Sterol 14α-Demethylase Ligand-Binding Pocket-Mediated Acquired and Intrinsic Azole Resistance in Fungal Pathogens




Abstract
:1. Introduction
2. Azole Resistance in Fungal Pathogens
2.1. Structure of the SDM Ligand-Binding Pocket
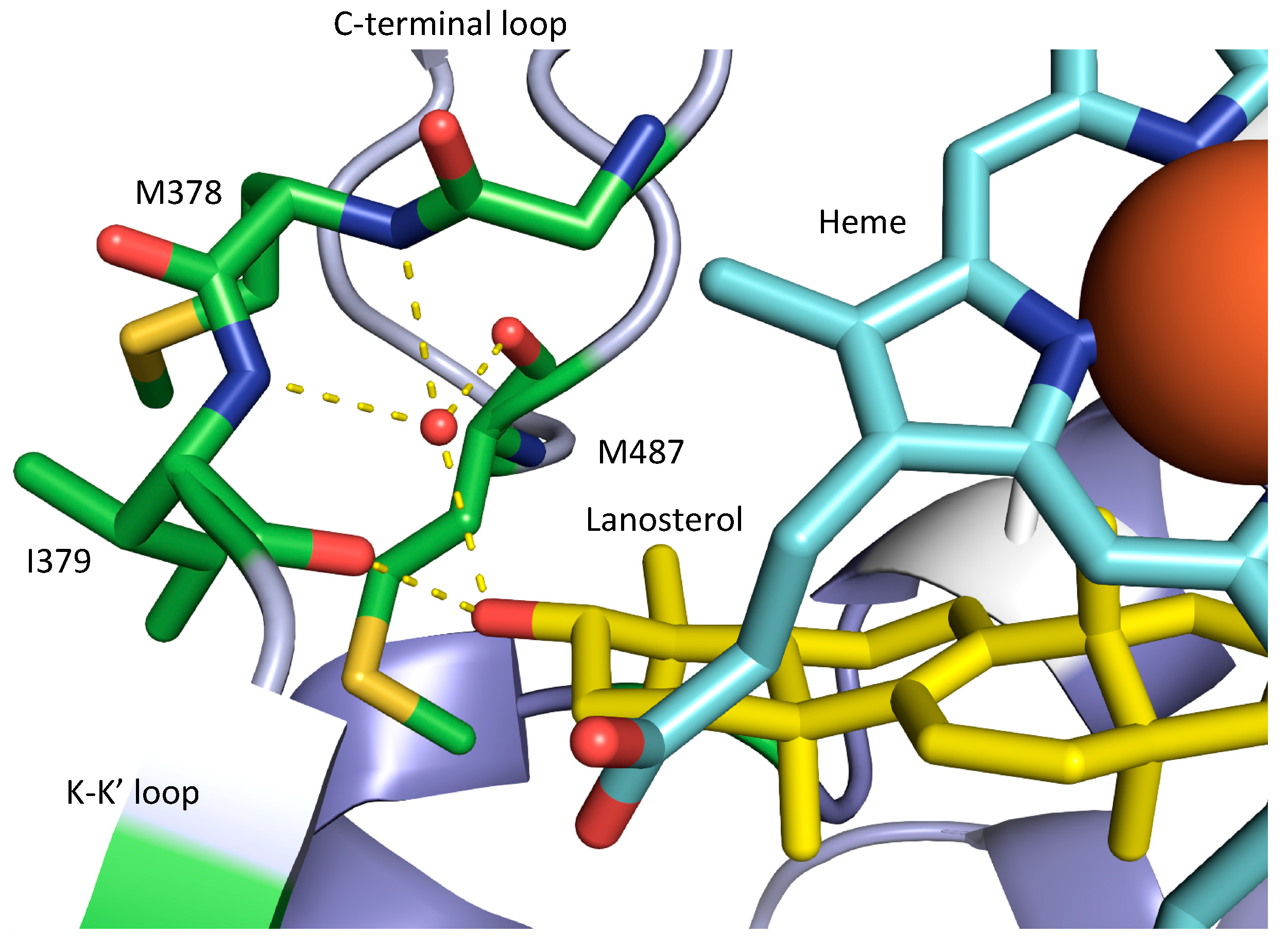
2.2. G73 in S. cerevisiae SDM
2.3. Y140 in S. cerevisiae SDM
2.3.1. Aspergillus Species
2.3.2. Candida Species
2.3.3. Cryptococcus Species
2.3.4. Saccharomyces cerevisiae
2.3.5. Scedosporium species
2.3.6. Ajellomyces capsulatum
2.3.7. Innate Azole Resistance of Mucormycetes
2.3.8. Agricultural Pathogens
2.4. K151 (According to S. cerevisiae SDM)
2.5. T322 (According to S. cerevisiae SDM)
2.5.1. Aspergillus fumigatus
2.5.2. Candida albicans
2.6. G464 (According to S. cerevisiae SDM)
2.6.1. Glycine G464S/D
2.6.2. Glycine G460 in M. graminicola
3. Discussion
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kathiravan, M.K.; Salake, A.B.; Chothe, A.S.; Dudhe, P.B.; Watode, R.P.; Mukta, M.S.; Gadhwe, S. The biology and chemistry of antifungal agents: A review. Bioorg. Med. Chem. 2012, 20, 5678–5698. [Google Scholar] [CrossRef] [PubMed]
- Mast, N.; Zheng, W.; Stout, C.D.; Pikuleva, I.A. Antifungal azoles: Structural insights into undesired tight binding to cholesterol-metabolizing cyp46a1. Mol. Pharmacol. 2013, 84, 86–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashyap, A.; Silakari, O. Chapter 9—Triazoles: Multidimensional 5-membered nucleus for designing multitargeting agents. In Key Heterocycle Cores for Designing Multitargeting Molecules; Silakari, O., Ed.; Elsevier: Amsterdam, The Netherlands, 2018; pp. 323–342. [Google Scholar]
- Fisher, M.C.; Hawkins, N.J.; Sanglard, D.; Gurr, S.J. Worldwide emergence of resistance to antifungal drugs challenges human health and food security. Science 2018, 360, 739–742. [Google Scholar] [CrossRef] [Green Version]
- Barber, A.E.; Riedel, J.; Sae-Ong, T.; Kang, K.; Brabetz, W.; Panagiotou, G.; Deising, H.B.; Kurzai, O. Effects of agricultural fungicide use on Aspergillus fumigatus abundance, antifungal susceptibility, and population structure. BioRxiv 2020. [Google Scholar] [CrossRef]
- Brauer, V.S.; Rezende, C.P.; Pessoni, A.M.; De Paula, R.G.; Rangappa, K.S.; Nayaka, S.C.; Gupta, V.K.; Almeida, F. Antifungal agents in agriculture: Friends and foes of public health. Biomolecules 2019, 9, 521. [Google Scholar] [CrossRef] [Green Version]
- Warrilow, A.G.S.; Hull, C.M.; Rolley, N.J.; Parker, J.E.; Nes, W.D.; Smith, S.N.; Kelly, D.E.; Kelly, S.L. Clotrimazole as a potent agent for treating the oomycete fish pathogen Saprolegnia parasiticathrough inhibition of sterol 14α-demethylase (cyp51). Appl. Environ. Microbiol. 2014, 80, 6154–6166. [Google Scholar] [CrossRef] [Green Version]
- Baumgardner, D.J. Fungal infections from human and animal contact. J. Patient-Cent. Res. Rev. 2017, 4, 78–89. [Google Scholar]
- Pham, D.K.; Chu, J.; Do, N.T.; Brose, F.; Degand, G.; Delahaut, P.; De Pauw, E.; Douny, C.; Van Nguyen, K.; Vu, T.D.; et al. Monitoring antibiotic use and residue in freshwater aquaculture for domestic use in vietnam. EcoHealth 2015, 12, 480–489. [Google Scholar] [CrossRef] [Green Version]
- Barber, A.E.; Scheufen, S.; Walther, G.; Kurzai, O.; Schmidt, V. Low rate of azole resistance in cases of avian aspergillosis in germany. Med. Mycol. 2020, 58, 1187–1190. [Google Scholar] [CrossRef]
- Kano, R.; Okubo, M.; Yanai, T.; Hasegawa, A.; Kamata, H. First isolation of azole-resistant Cryptococcus neoformans from feline cryptococcosis. Mycopathologia 2015, 180, 427–433. [Google Scholar] [CrossRef]
- Seyedmousavi, S.; Bosco, S.M.G.; de Hoog, S.; Ebel, F.; Elad, D.; Gomes, R.R.; Jacobsen, I.D.; Jensen, H.E.; Martel, A.; Mignon, B.; et al. Fungal infections in animals: A patchwork of different situations. Med. Mycol. 2018, 56, 165–187. [Google Scholar] [CrossRef]
- Caldas, S.S.; Arias, J.L.O.; Rombaldi, C.; Mello, L.L.; Cerqueira, M.B.R.; Martins, A.F.; Primel, E.G. Occurrence of pesticides and PPCPS in surface and drinking water in southern brazil: Data on 4-year monitoring. J. Braz. Chem. Soc. 2019, 30, 71–80. [Google Scholar] [CrossRef]
- Campestre, C.; Locatelli, M.; Guglielmi, P.; De Luca, E.; Bellagamba, G.; Menta, S.; Zengin, G.; Celia, C.; Di Marzio, L.; Carradori, S. Analysis of imidazoles and triazoles in biological samples after microextraction by packed sorbent. J. Enzym. Inhib. Med. Chem. 2017, 32, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shafiei, M.; Peyton, L.; Hashemzadeh, M.; Foroumadi, A. History of the development of antifungal azoles: A review on structures, sar, and mechanism of action. Bioorganic Chem. 2020, 104, 104240. [Google Scholar] [CrossRef] [PubMed]
- Warrilow, A.G.; Nishimoto, A.T.; Parker, J.E.; Price, C.L.; Flowers, S.A.; Kelly, D.E.; Rogers, P.D.; Kelly, S.L. The evolution of azole resistance in Candida albicans sterol 14α-demethylase (cyp51) through incremental amino acid substitutions. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef] [Green Version]
- Schoustra, S.; Debets, A.J.M.; Rijs, A.J.M.M.; Zhang, J.; Snelders, E.; Leendertse, P.; Melchers, W.J.G.; Rietveld, A.; Zwaan, B.; Verweij, P. Environmental hotspots for azole resistance selection of Aspergillus fumigatus, the netherlands. Emerg. Infect. Dis. J. 2019, 25, 1347. [Google Scholar] [CrossRef] [Green Version]
- Verweij, P.E.; Chowdhary, A.; Melchers, W.J.; Meis, J.F. Azole resistance in Aspergillus fumigatus: Can we retain the clinical use of mold-active antifungal azoles? Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2016, 62, 362–368. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yang, H.; Turra, D.; Zhou, S.; Ayhan, D.H.; DeIulio, G.A.; Guo, L.; Broz, K.; Wiederhold, N.; Coleman, J.J.; et al. The genome of opportunistic fungal pathogen fusarium oxysporum carries a unique set of lineage-specific chromosomes. Commun. Biol. 2020, 3, 50. [Google Scholar] [CrossRef] [Green Version]
- Keniya, M.V.; Sabherwal, M.; Wilson, R.K.; Woods, M.A.; Sagatova, A.A.; Tyndall, J.D.A.; Monk, B.C. Crystal structures of full-length lanosterol 14α-demethylases of prominent fungal pathogens Candida albicans and Candida glabrata provide tools for antifungal discovery. Antimicrob. Agents Chemother. 2018, 62, e01134-18. [Google Scholar] [CrossRef] [Green Version]
- Keniya, M.V.; Ruma, Y.N.; Tyndall, J.D.A.; Monk, B.C. Heterologous expression of full-length lanosterol 14α-demethylases of prominent fungal pathogens Candida albicans and Candida glabrata provides tools for antifungal discovery. Antimicrob. Agents Chemother. 2018, 62, e01131-18. [Google Scholar]
- Balkis, M.M.; Leidich, S.D.; Mukherjee, P.K.; Ghannoum, M.A. Mechanisms of fungal resistance: An overview. Drugs 2002, 62, 1025–1040. [Google Scholar] [CrossRef]
- Monk, B.C.; Sagatova, A.A.; Hosseini, P.; Ruma, Y.N.; Wilson, R.K.; Keniya, M.V. Fungal lanosterol 14α-demethylase: A target for next-generation antifungal design. Biochim. Biophys. Acta Proteins Proteom. 2020, 1868, 140206. [Google Scholar] [CrossRef] [PubMed]
- Lackner, M.; Tscherner, M.; Schaller, M.; Kuchler, K.; Mair, C.; Sartori, B.; Istel, F.; Arendrup, M.C.; Lass-Flörl, C. Positions and numbers of fks mutations in Candida albicans selectively influence in vitro and in vivo susceptibilities to echinocandin treatment. Antimicrob. Agents Chemother. 2014, 58, 3626–3635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Cantero, A.; López-Fernández, L.; Guarro, J.; Capilla, J. Azole resistance mechanisms in Aspergillus: Update and recent advances. Int. J. Antimicrob. Agents 2020, 55, 105807. [Google Scholar] [CrossRef] [PubMed]
- Teo, J.Q.; Lee, S.J.; Tan, A.L.; Lim, R.S.; Cai, Y.; Lim, T.P.; Kwa, A.L. Molecular mechanisms of azole resistance in Candida bloodstream isolates. Bmc Infect. Dis. 2019, 19, 63. [Google Scholar] [CrossRef] [PubMed]
- Magobo, R.E.; Lockhart, S.R.; Govender, N.P. Fluconazole-resistant Candida parapsilosis strains with a Y132F substitution in the ERG11 gene causing invasive infections in a neonatal unit, south africa. Mycoses 2020, 63, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Monk, B.C.; Tomasiak, T.M.; Keniya, M.V.; Huschmann, F.U.; Tyndall, J.D.; O’Connell, J.D., 3rd; Cannon, R.D.; McDonald, J.G.; Rodriguez, A.; Finer-Moore, J.S.; et al. Architecture of a single membrane spanning cytochrome p450 suggests constraints that orient the catalytic domain relative to a bilayer. Proc. Natl. Acad. Sci. USA 2014, 111, 3865–3870. [Google Scholar] [CrossRef] [Green Version]
- Patel, S.B. Plant sterols and stanols: Their role in health and disease. J. Clin. Lipidol. 2008, 2, S11–S19. [Google Scholar] [CrossRef] [Green Version]
- Sagatova, A.A.; Keniya, M.V.; Wilson, R.K.; Monk, B.C.; Tyndall, J.D. Structural insights into binding of the antifungal drug fluconazole to Saccharomyces cerevisiae lanosterol 14α-demethylase. Antimicrob. Agents Chemother. 2015, 59, 4982–4989. [Google Scholar] [CrossRef] [Green Version]
- Snelders, E.; Camps, S.M.; Karawajczyk, A.; Rijs, A.J.; Zoll, J.; Verweij, P.E.; Melchers, W.J. Genotype-phenotype complexity of the TR46/Y121F/T289A cyp51A azole resistance mechanism in Aspergillus fumigatus. Fungal Genet. Biol. Fg B 2015, 82, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.J.; Liu, W.L.; Lai, C.C.; Chao, C.M.; Ko, W.C.; Wang, H.C.; Dai, C.T.; Hsieh, M.I.; Choi, P.C.; Yang, J.L.; et al. Multicenter study of azole-resistant Aspergillus fumigatus clinical isolates, taiwan (1). Emerg. Infect. Dis. 2020, 26, 804–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lavergne, R.A.; Morio, F.; Danner-Boucher, I.; Horeau-Langlard, D.; David, V.; Hagen, F.; Meis, J.F.; Le Pape, P. One year prospective survey of azole resistance in Aspergillus fumigatus at a french cystic fibrosis reference centre: Prevalence and mechanisms of resistance. J. Antimicrob. Chemother. 2019, 74, 1884–1889. [Google Scholar] [CrossRef] [PubMed]
- Resendiz-Sharpe, A.; Mercier, T.; Lestrade, P.P.A.; van der Beek, M.T.; von dem Borne, P.A.; Cornelissen, J.J.; De Kort, E.; Rijnders, B.J.A.; Schauwvlieghe, A.; Verweij, P.E.; et al. Prevalence of voriconazole-resistant invasive Aspergillosis and its impact on mortality in haematology patients. J. Antimicrob. Chemother. 2019, 74, 2759–2766. [Google Scholar] [CrossRef] [PubMed]
- Rivero-Menendez, O.; Soto-Debran, J.C.; Medina, N.; Lucio, J.; Mellado, E.; Alastruey-Izquierdo, A. Molecular identification, antifungal susceptibility testing, and mechanisms of azole resistance in Aspergillus species received within a surveillance program on antifungal resistance in spain. Antimicrob. Agents Chemother. 2019, 63, e00865-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonardelli, F.; Macedo, D.; Dudiuk, C.; Cabeza, M.S.; Gamarra, S.; Garcia-Effron, G. Aspergillus fumigatus intrinsic fluconazole resistance is due to the naturally occurring T301I substitution in cyp51ap. Antimicrob. Agents Chemother. 2016, 60, 5420–5426. [Google Scholar] [CrossRef] [Green Version]
- Alcazar-Fuoli, L.; Cuesta, I.; Rodriguez-Tudela, J.L.; Cuenca-Estrella, M.; Sanglard, D.; Mellado, E. Three-dimensional models of 14α-sterol demethylase (cyp51a) from Aspergillus lentulus and Aspergillus fumigatus: An insight into differences in voriconazole interaction. Int. J. Antimicrob. Agents 2011, 38, 426–434. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Perlin, D.S. Establishing surrogate markers for fluconazole resistance in Candida albicans. Microb. Drug Resist. (Larchmt. N. Y.) 2005, 11, 232–238. [Google Scholar] [CrossRef]
- Whaley, S.G.; Berkow, E.L.; Rybak, J.M.; Nishimoto, A.T.; Barker, K.S.; Rogers, P.D. Azole antifungal resistance in Candida albicans and emerging non-albicans Candida species. Front. Microbiol. 2016, 7, 2173. [Google Scholar] [CrossRef] [Green Version]
- Accoceberry, I.; Rougeron, A.; Biteau, N.; Chevrel, P.; Fitton-Ouhabi, V.; Noël, T. A CTG Candida yeast genetically engineered for the genotype-phenotype characterization of azole antifungal resistance in human pathogenic yeasts. Antimicrob. Agents Chemother. 2017, 62, e01483-17. [Google Scholar] [CrossRef] [Green Version]
- Lamb, D.C.; Kelly, D.E.; Schunck, W.H.; Shyadehi, A.Z.; Akhtar, M.; Lowe, D.J.; Baldwin, B.C.; Kelly, S.L. The mutation T315A in Candida albicans sterol 14alpha-demethylase causes reduced enzyme activity and fluconazole resistance through reduced affinity. J. Biol. Chem. 1997, 272, 5682–5688. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, J.F.; Gade, L.; Chow, N.A.; Loparev, V.N.; Juieng, P.; Berkow, E.L.; Farrer, R.A.; Litvintseva, A.P.; Cuomo, C.A. Genomic insights into multidrug-resistance, mating and virulence in Candida auris and related emerging species. Nat. Commun. 2018, 9, 5346. [Google Scholar] [CrossRef] [Green Version]
- Healey, K.R.; Kordalewska, M.; Jiménez Ortigosa, C.; Singh, A.; Berrío, I.; Chowdhary, A.; Perlin, D.S. Limited ERG11 mutations identified in isolates of Candida auris directly contribute to reduced azole susceptibility. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sionov, E.; Chang, Y.C.; Garraffo, H.M.; Dolan, M.A.; Ghannoum, M.A.; Kwon-Chung, K.J. Identification of a Cryptococcus neoformans cytochrome p450 lanosterol 14α-demethylase (ERG11) residue critical for differential susceptibility between fluconazole/voriconazole and itraconazole/posaconazole. Antimicrob. Agents Chemother. 2012, 56, 1162–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, M.J.; Liu, J.Y.; Ni, P.H.; Wang, S.; Shi, C.; Wei, B.; Ni, Y.X.; Ge, H.L. Erg11 mutations associated with azole resistance in clinical isolates of Candida albicans. Fems Yeast Res. 2013, 13, 386–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martini, C.; Torelli, R.; de Groot, T.; De Carolis, E.; Morandotti, G.A.; De Angelis, G.; Posteraro, B.; Meis, J.F.; Sanguinetti, M. Prevalence and clonal distribution of azole-resistant Candida parapsilosis isolates causing bloodstream infections in a large italian hospital. Front. Cell. Infect. Microbiol. 2020, 10, 232. [Google Scholar] [CrossRef] [PubMed]
- Bosco-Borgeat, M.E.; Mazza, M.; Taverna, C.G.; Córdoba, S.; Murisengo, O.A.; Vivot, W.; Davel, G. Amino acid substitution in Cryptococcus neoformans lanosterol 14-α-demethylase involved in fluconazole resistance in clinical isolates. Rev. Argent. Microbiol. 2016, 48, 137–142. [Google Scholar] [CrossRef] [Green Version]
- Couzigou, C.; Gabriel, F.; Biteau, N.; Fitton-Ouhabi, V.; Noël, T.; Accoceberry, I. Two missense mutations, E123Q and K151E, identified in the ERG11 allele of an azole-resistant isolate of Candida kefyr recovered from a stem cell transplant patient for acute myeloid leukemia. Med. Mycol. Case Rep. 2014, 5, 12–15. [Google Scholar] [CrossRef]
- Caramalho, R.; Tyndall, J.D.A.; Monk, B.C.; Larentis, T.; Lass-Flörl, C.; Lackner, M. Intrinsic short-tailed azole resistance in mucormycetes is due to an evolutionary conserved aminoacid substitution of the lanosterol 14α-demethylase. Sci. Rep. 2017, 7, 15898. [Google Scholar] [CrossRef]
- Bernhardt, A.; Meyer, W.; Rickerts, V.; Aebischer, T.; Tintelnot, K. Identification of 14-α-lanosterol demethylase (cyp51) in Scedosporium species. Antimicrob. Agents Chemother. 2018, 62, e02599-17. [Google Scholar] [CrossRef] [Green Version]
- Zulak, K.G.; Cox, B.A.; Tucker, M.A.; Oliver, R.P.; Lopez-Ruiz, F.J. Improved detection and monitoring of fungicide resistance in Blumeria graminis f. sp. hordei with high-throughput genotype quantification by digital PCR. Front. Microbiol. 2018, 9, 706. [Google Scholar] [CrossRef]
- Mullins, J.G.; Parker, J.E.; Cools, H.J.; Togawa, R.C.; Lucas, J.A.; Fraaije, B.A.; Kelly, D.E.; Kelly, S.L. Molecular modelling of the emergence of azole resistance in Mycosphaerella graminicola. PLoS ONE 2011, 6, e20973. [Google Scholar] [CrossRef]
- Heick, T.M.; Justesen, A.F.; Jørgensen, L.N. Resistance of wheat pathogen zymoseptoria tritici to DMI and QOI fungicides in the nordic-baltic region—A status. Eur. J. Plant Pathol. 2017, 149, 669–682. [Google Scholar] [CrossRef]
- Pereira, D.A.; McDonald, B.A.; Brunner, P.C. Mutations in the cyp51 gene reduce DMI sensitivity in Parastagonospora nodorum populations in europe and china. Pest Manag. Sci. 2017, 73, 1503–1510. [Google Scholar] [CrossRef] [PubMed]
- Pandey, B.; Sharma, P. Structural insights into impact of y134f mutation and discovery of novel fungicidal compounds against cyp51 in puccinia triticina. J. Cell. Biochem. 2018, 119, 2588–2603. [Google Scholar] [CrossRef]
- Frenkel, O.; Cadle-Davidson, L.; Wilcox, W.F.; Milgroom, M.G. Mechanisms of resistance to an azole fungicide in the grapevine powdery mildew fungus, Erysiphe necator. Phytopathology 2015, 105, 370–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monk, B.C.; Keniya, M.V.; Sabherwal, M.; Wilson, R.K.; Graham, D.O.; Hassan, H.F.; Chen, D.; Tyndall, J.D.A. Azole resistance reduces susceptibility to the tetrazole antifungal VT-1161. Antimicrob. Agents Chemother. 2019, 63, e02114-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagatova, A.A.; Keniya, M.V.; Tyndall, J.D.A.; Monk, B.C. Impact of homologous resistance mutations from pathogenic yeast on Saccharomyces cerevisiae lanosterol 14α-demethylase. Antimicrob. Agents Chemother. 2018, 62, e02242-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagatova, A.A.; Keniya, M.V.; Wilson, R.K.; Sabherwal, M.; Tyndall, J.D.; Monk, B.C. Triazole resistance mediated by mutations of a conserved active site tyrosine in fungal lanosterol 14α-demethylase. Sci. Rep. 2016, 6, 26213. [Google Scholar] [CrossRef] [Green Version]
- Lockhart, S.R. Candida auris and multidrug resistance: Defining the new normal. Fungal Genet. Biol. Fg B 2019, 131, 103243. [Google Scholar] [CrossRef]
- Mann, P.A.; Parmegiani, R.M.; Wei, S.Q.; Mendrick, C.A.; Li, X.; Loebenberg, D.; DiDomenico, B.; Hare, R.S.; Walker, S.S.; McNicholas, P.M. Mutations in Aspergillus fumigatus resulting in reduced susceptibility to posaconazole appear to be restricted to a single amino acid in the cytochrome p450 14alpha-demethylase. Antimicrob. Agents Chemother. 2003, 47, 577–581. [Google Scholar] [CrossRef] [Green Version]
- Tashiro, M.; Izumikawa, K.; Hirano, K.; Ide, S.; Mihara, T.; Hosogaya, N.; Takazono, T.; Morinaga, Y.; Nakamura, S.; Kurihara, S.; et al. Correlation between triazole treatment history and susceptibility in clinically isolated Aspergillus fumigatus. Antimicrob. Agents Chemother. 2012, 56, 4870–4875. [Google Scholar] [CrossRef] [Green Version]
- Alcazar-Fuoli, L.; Mellado, E.; Cuenca-Estrella, M.; Sanglard, D. Probing the role of point mutations in the cyp51a gene from Aspergillus fumigatus in the model yeast Saccharomyces cerevisiae. Med. Mycol. 2011, 49, 276–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prigitano, A.; Esposto, M.C.; Romanò, L.; Auxilia, F.; Tortorano, A.M. Azole-resistant Aspergillus fumigatus in the italian environment. J. Glob. Antimicrob. Resist. 2019, 16, 220–224. [Google Scholar] [CrossRef] [PubMed]
- Meis, J.F.; Chowdhary, A.; Rhodes, J.L.; Fisher, M.C.; Verweij, P.E. Clinical implications of globally emerging azole resistance in Aspergillus fumigatus. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2016, 371, 20150460. [Google Scholar] [CrossRef] [Green Version]
- Assress, H.A.; Nyoni, H.; Mamba, B.B.; Msagati, T.A.M. Occurrence and risk assessment of azole antifungal drugs in water and wastewater. Ecotoxicol. Environ. Saf. 2020, 187, 109868. [Google Scholar] [CrossRef] [PubMed]
- Tyndall, J.D.; Sabherwal, M.; Sagatova, A.A.; Keniya, M.V.; Negroni, J.; Wilson, R.K.; Woods, M.A.; Tietjen, K.; Monk, B.C. Structural and functional elucidation of yeast lanosterol 14α-demethylase in complex with agrochemical antifungals. PLoS ONE 2016, 11, e0167485. [Google Scholar] [CrossRef]
- Ullmann, A.J.; Aguado, J.M.; Arikan-Akdagli, S.; Denning, D.W.; Groll, A.H.; Lagrou, K.; Lass-Flörl, C.; Lewis, R.E.; Munoz, P.; Verweij, P.E.; et al. Diagnosis and management of aspergillus diseases: Executive summary of the 2017 escmid-ecmm-ers guideline. Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2018, 24 (Suppl. 1), e1–e38. [Google Scholar] [CrossRef] [Green Version]
- Wiederhold, N.P.; Patterson, T.F. Emergence of azole resistance in Aspergillus. Semin. Respir. Crit. Care Med. 2015, 36, 673–680. [Google Scholar] [CrossRef]
- Wiederhold, N.P. Antifungal resistance: Current trends and future strategies to combat. Infect. Drug Resist. 2017, 10, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Geneme Grima, M.A.; Zemene, M.; Mamuye, Y.; Getaneh, G. A review on aspergillosis in poultry. J. Vet. Sci. Technol. 2016, 7, 382. [Google Scholar]
- Ainsworth, G.C.; Rewell, R.E. The incidence of aspergillosis in captive wild birds. J. Comp. Pathol. Ther. 1949, 59, 213–IN217. [Google Scholar] [CrossRef]
- Beernaert, L.A.; Pasmans, F.; Van Waeyenberghe, L.; Haesebrouck, F.; Martel, A. Aspergillus infections in birds: A review. Avian Pathol. 2010, 39, 325–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Department of Conservation. Available online: https://www.doc.govt.nz/our-work/kakapo-recovery/what-we-do/kakapo-aspergillosis-outbreak/ (accessed on 30 November 2020).
- Kermani, F.; Shams-Ghahfarokhi, M.; Gholami-Shabani, M.; Razzaghi-Abyaneh, M. Diversity, molecular phylogeny and fingerprint profiles of airborne Aspergillus species using random amplified polymorphic DNA. World J. Microbiol. Biotechnol. 2016, 32, 96. [Google Scholar] [CrossRef] [PubMed]
- Godeau, C.; Reboux, G.; Scherer, E.; Laboissiere, A.; Lechenault-Bergerot, C.; Millon, L.; Rocchi, S. Azole-resistant Aspergillus fumigatus in the hospital: Surveillance from flower beds to corridors. Am. J. Infect. Control 2020, 48, 702–704. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, D. Isolation of azole-resistant Aspergillus fumigatus from imported plant bulbs in japan and the effect of fungicide treatment. J. Pestic. Sci. 2020, 45, 147–150. [Google Scholar] [CrossRef]
- Nakano, Y.; Tashiro, M.; Urano, R.; Kikuchi, M.; Ito, N.; Moriya, E.; Shirahige, T.; Mishima, M.; Takazono, T.; Miyazaki, T.; et al. Characteristics of azole-resistant Aspergillus fumigatus attached to agricultural products imported to japan. J. Infect. Chemother. Off. J. Jpn. Soc. Chemother. 2020, 26, 1021–1025. [Google Scholar] [CrossRef]
- Buil, J.B.; Zoll, J.; Verweij, P.E.; Melchers, W.J.G. Molecular detection of azole-resistant Aspergillus fumigatus in clinical samples. Front. Microbiol. 2018, 9, 515. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Snelders, E.; Zwaan, B.J.; Schoustra, S.E.; Meis, J.F.; van Dijk, K.; Hagen, F.; van der Beek, M.T.; Kampinga, G.A.; Zoll, J.; et al. A novel environmental azole resistance mutation in Aspergillus fumigatus and a possible role of sexual reproduction in its emergence. mBio 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Jeanvoine, A.; Rocchi, S.; Bellanger, A.P.; Reboux, G.; Millon, L. Azole-resistant Aspergillus fumigatus: A global phenomenon originating in the environment? Med. Mal. Infect. 2020, 50, 389–395. [Google Scholar] [CrossRef]
- Dos Santos, R.A.C.; Rivero-Menendez, O.; Steenwyk, J.L.; Mead, M.E.; Goldman, G.H.; Alastruey-Izquierdo, A.; Rokas, A. Draft genome sequences of four Aspergillus section fumigati clinical strains. Microbiol. Resour. Announc. 2020, 9, e00856-20. [Google Scholar] [CrossRef]
- Samson, R.A.; Hong, S.; Peterson, S.W.; Frisvad, J.C.; Varga, J. Polyphasic taxonomy of Aspergillus section fumigati and its teleomorph Neosartorya. Stud. Mycol. 2007, 59, 147–203. [Google Scholar] [CrossRef]
- Mellado, E.; Alcazar-Fuoli, L.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L. Role of Aspergillus lentulus 14-α sterol demethylase (cyp51a) in azole drug susceptibility. Antimicrob. Agents Chemother. 2011, 55, 5459–5468. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcazar-Fuoli, L.; Mellado, E.; Alastruey-Izquierdo, A.; Cuenca-Estrella, M.; Rodriguez-Tudela, J.L. Aspergillus section fumigati: Antifungal susceptibility patterns and sequence-based identification. Antimicrob. Agents Chemother. 2008, 52, 1244–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chong, G.M.; Vonk, A.G.; Meis, J.F.; Dingemans, G.J.; Houbraken, J.; Hagen, F.; Gaajetaan, G.R.; van Tegelen, D.W.; Simons, G.F.; Rijnders, B.J. Interspecies discrimination of A fumigatus and siblings A. lentulus and A. felis of the Aspergillus section fumigati using the aspergenius (®) assay. Diagn. Microbiol. Infect. Dis. 2017, 87, 247–252. [Google Scholar] [CrossRef] [PubMed]
- Bassetti, M.; Vena, A.; Bouza, E.; Peghin, M.; Muñoz, P.; Righi, E.; Pea, F.; Lackner, M.; Lass-Flörl, C. Antifungal susceptibility testing in candida, aspergillus and cryptococcus infections: Are the mics useful for clinicians? Clin. Microbiol. Infect. Off. Publ. Eur. Soc. Clin. Microbiol. Infect. Dis. 2020, 26, 1024–1033. [Google Scholar] [CrossRef]
- Schelenz, S.; Hagen, F.; Rhodes, J.L.; Abdolrasouli, A.; Chowdhary, A.; Hall, A.; Ryan, L.; Shackleton, J.; Trimlett, R.; Meis, J.F.; et al. First hospital outbreak of the globally emerging Candida auris in a european hospital. Antimicrob. Resist. Infect. Control 2016, 5, 35. [Google Scholar] [CrossRef] [Green Version]
- CDC. CDC. Available online: https://www.cdc.gov/fungal/candida-auris/index.html (accessed on 30 November 2020).
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical practice guideline for the management of candidiasis: 2016 update by the infectious diseases society of America. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2016, 62, e1–e50. [Google Scholar] [CrossRef]
- Arastehfar, A.; Daneshnia, F.; Hilmioğlu-Polat, S.; Fang, W.; Yaşar, M.; Polat, F.; Metin, D.Y.; Rigole, P.; Coenye, T.; Ilkit, M.; et al. First report of candidemia clonal outbreak caused by emerging fluconazole-resistant Candida parapsilosis isolates harboring Y132F and/or Y132F+K143R in turkey. Antimicrob. Agents Chemother. 2020, 64, e01001-20. [Google Scholar] [CrossRef]
- Kuhn, D.M.; Mikherjee, P.K.; Clark, T.A.; Pujol, C.; Chandra, J.; Hajjeh, R.A.; Warnock, D.W.; Soil, D.R.; Ghannoum, M.A. Candida parapsilosis characterization in an outbreak setting. Emerg. Infect. Dis. 2004, 10, 1074–1081. [Google Scholar] [CrossRef]
- Chow, N.A.; Gade, L.; Tsay, S.V.; Forsberg, K.; Greenko, J.A.; Southwick, K.L.; Barrett, P.M.; Kerins, J.L.; Lockhart, S.R.; Chiller, T.M.; et al. Multiple introductions and subsequent transmission of multidrug-resistant Candida auris in the USA: A molecular epidemiological survey. Lancet Infect. Dis. 2018, 18, 1377–1384. [Google Scholar] [CrossRef]
- Chaabane, F.; Graf, A.; Jequier, L.; Coste, A.T. Review on antifungal resistance mechanisms in the emerging pathogen Candida auris. Front. Microbiol. 2019, 10, 2788. [Google Scholar] [CrossRef]
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.L.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A.; et al. Simultaneous emergence of multidrug-resistant Candida auris on 3 continents confirmed by whole-genome sequencing and epidemiological analyses. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2017, 64, 134–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, S.; Khan, Z.; Al-Sweih, N.; Alfouzan, W.; Joseph, L. Candida auris in various hospitals across kuwait and their susceptibility and molecular basis of resistance to antifungal drugs. Mycoses 2020, 63, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Chow, N.A.; Muñoz, J.F.; Gade, L.; Berkow, E.L.; Li, X.; Welsh, R.M.; Forsberg, K.; Lockhart, S.R.; Adam, R.; Alanio, A.; et al. Tracing the evolutionary history and global expansion of Candida auris using population genomic analyses. mBio 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Köhler, J.R.; Hube, B.; Puccia, R.; Casadevall, A.; Perfect, J.R. Fungi that infect humans. Microbiol. Spectr. 2017, 5. [Google Scholar] [CrossRef]
- Saag, M.S.; Graybill, R.J.; Larsen, R.A.; Pappas, P.G.; Perfect, J.R.; Powderly, W.G.; Sobel, J.D.; Dismukes, W.E. Practice guidelines for the management of cryptococcal disease. Infectious diseases society of America. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2000, 30, 710–718. [Google Scholar] [CrossRef]
- Firacative, C.; Torres, G.; Meyer, W.; Escandón, P. Clonal dispersal of Cryptococcus gattii VGII in an endemic region of cryptococcosis in Colombia. J. Fungi (Basel, Switz.) 2019, 5, 32. [Google Scholar] [CrossRef] [Green Version]
- Carriconde, F.; Gilgado, F.; Arthur, I.; Ellis, D.; Malik, R.; van de Wiele, N.; Robert, V.; Currie, B.J.; Meyer, W. Clonality and α-a recombination in the australian Cryptococcus gattii VGII population--an emerging outbreak in Australia. PLoS ONE 2011, 6, e16936. [Google Scholar] [CrossRef] [Green Version]
- Duncan, C.; Schwantje, H.; Stephen, C.; Campbell, J.; Bartlett, K. Cryptococcus gattii in wildlife of Vancouver island, British Columbia, Canada. J. Wildl. Dis. 2006, 42, 175–178. [Google Scholar] [CrossRef] [Green Version]
- Sionov, E.; Chang, Y.C.; Kwon-Chung, K.J. Azole heteroresistance in Cryptococcus neoformans: Emergence of resistant clones with chromosomal disomy in the mouse brain during fluconazole treatment. Antimicrob. Agents Chemother. 2013, 57, 5127–5130. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.C.; Chang, T.Y.; Liu, J.W.; Chen, F.J.; Chien, C.C.; Lee, C.H.; Lu, C.H. Increasing trend of fluconazole-non-susceptible Cryptococcus neoformans in patients with invasive cryptococcosis: A 12-year longitudinal study. Bmc Infect. Dis. 2015, 15, 277. [Google Scholar] [CrossRef] [Green Version]
- Mpoza, E.; Rhein, J.; Abassi, M. Emerging fluconazole resistance: Implications for the management of cryptococcal meningitis. Med. Mycol. Case Rep. 2018, 19, 30–32. [Google Scholar] [CrossRef]
- Ventoulis, I.; Sarmourli, T.; Amoiridou, P.; Mantzana, P.; Exindari, M.; Gioula, G.; Vyzantiadis, T.A. Bloodstream infection by Saccharomyces cerevisiae in two covid-19 patients after receiving supplementation of Saccharomyces in the ICU. J. Fungi (Basel, Switzerland) 2020, 6, 98. [Google Scholar] [CrossRef]
- Parapouli, M.; Vasileiadis, A.; Afendra, A.S.; Hatziloukas, E. Saccharomyces cerevisiae and its industrial applications. Aims Microbiol. 2020, 6, 1–31. [Google Scholar] [CrossRef]
- Lin, C.Y.; Alexander, C.; Steelman, A.J.; Warzecha, C.M.; de Godoy, M.R.C.; Swanson, K.S. Effects of a Saccharomyces cerevisiae fermentation product on fecal characteristics, nutrient digestibility, fecal fermentative end-products, fecal microbial populations, immune function, and diet palatability in adult dogs1. J. Anim. Sci. 2019, 97, 1586–1599. [Google Scholar] [CrossRef] [PubMed]
- van der Hoek, S.A.; Darbani, B.; Zugaj, K.E.; Prabhala, B.K.; Biron, M.B.; Randelovic, M.; Medina, J.B.; Kell, D.B.; Borodina, I. Engineering the yeast Saccharomyces cerevisiae for the production of l-(+)-ergothioneine. Front. Bioeng. Biotechnol. 2019, 7, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baeshen, N.A.; Baeshen, M.N.; Sheikh, A.; Bora, R.S.; Ahmed, M.M.; Ramadan, H.A.; Saini, K.S.; Redwan, E.M. Cell factories for insulin production. Microb. Cell Factories 2014, 13, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karathia, H.; Vilaprinyo, E.; Sorribas, A.; Alves, R. Saccharomyces cerevisiae as a model organism: A comparative study. PLoS ONE 2011, 6, e16015. [Google Scholar] [CrossRef] [Green Version]
- Cornely, O.A.; Alastruey-Izquierdo, A.; Arenz, D.; Chen, S.C.A.; Dannaoui, E.; Hochhegger, B.; Hoenigl, M.; Jensen, H.E.; Lagrou, K.; Lewis, R.E.; et al. Global guideline for the diagnosis and management of mucormycosis: An initiative of the european confederation of medical mycology in cooperation with the mycoses study group education and research consortium. Lancet Infect. Dis. 2019, 19, e405–e421. [Google Scholar] [CrossRef]
- Lackner, M.; de Hoog, G.S.; Verweij, P.E.; Najafzadeh, M.J.; Curfs-Breuker, I.; Klaassen, C.H.; Meis, J.F. Species-specific antifungal susceptibility patterns of Scedosporium and Pseudallescheria species. Antimicrob. Agents Chemother. 2012, 56, 2635–2642. [Google Scholar] [CrossRef] [Green Version]
- Spec, A.; Connolly, P.; Montejano, R.; Wheat, L.J. In vitro activity of isavuconazole against fluconazole-resistant isolates of Histoplasma capsulatum. Med. Mycol. 2018, 56, 834–837. [Google Scholar] [CrossRef] [Green Version]
- Wheat, L.J.; Connolly, P.; Smedema, M.; Rogers, P.D. Antifungal drug resistance in histoplasmosis. In Antimicrobial Drug Resistance: Clinical and Epidemiological Aspects; Mayers, D.L., Ed.; Humana Press: Totowa, NJ, USA, 2009; pp. 987–992. [Google Scholar]
- Walther, G.; Wagner, L.; Kurzai, O. Updates on the taxonomy of mucorales with an emphasis on clinically important taxa. J. Fungi (Basel, Switz.) 2019, 5, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, J.P.F.; Feliciano, L.M.; Santos, D.C.S.; Ramos, S.; Oliveira, R.A.; Attili-Angelis, D.; Rodrigues, N.R.; Sampaio, J.L.M.; dos Anjos Martins, M.; Melhem, M.S.C. Could fungicides lead to azole drug resistance in a cross-resistance manner among environmental Cryptococcus strains? Curr. Fungal Infect. Rep. 2020, 14, 9–14. [Google Scholar] [CrossRef]
- Kleinkauf, N.; Verweij, P.E.; Arendrup, M.C.; Donnelly, P.J.; Cuenca-Estrella, M.; Fraaije, B.; Melchers, W.J.G.; Adriaenssens, N.; Kema, G.H.J.; Ullmann, A.; et al. Risk Assessment on the Impact of Environmental Usage of Triazoles on the Development and Spread of Resistance to Medical Triazoles in Aspergillus Species; ECDC: Stockholm, Sweden, 2013. [Google Scholar]
- Tucker, M.A.; Lopez-Ruiz, F.; Cools, H.J.; Mullins, J.G.; Jayasena, K.; Oliver, R.P. Analysis of mutations in west australian populations of Blumeria graminis f. sp. hordei cyp51 conferring resistance to DMI fungicides. Pest Manag. Sci. 2020, 76, 1265–1272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Draz, I.S.; Abou-Elseoud, M.S.; Kamara, A.-E.M.; Alaa-Eldein, O.A.-E.; El-Bebany, A.F. Screening of wheat genotypes for leaf rust resistance along with grain yield. Ann. Agric. Sci. 2015, 60, 29–39. [Google Scholar] [CrossRef] [Green Version]
- Stammler, G.; Cordero, J.; Koch, A.; Semar, M.; Schlehuber, S. Role of the Y134F mutation in cyp51 and overexpression of cyp51 in the sensitivity response of Puccinia triticina to epoxiconazole. Crop Prot. 2009, 28, 891–897. [Google Scholar] [CrossRef]
- Couch, R.B.; Webster, R.G.; Kasel, J.A.; Cate, T.R. Efficacy of purified influenza subunit vaccines and relation to the major antigenic determinants on the hemagglutinin molecule. J. Infect. Dis. 1979, 140, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Savocchia, S.; Stummer, B.E.; Wicks, T.J.; van Heeswijck, R.; Scott, E.S. Reduced sensitivity of Uncinula necator to sterol demethylation inhibiting fungicides in southern australian vineyards. Australas. Plant Pathol. 2004, 33, 465–473. [Google Scholar] [CrossRef]
- Xisto, M.I.; Caramalho, R.D.; Rocha, D.A.; Ferreira-Pereira, A.; Sartori, B.; Barreto-Bergter, E.; Junqueira, M.L.; Lass-Flörl, C.; Lackner, M. Pan-azole-resistant Candida tropicalis carrying homozygous ERG11 mutations at position k143r: A new emerging superbug? J. Antimicrob. Chemother. 2017, 72, 988–992. [Google Scholar]
- Sabino, R.; Veríssimo, C.; Pereira, Á.A.; Antunes, F. Candida auris, an agent of hospital-associated outbreaks: Which challenging issues do we need to have in mind? Microorganisms 2020, 8, 181. [Google Scholar] [CrossRef] [Green Version]
- Kordalewska, M.; Lee, A.; Zhao, Y.; Perlin, D.S. Detection of Candida auris antifungal drug resistance markers directly from clinical skin swabs. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef]
- Eyre, D.W.; Sheppard, A.E.; Madder, H.; Moir, I.; Moroney, R.; Quan, T.P.; Griffiths, D.; George, S.; Butcher, L.; Morgan, M.; et al. A Candida auris outbreak and its control in an intensive care setting. N. Engl. J. Med. 2018, 379, 1322–1331. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Gaitán, A.; Martínez, H.; Moret, A.M.; Calabuig, E.; Tasias, M.; Alastruey-Izquierdo, A.; Zaragoza, Ó.; Mollar, J.; Frasquet, J.; Salavert-Lletí, M.; et al. Detection and treatment of Candida auris in an outbreak situation: Risk factors for developing colonization and candidemia by this new species in critically ill patients. Expert Rev. Anti-Infect. Ther. 2019, 17, 295–305. [Google Scholar]
- Forastiero, A.; Mesa-Arango, A.C.; Alastruey-Izquierdo, A.; Alcazar-Fuoli, L.; Bernal-Martinez, L.; Pelaez, T.; Lopez, J.F.; Grimalt, J.O.; Gomez-Lopez, A.; Cuesta, I.; et al. Candida tropicalis antifungal cross-resistance is related to different azole target (erg11p) modifications. Antimicrob. Agents Chemother. 2013, 57, 4769–4781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flowers, S.A.; Colón, B.; Whaley, S.G.; Schuler, M.A.; Rogers, P.D. Contribution of clinically derived mutations in ERG11 to azole resistance in Candida albicans. Antimicrob. Agents Chemother. 2015, 59, 450–460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cools, H.J.; Fraaije, B.A. Update on mechanisms of azole resistance in Mycosphaerella graminicola and implications for future control. Pest Manag. Sci. 2013, 69, 150–155. [Google Scholar] [CrossRef]
- Perlin, D.S.; Rautemaa-Richardson, R.; Alastruey-Izquierdo, A. The global problem of antifungal resistance: Prevalence, mechanisms, and management. Lancet Infect. Dis. 2017, 17, e383–e392. [Google Scholar] [CrossRef]
- Chen, Y.; Lu, Z.; Zhao, J.; Zou, Z.; Gong, Y.; Qu, F.; Bao, Z.; Qiu, G.; Song, M.; Zhang, Q.; et al. Epidemiology and molecular characterizations of azole resistance in clinical and environmental Aspergillus fumigatus isolates from China. Antimicrob. Agents Chemother. 2016, 60, 5878–5884. [Google Scholar] [CrossRef] [Green Version]
- Chowdhary, A.; Sharma, C.; van den Boom, M.; Yntema, J.B.; Hagen, F.; Verweij, P.E.; Meis, J.F. Multi-azole-resistant Aspergillus fumigatus in the environment in Tanzania. J. Antimicrob. Chemother. 2014, 69, 2979–2983. [Google Scholar] [CrossRef]
- Bromley, M.J.; van Muijlwijk, G.; Fraczek, M.G.; Robson, G.; Verweij, P.E.; Denning, D.W.; Bowyer, P. Occurrence of azole-resistant species of Aspergillus in the UK environment. J. Glob. Antimicrob. Resist. 2014, 2, 276–279. [Google Scholar] [CrossRef]
- Cao, D.; Wu, R.; Dong, S.; Wang, F.; Ju, C.; Yu, S.; Xu, S.; Fang, H.; Yu, Y. Five-year survey (2014 to 2018) of azole resistance in environmental Aspergillus fumigatus isolates from china. Antimicrob. Agents Chemother. 2020, 64. [Google Scholar] [CrossRef]
- Caetano, L.A.; Faria, T.; Springer, J.; Loeffler, J.; Viegas, C. Antifungal-resistant mucorales in different indoor environments. Mycology 2019, 10, 75–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomlin, C. The Pesticide Manual: A World Compendium, 11th ed.; British Crop Protection Council: Farnham, Surrey, UK, 1997. [Google Scholar]
- Cairns, T.; Siegmund, E.G.; Chiu, K.S.; Nelson, R. Residue characterization of triadimefon in grapes by gas chromatography and mass spectrometry/mass spectrometry. Biomed. Environ. Mass Spectrom. 1989, 18, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Hay, R. Therapy of skin, hair and nail fungal infections. J. Fungi 2018, 4, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caetano, L.A.; Faria, T.; Batista, A.C.; Viegas, S.; Viegas, C. Assessment of occupational exposure to azole resistant fungi in 10 portuguese bakeries. AIMS Microbiol. 2017, 3, 960–975. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
| Genus | Species | Phylum | AA Change in Active Site | 2nd Mutation | Gene | Correlated Resistance * | Ref. | |
|---|---|---|---|---|---|---|---|---|
| animal-pathogenic fungi | Ajellomyces | capsulatus | Ascomycota | Y136F | ERG11 | FLC, ITC | [28] | |
| Aspergillus | fumigatus | Ascomycota | Y121F | (TR46) T289A | cyp51A | VCZ | [31,32,33,34,35] | |
| Aspergillus | fumigatus | Ascomycota | I301 | cyp51A | FLC | [36] | ||
| Aspergillus | lentulus | Ascomycota | n.a. | cyp51A | VCZ | [37] | ||
| Candida | albicans | Ascomycota | G464S | ERG11 | FLC | [38] | ||
| Candida | albicans | Ascomycota | Y132F, K143R | ERG11 | FLC, VCZ | [39,40] | ||
| Candida | albicans | Ascomycota | T315A | ERG11 | FLC | [41] | ||
| Candida | auris | Ascomycota | Y132F, K143R | ERG11 | FLC | [41] | ||
| Candida | orthopsilosis | Ascomycota | Y132F, K143R | ERG11 | FLC, VCZ | [42] | ||
| Candida | parapsilosis | Ascomycota | Y132F | P6S, C45G, G50L, D460T | ERG11 | FLC, VCZ | [43,44] | |
| Candida | parapsilosis | Ascomycota | Y132F | R398I | ERG11 | FLC, VCZ | [45,46] | |
| Candida | tropicalis | Ascomycota | Y132F, K143R | ERG11 | FLC, VCZ | [40,43] | ||
| Cryptococcus | neoformans | Basidiomycota | Y145F | ERG11 | VCZ | [47] | ||
| Kluyveromyces | marxianus | Ascomycota | K151 | ERG11 | FLC, VCZ, ITC, PCZ | [48] | ||
| Mucor | circinelloides | Mucormycota | F129 | A291 | CYP51 F5 | FLC, VCZ | [49] | |
| Rhizopus | arrhizus | Mucormycota | F129 | A291 | CYP51 F5 | FLC, VCZ | [49] | |
| Rhizopus | microsporus | Mucormycota | F129 | A291 | CYP51 F5 | FLC, VCZ | [49] | |
| Scedosporium | apiospermum | Ascomycota | Y136F | cyp51 | VCZ | [50] | ||
| plant-pathogenic fungi | Blumeria | graminis | Ascomycota | Y136F/Y137F | Cyp51 | TBC | [51] | |
| Mycosphaerella | graminicola | Ascomycota | Y137F, G460 | Cyp51 | TDM, TBC, EPC | [52,53] | ||
| Parastagonospora | nodorum | Ascomycota | Y144F/H | Cyp51 | PPC | [54] | ||
| Puccinia | triticina | Basidiomycota | Y134F | Cyp51 | EPC | [55] | ||
| Uncinula | necator | Ascomycota | Y136F | Cyp51 | TBC | [56] |
| Saccharomyces | Cerevisiae | Y140 | T322 | G464 | K151 | G73 | V311 |
|---|---|---|---|---|---|---|---|
| Ajellomyces | capsulatus | Y136F | - | - | - | - | - |
| Aspergillus | fumigatus | Y121F | I301 | - | - | G54 | A289 |
| Aspergillus | lentulus | - | - | G448S | - | - | - |
| Candida | albicans | Y132F | T315A | G464S | K143R | - | - |
| Candida | auris | Y132F | - | - | K143R | - | - |
| Candida | orthopsilosis | Y132F | - | - | K143R | - | - |
| Candida | parapsilosis | Y132F | - | - | - | - | - |
| Candida | tropicalis | Y132F | - | - | K143R | - | - |
| Cryptococcus | neoformans | Y145F | - | - | - | - | - |
| Kluyveromyces | marxianus | - | - | - | K151 | - | - |
| Mucor | circinelloides | F129 | - | - | - | - | A291 |
| Rhizopus | arrhizus | F129 | - | - | - | - | A291 |
| Rhizopus | microsporus | F129 | - | - | - | - | A291 |
| Scedosporium | apiospermum | Y136F | - | G464S | - | - | - |
| Blumeria | graminis | Y136F | - | - | - | - | - |
| Mycosphaerella | graminicola | Y137F | - | G460 | - | - | - |
| Parastagonospora | nodorum | Y144F/H | - | - | - | - | - |
| Puccinia | triticina | Y134F | - | - | - | - | - |
| Uncinula | necator | Y136F | - | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosam, K.; Monk, B.C.; Lackner, M. Sterol 14α-Demethylase Ligand-Binding Pocket-Mediated Acquired and Intrinsic Azole Resistance in Fungal Pathogens. J. Fungi 2021, 7, 1. https://doi.org/10.3390/jof7010001
Rosam K, Monk BC, Lackner M. Sterol 14α-Demethylase Ligand-Binding Pocket-Mediated Acquired and Intrinsic Azole Resistance in Fungal Pathogens. Journal of Fungi. 2021; 7(1):1. https://doi.org/10.3390/jof7010001
Chicago/Turabian StyleRosam, Katharina, Brian C. Monk, and Michaela Lackner. 2021. "Sterol 14α-Demethylase Ligand-Binding Pocket-Mediated Acquired and Intrinsic Azole Resistance in Fungal Pathogens" Journal of Fungi 7, no. 1: 1. https://doi.org/10.3390/jof7010001
APA StyleRosam, K., Monk, B. C., & Lackner, M. (2021). Sterol 14α-Demethylase Ligand-Binding Pocket-Mediated Acquired and Intrinsic Azole Resistance in Fungal Pathogens. Journal of Fungi, 7(1), 1. https://doi.org/10.3390/jof7010001