Role of Volatiles from the Endophytic Fungus Trichoderma asperelloides PSU-P1 in Biocontrol Potential and in Promoting the Plant Growth of Arabidopsis thaliana


 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Trichoderma Species and Pathogen Sources
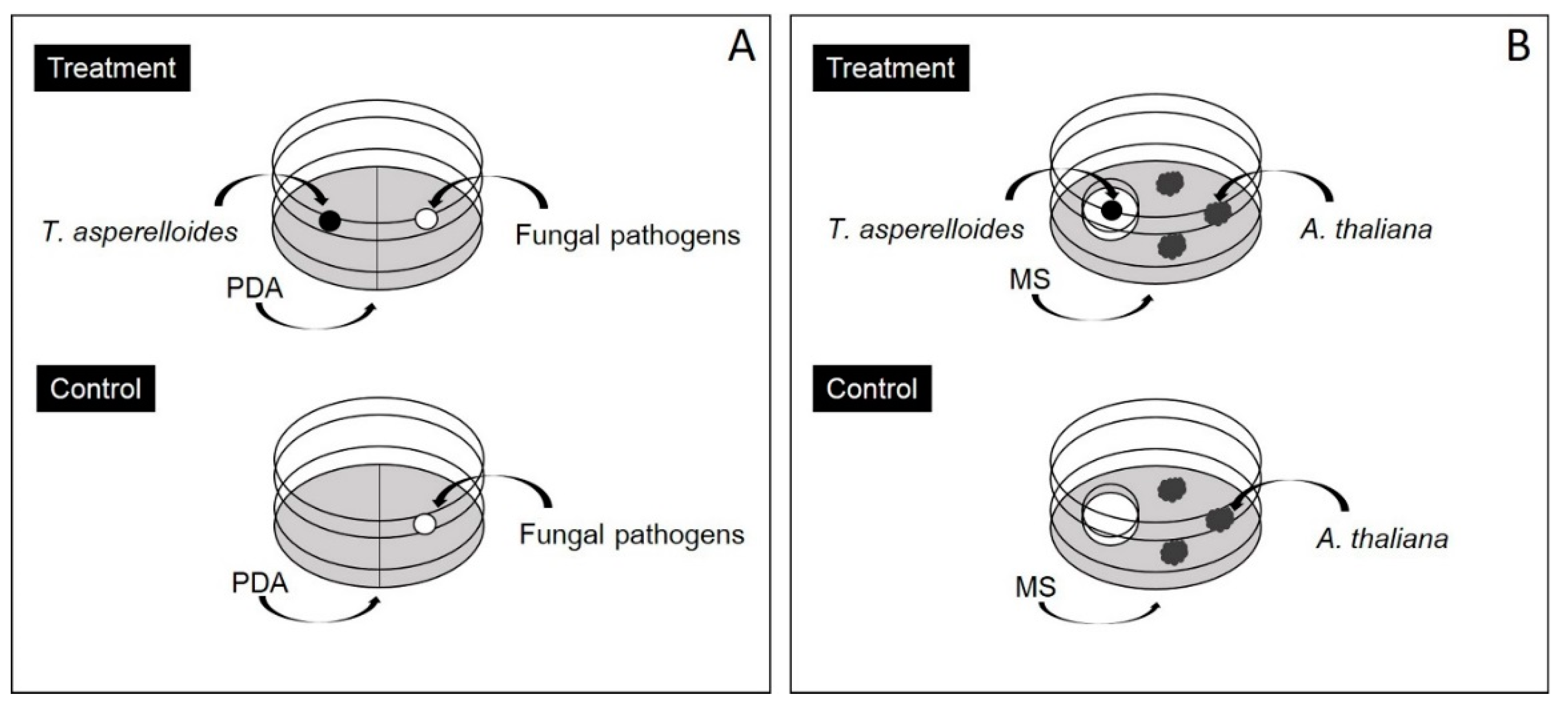
2.2. Volatile Antifungal Bioassay
2.3. Plant and Growth Conditions
2.4. Volatile Exposure Bioassay
2.5. Plant Chlorophyll Measurements
2.6. RNA Extraction and Quantitative Reverse Transcription PCR Analysis
2.7. Enzyme Assay
2.8. Gas Chromatography-Mass Spectrometryanalysis
2.9. Effect of Commercial VOCs on the Antifungal Activity, Defense Responses, and Growth of A. thaliana
2.10. Statistical Analysis
3. Results
3.1. VOCs Emitted by T. asperelloides PSU-P1 Inhibit Fungal Growth
3.2. VOCs Emitted by T. asperelloides PSU-P1 Increase Growth in A. thaliana
3.3. VOCs Induce Gene Expression of Defense-Related Genes
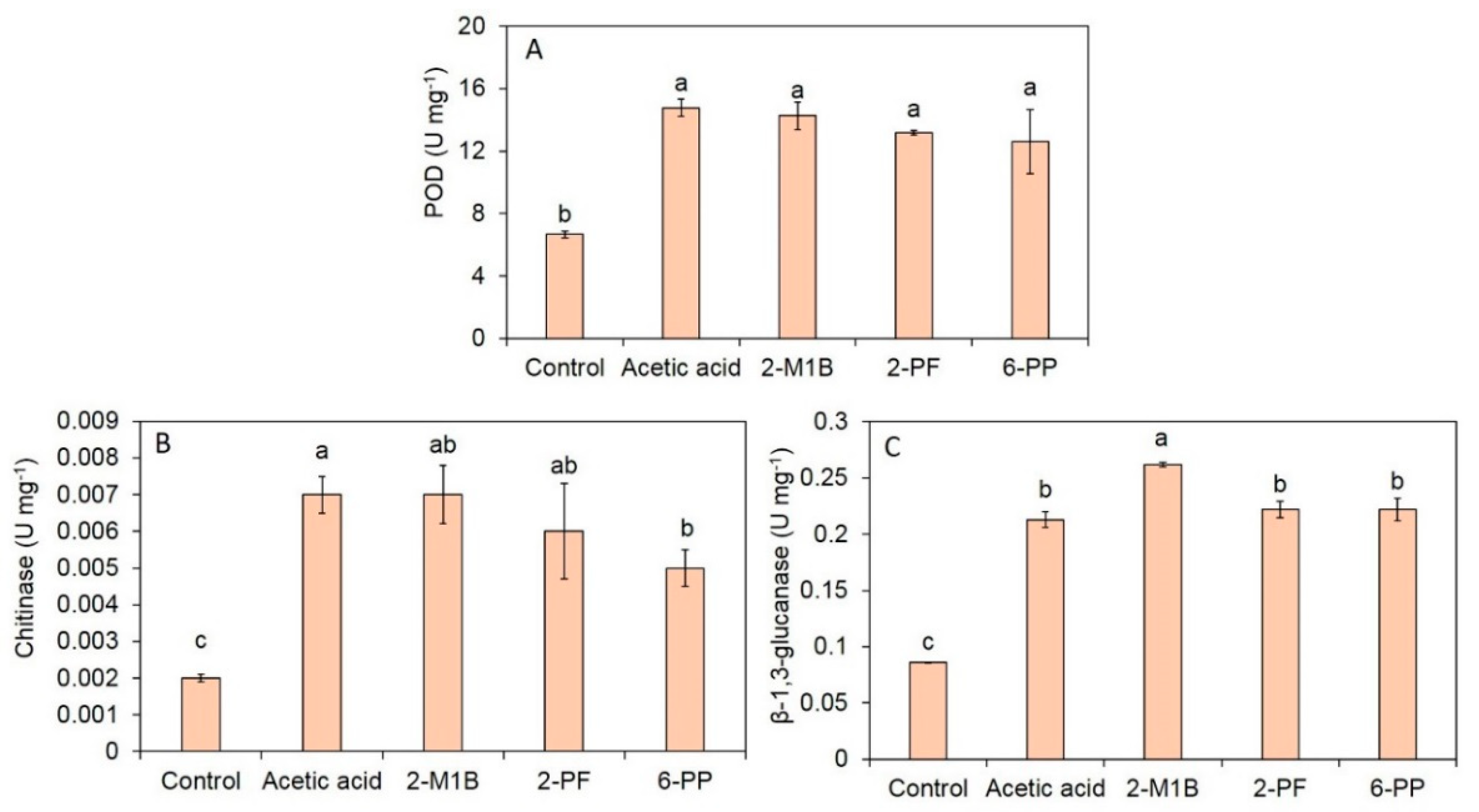
3.4. Induction of Defense-Related Enzymes and Enzyme Activities by Fungal VOCs
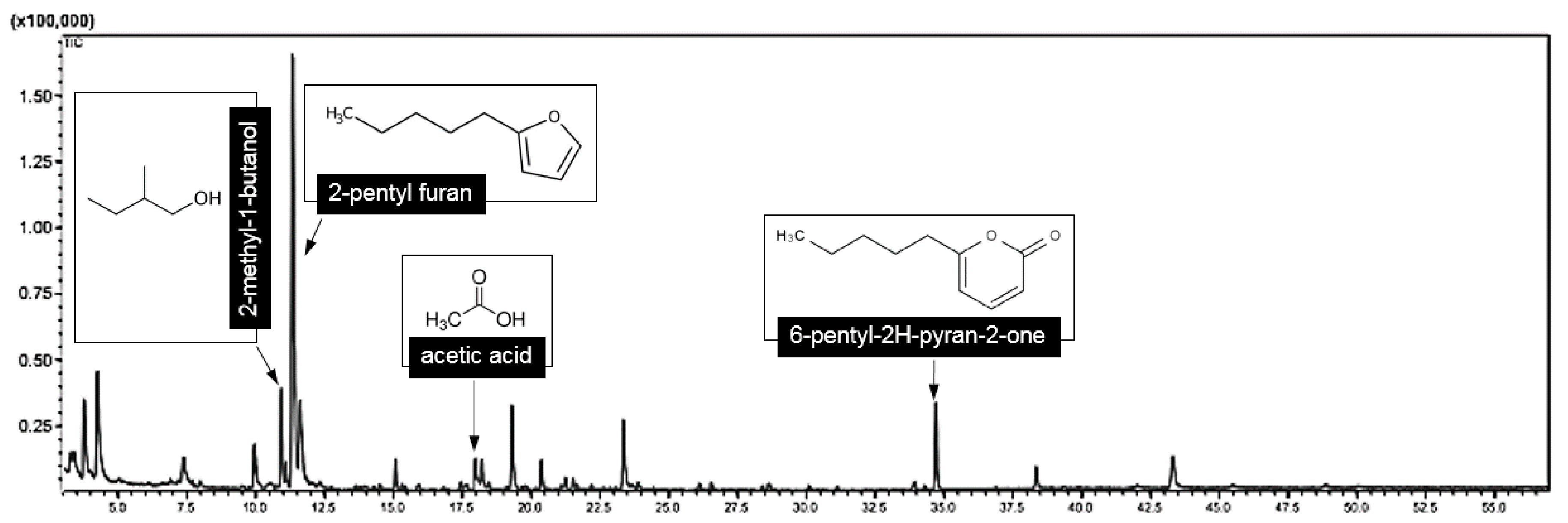
3.5. Identification of VOCs through SPME GC/MS
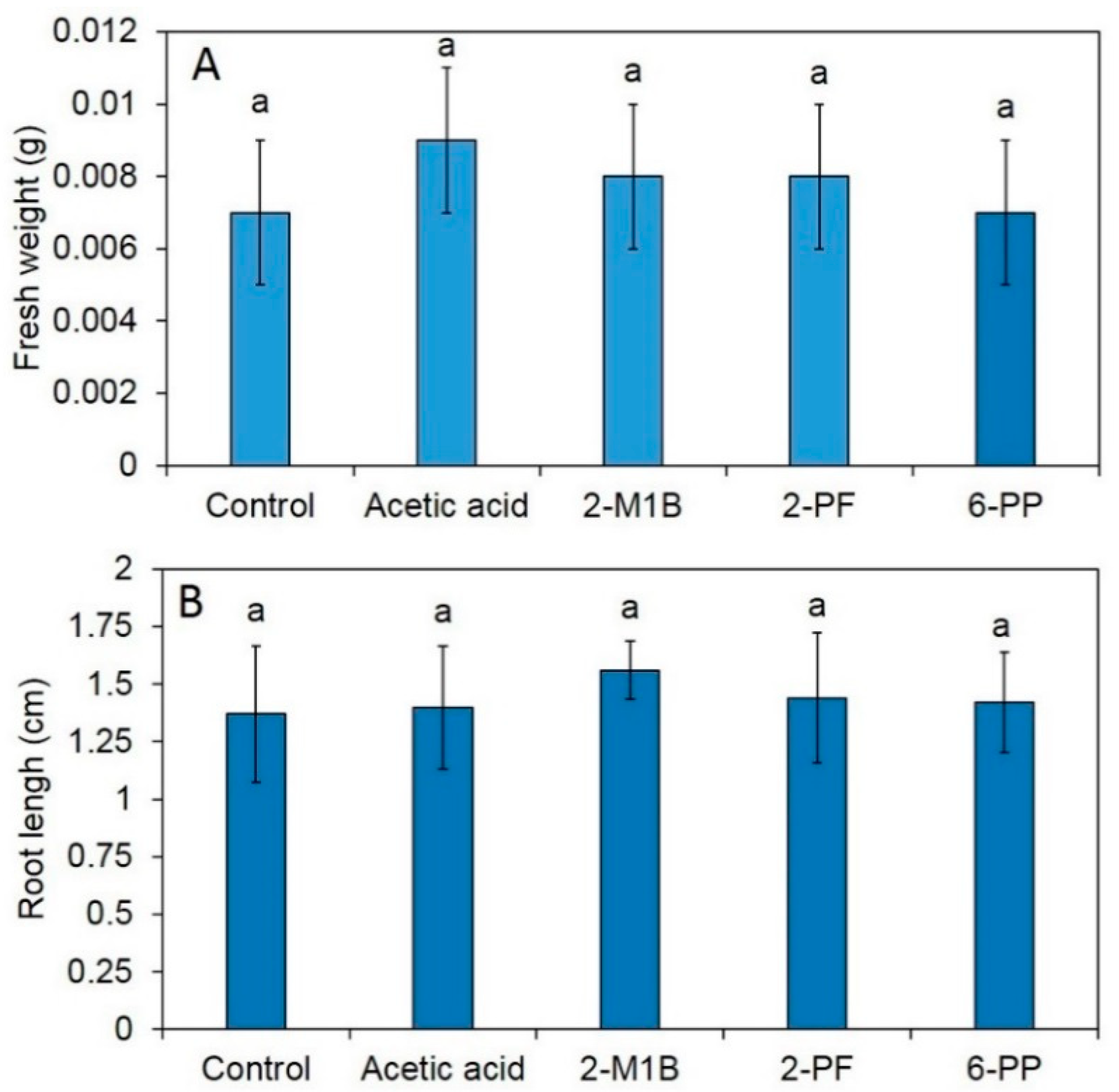
3.6. Effects of Commercial Volatile Compounds on Antifungal Activity, Induced Enzyme Activity, and Plant Phenotype
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shi, M.; Chen, L.; Wang, X.W.; Zhang, T.; Zhao, P.B.; Song, X.Y.; Sun, C.Y.; Chen, X.L.; Zhou, B.C.; Zhang, Y.Z. Antimicrobial peptaibols from Trichoderma pseudokoningii induce programmed cell death in plant fungal pathogens. Microbiology 2012, 158, 166–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunpapao, A.; Chairin, T.; Ito, S. The biocontrol by Streptomyces and Trichoderma of leaf spot disease caused by Curvularia oryzae in oil palm seedlings. Biol. Control 2018, 123, 36–42. [Google Scholar] [CrossRef]
- Wonglom, P.; Daengsuwan, W.; Ito, S.; Sunpapao, A. Biological control of Sclerotium fruit rot of snake fruit and stem rot of lettuce by Trichoderma sp. T76-12/2 and the mechanism involved. Physiol. Molec. Plant Pathol. 2019, 107, 1–7. [Google Scholar] [CrossRef]
- Baiyee, B.; Ito, S.; Sunapapo, A. Trichoderma asperellum T1 mediated antifungal activity and induced defense response against leaf spot fungi in lettuce (Lactuca sativa L.). Physiol. Molec. Plant Pathol. 2019, 106, 96–101. [Google Scholar] [CrossRef]
- Wonglom, P.; Ito, S.; Sunpapao, A. Volatile organic compounds emitted from endophytic fungus Trichoderma asperellum T1 mediate antifungal activity, defense response and promote plant growth in lettuce (Lactuca sativa). Fungal Ecol. 2020, 43, 100867. [Google Scholar] [CrossRef]
- Vinodkumar, S.; Indumathi, T.; Nakkeeran, S. Trichoderma asperellum (NVTA2) as a potential antagonist for the management of stem rot in carnation under protected cultivation. Biol. Control 2017, 113, 58–64. [Google Scholar] [CrossRef]
- Baiyee, B.; Pornsuriya, C.; Ito, S.; Sunapapo, A. Trichoderma spirale T76-1 displays biocontrol activity on lettuce (Lactuca sativa L.) caused by Corynespora cassiicola or Curvularia aeria. Biol. Control 2019, 129, 195–200. [Google Scholar] [CrossRef]
- Monteiro, V.; Silva, R.N.; Steindorff, A.; Costa, F.; Noronha, E.; Ricart, C.; de Sousa, M.; Vainstein, M.; Monteiro, V.; Ulhoa, C. New insight in; Trichoderma harzianum antagonism of fungal plant pathogens by secreted protein analysis. Curr. Microbiol. 2010, 61, 298–305. [Google Scholar] [CrossRef]
- Carreras-Villaseñor, N.; Sánchez-Arreguín, J.A.; Herrera-Estrella, A.H. Trichoderma: Sensing the environment for survival and dispersal. Microbiology 2012, 158, 3–16. [Google Scholar] [CrossRef]
- Evans, H.C.; Holmes, K.A.; Thomas, S.E. Endophytes and mycoparasites associated with an indigenous forest tree, Theobroma gileri, in Ecuador and a preliminary assessment of their potentials biocontrol agents of cocoa diseases. Mycol. Prog. 2003, 2, 149–160. [Google Scholar] [CrossRef]
- Brader, G.; Compant, S.; Mitter, B.; Trognitz, F.; Sessitsch, A. Metabolic potential of endophytic bacteria. Curr. Opin. Biotechnol. 2014, 27, 30–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macías-Rubalcava, M.L.; Hernández-Bautista, B.E.; Oropeza, F.; Duarte, G.; González, M.C.; Glenn, A.E.; Hanlin, R.T.; Anaya, A.L. Allelochemical effects of volatile compounds and organic extracts from Muscodor yucatanensis, a tropical endophytic fungus from Bursera simaruta. J. Chem. Ecol. 2010, 36, 1122–1131. [Google Scholar] [CrossRef] [PubMed]
- Insam, H.; Seewald, S.A. Volatile organic compounds (VOCs) in soils. Biol. Fertil. Soils 2010, 46, 199–213. [Google Scholar] [CrossRef] [Green Version]
- Dudareva, N.; Negre, F.; Nagegowda, D.A.; Orlova, I. Plant volatiles: Recent advances and future perspectives. Crit. Rev. Plant Sci. 2006, 25, 417–440. [Google Scholar] [CrossRef]
- Campos, V.P.; Pinho, R.S.C.; Freire, E.S. Volatiles produced by interacting microorganisms potentially useful for the control of plant pathogens. Cienc. Agrotec. 2010, 34, 525–535. [Google Scholar] [CrossRef] [Green Version]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma-plant pathogens interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Sumida, C.H.; Daniel, J.F.S.; Araujod, A.P.C.S.; Peitl, D.C.; Abreu, L.M.; Dekker, R.F.H.; Canteri, M.G. Trichoderma asperelloides antagonism to nine Sclerotinia sclerotiorum strains and biological control of white mold disease in soybean plants. Biocontrol Sci. Technol. 2018, 28, 142–156. [Google Scholar] [CrossRef]
- Ruangwong, O.; Wonglom, P.; Phoka, N.; Suwannarach, N.; Lumyong, S.; Ito, S.; Sunpapao, A. Biological control activity of Trichoderma asperelloides PSU-P1 against gummy stem blight in muskmelon plant (Cucumis melo). Physiol. Molec. Plant Pathol. under review.
- Chairin, T.; Pornsuriya, C.; Thaochan, N.; Sunpapao, A. Corynespora cassiicola causes leaf spot disease on lettuce (Lactuca sativa) cultivated in hydroponic systems in Thailand. Australas. Plant Dis. Notes 2017, 12, 1–3. [Google Scholar] [CrossRef]
- Pornsuriya, C.; Ito, S.; Sunpapao, A. First report of leaf spot on lettuce caused by Curvularia aeria. J. Gen. Plant Pathol. 2018, 84, 296–299. [Google Scholar] [CrossRef]
- Pornsuriya, C.; Chitphithak, J. Blue mold caused by Penicillium oxalicum on muskmelon (Cucumis melo) in Thailand. Australas. Plant. Dis. Notes 2018, 13, 46. [Google Scholar] [CrossRef] [Green Version]
- Daengsuwan, W.; Wonglom, P.; Arikit, S.; Sunpapao, A. Morphological and molecular identification of the Neopestalotiopsis clavispora associated with flower blight on Anthurium andraeanum in Thailand. Hort. Plant J. accepted for publication.
- Koike, S.T. Southern blight of Jerusalem artichoke caused by Sclerotium rolfsii in California. Plant Dis. 2004, 88, 769. [Google Scholar] [CrossRef] [PubMed]
- Aveskamp, M.M.; de Gruyter, J.; Woudenberg, J.H.C.; Verkleyand, G.J.M.; Crous, P.W. Highlights of the Didymellaceae: A polyphasic approach to characterize Phoma and related pleosporalean genera. Stud. Mycol. 2010, 65, 1–60. [Google Scholar] [CrossRef] [PubMed]
- Dennis, C.; Webster, J. Antagonistic properties of species-groups of Trichoderma, II. Production of volatile antibiotic. Trans. Br. Mycol. Soc. 1971, 57, 41–48. [Google Scholar] [CrossRef]
- Prapagdee, B.; Akrapikulchart, U.; Mongkolsuk, S. Potential of a soil-borne Streptomyces hygroscopicus for biocontrol of Anthracnose disease caused by Colletotrichum gloeosporioides in orchard. J. Biol. Sci. 2008, 8, 1187–1192. [Google Scholar] [CrossRef] [Green Version]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assay with tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Moran, R. Formulae for determination of chlorophyllous pigments extracted with N,N-dimethylformamide. Plant Physiol. 1982, 69, 1376–1381. [Google Scholar] [CrossRef] [Green Version]
- Kaewsuksaeng, S.; Tatmala, N.; Shigyo, M.; Tanaka, S.; Yamauchi, N. Application of electrostatic atomized water particle suppress calyx discoloration in relation to postharvest quality of mangosteen (Garcinia mangostana L.). Sci. Hort. 2019, 250, 380–387. [Google Scholar] [CrossRef]
- Dumhai, R.; Wanchana, S.; Saensuk, C.; Choowongkomon, K.; Mahatheeranont, S.; Kraithong, T.; Toojinda, T.; Vanavichit, A.; Arikit, S. Discovery of a novel CnAMADH2 allele associated with higher levels of 2-acetyl-1-pyroline (2AP) in yellow dwarf coconut (Cocos nucifera L.). Sci. Hort. 2019, 243, 490–497. [Google Scholar] [CrossRef]
- Chairin, T.; Petcharat, V. Induction of defense responses in longkong fruit (Aglaia dookkoo Griff.) against fruit rot fungi by Metarhizium guizhouense. Biol. Control 2017, 111, 40–44. [Google Scholar] [CrossRef]
- Vetter, J.L.; Steinberg, M.P.; Nelson, A.I. Quantitative determination of peroxidase in sweet corn. J. Agric. Food Chem. 1958, 6, 39–41. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Annl. Biochem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Suwannarach, N.; Kaewyana, C.; Yodmeeklin, A.; Kumla, J.; Lumyong, S. Evaluation of Muscodor cinnamomi as an egg biofumigant for the reduction of microorganisms on egg shell surfaces and its effect on egg quality. Int. J. Food Microbiol. 2017, 244, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Parker, S.R.; Cutler, H.G.; Jacyno, J.M.; Hill, R.A. Biological activities of 6-pentyl-2H-pyran-2-one and its analogs. J. Agric. Food Chem. 1997, 45, 2774–2776. [Google Scholar] [CrossRef]
- Naznin, H.A.; Kiyohara, D.; Kimura, M.; Miyazawa, M.; Shimizu, M.; Hyakumachi, M. Systemic resisatnce induced by volatile organic compounds emitted by plant growth-promoting fungi in Arabidopsis thaliana. PLoS ONE 2014, 9, e86882. [Google Scholar] [CrossRef] [Green Version]
- Sunpapao, A. Antagonistic Microorganisms: Current Research and Innovations; Lambert Academic Publishing: Saarbrücken, Germany, 2020; p. 120. [Google Scholar]
- Rogawansmamy, S.; Gaskin, S.; Taylor, M.; Pisaniello, D. An evaluation of antifungal agents for the treatment of fungal contamination in indoor air environments. Int. J. Environ. Res. Public Health 2015, 12, 6319–6332. [Google Scholar] [CrossRef] [Green Version]
- Rezende, D.C.; Fialho, M.B.; Brand, S.C.; Blumer, S.; Pascholati, S.F. Antimicrobial activity of volatile organic compounds and their effect on lipid peroxidation and electrolyte loss in Colletotrichum gloeosporioides and Colletotrichum acutatum mycelia. Afr. J. Microbiol. Res. 2015, 9, 1527–1535. [Google Scholar]
- Andriamialisoa, Z.; Giraud, M.; Labia, R.; Valla, A. Chemical synthesis of 6-pentyl-2H-pyran-2-one: A natural antifungal biosynthesized by Trichoderma spp. Chem. Ecol. 2010, 20, 55–59. [Google Scholar] [CrossRef]
- Collins, R.P.; Halim, A.F. Characterization of the major aroma constituent of the fungus Trichoderma viride (Pers.). J. Agric. Food Chem. 1972, 20, 437–438. [Google Scholar] [CrossRef]
- Bonnarme, P.; Djian, A.; Latrasse, A.; Feron, G.; Ginies, C.; Durand, A.; Le Quéré, J.L. Production of 6-pentyl-a-pyrone by Trichoderma sp. from vegetable oils. J. Biotechnol. 1997, 156, 143–150. [Google Scholar] [CrossRef]
- Simon, A.; Dunlop, R.W.; Ghisalberti, E.L.; Sivasithamparam, K. Trichoderma koningii produces a pyrone compound with antibiotic properties. Soil Biol. Chem. 1988, 20, 263–264. [Google Scholar] [CrossRef]
- Reithner, B.; Brunner, K.; Schuhmacher, R.; Peissl, P.; Seidl, V.; Krska, R.; Zeilinger, S. The G protein α subunit Tga1 of Trichoderma atroviride is involved in chitinase formation and differential production of antifungal metabolites. Fungal Genet. Biol. 2005, 42, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Reithner, B.; Schuhmacher, R.; Stoppacher, N.; Pucher, M.; Brunner, K.; Zeilinger, S. Signaling via the Trichoderma atroviride mitogen-activated protein kinase Tmk1 differentially affects mycoparasitism and plant protection. Fungal Genet. Biol. 2007, 44, 1123–1133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.; Behringer, G.; Hung, R.; Bennett, J. Effects of fungal volatile organic compounds on Arabidopsis thaliana growth and gene expression. Fungal Ecol. 2019, 37, 1–9. [Google Scholar] [CrossRef]
- Lee, S.; Yap, M.; Behringer, G.; Hung, R.; Bennett, J.W. Volatile organic compounds emitted by Trichoderma species mediate plant growth. Fungal Biol. Biotechnol. 2016, 3, 7. [Google Scholar] [CrossRef] [Green Version]
- Jalali, F.; Zafari, D.; Salari, H. Volatile organic compounds of some Trichoderma spp. increase growth and induce salt tolerance in Arabidopsis thaliana. Fungal Ecol. 2017, 29, 67–75. [Google Scholar] [CrossRef]
- Zou, C.; Li, Z.; Yu, D. Bacillus megaterium strain XTBG34 promotes plant growth by producing 2-pentylfuran. J. Microbiol. 2010, 48, 460–466. [Google Scholar] [CrossRef]
- El-Hassan, A.; Buchennauer, H. Action of 6-penthyl-alpha pyrone in controlling seedling blight incited by Fusarium moniliforme and inducing defense responses in maize. J. Phytopathol. 2009, 157, 697–707. [Google Scholar] [CrossRef]
- Kim, J.-S.; Lee, J.; Lee, C.-H.; Woo, S.Y.; Kang, H.; Seo, S.-G.; Kim, S.-H. Activation of pathogenesis-related genes by the rhizobacterium, Bacillus sp. JS, which induces systemic resistance in tobacco plants. Plant. Pathol. J. 2015, 31, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.; Vaishnav, A.; Varma, A.; Choudhary, D.W. Comparative expression analysis of defence-related genes in Bacillus-treated Glycine max upon challenge inoculation with selective fungal phytopathogens. Curr. Sci. 2018, 115, 1950–1956. [Google Scholar] [CrossRef]
- Kottb, M.; Gigolashvili, T.; Großkinsky, D.K.; Piechulla, B. Trichoderma volatiles effecting Arabidopsis: From inhibition to protection against phytopathogenic fungi. Front. Microbial. 2015, 6, 995. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Disease | Reference | |
|---|---|---|---|
| Antagonist | Fungal Pathogen | ||
| Trichoderma asperelloides PSU-P1 | – | – | [18] |
| Colletotrichum sp. | Leaf blight | CCPM | |
| Corynespora cassiicola | Leaf spot | [19] | |
| Curvularia lunata | Leaf spot | [20] | |
| Ganoderma sp. | Basal stem rot | CCPM | |
| Penicillium oxalicum | Blue mold | [21] | |
| Neopestalotiopsis clavispora | Flower blight | [22] | |
| Sclerotium rolfsii | Southern blight | [23] | |
| Stagonosporopsis cucurbitacearum | Gummy stem blight | [24] | |
| Gene | Accession Number | Primer | Sequence (5′ → 3′) | Product Size (bp) |
|---|---|---|---|---|
| ACT | NM001036427 | ACT-F | CTCCCATTCCCTTCTCCTTC | 247 |
| ACT-R | CGAGGACGACCCACAATACT | |||
| CHI | NM129919 | Chi-F | TAGCTTCGGTGCTTCCATCT | 159 |
| Chi-R | GCACATGGGAACTCTGGTTT | |||
| GUL | NM115586 | Glu-F | TGGTGTCAGATTCCGGTACA | 192 |
| Glu-R | TCATCCCTGAACCTTCCTTG | |||
| POD | BT001238 | POD-F | ACCAACAGACCAGACCCAAG | 244 |
| POD-R | CGAACGTGTTGCTGCTGTAT |
| RT (min) | Volatile Compound | m/z | Formula | Similarity | Total Area |
|---|---|---|---|---|---|
| 10.93 | 2-methyl-1-butanol * | 88 | C5H12O | 95 | 6.96 |
| 11.34 | 2-pentyl furan | 138 | C9H14O | 96 | 30.59 |
| 17.98 | Acetic acid | 60 | C2H4O2 | 97 | 1.96 |
| 34.71 | 6-penthyl-2H-pyran-2-one | 108 | C10H14O2 | 97 | 5.45 |
| Fungal Pathogen | Disease | Antifungal Ability 1 | |||
|---|---|---|---|---|---|
| Acetic Acid | 2-PF | 2M1B | 6-PP | ||
| Colletotrichum sp. | Leaf blight | − | − | − | − |
| C. cassiicola | Leaf spot | − | − | − | − |
| C. lunata | Leaf spot | − | − | − | − |
| Ganoderma sp. | Basal stem rot | + | + | + | + |
| P. oxalicum | Blue mold | − | + | + | + |
| N. clavispora | flower blight | − | − | − | − |
| S. rolfsii | Southern blight | − | − | − | + |
| S. cucurbitacearum | Gummy stem blight | − | − | − | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Phoka, N.; Suwannarach, N.; Lumyong, S.; Ito, S.-i.; Matsui, K.; Arikit, S.; Sunpapao, A. Role of Volatiles from the Endophytic Fungus Trichoderma asperelloides PSU-P1 in Biocontrol Potential and in Promoting the Plant Growth of Arabidopsis thaliana. J. Fungi 2020, 6, 341. https://doi.org/10.3390/jof6040341
Phoka N, Suwannarach N, Lumyong S, Ito S-i, Matsui K, Arikit S, Sunpapao A. Role of Volatiles from the Endophytic Fungus Trichoderma asperelloides PSU-P1 in Biocontrol Potential and in Promoting the Plant Growth of Arabidopsis thaliana. Journal of Fungi. 2020; 6(4):341. https://doi.org/10.3390/jof6040341
Chicago/Turabian StylePhoka, Nongnat, Nakarin Suwannarach, Saisamorn Lumyong, Shin-ichi Ito, Kenji Matsui, Siwaret Arikit, and Anurag Sunpapao. 2020. "Role of Volatiles from the Endophytic Fungus Trichoderma asperelloides PSU-P1 in Biocontrol Potential and in Promoting the Plant Growth of Arabidopsis thaliana" Journal of Fungi 6, no. 4: 341. https://doi.org/10.3390/jof6040341