quantification of microbial biofilms grown in microtiter plates. J. Microbiol. Methods 2008, 72, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Ramos, L.S.; Mello, T.P.; Branquinha, M.H.; Santos, A.L.S. Biofilm formed by Candida haemulonii species complex: Structural analysis and extracellular matrix composition. J. Fungi 2020, 6, 46. [Google Scholar] [CrossRef] [Green Version]
- Chandra, J.; Mukherjee, P.K.; Ghannoum, M.A. In vitro growth and analysis of Candida biofilms. Nat. Protoc. 2008, 3, 1909–1924. [Google Scholar] [CrossRef]
- Ramage, G.; Rajendran, R.; Sherry, L.; Williams, C. Fungal biofilm resistance. Int. J. Microbiol. 2012, 2012, 528521. [Google Scholar] [CrossRef]
- Beauvais, A.; Schmidt, C.; Guadagnini, S.; Roux, P.; Perret, E.; Henry, C.; Paris, S.; Mallet, A.; Prevost, M.C.; Latge, J.P. An extracellular matrix glues together the aerial-grown hyphae of Aspergillus fumigatus. Cell. Microbiol. 2007, 9, 1588–1600. [Google Scholar] [CrossRef]
- Reséndiz-Sánchez, J.; Ortiz-Alvarez, J.; Casimiro-Ramos, A.; Hernandez-Rodriguez, C.; Villa-Tanaca, L. First report of a catheter-related bloodstream infection by Candida haemulonii in a children’s hospital in Mexico City. Int. J. Infect. Dis. 2020, 92, 123–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.N.; Shin, J.H.; Sung, H.; Lee, K.; Kim, E.C.; Ryoo, N.; Lee, J.S.; Jung, S.I.; Park, K.H.; Kee, S.J.; et al. Candida haemulonii and closely related species at 5 university hospitals in Korea: Identification, antifungal susceptibility, and clinical features. Clin. Infect. Dis. 2009, 48, e57–e61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruan, S.Y.; Kuo, Y.W.; Huang, C.T.; Hsiue, H.C.; Hsueh, P.R. Infections due to Candida haemulonii: Species identification, antifungal susceptibility and outcomes. Int. J. Antimicrob. Agents 2010, 35, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Crouzet, J.; Sotto, A.; Picard, E.; Lachaud, L.; Bourgeois, N. A case of Candida haemulonii osteitis: Clinical features, biochemical characteristics, and antifungal resistance profile. Clin. Microbiol. Infect. 2011, 17, 1068–1070. [Google Scholar] [CrossRef] [Green Version]
- De Almeida, J.N., Jr.; Assy, J.G.; Levin, A.S.; Del Negro, G.M.; Giudice, M.C.; Tringoni, M.P.; Thomaz, D.Y.; Motta, A.L.; Abdala, E.; Pierroti, L.C.; et al. Candida haemulonii complex species, Brazil, January 2010–March 2015. Emerg. Infect. Dis. 2016, 22, 561–563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muro, M.D.; Motta Fde, A.; Burger, M.; Melo, A.S.; Dalla-Costa, L.M. Echinocandin resistance in two Candida haemulonii isolates from pediatric patients. J. Clin. Microbiol. 2012, 50, 3783–3785. [Google Scholar] [CrossRef] [Green Version]
- Almeida, J.N., Jr.; Motta, A.L.; Rossi, F.; Abdala, E.; Pierrotti, L.C.; Kono, A.S.; Diz Mdel, P.; Benard, G.; Del Negro, G.M. First report of a clinical isolate of Candida haemulonii in Brazil. Clinics 2012, 67, 1229–1231. [Google Scholar] [CrossRef]
- Kathuria, S.; Singh, P.K.; Sharma, C.; Prakash, A.; Masih, A.; Kumar, A.; Meis, J.F.; Chowdhary, A. Multidrug-resistant Candida auris misidentified as Candida haemulonii: Characterization by matrix-assisted laser desorption ionization-time of flight mass spectrometry and DNA sequencing and its antifungal susceptibility profile variability by Vitek 2, CLSI broth microdilution, and Etest method. J. Clin. Microbiol. 2015, 53, 1823–1830. [Google Scholar] [CrossRef] [Green Version]
- Kumar, A.; Prakash, A.; Singh, A.; Kumar, H.; Hagen, F.; Meis, J.F.; Chowdhary, A. Candida haemulonii species complex: An emerging species in India and its genetic diversity assessed with multilocus sequence and amplified fragment-length polymorphism analyses. Emerg Microbes Infect. 2016, 5, e49. [Google Scholar] [CrossRef] [Green Version]
- Fang, S.Y.; Wei, K.C.; Chen, W.C.; Lee, S.J.; Yang, K.C.; Wu, C.S.; Sun, P.L. Primary deep cutaneous candidiasis caused by Candida duobushaemulonii in a 68-year-old man: The first case report and literature review. Mycoses 2016, 59, 818–821. [Google Scholar] [CrossRef]
- Ben-Ami, R.; Berman, J.; Novikov, A.; Bash, E.; Shachor-Meyouhas, Y.; Zakin, S.; Maor, Y.; Tarabia, J.; Schechner, V.; Adler, A.; et al. Multidrug-resistant Candida haemulonii and C. auris, Tel Aviv, Israel. Emerg. Infect. Dis. 2017, 23. [Google Scholar] [CrossRef] [PubMed]
- Isla, G.; Taverna, C.G.; Szusz, W.; Vivot, W.; García-Effron, G.; Davel, G. Candida haemulonii sensu lato: Update of the determination of susceptibility profile in Argentina and literature review. Curr. Fungal Infect. Rep. 2017, 11, 203–208. [Google Scholar] [CrossRef]
- Ramos, R.; Caceres, D.H.; Perez, M.; Garcia, N.; Castillo, W.; Santiago, E.; Borace, J.; Lockhart, S.R.; Berkow, E.L.; Hayer, L.; et al. Emerging multidrug-resistant Candida duobushaemulonii infections in Panama hospitals: Importance of laboratory surveillance and accurate identification. J. Clin. Microbiol. 2018, 56, e00371-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz, J.F.; Gade, L.; Chow, N.A.; Loparev, V.N.; Juieng, P.; Berkow, E.L.; Farrer, R.A.; Litvintseva, A.P.; Cuomo, C.A. Genomic insights into multidrug-resistance, mating and virulence in Candida auris and related emerging species. Nat. Commun. 2018, 9, 5346. [Google Scholar] [CrossRef] [Green Version]
- Frías-De-León, M.G.; Martinez-Herrera, E.; Acosta-Altamirano, G.; Arenas, R.; Rodriguez-Cerdeira, C. Superficial candidosis by Candida duobushaemulonii: An emerging microorganism. Infect. Genet. Evol. 2019, 75, 103960. [Google Scholar] [CrossRef]
- Zhang, H.; Niu, Y.; Tan, J.; Liu, W.; Sun, M.A.; Yang, E.; Wang, Q.; Li, R.; Wang, Y.; Liu, W. Global screening of genomic and transcriptomic factors associated with phenotype differences between multidrug-resistant and -susceptible Candida haemulonii strains. mSystems 2019, 4. [Google Scholar] [CrossRef] [Green Version]
- Di Luca, M.; Koliszak, A.; Karbysheva, S.; Chowdhary, A.; Meis, J.F.; Trampuz, A. Thermogenic characterization and antifungal susceptibility of Candida auris by microcalorimetry. J. Fungi 2019, 5, 103. [Google Scholar] [CrossRef] [Green Version]
- Bastos, R.W.; Rossato, L.; Valero, C.; Lagrou, K.; Colombo, A.L.; Goldman, G.H. Potential of Gallium as an antifungal agent. Front. Cell Infect. Microbiol. 2019, 9, 414. [Google Scholar] [CrossRef] [Green Version]
- Xiao, M.; Chen, S.C.; Kong, F.; Xu, X.L.; Yan, L.; Kong, H.S.; Fan, X.; Hou, X.; Cheng, J.W.; Zhou, M.L.; et al. Distribution and antifungal susceptibility of Candida species causing candidemia in China: An update from the CHIF-NET Study. J. Infect. Dis. 2020, 221, S139–S147. [Google Scholar] [CrossRef]
- Gade, L.; Muñoz, J.F.; Sheth, M.; Wagner, D.; Berkow, E.L.; Forsberg, K.; Jackson, B.R.; Ramos-Castro, R.; Escandón, P.; Dolande, M.; et al. Understanding the emergence of multidrug-resistant Candida: Using Whole-Genome Sequencing to describe the population structure of Candida haemulonii species complex. Front. Genet. 2020, 11, 554. [Google Scholar] [CrossRef]
- Lima, S.L.; Francisco, E.C.; De Almeida Júnior, J.N.; Santos, D.W.C.L.; Carlesse, F.; Queiroz-Telles, F.; Melo, A.S.A.; Colombo, A.L. Increasing prevalence of multidrug-resistant Candida haemulonii species complex among all yeast cultures collected by a reference laboratory over the past 11 years. J. Fungi 2020, 6, 110. [Google Scholar] [CrossRef] [PubMed]
- Coles, M.; Cox, K.; Chao, A. Candida haemulonii: An emerging opportunistic pathogen in the United States? IDCases 2020, 21, e00900. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, L.S.; Gazara, R.K.; Passarelli-Araujo, H.; Valengo, A.E.; Pontes, P.V.M.; Nunes-da-Fonseca, R.; De Souza, R.F.; Venancio, T.M.; Dalla-Costa, L.M. First genome sequences of two multidrug-resistant Candida haemulonii var. vulnera isolates from pediatric patients with candidemia. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Kordalewska, M.; Lee, A.; Park, S.; Berrio, I.; Chowdhary, A.; Zhao, Y.; Perlin, D.S. Understanding echinocandin resistance in the emerging pathogen Candida auris. Antimicrob. Agents Chemother. 2018, 62, e00238-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sherry, L.; Ramage, G.; Kean, R.; Borman, A.; Johnson, E.M.; Richardson, M.D.; Rautemaa-Richardson, R. Biofilm-forming capability of highly virulent, multidrug-resistant Candida auris. Emerg. Infect. Dis. 2017, 23, 328–331. [Google Scholar] [CrossRef] [Green Version]
- Kean, R.; Delaney, C.; Sherry, L.; Borman, A.; Johnson, E.M.; Richardson, M.D.; Rautemaa-Richardson, R.; Williams, C.; Ramage, G. Transcriptome assembly and profiling of Candida auris reveals novel insights into biofilm-mediated resistance. mSphere 2018, 3, e00334-18. [Google Scholar] [CrossRef] [Green Version]
- Guembe, M.; Guinea, J.; Marcos-Zambrano, L.J.; Fernandez-Cruz, A.; Pelaez, T.; Munoz, P.; Bouza, E. Micafungin at physiological serum concentrations shows antifungal activity against Candida albicans and Candida parapsilosis biofilms. Antimicrob. Agents Chemother. 2014, 58, 5581–5584. [Google Scholar] [CrossRef] [Green Version]
- Ziccardi, M.; Souza, L.O.; Gandra, R.M.; Galdino, A.C.; Baptista, A.R.; Nunes, A.P.; Ribeiro, M.A.; Branquinha, M.H.; Santos, A.L.S. Candida parapsilosis (sensu lato) isolated from hospitals located in the Southeast of Brazil: Species distribution, antifungal susceptibility and virulence attributes. Int. J. Med. Microbiol. 2015, 305, 848–859. [Google Scholar] [CrossRef]
- Simitsopoulou, M.; Peshkova, P.; Tasina, E.; Katragkou, A.; Kyrpitzi, D.; Velegraki, A.; Walsh, T.J.; Roilides, E. Species-specific and drug-specific differences in susceptibility of Candida biofilms to echinocandins: Characterization of less common bloodstream isolates. Antimicrob. Agents Chemother. 2013, 57, 2562–2570. [Google Scholar] [CrossRef] [Green Version]
- Melo, A.S.; Colombo, A.L.; Arthington-Skaggs, B.A. Paradoxical growth effect of caspofungin observed on biofilms and planktonic cells of five different Candida species. Antimicrob. Agents Chemother. 2007, 51, 3081–3088. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, J.A.; Carr, J.H.; Starling, C.E.; De Resende, M.A.; Donlan, R.M. Biofilm formation and effect of caspofungin on biofilm structure of Candida species bloodstream isolates. Antimicrob. Agents Chemother. 2009, 53, 4377–4384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsen, M.D.; Whyte, J.A.; Odds, F.C. Candida albicans and Candida dubliniensis respond differently to echinocandin antifungal agents in vitro. Antimicrob. Agents Chemother. 2007, 51, 1882–1884. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Fungal Species | MIC (mg/L) | |
|---|---|---|
| Isolates | Caspofungin b | Micafungin |
| C. haemulonii | ||
| LIPCh2 | 0.5 | 0.25 |
| LIPCh3 | 0.5 | 0.5 |
| LIPCh4 | 0.5 | 0.5 |
| LIPCh7 | 0.25 | 0.25 |
| LIPCh12 | 0.125 | 0.25 |
| GM-MIC a | 0.33 | 0.33 |
| Arithmetic mean | 0.37 | 0.35 |
| C. duobushaemulonii | ||
| LIPCh1 | 0.125 | 0.25 |
| LIPCh6 | 0.25 | 0.5 |
| LIPCh8 | 0.125 | 0.25 |
| LIPCh10 | 0.25 | 0.25 |
| GM-MIC | 0.18 | 0.30 |
| Arithmetic mean | 0.19 | 0.31 |
| C. haemulonii var. vulnera | ||
| LIPCh5 | 0.25 | 0.25 |
| LIPCh9 | 0.25 | 0.25 |
| LIPCh11 | 0.5 | 0.25 |
| GM-MIC | 0.32 | 0.25 |
| Arithmetic mean | 0.33 | 0.25 |
| Overall GM-MIC | 0.26 | 0.30 |
| Overall arithmetic mean | 0.30 | 0.31 |
| Database | Total Number of Papers | Number of Selected Papers * | References of the Selected Papers * |
|---|---|---|---|
| Web of Science | 148 | 18 | [5,7,25,29,31,32,33,34,35,36,37,38,39,40,41,42,43,44] |
| PubMed | 63 | 16 | [5,7,25,29,31,32,33,34,35,39,40,41,43,44,45,46] |
| Google Scholar | 46 | 13 | [5,7,25,29,31,32,33,35,36,40,44,46,47] |
| Scielo | 5 | 1 | [31] |
| Fungal Species | Susceptibility Profile (%) * | |||||
|---|---|---|---|---|---|---|
| Caspofungin | Micafungin | Anidulafungin | ||||
| S | NS | S | NS | S | NS | |
| C. haemulonii | 89.8 | 10.2 | 96.3 | 3.7 | 98.4 | 1.6 |
| n = 157 | n = 136 | n = 185 | ||||
| C. duobushaemulonii | 95.5 | 4.5 | 100 | 0 | 99.1 | 0.9 |
| n = 111 | n = 105 | n = 110 | ||||
| C. haemulonii var. vulnera | 85.0 | 15.0 | 91.7 | 8.3 | 97.1 | 2.9 |
| n = 20 | n = 12 | n = 35 | ||||
| Drug a Species | MIC (mg/L) | MIC50 b | MIC90 c | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ≤0.015 | 0.03 | 0.06 | 0.12 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | >16 | Range | |||
| CAS | |||||||||||||||
| Ch | 19 | 17 | 14 | 12 | 6 | 1 | 1 | 1 | 14 | 0.03–>16 | 0.12 | >16 | |||
| Cd | 3 | 14 | 18 | 20 | 9 | 4 | 1 | 1 | 1 | 3 | ≤0.015–>16 | 0.12 | 0.5 | ||
| Chv | 2 | 5 | 4 | 3 | 0.12–>16 | 0.25 | >16 | ||||||||
| MCF | |||||||||||||||
| Ch | 8 | 12 | 28 | 8 | 4 | 1 | 4 | ≤0.015–>16 | 0.06 | 0.25 | |||||
| Cd | 2 | 12 | 36 | 12 | 3 | 1 | 0.06–0.5 | 0.06 | 0.12 | ||||||
| Chv | 1 | 4 | 1 | 0.06–>16 | 0.12 | 0.12 | |||||||||
| ANF | |||||||||||||||
| Ch | 27 | 14 | 19 | 10 | 3 | 1 | 1 | 2 | ≤0.015–>16 | 0.03 | 0.12 | ||||
| Cd | 11 | 8 | 17 | 16 | 15 | 4 | 3 | 1 | 1 | ≤0.015–4 | 0.12 | 0.5 | |||
| Chv | 8 | 1 | 4 | 1 | ≤0.015–>16 | ≤0.015 | 0.06 | ||||||||
| Reference Number | Fungal Species (Number of Isolates) | GM-MIC (Range) * | ||
|---|---|---|---|---|
| Caspofungin | Micafungin | Anidulafungin | ||
| [5] • | Ch (n = 19) | 11.10 (#) (0.25–>16) | 0.17 (#) (<0.03–>16) | 0.06 (#) (<0.03–>16) |
| Cd (n = 7) | 5.38 (#) (0.5–>16) | 0.06 (0.06–0.12) | 0.08 (#) (<0.03–4) | |
| Chv (n = 4) | 11.31 (#) (0.5–16) | 0.40 (#) (0.06–>16) | 0.20 (#) (<0.03–>16) | |
| [29] • | Ch (n = 14) | 0.12 (0.125–0.5) | - | 0.015 (0.015–0.015) |
| Cd (n = 9) | 0.22 (#) (0.06–16) | - | 0.06 (0.015–0.5) | |
| Chv (n = 8) | 0.26 (0.125–0.5) | - | 0.016 (0.015–0.03) | |
| [33] • | Ch (n = 6)/Chv (n = 1) | 0.18 (0.06–1) | 0.27 (0.125–1) | 0.45 (0.25–1) |
| Cd (n = 8) | 0.13 (0.06–0.25) | 0.38 (0.125–1) | 0.54 (0.5–1) | |
| [7] • | Ch (n = 26) | ND (0.03–0.5) | ND (0.06–0.5) | ND (0.015–0.5) |
| Cd (n = 5) | ND (0.06–0.12) | ND (0.06–0.12) | ND (0.06–0.25) | |
| [35] ◦ | Ch (n = 3) | 0.5 (0.5–0.5) | 0.19 (0.12–0.5) | 0.03 (0.03–0.03) |
| [39] ◦ | Cd (n = 2) | - | 0.12 (0.06–0.25) | 0.04 (0.03–0.06) |
| [40] ◦ | Ch (n = 3) | 0.10 (0.06–0.125) | 0.20 (0.125–0.25) | - |
| [44] ◦ | Ch (n = 38) | 0.06 (#) (0.03–16) | 0.04 (<0.08–0.12) | 0.05 (0.03–0.25) |
| Cd (n = 55) | 0.07 (0.016–0.5) | 0.06 (0.016–0.25) | 0.13 (0.016–2) | |
| [32] • | Ch (n = 7) | 0.19 (0.06–1) | 0.28 (0.125–1) | 0.44 (0.25–1) |
| Cd (n = 5) | 0.14 (0.06–0.25) | 0.35 (0.125–1) | 0.56 (0.5–1) | |
| [45] • | Ch (n = 21) | - | - | 0.10 (0.06–0.25) |
| Cd (n = 13) | - | - | 0.10 (0.03–0.5) | |
| Chv (n = 15) | - | - | 0.13 (0.03–0.25) | |
| [43] • | Ch (n = 32) | 0.104 (ND) | 0.106 (ND) | 0.103 (ND) |
| [36] • | Ch (n = 16) | 0.13(#) (0.015–8) | 0.11(#) (0.03–8) | 0.09 (0.015–0.5) |
| Cd (n = 3) | 5.03(#) (1–16) | 0.06 (0.015–0.06) | 0.79 (0.5–2) | |
| Chv (n = 5) | 0.12 (0.06–0.25) | 0.14 (0.12–0.25) | 0.05 (0.015–0.12) | |
| [42] ◦ | Ch (n = 4) | 0.06 (0.03–0.12) | - | - |
| [47] ◦ | Chv (n = 2) | 0.25 (0.25–0.25) | 0.12 (0.12–0.12) | 0.06 (0.06–0.06) |
| Arithmetic mean of overall GM-MIC, except for the resistant strains(#) | ||||
| Ch | 0.18 ± 0.15 | 0.18 ± 0.09 | 0.16 ± 0.18 | |
| Cd | 0.11 ± 0.04 | 0.17 ± 0.15 | 0.32 ± 0.30 | |
| Chv | 0.21 ± 0.08 | 0.13 ± 0.01 | 0.06 ± 0.05 | |
| Echinocandins | MBEC (mg/L) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C. haemulonii Isolates | C. duobushaemulonii Isolates | C. haemulonii var. vulnera Isolates | ||||||||||
| Ch2 | Ch3 | Ch4 | Ch7 | Ch12 | Ch1 | Ch6 | Ch8 | Ch10 | Ch5 | Ch9 | Ch11 | |
| Caspofungin | 0.5 | 2 | ˃8 | 0.25 | 0.25 | ˂0.25 | ˂0.25 | ˂0.25 | 0.25 | 0.25 | 0.25 | 0.25 |
| Micafungin | 0.25 | 0.5 | 8 | ˂0.25 | ˂0.25 | ˂0.25 | 0.5 | 0.25 | 0.25 | 0.25 | ˂0.25 | ˂0.25 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramos, L.S.; Silva, L.N.; Branquinha, M.H.; Santos, A.L.S. Susceptibility of the Candida haemulonii Complex to Echinocandins: Focus on Both Planktonic and Biofilm Life Styles and a Literature Review. J. Fungi 2020, 6, 201. https://doi.org/10.3390/jof6040201
Ramos LS, Silva LN, Branquinha MH, Santos ALS. Susceptibility of the Candida haemulonii Complex to Echinocandins: Focus on Both Planktonic and Biofilm Life Styles and a Literature Review. Journal of Fungi. 2020; 6(4):201. https://doi.org/10.3390/jof6040201
Chicago/Turabian StyleRamos, Lívia S., Laura N. Silva, Marta H. Branquinha, and André L. S. Santos. 2020. "Susceptibility of the Candida haemulonii Complex to Echinocandins: Focus on Both Planktonic and Biofilm Life Styles and a Literature Review" Journal of Fungi 6, no. 4: 201. https://doi.org/10.3390/jof6040201
APA StyleRamos, L. S., Silva, L. N., Branquinha, M. H., & Santos, A. L. S. (2020). Susceptibility of the Candida haemulonii Complex to Echinocandins: Focus on Both Planktonic and Biofilm Life Styles and a Literature Review. Journal of Fungi, 6(4), 201. https://doi.org/10.3390/jof6040201