
Sand Particle Size and Phosphorus Amount Affect Rhizophagus irregularis Spore Production Using In Vitro Propagated Spore as a Starter Inoculum in Rhizosphere of Maize (Zea mays) Plantlets

 , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Isolation of AM Fungal Spore
2.2. Establishment of Monoxenic Culture of AM Fungi
2.3. Molecular Identification of Monoxenic Mycorrhizae
2.4. Optimization of M-Media for R. irregularis Isolate Apex-MYK-01
2.5. Sand Substrate-Based Inoculum Production of R. irregularis Isolate Apex-MYK-01 under Maize Plant System
2.5.1. Sand Processing and Grading
2.5.2. Inoculation of Maize Seedlings in Sand with R. irregularis Isolate Apex-MYK-01
2.6. Harvest and Data Measurement
2.7. Statistical Analysis
3. Results
3.1. Monoxenic Culture and Identification of AM Fungal Strain
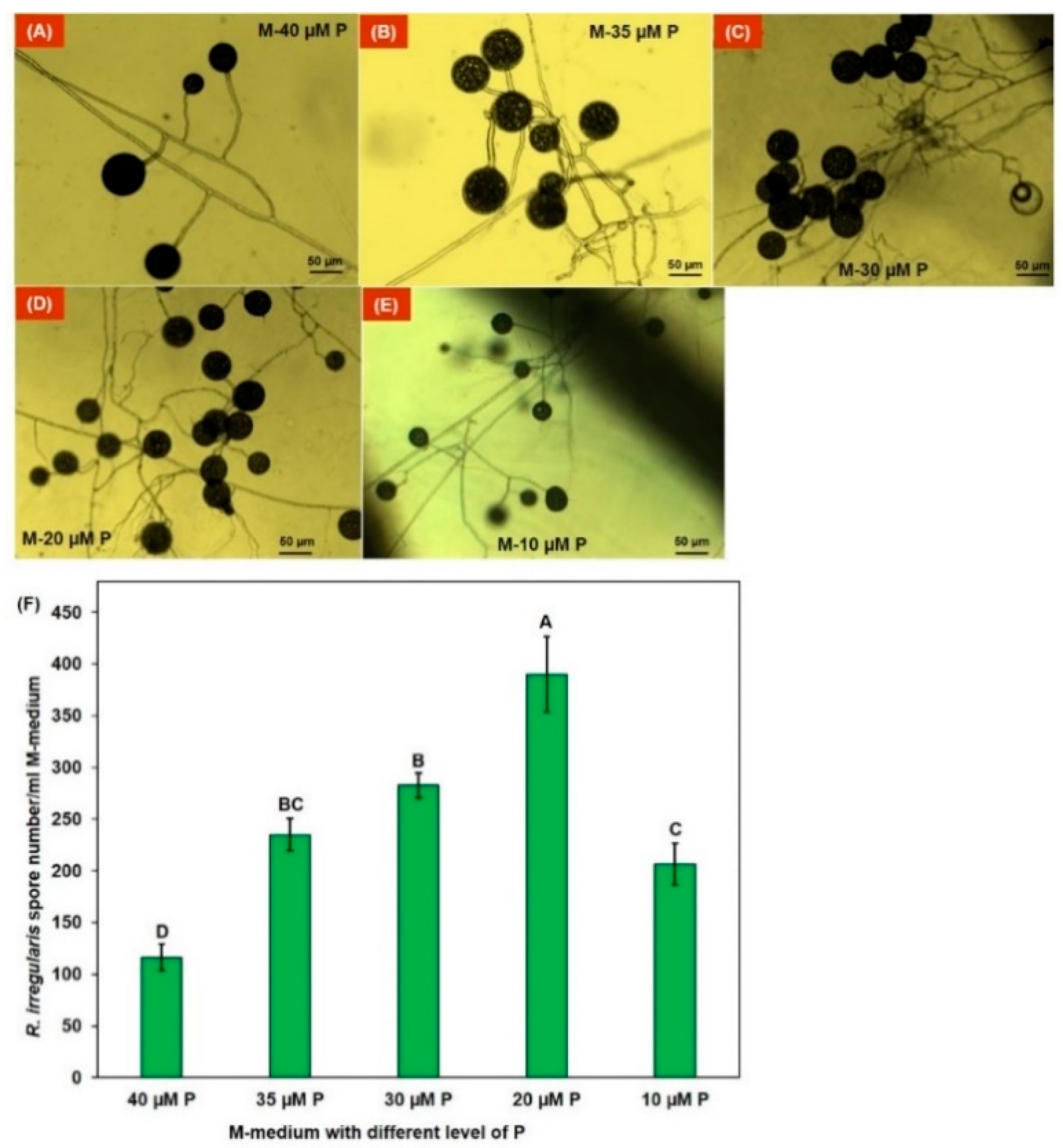
3.2. AM Fungal Spore Number in Different Level of P in Monoxenic Culture
3.3. R. irregularis Spore Density and Colonization Rate in Sand Substrate-Based Culture
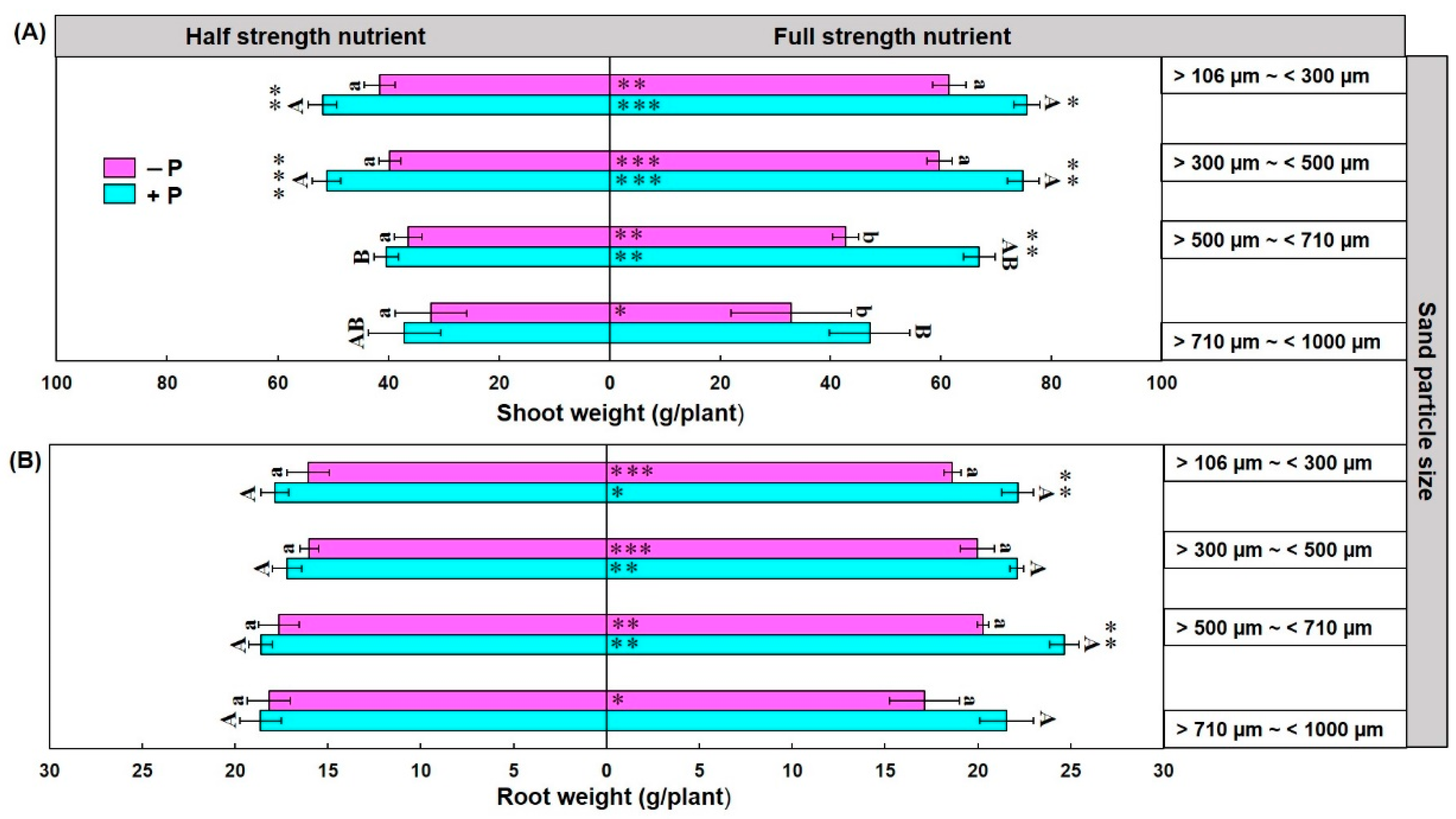
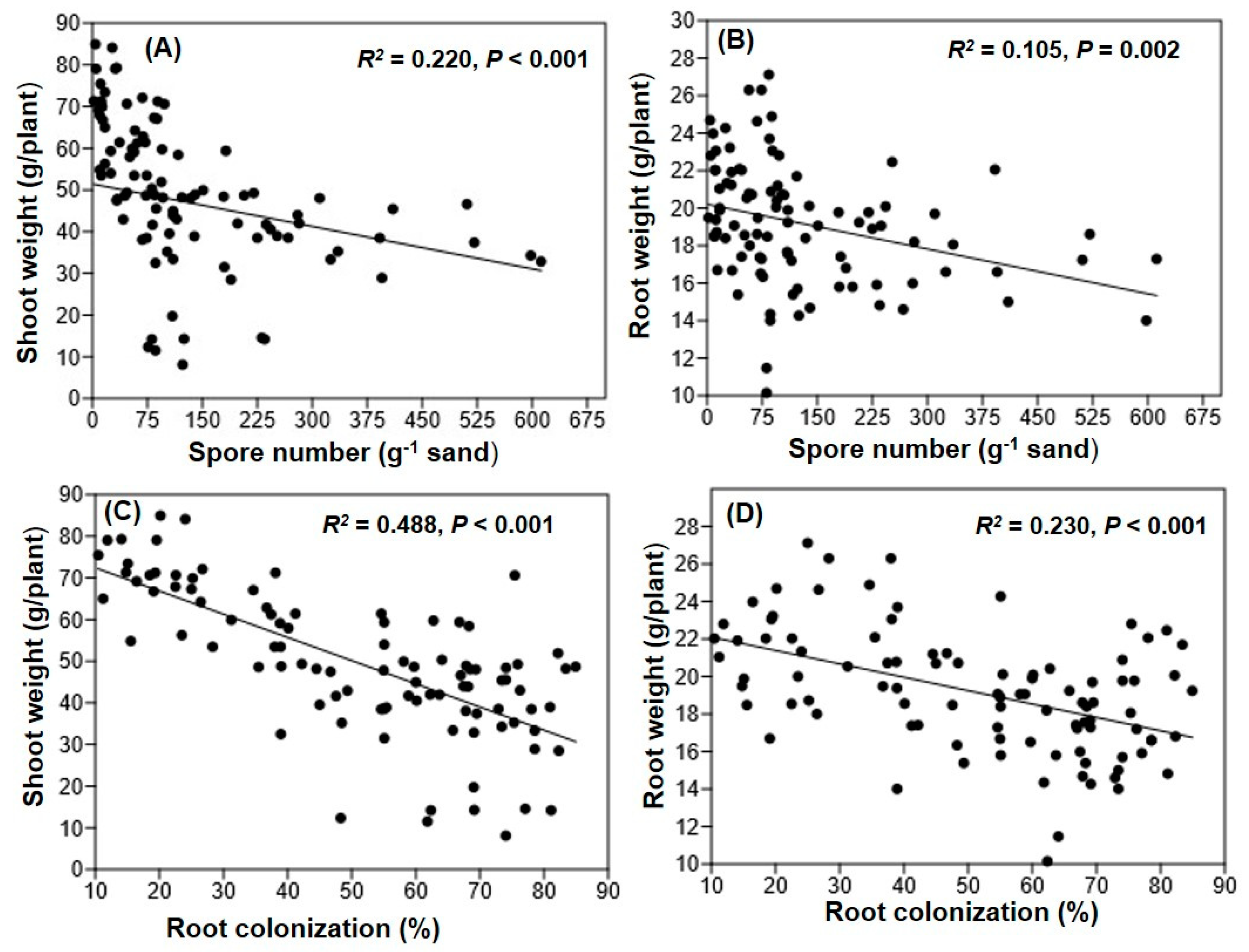
3.4. Maize Shoot and Root Weight and Their Relation with Spore Density and Colonization
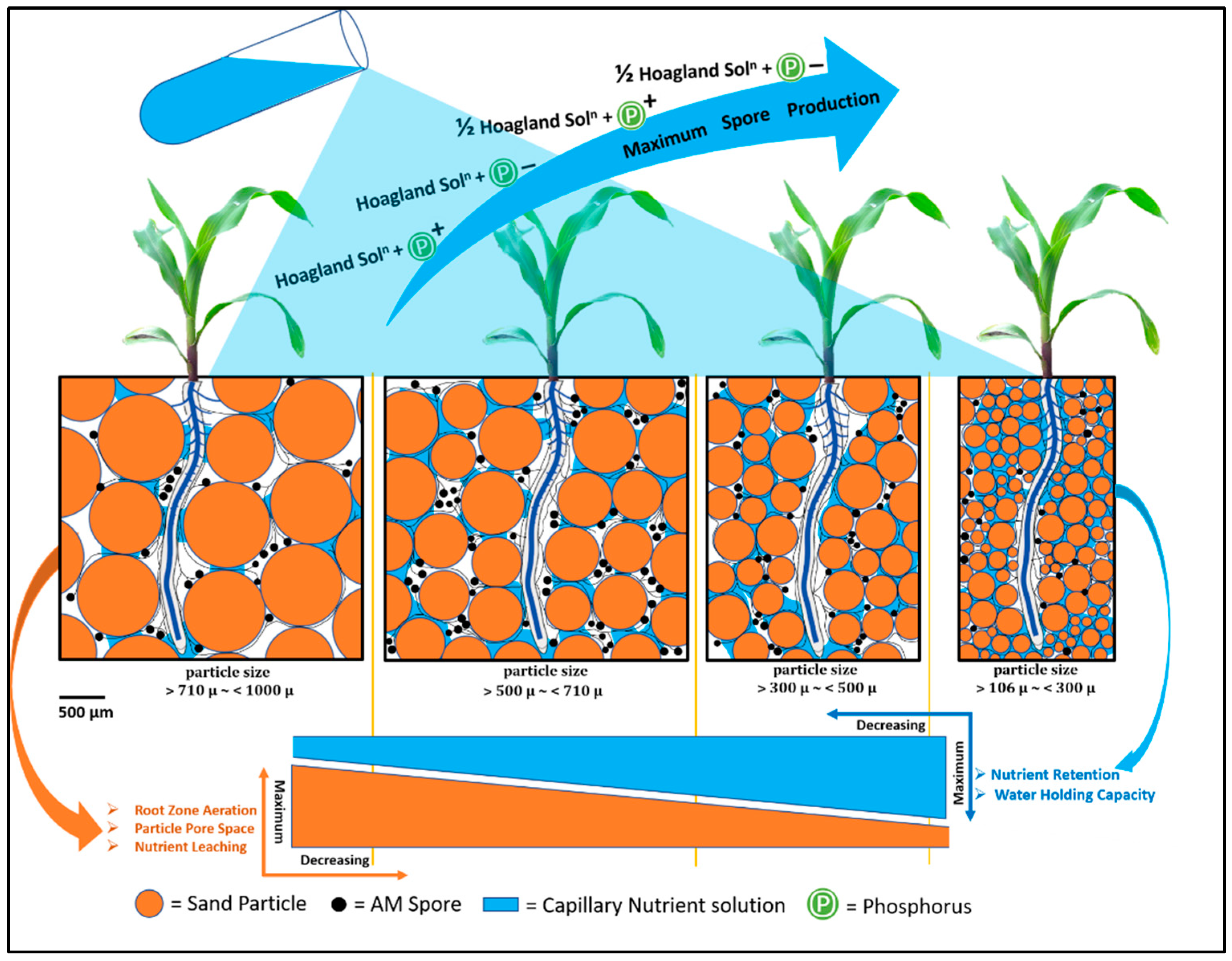
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis, 3rd ed.; Academic Press: London, UK, 2008. [Google Scholar]
- Smith, F.A.; Jakobsen, I.; Smith, S.E. Spatial differences in acquisition of soil phosphate between two arbuscular mycorrhizal fungi in symbiosis with Medicago truncatula. New Phytol. 2000, 147, 357–366. [Google Scholar] [CrossRef]
- Shi, S.; Luo, X.; Wen, M.; Dong, X.; Sharifi, S.; Xie, D.; He, X. Funneliformis mosseae improves growth and nutrient accumulation in wheat by facilitating soil nutrient uptake under elevated CO2 at daytime, not nighttime. J. Fungi 2021, 7, 458. [Google Scholar] [CrossRef] [PubMed]
- Tedersoo, L.; Bahram, M. Mycorrhizal types differ in ecophysiology and alter plant nutrition and soil processes. Biol. Rev. 2019, 94, 1857–1880. [Google Scholar] [CrossRef]
- Selvakumar, G.; Shagol, C.C.; Kang, Y.; Chung, B.N.; Han, S.G.; Sa, T. Arbuscular mycorrhizal fungi spore propagation using single spore as starter inoculum and a plant host. J. Appl. Microbiol. 2018, 124, 1556–1565. [Google Scholar] [CrossRef] [PubMed]
- Rosikiewicz, P.; Bonvin, J.; Sanders, I.R. Cost-efficient production of in vitro Rhizophagus irregularis. Mycorrhiza 2017, 27, 477–486. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Wei, S.; Hu, W.; Xiao, L.; Tang, M. Arbuscular mycorrhizal fungus Rhizophagus irregularis increased potassium content and expression of genes encoding potassium channels in Lycium barbarum. Front. Plant Sci. 2017, 8, 440. [Google Scholar] [CrossRef] [Green Version]
- Ijdo, M.; Cranenbrouck, S.; Declerck, S. Methods for large-scale production of AM fungi: Past, present, and future. Mycorrhiza 2010, 21, 1–16. [Google Scholar] [CrossRef]
- Ruiz-Lozano, J.; Porcel, R.; Bárzana, G.; Azcón, R.; Aroca, R. Contribution of Arbuscular Mycorrhizal Symbiosis to Plant Drought Tolerance: State of the Art. In Plant Responses to Drought Stress; Gabler: Wiesbaden, Germany, 2012; pp. 335–362. [Google Scholar]
- Jabborova, D.; Annapurna, K.; Paul, S.; Kumar, S.; Saad, H.A.; Desouky, S.; Ibrahim, M.F.M.; Elkelish, A. Beneficial features of biochar and arbuscular mycorrhiza for improving spinach plant growth, root morphological traits, physiological properties, and soil enzymatic activities. J. Fungi 2021, 7, 571. [Google Scholar] [CrossRef]
- Xia, L.; Shao, C.; Zhang, N.; Wu, A.; Xie, J.; Qiu, Y.; He, X.; Pei, J.; Wang, X.; Wang, Y. Improved tolerance of mycorrhizal Torreya grandis seedlings to sulfuric acid rain related to phosphorus and zinc contents in shoots. J. Fungi 2021, 7, 296. [Google Scholar] [CrossRef]
- Rillig, M.C.; Aguilar-Trigueros, C.A.; Bergmann, J.; Verbruggen, E.; Veresoglou, S.; Lehmann, A. Plant root and mycorrhizal fungal traits for understanding soil aggregation. New Phytol. 2015, 205, 1385–1388. [Google Scholar] [CrossRef]
- Chandrasekaran, M.; Boopathi, T.; Manivannan, P. Comprehensive assessment of ameliorative effects of AM fungi in alleviating abiotic stress in tomato plants. J. Fungi 2021, 7, 303. [Google Scholar] [CrossRef] [PubMed]
- Shi, Z.; Zhang, J.; Lu, S.; Li, Y.; Wang, F. Arbuscular mycorrhizal fungi improve the performance of sweet sorghum grown in a mo-contaminated soil. J. Fungi 2020, 6, 44. [Google Scholar] [CrossRef] [Green Version]
- Chen, M.; Arato, M.; Borghi, L.; Nouri, E.; Reinhardt, D. Beneficial services of arbuscular mycorrhizal fungi—From ecology to application. Front. Plant Sci. 2018, 9, 1270. [Google Scholar] [CrossRef]
- Pellegrino, E.; Piazza, G.; Arduini, I.; Ercoli, L. Field inoculation of bread wheat with Rhizophagus irregularis under organic farming: Variability in growth response and nutritional uptake of eleven old genotypes and a modern variety. Agronomy 2020, 10, 333. [Google Scholar] [CrossRef] [Green Version]
- Bago, B.; Bécard, G. Bases of the obligate biotrophy of arbuscular mycorrhizal fungi. In Mycorrhizal Technology in Agriculture; Gianinazzi, S., Schüepp, H., Barea, J.M., Haselwandter, K., Eds.; Birkhäusel: Basel, Switzerland, 2002. [Google Scholar]
- Fortin, J.A.; Bécard, G.; Declerck, S.; Dalpé, Y.; St-Arnaud, M.; Coughlan, A.P.; Piché, Y. Arbuscular mycorrhiza on root-organ cultures. Can. J. Bot. 2002, 80, 1–20. [Google Scholar] [CrossRef]
- Adholeya, A.; Tiwari, P.; Singh, R. Large-scale production of arbuscular mycorrhizal fungi on root organs and inoculation strategies. In In Vitro Culture of Mycorrhizas; Declerck, S., Strullu, D.G., Fortin, J.A., Eds.; Springer: Heidelberg/Berlin, Germany, 2005; pp. 315–338. [Google Scholar]
- Trejo-Aguilar, D.; Banuelos, J. Isolation and culture of arbuscular mycorrhizal fungi from field samples. In Arbuscular Mycorrhizal Fungi. Methods in Molecular Biology; Ferrol, N., Lanfranco, L., Eds.; Humana: New York, NY, USA, 2020; Volume 2146. [Google Scholar]
- Saito, M.; Marumoto, T. Inoculation with arbuscular mycorrhizal fungi: The status quo in Japan and the future prospects. Plant Soil 2002, 244, 273–279. [Google Scholar] [CrossRef]
- Huang, L.L.; Sylvia, D.M. Production of vesicular arbuscular mycorrhizal fungus inoculum in aeroponic culture. Appl. Enviorn. Microbiol. 1988, 54, 553–555. [Google Scholar] [CrossRef] [Green Version]
- Douds, D.D., Jr.; Nagahashi, G.; Pfeffer, P.E.; Reider, C.; Kayser, W.M. On-farm production of AM fungus inoculum in mixtures of compost and vermiculite. Biores. Tech. 2006, 97, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Gaur, A.; Varma, A. Research methods in arbuscular mycorrhizal fungi. In Advanced Techniques in Soil Microbiology; Gabler: Wiesbaden, Germany, 2007; pp. 377–396. [Google Scholar]
- Gaur, A.; Adholeya, A. Arbuscular-mycorrhizal inoculation of five tropical fodder crops and inoculum production in marginal soil amended with organic matter. Biol. Fertil. Soils 2002, 35, 214–218. [Google Scholar] [CrossRef]
- Jarstfer, A.G.; Sylvia, D.M. Inoculum production and inoculation strategies for vesicular-arbuscular mycorrhizal fungi. In Soil Microbial Tecnologies: Applications in Agriculture, Forestry and Environmental Management; Dekker, M.B.M., Ed.; Marcel Dekker: New York, NY, USA, 1992; pp. 349–377. [Google Scholar]
- Silva, F.S.B.; Yano-Melo, A.M.; Brandão, J.A.C.; Maia, L.C. Sporulation of arbuscular mycorrhizal fungi using Tris-HCl buffer in addition to nutrient solutions. Braz. J. Microbiol. 2005, 36, 327–332. [Google Scholar] [CrossRef]
- Gaur, A.; Adholeya, A. Effects of the particle size of soil-less substrates upon AM fungus inoculum production. Mycorrhiza 2000, 10, 43–48. [Google Scholar] [CrossRef]
- Baranski, R. Genetic transformation of carrot (Daucus carota) and other apiaceae species. Transgenic Plant J. 2008, 2, 18–38. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Fortin, J.A.; St-Arnaud, M.; Hamel, C.; Chaverie, C.; Jolicoeur, M. Aseptic In Vitro Endomycorrhizal Spore Mass Production. 1996. US Patent 5554530, 6 August 1993. [Google Scholar]
- Bécard, G.; Fortin, J.A. Early events of vesicular–arbuscular mycorrhiza formation on Ri T-DNA transformed roots. New Phytol. 1988, 108, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Maitra, P.; Zheng, Y.; Chen, L.; Wang, Y.-L.; Ji, N.-N.; Lü, P.-P.; Gan, H.-Y.; Li, X.C.; Sun, X.; Zhou, X.H.; et al. Effect of drought and season on arbuscular mycorrhizal fungi in a subtropical secondary forest. Fungal Ecol. 2019, 41, 107–115. [Google Scholar] [CrossRef]
- Hoagland, D.R. Optimum nutrient solutions for plants. Science 1920, 52, 562–564. [Google Scholar] [CrossRef] [Green Version]
- Maitra, P.; Zheng, Y.; Wang, Y.-L.; Mandal, D.; Lü, P.-P.; Gao, C.; Babalola, B.J.; Ji, N.-N.; Li, X.-C.; Guo, L.-D. Phosphorus fertilization rather than nitrogen fertilization, growing season and plant successional stage structures arbuscular mycorrhizal fungal community in a subtropical forest. Biol. Fertil. Soils 2021, 57, 685–697. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 10 September 2021).
- Pohlert, T. The Pairwise Multiple Comparison of Mean Ranks Package (PCMR). 2 R Package Version 4.1. 2014. Available online: http://cran.r-project/package=PCMR (accessed on 14 April 2021).
- Zeng, T.; Holmer, R.; Hontelez, J.; Lintel-Hekkert, B.T.; Marufu, L.; De Zeeuw, T.; Wu, F.; Schijlen, E.; Bisseling, T.; Limpens, E. Host- and stage-dependent secretome of the arbuscular mycorrhizal fungus Rhizophagus irregularis. Plant J. 2018, 94, 411–425. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.; Wang, Y.; Liu, M.; Li, Q.; Xiao, W.; Song, X. Effects of nitrogen deposition and phosphorus addition on arbuscular mycorrhizal fungi of Chinese fir (Cunninghamia lanceolata). Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef]
- Millner, P.D.; Kitt, D.G. The Beltsville method for soilless production of vesicular-arbuscular mycorrhizal fungi. Mycorrhiza 1992, 2, 9–15. [Google Scholar] [CrossRef]
- Nouri, E.; Breuillin-Sessoms, F.; Feller, U.; Reinhardt, D. Phosphorus and nitrogen regulate arbuscular mycorrhizal symbiosis in petunia hybrida. PLoS ONE 2014, 9, e90841. [Google Scholar] [CrossRef] [PubMed]
- Lü, P.-P.; Zheng, Y.; Chen, L.; Ji, N.-N.; Yao, H.; Maitra, P.; Hu, H.-W.; Li, X.-C.; Guo, L.-D. Irrigation and fertilization effects on arbuscular mycorrhizal fungi depend on growing season in a dryland maize agroecosystem. Pedobiologia 2020, 83, 150687. [Google Scholar] [CrossRef]
- Breuillin, F.; Schramm, J.; Hajirezaei, M.; Ahkami, A.; Favre, P.; Druege, U.; Hause, B.; Bucher, M.; Kretzschmar, T.; Bossolini, E.; et al. Phosphate systemically inhibits development of arbuscular mycorrhiza in Petunia hybrida and represses genes involved in mycorrhizal functioning. Plant J. 2010, 64, 1002–1017. [Google Scholar] [CrossRef] [PubMed]
- Blanke, V.; Renker, C.; Wagner, M.; Füllner, K.; Held, M.; Kuhn, A.J.; Buscot, F. Nitrogen supply affects arbuscular mycorrhizal colonization of Artemisia vulgaris in a phosphate-polluted field site. New Phytol. 2005, 166, 981–992. [Google Scholar] [CrossRef]
- Douds, D.D., Jr.; Schenck, N.C. Increased sporulation of vesicular–arbuscular mycorrhizal fungi by manipulation of nutrient regimens. Appl. Environ. Microbiol. 1990, 56, 413–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douds, D.D., Jr.; Schenck, N.C. Relationship of colonization and sporulation by VA mycorrhizal fungi to plant nutrient and carbohydrate contents. New Phytol. 1990, 116, 621–627. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | d.f. | Spore Density | Colonization | Shoot Weight | Root Weight | ||||
|---|---|---|---|---|---|---|---|---|---|
| χ2 | P | χ2 | P | F | P | F | P | ||
| Sand particle size (SP) | 3 | 39.10 | <0.001 | 27.52 | <0.001 | 17.16 | <0.001 | 2.440 | 0.070 |
| Nutrient strength (NS) | 1 | 38.48 | <0.001 | 47.64 | <0.001 | 51.22 | <0.001 | 45.33 | <0.001 |
| Phosphorus omission (PO) | 1 | 9.360 | 0.002 | 7.753 | 0.005 | 28.72 | <0.001 | 23.53 | <0.001 |
| SP × NS | 3 | na | na | na | na | 2.196 | 0.039 | 2.804 | 0.045 |
| SP × PO | 3 | na | na | na | na | 0.195 | 0.899 | 0.242 | 0.866 |
| NS × PO | 1 | na | na | na | na | 4.144 | 0.045 | 6.591 | 0.012 |
| SP × NS × PO | 3 | na | na | na | na | 0.721 | 0.542 | 0.533 | 0.661 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maitra, P.; Al-Rashid, J.; Barman, N.C.; Khan, M.N.M.; Mandal, D.; Rasul, N.M.; Chowdhury, A.; El-Sappah, A.H.; Li, J. Sand Particle Size and Phosphorus Amount Affect Rhizophagus irregularis Spore Production Using In Vitro Propagated Spore as a Starter Inoculum in Rhizosphere of Maize (Zea mays) Plantlets. J. Fungi 2021, 7, 846. https://doi.org/10.3390/jof7100846
Maitra P, Al-Rashid J, Barman NC, Khan MNM, Mandal D, Rasul NM, Chowdhury A, El-Sappah AH, Li J. Sand Particle Size and Phosphorus Amount Affect Rhizophagus irregularis Spore Production Using In Vitro Propagated Spore as a Starter Inoculum in Rhizosphere of Maize (Zea mays) Plantlets. Journal of Fungi. 2021; 7(10):846. https://doi.org/10.3390/jof7100846
Chicago/Turabian StyleMaitra, Pulak, Jubair Al-Rashid, Nirmal Chandra Barman, Md. Niuz Morshed Khan, Dipa Mandal, Noorain Munim Rasul, Apu Chowdhury, Ahmed H. El-Sappah, and Jia Li. 2021. "Sand Particle Size and Phosphorus Amount Affect Rhizophagus irregularis Spore Production Using In Vitro Propagated Spore as a Starter Inoculum in Rhizosphere of Maize (Zea mays) Plantlets" Journal of Fungi 7, no. 10: 846. https://doi.org/10.3390/jof7100846
APA StyleMaitra, P., Al-Rashid, J., Barman, N. C., Khan, M. N. M., Mandal, D., Rasul, N. M., Chowdhury, A., El-Sappah, A. H., & Li, J. (2021). Sand Particle Size and Phosphorus Amount Affect Rhizophagus irregularis Spore Production Using In Vitro Propagated Spore as a Starter Inoculum in Rhizosphere of Maize (Zea mays) Plantlets. Journal of Fungi, 7(10), 846. https://doi.org/10.3390/jof7100846