
Low Dose of Fluoride in the Culture Medium of Cordyceps militaris Promotes Its Growth and Enhances Bioactives with Antioxidant and Anticancer Properties

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Lines
2.3. Cordyceps militaris Cultivation
2.4. Fluoride Detection and Quantification
- (1)
- The sample m (x)g and the internal standard 1,1,1,3,3,3-hexafluoroisopropanol n(std) mol were mixed in 1 mL of MeOD-d4.
- (2)
- The above mixture (600 μL) was analyzed using 19F NMR, and the ratio of the area of the sample to the internal standard was obtained, which was equal to the molar ratio of fluorine in the sample and 1,1,1,3,3,3-hexafluoroisopropanol: Area(std)/Area(x) = 6 × n(std)/n(x)
- (3)
- The quantification of fluorine in C. militaris was estimated as follows: 19 × n(x)/m(x) × 100%
2.5. Carotenoid Quantification
2.6. Preparation of Aqueous Extracts from C. militaris Fruiting Bodies
2.7. Antioxidant Activity Assay
2.7.1. Assay for SOD-Like Activity
2.7.2. DPPH Radical-Scavenging Activity Assay
2.8. Anticancer Activity Assays
2.8.1. Cell Viability Assay
2.8.2. Apoptosis Assay
2.8.3. Cell Cycle Assay
2.8.4. Western Blotting Assay
2.9. Statistical Analysis
3. Results and Discussion
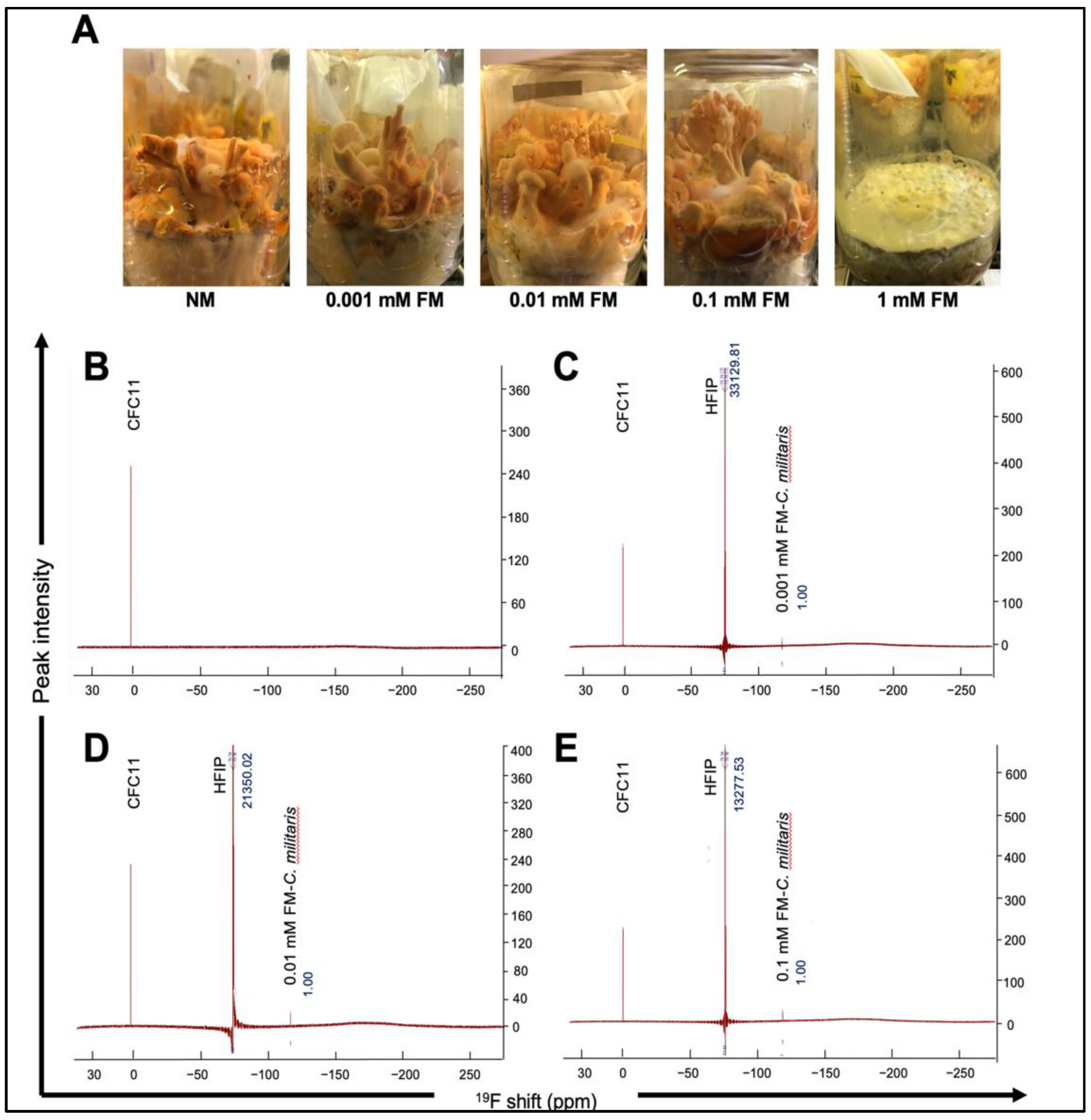
3.1. Dry Weight and Total Carotenoid Content of C. militaris Raised in the Normal (NM) and Flouride-Supplemented (FM) Medium
3.2. 19F NMR Spectra of NM and FM Raised C. militaris
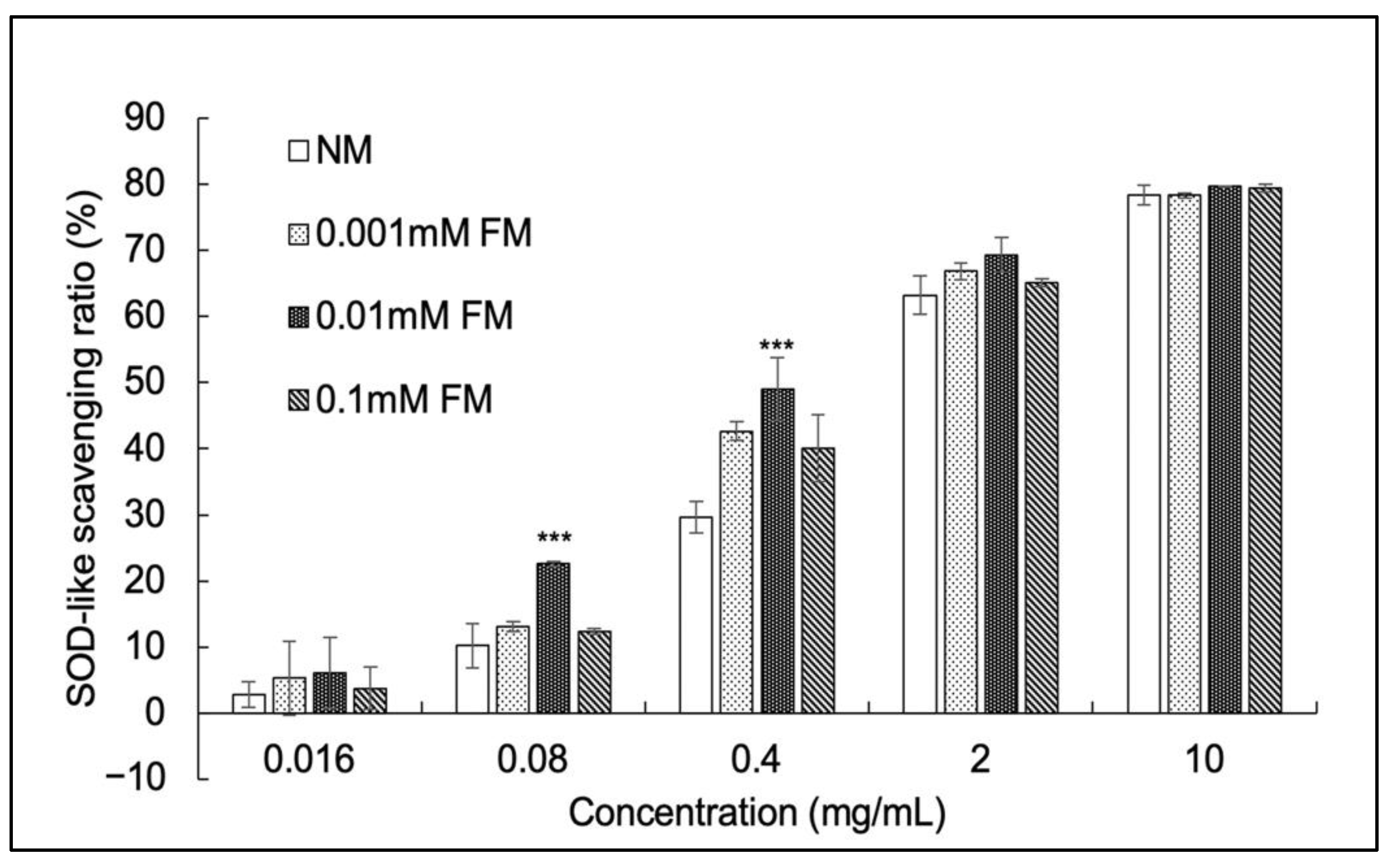
3.3. SOD-Like Activity Assay
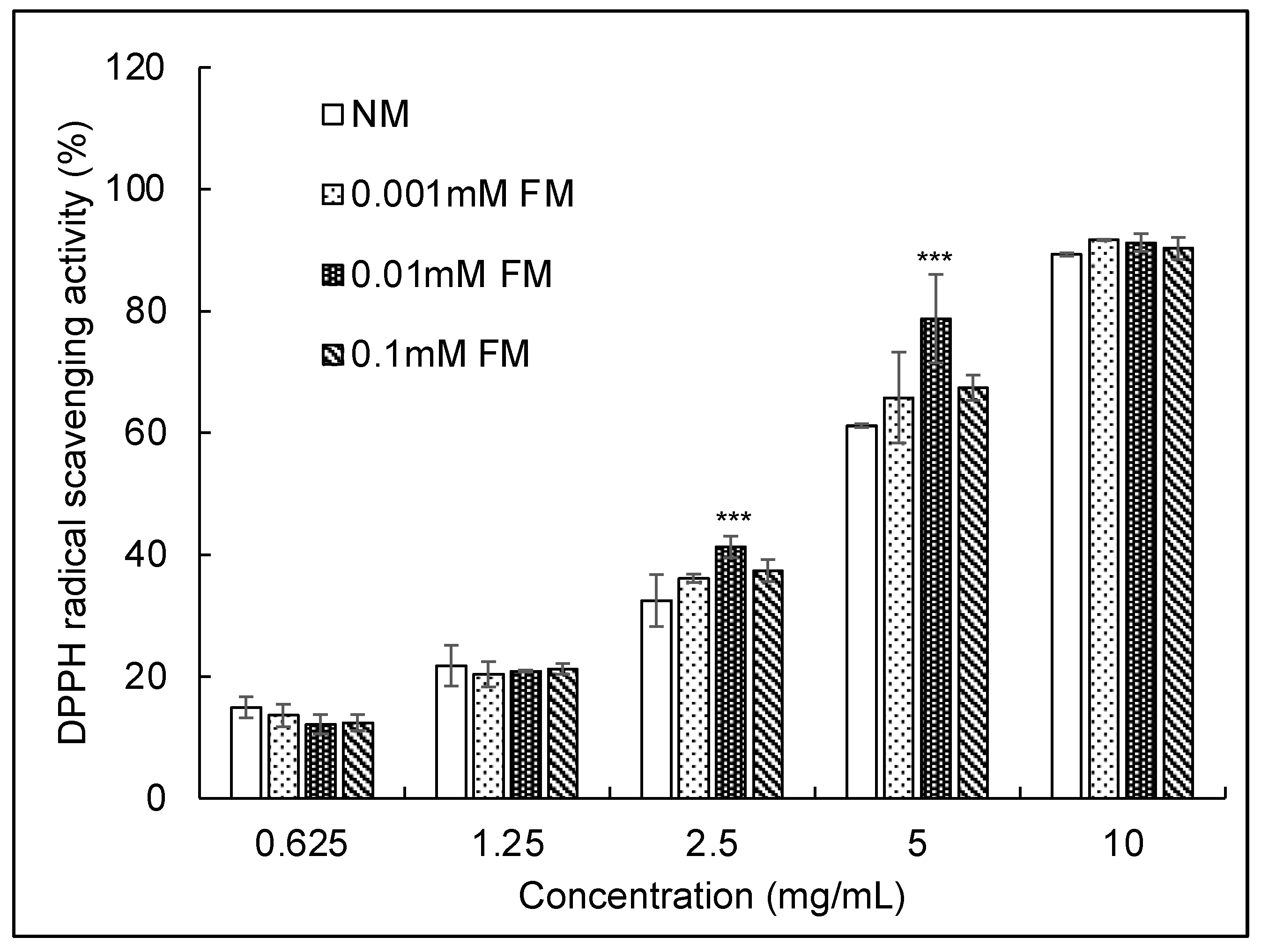
3.4. DPPH Radical Scavenging Activity Assay
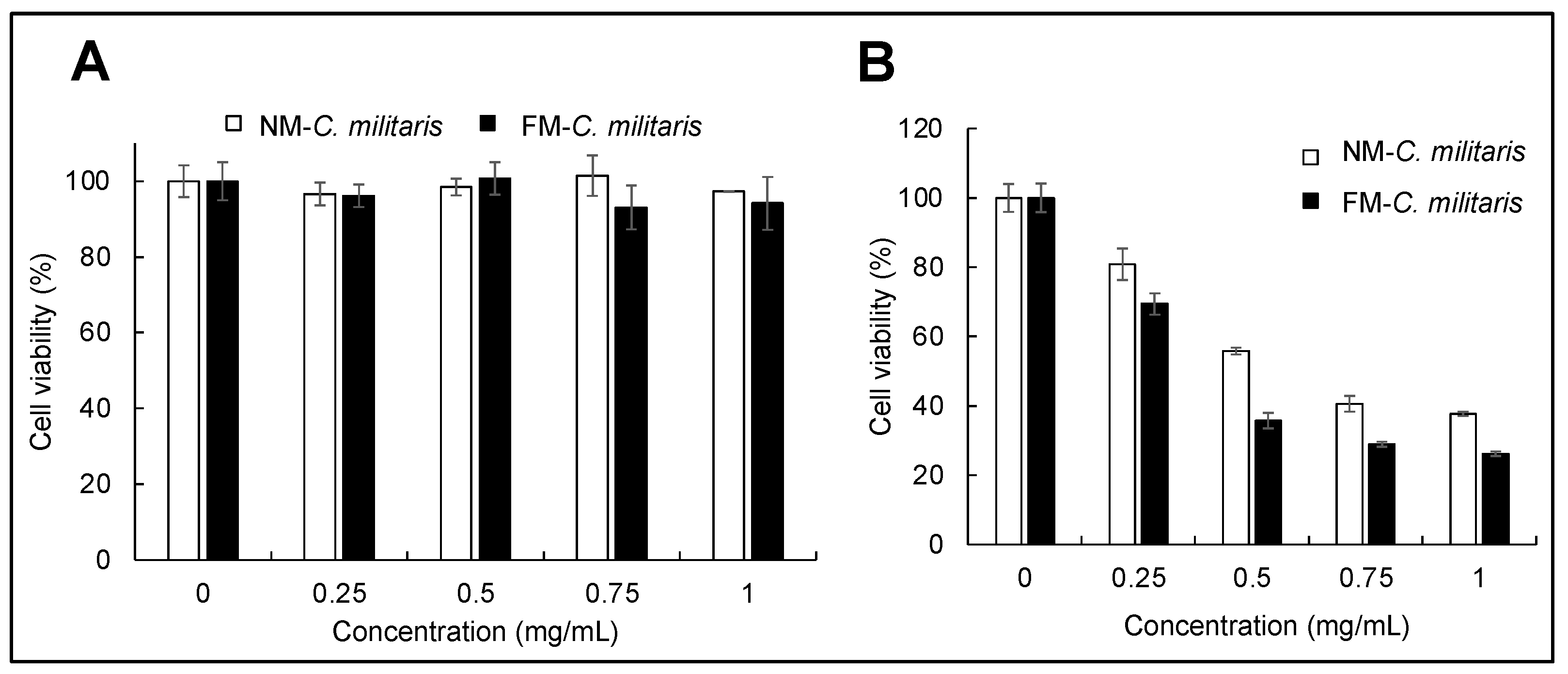
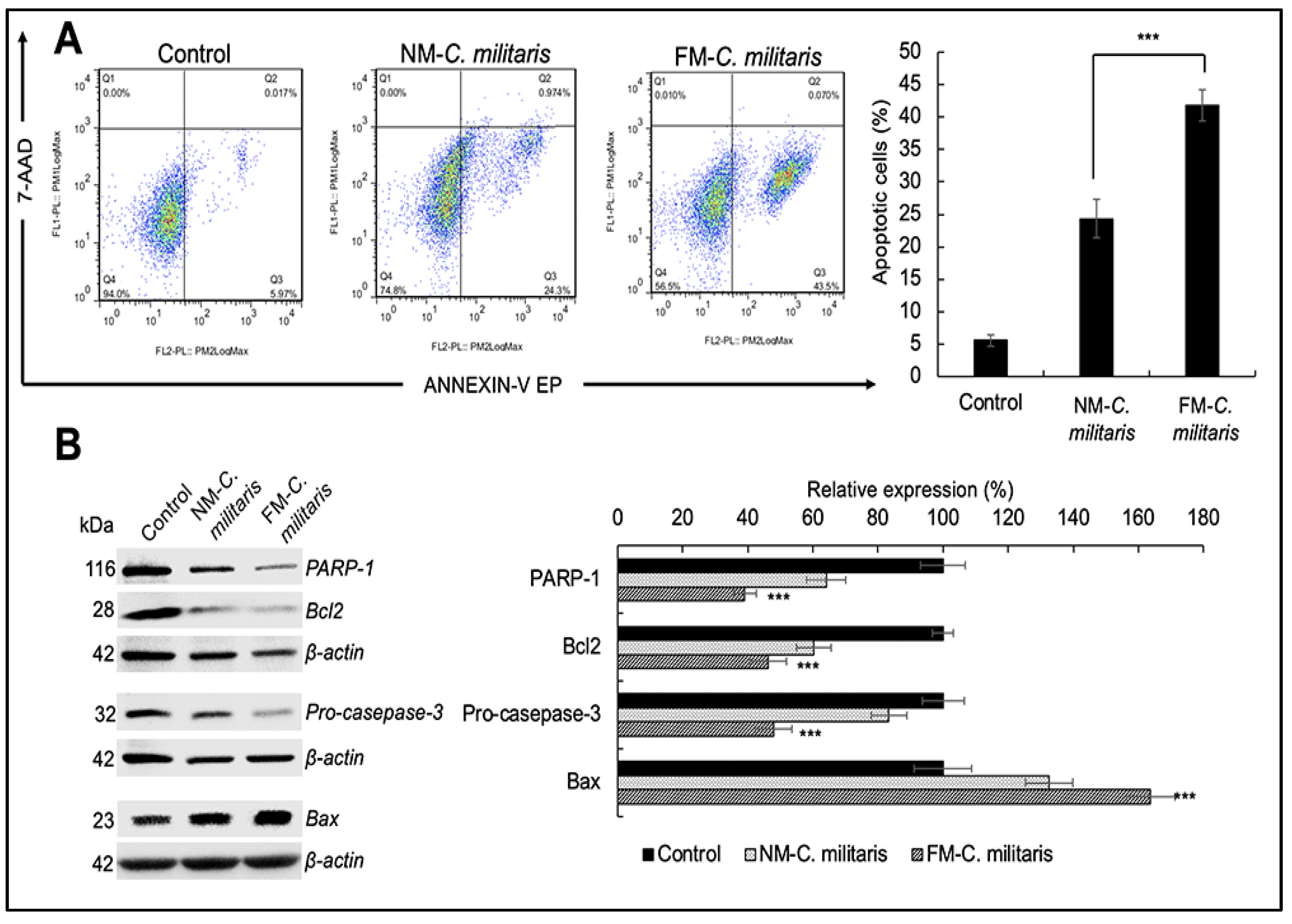
3.5. FM-C. militaris Extracts Showed Enhanced Cytotoxicity in Cancer Cells
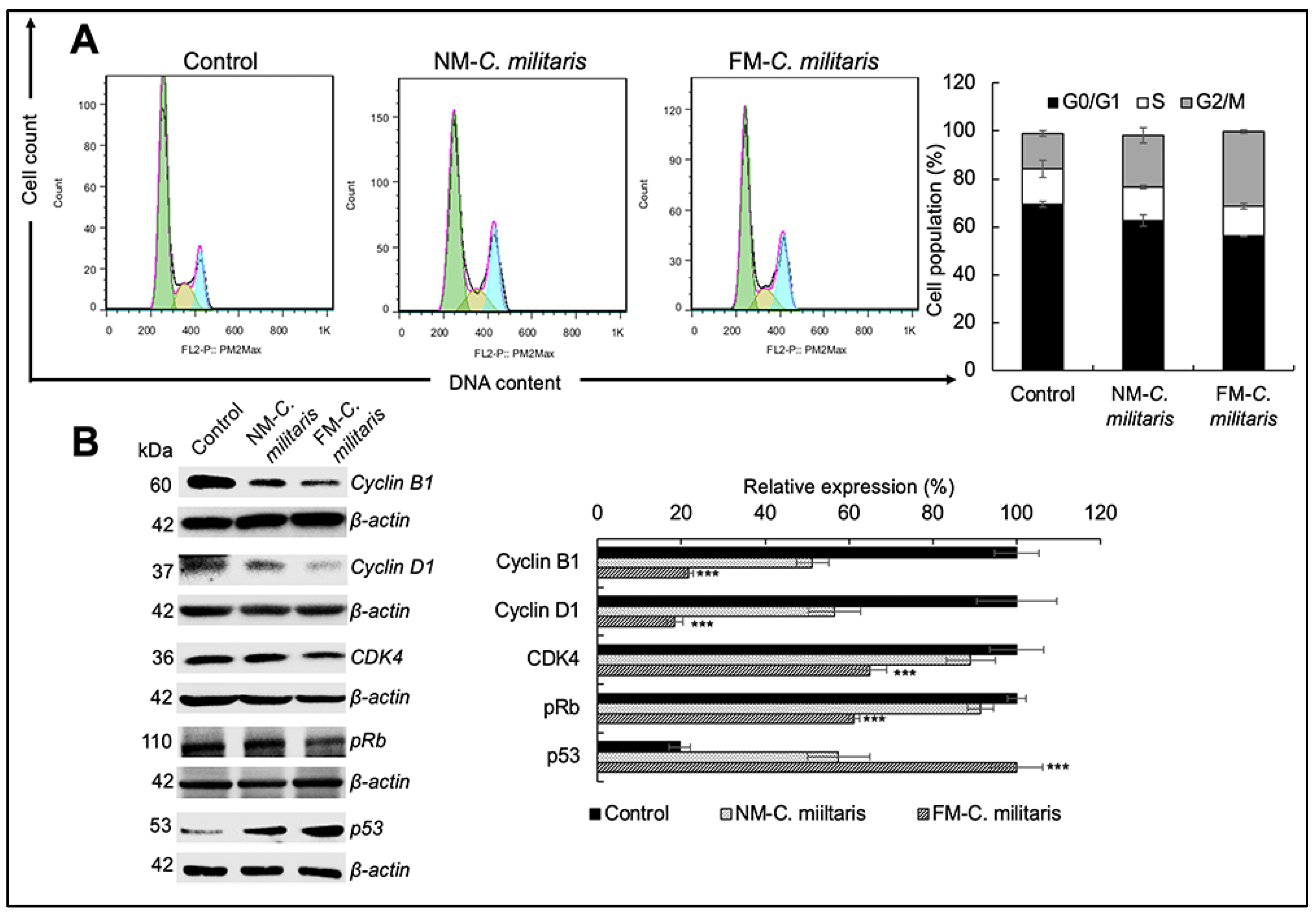
3.6. FM-C. militaris Extracts Caused Enhanced Apoptosis and Cell Cycle Arrest in Cancer Cells
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Wu, L.; Sun, H.; Hao, Y.; Zheng, X.; Song, Q.; Dai, S.; Zhu, Z. Chemical structure and inhibition on α-glucosidase of the polysaccharides from Cordyceps militaris with different developmental stages. Int. J. Biol. Macromol. 2020, 148, 722–736. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, B.; Lin, R.; Jia, L.; Deng, P.; Fan, K.; Wang, G.; Wang, L.; Zhang, J. Extraction and antioxidant activities of intracellular polysaccharide from Pleurotus sp. Mycelium. Int. J. Biol. Macromol. 2010, 47, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Aida, F.M.N.A.; Shuhaimi, M.; Yazid, M.; Maaruf, A.G. Mushroom as a potential source of prebiotics: A review. Trends Food Sci. Technol. 2009, 20, 567–575. [Google Scholar] [CrossRef]
- Ng, T.B.; Wang, H.X. Pharmacological actions of Cordyceps, a prized folk medicine. J. Pharm. Pharmacol. 2005, 57, 1509–1519. [Google Scholar] [CrossRef]
- Lin, L.; Xu, J. Fungal Pigments and Their Roles Associated with Human Health. J. Fungi 2020, 6, 280. [Google Scholar] [CrossRef] [PubMed]
- Nabi, F.; Arain, M.A.; Rajput, N.; Alagawany, M.; Soomro, J.; Umer, M.; Soomro, F.; Wang, Z.; Ye, R.; Liu, J. Health benefits of carotenoids and potential application in poultry industry: A review. J. Anim. Physiol. Anim. Nutr. 2020, 104, 1809–1818. [Google Scholar] [CrossRef]
- Venil, C.K.; Velmurugan, P.; Dufosse, L.; Devi, P.R.; Ravi, A.V. Fungal Pigments: Potential Coloring Compounds for Wide Ranging Applications in Textile Dyeing. J. Fungi 2020, 6, 68. [Google Scholar] [CrossRef]
- Bhatt, T.; Patel, K. Carotenoids: Potent to Prevent Diseases Review. Nat. Prod. Bioprospect. 2020, 10, 109–117. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoids as natural functional pigments. J. Nat. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Li, S.P.; Yang, F.Q.; Tsim, K.W. Quality control of Cordyceps sinensis, a valued traditional Chinese medicine. J. Pharm. Biomed. Anal. 2006, 41, 1571–1584. [Google Scholar] [CrossRef] [PubMed]
- Yue-Qin, C.; Ning, W.; Hui, Z.; Liang-Hu, Q. Differentiation of medicinal Cordyceps species by rDNA ITS sequence analysis. Planta Med. 2002, 68, 635–639. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.S.; Yang, C.H.; Lu, K.; Huang, K.S.; Zheng, Y.Z. Synthesis of agar microparticles using temperature-controlled microfluidic devices for Cordyceps militaris cultivation. Electrophoresis 2011, 32, 3157–3163. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Dai, X.; Xu, F.; Wang, F.; Gong, B.; Wei, Y. Preparation of imidazole-functionalized silica by surface-initiated atom transfer radical polymerization and its application for hydrophilic interaction chromatography. Anal. Bioanal. Chem. 2012, 404, 1477–1484. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zeng, Y.; Cui, Y.; Liu, H.; Dong, C.; Sun, Y. Structural characterization, antioxidant and immunomodulatory activities of a neutral polysaccharide from Cordyceps militaris cultivated on hull-less barley. Carbohydr. Polym. 2020, 235, 115969. [Google Scholar] [CrossRef]
- Tang, J.; Qian, Z.; Wu, H. Enhancing cordycepin production in liquid static cultivation of Cordyceps militaris by adding vegetable oils as the secondary carbon source. Bioresour. Technol. 2018, 268, 60–67. [Google Scholar] [CrossRef]
- Dong, J.Z.; Lei, C.; Ai, X.R.; Wang, Y. Selenium enrichment on Cordyceps militaris link and analysis on its main active components. Appl. Biochem. Biotechnol. 2012, 166, 1215–1224. [Google Scholar] [CrossRef]
- Shih, I.L.; Tsai, K.L.; Hsieh, C. Effects of culture conditions on the mycelial growth and bioactive metabolite production in submerged culture of Cordyceps militaris. Biochem. Eng. J. 2007, 33, 193–201. [Google Scholar] [CrossRef]
- Xie, C.; Gu, Z.; Fan, G.; Gu, F.; Han, Y.; Chen, Z. Production of cordycepin and mycelia by submerged fermentation of Cordyceps militaris in mixture natural culture. Appl. Biochem. Biotechnol. 2009, 158, 483–492. [Google Scholar] [CrossRef]
- Hong, B.D.; Joo, R.N.; Lee, K.S.; Lee, D.S.; Rhie, J.H.; Min, S.W.; Song, S.G.; Chung, D.Y. Fluoride in soil and plant. Korean J. Agric. Sci. 2016, 43, 522–536. [Google Scholar] [CrossRef]
- Ken, J.; Gordon, S. Fluoroproducts-the extremophiles. J. Fluor. Chem. 2000, 104, 5–18. [Google Scholar] [CrossRef]
- Böhm, H.J.; Banner, D.; Bendels, S.; Kansy, M.; Kuhn, B.; Müller, K.; Ulrike, O.S.; Stahl, M. Fluorine in medicinal chemistry. ChemBioChem 2004, 5, 637–643. [Google Scholar] [CrossRef]
- Gillis, E.P.; Eastman, K.J.; Hill, M.D.; Donnelly, D.J.; Meanwell, N.A. Applications of fluorine in medicinal chemistry. J. Med. Chem. 2015, 58, 8315–8359. [Google Scholar] [CrossRef]
- Barbier, O.; Laura, A.M.; Del Razo, L.M. Molecular mechanisms of fluoride toxicity. Chem. Biol. Interact. 2010, 188, 319–333. [Google Scholar] [CrossRef]
- Jha, S.K.; Nayak, A.K.; Sharma, Y.K. Fluoride toxicity effects in onion (Allium cepa L.) grown in contaminated soils. Chemosphere 2009, 76, 353–356. [Google Scholar] [CrossRef]
- Doley, D. Fluoride-induced enhancement and inhibition of shoot growth in four taxa of Pinus. New Phytol. 1989, 112, 543–552. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Li, P.; Zhao, X. Effect of fluoride on carbonic anhydrase activity and photosynthetic oxygen evolution of the algae Chlamydomonas reinhardtii. Fluoride 2007, 40, 51–54. [Google Scholar]
- Zhan, X.A.; Xu, Z.R.; Li, J.X.; Wang, M. Effects of fluorosis on lipid peroxidation and antioxidant systems in young pigs. Fluoride 2005, 38, 157–161. [Google Scholar]
- Chen, Y. Differences in fluoride effects on fecundity among varieties of the silkworm Bombyx mori. Fluoride 2003, 36, 163–169. [Google Scholar]
- Burgstahler, A.W. Paradoxical dose-response effects of fluoride. Fluoride 2002, 35, 143–147. [Google Scholar]
- Calabrese, E.J.; Blain, R. The hormesis database: The occurrence of hormetic dose responses in the toxicological literature. Regul. Toxicol. Pharmacol. 2011, 61, 73–81. [Google Scholar] [CrossRef]
- Hasegawa, J. The Determination of carotene content in foods. Agric. Biol. Chem. 1990, 54, 1819–1827. [Google Scholar]
- Yuan, X.; Sun, H.; Liu, Y.; Shiroshita, T.; Kawano, S.; Takeshi, S.; Ma, J.; Zhang, Z. Anti-cancer activity comparisons of aqueous extracts from Inonotus obliquus, Cordyceps militaris and Uncaria tomentosa in vitro and in vivo. J. Pharmacogn. Phytochem. 2014, 2, 19–25. [Google Scholar]
- Chae, Y.; Kim, D.; An, Y.J. Effects of fluorine on crops, soil exoenzyme activities, and earthworms in terrestrial ecosystems. Ecotoxicol. Environ. Saf. 2018, 151, 21–27. [Google Scholar] [CrossRef]
- Yang, T.; Sun, J.; Lian, T.; Wang, W.; Dong, C. Process optimization for extraction of carotenoids from medicinal caterpillar fungus, Cordyceps militaris (Ascomycetes). Int. J. Med. Mushrooms 2014, 16, 125–135. [Google Scholar] [CrossRef]
- Milani, A.; Basirnejad, M.; Shahbazi, S.; Bolhassani, A. Carotenoids: Biochemistry, pharmacology and treatment. Br. J. Pharmacol. 2017, 174, 1290–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Q.; Wei, T.; Lin, Y.; Ye, Z.W.; Lin, J.F.; Guo, L.Q.; Yun, F.; Kang, L. Developing a novel two-stage process for carotenoid production by Cordyceps militaris (Ascomycetes). Int. J. Med. Mushrooms 2019, 21, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Ye, Z.W.; Liu, C.; Guo, L.Q.; Lin, J.F.; Wang, H.; Fan, Y.; Kang, L. Increasing of the contain of carotenoids in Caterpillar mushroom, Cordyceps militaris (Ascomycetes) by using the fungal elicitors cultivation. Int. J. Med. Mushrooms 2019, 21, 1181–1191. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.Z.; Lei, C.; Zheng, X.J.; Ai, X.R.; Wang, Y.; Wang, Q. Light Wavelengths Regulate Growth and Active Components of Cordyceps militaris Fruit Bodies. J. Nutr. Biochem. 2013, 37, 578–584. [Google Scholar] [CrossRef]
- Reddy, M.P.; Kaur, M. Sodium fluoride induced growth and metabolic changes in Salicornia brachiata Roxb. Water Air Soil Pollut. 2008, 188, 171–179. [Google Scholar] [CrossRef]
- Janiszewska, J.; Balcerzak, M. Analytical problems with the evaluation of human exposure to fluorides from tea products. Food Anal. Methods 2013, 6, 1090–1098. [Google Scholar] [CrossRef] [Green Version]
- Gago, C.; Romar, A.; Fernández-Marcos, M.L.; Álvarez, E. Fluoride sorption and desorption on soils located in the surroundings of an aluminium smelter in Galicia (NW Spain). Environ. Earth Sci. 2014, 72, 4105–4114. [Google Scholar] [CrossRef]
- O’Hagan, D.; Harper, D.B. Fluorine-containing natural products. J. Fluor. Chem. 1999, 100, 127–133. [Google Scholar] [CrossRef]
- Walker, M.C.; Chang, M.C. Natural and engineered biosynthesis of fluorinated natural products. Chem. Soc. Rev. 2014, 43, 6527–6536. [Google Scholar] [CrossRef] [Green Version]
- Shamsipur, M.; Sarkouhi, M.; Hassan, J.; Haghgoo, S. Fluorine-19 nuclear magnetic resonance (19F NMR) as a powerful technique for the assay of anti-HIV drug efavirenz in human serum and pharmaceutical formulations. Afr. J. Pharm. Pharmacol. 2011, 5, 1573–1579. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Zhu, X.; Peng, C.; Xu, W.; Li, D.; Wang, Y.; Fang, S.; Li, Y.; Hu, S.; Wan, X. Critical factors determining fluoride concentration in tea leaves produced from Anhui province, China. Ecotoxicol. Environ. Saf. 2016, 131, 14–21. [Google Scholar] [CrossRef]
- Dong, J.Z.; Liu, M.R.; Lei, C.; Zheng, X.J.; Wang, Y. Effects of selenium and light wavelengths on liquid culture of Cordyceps militaris Link. Appl. Biochem. Biotechnol. 2012, 166, 2030–2036. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ban, L.; Meng, S.; Huang, L.; Sun, N.; Yang, H.; Wang, Y.; Wang, L. Bioactivities of crude polysaccharide extracted from fermented soybean curd residue by Cordyceps militaris. Biotechnol. Biotechnol. Equip. 2021, 35, 385–396. [Google Scholar] [CrossRef]
- Yadu, B.; Chandrakar, V.; Keshavkant, S. Responses of plants to fluoride: An overview of oxidative stress and defense mechanisms. Fluoride 2016, 49 (3 Pt 2), 293–302. [Google Scholar]
- Shivarajashankara, Y.M.; Shivashankara, A.R.; Bhat, P.G.; Rao, S.H. Effect of fluoride intoxication on lipid peroxidation and antioxidant systems in rats. Fluoride 2001, 34, 108–113. [Google Scholar]
- Chakrabarti, S.; Patra, P.K. Biochemical and antioxidant responses of paddy (Oryza sativa L.) to fluoride stress. Fluoride 2015, 48, 56–61. [Google Scholar]
- Chakrabarti, S.; Patra, P.K. Effect of fluoride on superoxide dismutase activity in four common crop plants. Fluoride 2013, 46, 59–62. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C. militaris Samples | Dry Weight (g/bottle) | TCC (µg/g) |
|---|---|---|
| NM | 1.07 ± 0.07 | 2561.27 ± 26.56 |
| 0.001 mM FM | 1.16 ± 0.11 | 2751.00 ± 21.36 |
| 0.01 mM FM | 1.55 ± 0.17 | 3161.38 ± 35.71 |
| 0.1 mM FM | 1.28 ± 0.14 | 3035.96 ± 53.30 |
| C. militaris Sample | Mass (x) g | n(std) mol | A(std)/A(x) | Fluorine (ppm) |
|---|---|---|---|---|
| 0.001 mM FM | 0.13 | 5.7 × 10−4 | 33,129.81 | 15.09 |
| 0.01 mM FM | 0.09 | 5.7 × 10−4 | 21,350.02 | 33.81 |
| 0.1 mM FM | 0.09 | 5.7 × 10−4 | 13,277.53 | 54.38 |
| C. militaris Sample | DPPH IC50 (mg/mL) | SOD IC50 (mg/mL) | SOD Vigor (U/mg) |
|---|---|---|---|
| NM | 4.16 | 1.13 | 44.25 |
| 0.001 mM FM | 2.92 | 0.81 | 61.37 |
| 0.01 mM FM | 2.59 | 0.59 | 84.75 |
| 0.1 mM FM | 2.89 | 0.87 | 57.47 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Wang, J.; Zhang, H.; Xiao, L.; Lei, Z.; Kaul, S.C.; Wadhwa, R.; Zhang, Z. Low Dose of Fluoride in the Culture Medium of Cordyceps militaris Promotes Its Growth and Enhances Bioactives with Antioxidant and Anticancer Properties. J. Fungi 2021, 7, 342. https://doi.org/10.3390/jof7050342
Li X, Wang J, Zhang H, Xiao L, Lei Z, Kaul SC, Wadhwa R, Zhang Z. Low Dose of Fluoride in the Culture Medium of Cordyceps militaris Promotes Its Growth and Enhances Bioactives with Antioxidant and Anticancer Properties. Journal of Fungi. 2021; 7(5):342. https://doi.org/10.3390/jof7050342
Chicago/Turabian StyleLi, Xiaoshuai, Jia Wang, Huayue Zhang, Long Xiao, Zhongfang Lei, Sunil C. Kaul, Renu Wadhwa, and Zhenya Zhang. 2021. "Low Dose of Fluoride in the Culture Medium of Cordyceps militaris Promotes Its Growth and Enhances Bioactives with Antioxidant and Anticancer Properties" Journal of Fungi 7, no. 5: 342. https://doi.org/10.3390/jof7050342
APA StyleLi, X., Wang, J., Zhang, H., Xiao, L., Lei, Z., Kaul, S. C., Wadhwa, R., & Zhang, Z. (2021). Low Dose of Fluoride in the Culture Medium of Cordyceps militaris Promotes Its Growth and Enhances Bioactives with Antioxidant and Anticancer Properties. Journal of Fungi, 7(5), 342. https://doi.org/10.3390/jof7050342