Abstract
The genus Cryptococcus comprises more than 80 species, including C. neoformans and C. gattii, which are pathogenic to humans, mainly affecting the central nervous system. The two species differ in geographic distribution and environmental niche. C. neoformans has a worldwide distribution and is often isolated from bird droppings. On the contrary, C. gattii is reported in tropical and subtropical regions and is associated with Eucalyptus species. This review aims to describe the distribution of environmental isolates of the Cryptococcus neoformans species complex and the Cryptococcus gattii species complex in Colombia. A systematic investigation was carried out using different databases, excluding studies of clinical isolates reported in the country. The complex of the species of C. gattii is recovered mainly from trees of the genus Eucalyptus spp., while the complex of the species of C. neoformans is recovered mainly from avian excrement, primarily Columba livia (pigeons) excrement. In addition, greater positivity was found at high levels of relative humidity. Likewise, an association was observed between the presence of the fungus in places with little insolation and cold or temperate temperatures compared to regions with high temperatures.
1. Introduction
The genus Cryptococcus includes yeast fungi that develop a specific biotrophic association with different host plants and comprises more than 80 species, with two being pathogenic to humans. Previously known as a single species, Cryptococcus neoformans was divided into two varieties, C. neoformans variety gattii and C. neoformans variety neoformans [1,2]; nowadays, these two varieties are classified as two different anamorphic species, C. neoformans species complex and C. gattii species complex [2,3,4,5]. C. neoformans complex has several varieties (C. neoformans variety grubii serotype A, with three genotypes VNI, VNB, VNII; and C. neoformans var. neoformans serotype D with genotype VNIV; the serotype hybrid AD is associated with the VNIII genotype), while the complex of the species of C. gattii is divided into four genotypes (VGI, VGII, VGIII, and VGIV) [6,7,8,9]. VGII has been subclassified into three associated genotypes (VGIIa, VGIIb, and VGIIc) [10,11]. Nevertheless, it has been suggested that the actual classifications of C. neoformans species (known as C. neoformans variety grubii serotype A with VNB genotype, and VNI / VNII genotypes), C. deneoformans (referred to as C. neoformans var. neoformans with serotype D y genotype VNIV), and a hybrid classification composed of C. neoformans and C. deneoformans (with serotype AD and genotype VNIII) be recognized. The complex of the species of C. gattii has been reorganized as five species: C. gattii with genotype VGI, C. deuterogattii with genotype VGII, C. bacillisporus with genotype VGIII, C. tetragattii with genotype VGIV, and finally C. decagattii with genotypes VGIV and VGIIIc, in addition to hybrids such as C. deneoformans with C. gattii, the hybrid C. neoformans with C. gattii, and the hybrid C. neoformans with C. deuterogattii [6,7]. These species can cause pulmonary effects and damage to the central nervous system (CNS) [8], with being C. neoformans the worldwide cause of cryptococcal meningoencephalitis [9]. The C. neoformans complex affects immunocompromised individuals, mainly those affected by the human immunodeficiency virus (HIV) [12,13,14,15]. In contrast, the C. gattii complex is associated with infections in apparently immunocompetent patients, indicating that even people with a healthy immune system can be affected by this pathogen [16,17]. It should be noted that Cryptococcus spp. is capable of remaining latent without affecting the host, which makes it highly adaptive [9].
The C. neoformans complex has a wide distribution in different departments of Colombia, such as Cauca [18], Córdoba [19], Cundinamarca [20,21], Huila [22], Nariño [23] Norte de Santander [24,25,26,27], and Valle del Cauca [28,29]. In the same way, other regions of the world, such as Malawi [30], Nigeria [31,32], China [33], South Africa [34], Brazil [35,36], United States [37], Italy [38], Guinea [39], Argentina [40,41], and Ecuador [42], among others [25,43,44,45,46,47], have reported this species in the environment. This complex is generally associated with birds’ excreta, especially from pigeons (C. livia) [48], saprophytic sources of the C. neoformans complex and the soil contaminated with it [49,50,51,52]. The high contents of nitrogen, creatinine, and salts present in these wastes generate an appropriate environment for the growth of the fungus [53,54]. The pigeon feces, having low moisture content and low exposure to sunlight, are reservoirs for the C. neoformans complex [23]. Additionally, the C. neoformans complex has been reported in tree species, with higher prevalence compared to the C. gattii complex, for example in Eucalyptus spp. [55,56], Terminalia catappa [22], Olea europea [57], and Ceratonia (carob tree) [55].
Conversely, the C. gattii complex has been described in temperate regions such as Canada, California, and Oregon [58]. Thus, in Colombia, this species has been identified in the regions of Norte de Santander [25,26], Cundinamarca [59,60], and Córdoba [19], and across the globe, including Australia [61], Africa [34], India [45,62], Italy [55], the United States [63] and southern California [64], Canada [58], Spain [65], and China [33], among other regions [45,66,67,68,69,70,71,72,73].
The habitats of C. gattii complex are associated with tree species, with reported isolation from flowers, bark, and leaves [46,74,75]. Despite this, this complex has been found in the environment to a lesser degree than the C. neoformans complex, especially when associated with the genus Eucalyptus [55]. However, at present, it has also been associated with flowers, hollows, and decomposing wood of different trees, including Acacia, Ficus, and Terminalia catappa, in several regions of the world [24,59,63,67]. In some cases, the C. gattii complex has also been isolated from other environments, such as soil, air, and water [10,76,77]. Research suggests that periods of higher humidity in temperate climates are more favorable for the proliferation of the C. gattii complex; however, different studies on the environmental conditions have reported that the favorable characteristics for the development of this yeast depend on the climatic conditions of the area [60,73,78,79].
Studies published from 2006 to 2016 in different regions of Colombia reported cases of cryptococcosis. The prevalent risk factor reported was HIV infection in 83.5% sufferers and an increase in cases in the female population compared to the previous period of 1997–2005 [78]. Thus, the average annual incidence in the general population is 2.4 per one million inhabitants, while in AIDS patients there is an increase of 1.1 people per 1000 [80]. These data reflect the importance of knowing the distribution of these pathogenic species in the environmental isolates to identify possible sources in which infectious propagules may grow.
The objective of this systematic review is to describe the distribution of environmental isolates of the C. neoformans species complex and C. gattii species complex in Colombia and their associations with environmental conditions.
2. Materials and Methods
A systematic search was carried out in eight online databases: BioMed, DialNet, DOAJ, Medline, PubMed, ScienceDirect, Redalyc, and SciELO, following the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines, which include identification, selection, and inclusion of the reviewed literature. The years of publication were 1990 to 2020. The descriptors used were: Cryptococcus spp., environment, meteorological conditions, and ecology. The articles were selected considering the following inclusion criteria: studies of environmental isolates of Cryptococcus neoformans and Cryptococcus gattii in Colombia, mainly articles in Spanish and English. Studies of clinical isolates reported in Colombia were excluded. The titles and abstracts of 103 articles were evaluated for relevance; after removing duplicate articles and study reviews, 53 articles underwent full-text review, taking into account environmental variables, microbiological identification methods, and molecular characterization, which were included in the total synthesis of the document.
3. Results
Colombia has different thermal floors, producing multiple climatic conditions throughout the national territory. The natural habitat of the C. neoformans species complex and C. gattii species complex is associated with different environments, from cold and temperate climates to tropical and subtropical climates. In this way, the studies carried out in Colombia aimed to identify the distribution of the infectious agent of cryptococcosis, and thus identify the ecological niche of this fungus.
Of the included studies, five reports showed associations between isolates of Cryptococcus spp. and environmental variables. Indeed, Granados et al. and Anacona et al. reported a correlation between high and mean temperature values and recovered isolates of C. neoformans [18,81]. Conversely, C. gattii serotype C had fewer isolates due to extreme minimum and maximum temperatures according to Granados et al. [81]. Furthermore, a negative correlation with relative humidity was shown for C. neoformans serotype A and C. gattii serotype C [23,81] in comparison with the favorable influence of humidity in recovered C. neoformans, as reported by Anacon 2018 et al. [18]. Additionally, C. gattii serotype B exhibited a positive correlation with relative humidity, associated with the high recovery under low evaporation conditions [81].
For the climatic variable precipitation, the C. neoformans serotype A evidenced a strong correlation [81]. For solar radiation, Granados et al. and Vallejo et al. reported that C. neoformans serotype A and C. gattii serotype C exhibited a positive correlation with high solar radiation values, in contrast to C. gattii serotype B, which showed a positive correlation with low values of solar radiation [23,81].
Regarding the sample source, Caicedo and Vallejo reported that the accumulation of excreta and the high presence of pigeons result in increases in the recovery of the C. neoformans fungus [23,28].
Finally, Caicedo reported that wooden boxes and flat ceilings were the nesting areas where there were more significant isolates of C. neoformans [28].
The studies carried out in Colombia confirm the presence of pathogenic species of the genus Cryptococcus in the region, with the C. gattii species complex being reported in tropical to temperate climates, demonstrating that this species is highly adaptable to different ecological conditions in the country [25,60]. Other isolates of C. neoformans have been reported in temperate and tropical climates, associated mainly with avian excreta [18,22,23,56,82]. Environmental isolates of these pathogenic species have been reported in different cities in Colombia, such as Popayán [18], Pasto [23], Cúcuta [26,27], Monteria [19], Bogotá [18,19], and municipalities of Cundinamarca [21,29,61], with Bogotá being the city with the highest number of positive samples [20,81]. These results are discussed throughout the article and are synthesized in Figure 1 and Table 1, where the record of environmental isolates of C. neoformans and C. gattii in different parts of Colombia are shown.
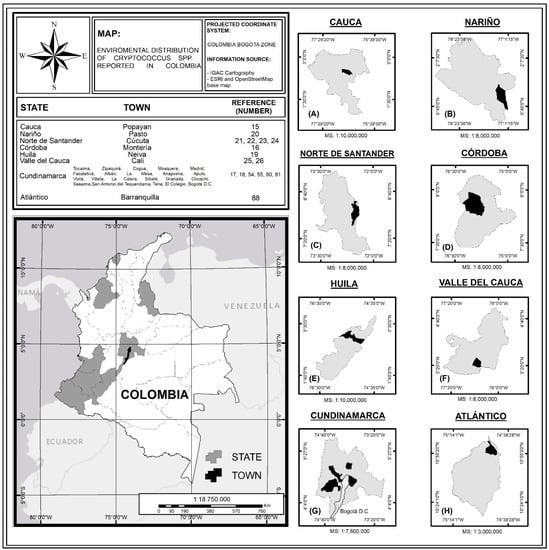
Figure 1.
Distribution of environmental isolations of Cryptococcus spp. in Colombia; MS: Map Scale. (A) Department of Cauca, (B) Department of Nariño, (C) Department of Norte de Santander, (D) Department of Córdoba, (E) Department of Huila, (F) Department of Valle del Cauca, (G) Department of Cundinamarca, and (H) Department of Atlántico. Source: The authors of this study, based on the information recorded in Table 1.

Table 1.
Environmental isolates of the Cryptococcus genus registered in Colombia.
3.1. Atlántico
The city of Barranquilla is the capital of the department of Atlántico, in which a warm climate predominates, with an annual temperature of 28 °C; this region has bimodal rainy seasons that go from May to June and from September to November, with average annual rainfall ranging between 500 and 1500 mm, with dry seasons from December to March and from June to July. Between 2012 and 2014, Noguera et al. conducted a clinical and environmental study, whereby 32 sampling points were recorded, consisting of local parks and areas with a high density of trees. From 1068 environmental samples, 0.4% were positive for C. neoformans molecular type VNI in almond trees and roses. The researchers concluded that C. neoformans, a molecular type VNI, had a higher prevalence than C. gattii and was associated with human exposure in this geographic region [83].
3.2. Bogotá
In Bogotá in 1994, Duarte et al. evaluated the association of Cryptococcus species with the eucalyptus trees E. camaldulensis and E. terericornis, distributed in 13 areas located in the northeast, east, west, and center of Bogotá. This city has an altitude of 2630 m above sea level (masl), while at the sampling time the city had a humidity of 65.5% and average temperature of 16.4 °C; the location is shown in Figure 1F. It should be noted that the study did not report the molecular characterization of the isolates. From the results of the 13 evaluated areas, 572 isolates were recovered, of which 27 (4.72%) were of the Cryptococcus genus, distributed as follows; 8 were C. laurentii; 5 were C. macerans; 4 were C. ater and 4 were C. uniguttulatus; 2 were C. neoformans; and 1 each was C. hungaricus, C. albidus, C. kuetzingii, and C. heveanesis. For the remaining 500 samples, the authors did not report any identification. An important finding reported in this study was the establishment of the leaves as the main sources of isolates for Cryptococcus [21].
Another environmental report by Castañeda and Castañeda in 2001 showed the association between isolates of Cryptococcus species and Eucalyptus trees in a park located in the northeast of Bogotá. No environmental data were recorded during the investigation; however, according to the climatic parameters reported by IDEAM in this period, Bogotá had an average temperature of 13.4 °C and an annual rainfall of 453.91 mm [84,85]. The study did not include a molecular characterization of the isolates. C. laurentii was the mainly isolated species, with 17 positive samples, followed by C. neoformans var. neoformans with two positive isolates, and finally one C. albidus isolate. Contrary to what is reported in the literature, where a close association between C. gattii and E. camaldulensis is described, in this study no isolates of this species were obtained [20].
The study published by Granados and Castañeda showed the results of sampling carried out in 2003 during February to May in 3 areas of Bogotá: The José Celestino Mutis Botanical Garden (JB) (4°41′ N, 74°06′ W), El Lago metropolitan park (PEL) (4°40′ N, 74°03′ W), and the campus of Universidad Nacional de Colombia (UN) (4°38′ N, 74°06′ W). Climatological data for this period showed an average long-term temperature of 19.1 °C, with precipitation being between 500 and 1000 mm [85,86]. In total, 480 samples associated with the trees (form the bark, the soil around the trees, and debris) and 89 samples of bird droppings were collected. The authors did not report molecular characterization for the isolates. Of the 480 samples obtained from the trees, 38 (7.9%) were C. neoformans.
It should be noted that in those trees that were inhabited by birds, the frequency of isolation was lower (21.87%) compared to samples isolated from the trees or in decomposing wood (48.75%), which were not frequented by birds. For the statistical analysis, ANOVA, MANOVA, canonical discriminant analysis, Pearson’s correlation, and Chi squared were performed, using the Statistical Analysis System (SAS) for Windows v. 8.02. The frequency of isolation in the excreta samples was 6.7% [87] for C. neoformans var. gattii and 1% (5) for C. neoformans var. grubii, which were significantly higher in the dry excrement than the humid excreta. Based on the results, exposure to sunlight does not represent a decisive factor influencing the recovery of yeast. In this study, the fungus showed more frequent colonization in the debris found around trees, supporting the concept that the main niche appears to be the result of the biodegradation of wood. In contrast, the rainy months (April and May), characterized by high rainfall, humidity, few hours of sunlight, and less extreme temperatures (an average temperature of 5 °C higher), favored the appearance and spread of C. neoformans in the environment more than the dry months. Finally, this study shows how C. neoformans can colonize several zones in a city, and how its population densities can vary within the zones; this distribution could be explained by the capacity of birds to disperse propagules in the wind [81].
In 2010, Escandón et al. collected samples over six months, between February and August 2007. The authors did not report environmental data during the investigation; however, according to the climatic parameters reported by IDEAM in this period, Bogotá had an average temperature of 19.124 °C and precipitation range of 500–1000 mm [86,88]. In total, 28 bark, 37 soil, 52 debris, 4 seed, and 7 flower samples were taken from a total of 91 trees. Isolates were typed using RFLP of the URA5 gene, while PCR was used for the determination of mating types. Of the 128 samples collected, 15 (11.7%) isolates were identified as C. gattii molecular type VGIII, mating type a; of these isolates, three were from the extract of the inflorescence of red flowers, while the rest were from the detritus of Corymbia ficifolia. Of the 113 remaining samples, the researchers did not specify any identification. The present study reports the isolation of C. gattii in C. ficifolia trees in Colombia, as represented in Figure 2. This is the first report to reveal this tree species as a habitat for the fungal species [59].
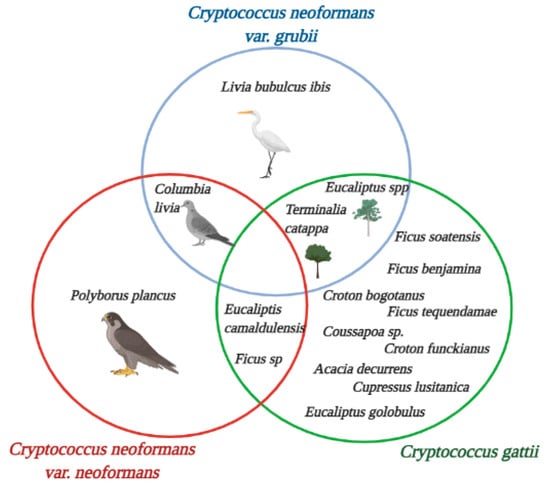
Figure 2.
Environmental isolate sources of C. neoformans and C. gattii species complexes in Colombia. Source: The authors of this study; image created in BioRender.com.
3.3. Cali
In 1996, 20 communes of Santiago de Cali were selected as sampling zones for the presence of nesting sites. Santiago de Cali is the capital city of Valle del Cauca department (Figure 1G), which has an average temperature of 28.55 °C and a precipitation range of 1000–1500 mm according to the reports by IDEAM [86,88]. In total, 119 samples of pigeon droppings (C. livia) from ceilings, roofs, and wooden boxes were recovered. All samples collected on roofs were negative, while all those obtained from ceilings were positive for C. neoformans. Molecular characterization was not performed. The statistical analysis consisted of descriptive statistics and the Chi square test, however the program used was not indicated. In total, 47 % of the samples were identified as C. neoformans. Researchers associated high exposure to ultraviolet rays with the absence of C. neoformans in roof samples because the sunlight exposure limited the growth of yeast. They also found that where there was a higher number of pigeons, the recovery rate of the fungus was higher (Figure 2). Finally, given the significant increase in cryptococcosis in AIDS patients, the authors noted the potential risk for these patients of acquiring cryptococcosis in urban environments in the city of Cali [28].
In addition, Caicedo et al. in 1996 published a study carried out at the Cali Zoo between 1994 and 1995. The researchers did not report environmental data during the sampling; however, according to the climatic parameters reported by IDEAM in this period, the city of Cali presented an average temperature of 28.55 °C, with precipitation ranging between 1000 and 1500 mm [86,88]. In total, 380 samples were recovered; 110 from bird droppings, 148 from the air inside the cages, and 122 from the air outside the cages. The study reported two isolates of the species C. neoformans var. neoformans, one (0.9%) from excreta of caracara (Polyborus plancus) and the other from a petri dish exposed in the area where the same species of falconiform bird lived (Figure 2); the latter corresponded to 0.3% of the air samples, while the other air sample results were negative. The results obtained in the zoo in the city of Cali showed a low frequency of isolates of this yeast, which was associated with the fact that the cleaning and disinfection procedures that were implemented were adequate [29].
3.4. Cúcuta
In 1998, Callejas et al. published a study that aimed to search the C. gattii habitat in the city of Cúcuta. The municipal area covers approximately 1176 km2. The altitude is 320 masl, the average temperature is 27 °C, and the average annual rainfall is 763 mm. This city is located in the northern region of Santander, as shown in Figure 1C. A total of 157 samples collected from May to September 1997 from 68 trees of T. catappa, 90 debris samples, 54 samples from other types of plant materials (38 from seeds, seven from bark, eight from leaves, and one from flowers), and 13 air samples were also collected. The authors did not report on the molecular characterization. The researchers were able to isolate four positive samples for C. gattii serotype C (2.54%), corresponding to plant debris; they did not report on the identification for the remaining 153 samples. The researchers concluded that there may be an association between plant dendrites and the positive isolates of C. gattii, this being a type of habitat in which yeast is found [25], as shown in Figure 2.
In 2001, Castañeda et al. published a study that sought to determine the association of the presence of C. gattii (at that time classified as C. neoformans var. gattii) with the species T. catappa (almond trees). The authors reported on a total of 370 samples, of which 160 were from almond debris and 210 were from 9 new almond trees, selected from January 1998 to July 1999. The study reported that 31 samples (19.3%) were positive for C. gattii serotype C (formerly C. neoformans var. gattii) of the 160 samples initially collected in the study, while of the 210 samples collected from the nine additional almond trees, only 1 (0.48%) was positive for C. gattii. The authors established that it was not possible to isolate the C. neoformans species complex from any of the almond trees, thus suggesting that there is a specific association of C. gattii with this tree species [24]. Another environmental report was made in 2011 in Cúcuta by Firacative et al. This area has an average temperature of 28 °C and an approximate average annual rainfall of 763 mm. In total, 4389 samples were processed, among which samples of soil, bark, debris, leaves, and fruits were collected, which were associated with 3634 trees from different species, including T. catappa (almond), Ficus sp. (Higuerón), Licania tomentosa (Oiti), Pithecellobium dulce (Chiminango), Fagara rhoifolia (Tachuelo), Melicoccus bijugatus (mamón), Enterolobium cyclocarpum (piñon), and Mangifera indica (mango) (Figure 2). Molecular typing of isolates was performed by PCR fingerprinting and RFLP of the URA5 gene. From 4389 samples, only six were positive—three (0.07%) for C. gattii and three (0.07%) for C. neoformans. From C. gattii-positive samples, seven isolates were C serotype, VGIII molecular type; and one was serotype B, VGI molecular type. Of the positive samples for C. neoformans, 13 isolates were classified as a C. neoformans var. grubii, serotype A, VNI molecular type. All recovered isolates were from the soil samples. Contrary to expectations for the study area, which has a high rate of cryptococcosis due to C. gattii, there was a low rate of environmental recovery for this species, suggesting the importance of establishing environmental variables that directly affect the survival of microorganisms in the environment, meaning samplings should be taken at specific times of the year [26].
In 2019, Angarita et al. carried out a study in the city of Cúcuta to identify and characterize Cryptococcus environmental isolates in ten public areas. The study did not report environmental data for the months of sample collection; however, as reported by IDEAM for this period, the average temperature was 24.95 °C, with annual rainfall ranging between 50 and 74 mm [85]. A total of 1300 samples from 446 trees belonging to 10 different species were found, including soil (442), bark (434), nuts (40), and leaves (384). PCR fingerprinting and RFLP of the URA5 gene were used to determine the molecular pattern. The results obtained in this study established the presence of C. neoformans in 19 trees (4.3%) and C. gattii in one tree (0.2%), for a total of 21 isolates and 20 positive trees. Santander Park, with 47.6% (10 out of 21) of the isolates, was the area that presented the most significant number of positive samples, followed by La Victoria Park with 23.8% (5 of 21 isolates), all of which were identified as C. neoformans. Using a different method, C. gattii was isolated from the Ficus benjamina tree located in the Mercedes Ábrego park (Figure 2). Genotypic analysis revealed the presence of C. neoformans var. grubii VNI molecular type in 85.7% of the environmental isolates, followed by VNII in 9.5% and C. gattii VGII molecular type in 4.8% of the isolates. This study revealed the importance of performing longitudinal sampling of the environmental niches reported in the literature for this fungus, thus allowing its presence to be established in the sampled areas, suggesting this type of sampling as a tool for permanent surveillance of this pathogenic fungus in endemic areas of the city of Cúcuta [27].
3.5. Cundinamarca
Cundinamarca department is in the central region of the country, located between 3°42′ and 5°51′ latitude north and 73°03′ and 74°54′ longitude west of the Greenwich meridian. The altitude is between 2000 and 3000 masl and the temperature ranges between 12 and 18 °C, with a predominant cold thermal floor. The Páramo regions have temperatures equal or less than 12 °C and are located at an elevation above 3000 masl. Quintero et al. carried out a study in different areas of Cundinamarca in order to evaluate under which climatic conditions the Cryptococcus species are best adapted, taking into account the topographic, natural, and climatological diversity of the department. Of the 765 samples processed, 437 corresponded to eucalyptus debris, 182 to almond debris, and 146 to pigeon droppings. The results indicated that C. neoformans was present in 12 (46%) of the municipalities studied, indicating that 104 (14%) samples were positive, whereby the pigeon droppings (21%) were the principal substrate. A total of 92 isolates (88%) were recovered from the cold thermal floor. In the Páramo area, the yeast was not isolated due to the environmental conditions, such as the precipitation and high relative humidity, few hours of sunlight, and temperate thermal floor between 18 and 24 °C, which favor the propagation of this yeast in the environment, as well as the high altitudes of between 1000 and 2000 masl. In conclusion, the authors confirmed the association between C. neoformans var. grubii serotype A and pigeon feces, in addition to confirming eucalyptus as an environmental niche for C. gattii serotypes B and C, while almonds were also established as possible hosts for serotype C (Figure 2). Finally, the importance of the cold thermal floor as a habitat for the fungus was established [56].
However, a study carried out by Escandón P. in the municipality of La Calera (Cundinamarca) from February to March of 2003 evaluated the presence and distribution of species of the C. neoformans complex, especially serotype B, which were associated with the study area. This region has two thermal floors, namely cold and temperate, in addition to having an altitude of 2728 masl, with an average temperature of 12.7 °C and annual precipitation of 250 mm. The researchers recovered 195 samples from trees debris, 167 from Eucalyptus spp., and 28 from other species, such as pines (Pinus sp.), white snakeroot tree (Ageratina altissima), Laurel (Laurus nobilis), Raque (Vallea stipularis), and Guaba (Phytolacca bogotensis). Of the 167 samples taken with a swab, C. gattii was found in February in 3 out of 40 samples (7.5%), on March 6 32 out of 40 samples (80%), and on March 12 in 11 out of 87 samples (12, 5%). Additionally, C. gattii serotype B was recovered from 46 of the 167 samples of Eucalyptus (27.5%). The presence of yeasts with small capsules was confirmed in 30 (65.2%) of these, while the remaining 16 were strains without capsules. This study was the first report on C. gattii serotype B. It should be noted that the fungus was found on different trees species, as observed in Figure 2 [60,89].
3.6. Huila
In 2018, Virviescas B. et al. performed a study in the city of Neiva to determine the distribution of the etiological agents of cryptococcosis in the region. The municipality has an average annual temperature of 27.7 °C, is located at a latitude of 2°59′55” north and a longitude of 75°18′16” west, and has an altitude of 442 masl, with an approximate population of 345,911 inhabitants. In total, 118 samples were collected—32 from the fruits, leaves, and debris of almond trees (T. catappa), and 86 from pigeon droppings (C. livia). The samples were processed using conventional techniques. Species were determined by seeding on Canavanine-Glycine-Bromothymol Blue (CGB) agar, and molecular typing was performed using PCR fingerprinting and RFLP of the URA5 gene. Of the total samples processed, 40 colonies obtained from eight (6.8%) samples were identified as C. neoformans var. grubii and isolated from bird droppings, which can be observed in Figure 2, contrary to tree samples, where no positive samples were recovered. Likewise, C. gattii was not isolated from the collected samples [22].
3.7. Monteria
Between 2008 and 2009, an investigation was carried out by Contreras et al. in Monteria, the capital of the department of Córdoba (Figure 1D), located in northern region of Colombia, to determine the ecological association of C. gattii with almond trees (T. catappa) present in the urban area of the city. Monteria has an altitude of 20 masl, a warm climate with temperatures ranging between 28 and 35 °C, a relative humidity of 85%, and pluvial precipitation of 1200 to 2500 mm annually. The researchers collected 2445 samples from 163 T. catappa trees. The samples were collected, taking into account the parts of the bark with grooves and hollows. The study did not include molecular characterization. The statistical analyses included non-parametric analysis, Tukey’s hypothesis test, and Chi-squared tests using Statgraphics Centurion version 15.2.
Of the total samples processed, C. gattii was isolated from 217 samples, which corresponded to 8.9%. The authors highlighted that the sample collection corresponded to a rainy period involving more humidity and a higher accumulation of debris. Given the obtained results, the researchers consider almond trees an ecological niche for C. gattii (mainly the fruits, which are essential), since in the region the ripe fruit is edible and could indicate a point of spread for the fungus. These findings were the first report of the isolation of C. gattii from environmental sources in the department of Córdoba [19].
3.8. Pasto
For the municipality of Pasto, Vallejo et al. in 2016 reported a study that carried out an analysis of the residential and populated areas with a high density of pigeons. The study did not report environmental data; however, according to the climatic parameters reported by the IDEAM, Pasto has an altitude of 2527 masl, an average temperature of 19.2 °C, and a maximum average annual rainfall of 106,267 mm [85]. The city is located on the Andes cordillera, as shown in Figure 1B. In total, 128 samples were recovered from the different areas; most of the samples were from feces without debris, some contained residues of soil and vegetables, while others contained feathers. The samples were exposed continuously to light and variably to the environment. Sampling areas were characterized and classified as high, medium, and low, according to the environmental conditions of each area and the characteristics of the samples.
Contrary to the procedure reported by Caicedo et al. [28], samples were exposed to long hours of sunlight, which is an excellent condition for obtaining positive samples, as it is different to the low humidity in the samples, which affects the recovery of the fungus. The study also showed that a higher number of pigeons per nest favored a higher percentage of isolates of the fungus being obtained [23]. The study did not perform molecular characterization of the obtained isolates. The fungus was isolated in 9 out of 10 sampled areas, whereby 26.5% of the total samples collected corresponded to positive isolates of C. neoformans. The number of isolates was lower in fresh samples, while wet samples (not necessarily fresh) provided a higher number of positive isolates, similar to the results reported by Granados and Castañeda in 2006 [90]. Descriptive statistics and the Chi-squared test were used for statistical analysis using SPSS Statistics 20.0 software under shareware license. This study established that of the pigeon droppings samples taken in urban areas, the prevalence of C. neoformans was 90%, demonstrating an association of the avian species with Cryptococcus isolates (Figure 2). Additionally, the importance of establishing the variables was confirmed, such as the exposure to sun or shade, and of checking whether the avian excreta were fresh, wet, or contaminated with plant residues, which could be associated with an increased probability to obtain isolations positive for this yeast [23].
3.9. Popayán
Popayán is the capital of the Colombian department of Cauca, with an elevation of 1735 masl, a subtropical highland climate with an annual average temperature of 18–19 °C, with maximum temperatures occurring from July to September (high temperature of 29 °C and minimum temperature of 10 °C) and a precipitation of 1941 mm. The city is located on the west of Colombia, as shown in Figure 1A in the distribution map. A study reporting environmental isolates was published in 2018 by Anacona et al.; they collected a total of 303 samples of bird droppings from pigeons (C. livia) and herons (Bubulcus ibis) between September 2012 and June 2013. These associations can be observed in Figure 2.
The authors collected and processed the samples using conventional techniques. The identification of presumptive isolates of the C. neoformans species complex was also carried out by means of conventional procedures, along with the molecular characterization of the isolates. One-way ANOVA, post hoc analysis, Tukey’s test, and the Chi-squared test were performed using SPSS version 15 for statistical analysis. A total of 118 (38.94%) samples were positive for Cryptococcus neoformans var. grubii, with 99.2% corresponding to the VNI molecular pattern and the remaining 0.8% to VNII. Contrary to the report for the pigeon excreta samples, no species of the C. neoformans species complex was isolated from B. ibis. In this way, the study allows us to confirm that bird droppings, especially of C. livia, are a fundamental environmental niche for C. neoformans var. grubii molecular type VNI, since they allow growth and reproduction of the fungus, representing a potential source of infection for the susceptible population in Popayán [18], as shown in Figure 2.
4. Discussion
The Cryptococcus genus comprises a group of fungal pathogens that frequently affects humans; without timely and adequate treatment, it can be fatal. In order to establish control measures for infectious diseases, it is highly relevant to provide a report on the fungal environmental distribution throughout a country.
According to the environmental studies reported in Colombia until 2019, climatic conditions are essential when analyzing the requirements that favor the appearance of the fungus (Supplementary Table S1). The studies report an extensive distribution of the fungus at different altitudes ranging from 20 masl (e.g., Monteria city [19]) up to 3000 masl (e.g., Cundinamarca department) [54]. The fungus can survive in temperatures as low as 12 °C, as demonstrated by Quintero [56], or in high temperatures reaching 28.8 °C, as reported by Cúcuta [24], thus demonstrating its wide distribution range in the country, as shown in Figure 3.
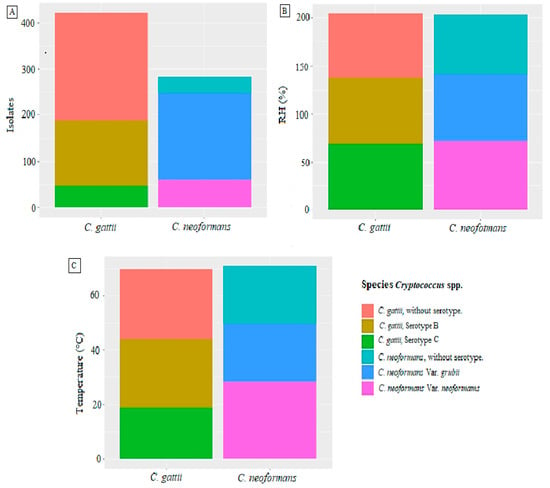
Figure 3.
Environmental conditions reported for each species of Cryptococcus spp. (A) Numbers of isolates reported for each species (C. gattii and C. neoformans), according to their respective varieties. (B) Percentages of relative humidity (RH) reported for the isolates of each species (C. gattii and C. neoformans), according to their respective varieties. (C) Temperatures (°C) reported for the isolates of each species (C. gattii and C. neoformans), according to their respective varieties. Source: The authors of this study; image created in RStudio.
Regarding the numbers of samples collected for the isolation of Cryptococcus spp., it can be said that the most significant number of samples collected was in the city of Cúcuta at 4338 samples, of which only 0.14% were positive, which may be due to the temperatures at this site (27.8 °C to 28.8 °C) [24]. This low frequency of positive isolates is in contrast with the results found in the city of Popayán, where the numbers of samples collected was 303, 38.94% of which were positive. This city has a temperature range of between 10 and 29 °C [18]. In the same way, this region was established by Mak et al. as an optimal ecological niche for isolation of C. neoformans species [89]. Similarly, studies carried out in the department of Cundinamarca resulted in 13.59% of positive samples for Cryptococcus from the 765 samples collected under temperature conditions ranging between 12 and 18 °C [56]. The frequency of positive Cryptococcus isolates is higher in environments with low temperatures, since such conditions promote their conservation [91].
The methods used for the identification and characterization of the Cryptococcus species complex have mainly involved extraction with saline phosphate buffer and identification using G. abyssinica seed agar. In some studies, Indian ink staining was used to identify the fungus capsule, being a simple and inexpensive method. The molecular methods generally used in the studies were restriction fragment length polymorphism (RFLP) of the URA5 gene and PCR fingerprinting with primer (GTG)5.
Data analysis was not performed in most of the studies, except when reporting the percentages of positive samples found for each species according to the nomenclature used by each study. However, some studies have reported statistical data analysis, including Granados et al. in Bogotá [81], Caicedo et al. in Cali [28], Quintero et al. in Cundinamarca [56], Contreras et al. [19], Vallejo et al. in Pasto [23], and Anacona et al. in Popayán [18]. According to the results of these statistical analyses, the climatic variables that influence the recovery of the fungus are precipitation, temperature, evaporation, solar radiation, and humidity (Supplementary Table S1).
The studies carried out in Colombia, as with the reports worldwide, demonstrate the association of the C. neoformans complex with excreta of C. livia pigeons and of some C. neoformans species with P. plancus, thus affirming the association of the fungus with different avian species. Bird droppings contain xanthine, urea, uric acid, and creatinine, substrates that the fungus can assimilate within [11,31], given the presence of enzymes such as creatinine that favor the adaptation of yeast to this type of environment [92]. This proves that bird droppings are a secondary and temporary niche for this species [55].
In addition, the probability of finding Cryptococcus is higher if the samples are wet, as shown by Quintero et al. In Cundinamarca region, 88% of the isolates were recovered from the cold thermal floor, with high precipitation and relatively humid environmental conditions, which favored the occurrence and spread of the fungus in the environment, as reported by Cogliati et al., who found a higher occurrence of the fungus in rainy seasons or wet periods [53,93]. Similar to the Cundinamarca study, a study carried out in Australia established that yeast could be isolated from eucalyptus debris and pigeon droppings, however the study had a particular focus on cold seasons with high relative humidity [54,94]. Contreras and collaborators in 2011 reported similar data, where they highlighted that the highest percentage of positive samples was because the collection was performed in rainy seasons, which promoted more significant accumulation of debris [19]. This was in line with the study by Vallejo et al., who established that wet samples contain higher numbers of isolates of Cryptococcus species.
Vallejo et al. in 2016 obtained positive samples for the Cryptococcus species complex in places exposed to constant sunlight, thus relating the amount of brightness to the presence of the fungus [22], since the ability to produce melanin not only helps the fungus resist solar radiation, but it can also use it as metabolic energy [23,95]. This aspect differed significantly from the rest of the studies, which mentioned that the solar brightness received by the fungus is a factor that prevents its growth. For example, in the study carried out in Cali, the authors did not collect positive samples from places with high sun exposure [29].
According to the samples registered for the different studies, the presence of the C. neoformans complex is generally associated with debris from trees of the genus Eucalyptus and Ficus sp., as stated by Lazéra et al. [96] and Cogliati et al., who found associations with arboreal specimens as niches and stable reservoirs [53,93]. Similar results were also found in the regions of Popayán, Cundinamarca, Bogotá, and Cali (Figure 4). Likewise, this species is strongly associated with avian droppings, generally from pigeons (C. livia) [87], which is consistent with the findings described Kwon-Chung and Bennett in 1984 [47], who established the primary environmental source of C. neoformans as being pigeon droppings, which was contrary to the findings reported by Cogliati et al., who that suggested that poultry feces is a secondary and transitory niche [53,93].
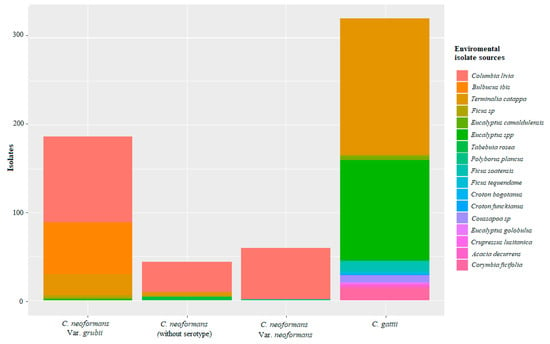
Figure 4.
Isolations reported according to their environmental source for the complexes of species of C. neoformans and C. gattii in Colombia. Source: The authors of this study; image created in RStudio.
C. gattii has been found in different regions, such as Cúcuta and Monteria, which have average temperatures of around 27 °C, as well as Cundinamarca, which has an average temperature of 13 °C. In agreement with the study by Cogliati et al., where identified that warm temperatures favors the distribution of C. gattii complex [53,93]. The trees species associated with this yeast are T. catappa and Ficus sp., as reported by Castañeda et al. [24] and corroborated by the study by Escandon et al., in which the survival of the C. gattii complex was shown for up to 12 months in these trees species [97]. Additionally, Firacative et al. recorded positive isolates from the soil close to the tree species Ficus sp. [26]. In the city of Monteria, Contreras et al. reported C. gattii in soil samples from Ficus spp. [19]. This association between the C. gattii complex and rubber and almond species has been reported in different parts of the world. For example, the study by Randhawa et al. in 2003 reported decomposed wood in Ficus religiosa trees as an environmental source of C. gattii [73]. These findings indicate that these tree species could be a natural habitat for C. gattii complex [94], describing them as reservoirs for this yeast species [65,97,98]. The C. gattii complex is difficult to isolate from the environment [99,100]. However, the findings from a few studies showed high numbers of positive samples for this species. For the C. neoformans complex, the number of positive isolates reported in each study was low, however the number of studies is much higher [19,26,27], as can be seen in Figure 4.
Both the C. neoformans complex and the C. gattii complex have been reported in clinical isolates; however, the C. neoformans complex is mainly isolated from the environment, indicating the presence of the two species in the environment [101]. Thus, by expanding the search for the C. gattii complex in the environment, the number of studies has increased, with the largest numbers of isolated samples being obtained for the species of C. gattii, especially serotype C in Cúcuta, which has a high incidence of C. gattii serotype B in immunocompetent patients [19,24,25,26,27]. In contrast, in La Calera, Cundinamarca, species of C. gattii with serotype B have been isolated in low proportions [60]. The two serotypes are considered primary pathogens, which therefore infect immunocompetent individuals, showing the need to establish the foci of infection for this species [102].
Regarding genotypes, C. gattii VGII is the most common in clinical isolates, while the VGIII genotype is less frequent [102]. However, in the study by Escandon et al., the most common phenotype determined in environmental isolates was VGIII, in agreement with the results reported by Firacative et al. in 2011, although to a lesser extent [26,57]. Additionally, the predominant genotype for clinical isolates of C. neoformans is VNI [102,103], which coincides with the genotype determined in environmental samples [18,26,27].
Finally, it should be mentioned that as in other regions, in Colombia the soil samples represented a poor environment for the recovery of the C. neoformans species, while the excreta of birds was considered an important reservoir for this species [73,99,100].
5. Conclusions
The studies carried out in Colombia have shown that the C. neoformans species complex and C. gattii species complex share some niches in terms of where they are established, being distributed in regions ranging from 20 to 3000 masl. Similarly, positive isolates were found in cold and temperate climates, with low sunlight and high relative humidity, thus showing that in wet samples, the probability of finding species of the genus Cryptococcus is higher. In contrast, although to a lesser extent, it has been established that in some cases these yeast species can grow in areas with a significant influence of sunlight, which may be related to the production of melanin as a protective factor against UV rays, giving this species adaptability to these environments [89] In this way, it is essential to investigate the production of this virulence factor in the isolates that recover under conditions of a strong influence of solar brightness in future studies.
Furthermore, the methods used for isolation and identification of the Cryptococcus complex species, such as G. abyssinica agar, urease test, Indian ink staining, CGB agar, RFLP of the URA5 gene, and PCR fingerprinting with primer (GTG)5, correspond to the methods mostly used in Colombian studies. It is worth highlighting their effectiveness for the characterization of these two species.
Finally, the reviewed studies allowed us to establish that the most significant numbers of Cryptococcus spp. species were isolated in urban areas, such as squares, parks, churches, and zoos, which could be related to human activity, therefore increasing the possibility of infection through these sources.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/jof7060410/s1, Table S1: Studies that reported the environmental conditions which favor the development of the fungus.
Author Contributions
Study design, M.F.-C., Z.A.S.-Q.; development and methodology, B.-N.S.-E., D.G.-S., M.F.-C., Z.A.S.-Q.; collection of data, B.-N.S.-E., D.G.-S.; formal analysis, B.-N.S.-E., M.F.-C., P.E., Z.A.S.-Q.; writing—original draft preparation, B.-N.S.-E., M.F.-C., Z.A.S.-Q.; writing—review and editing, M.F.-C., P.E., Z.A.S.-Q. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Acknowledgments
The authors thank Universidad de Boyacá, Universidad Pedagógica y Tecnológica de Colombia (UPTC) and Instituto Nacional de Salud, and the anonymous reviewers who participated in the critical review of the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vanbreuseghem, R.; Takashio, M. An atypical strain of Cryptococcus neoformans (San Felice) Vuillemin 1894. Part II. Cryptococcus neoformans var. gattii. nov. Med. Trop. 1970, 50, 695–702. [Google Scholar]
- Kwon-Chung, K.J.; Boekhout, T.; Fell, J.W.; Diaz, M. (1557) Proposal to conserve the name Cryptococcus gattii against C. hondurianus and C. bacillisporus (Basidiomycota, Hymenomycetes, Tremellomycetidae). Taxon 2002, 51, 804–806. [Google Scholar] [CrossRef]
- Xue, C.; Tada, Y.; Dong, X.; Heitman, J. The Human Fungal Pathogen Cryptococcus Can Complete Its Sexual Cycle during a Pathogenic Association with Plants. Cell Host Microbe 2007, 1, 263–273. [Google Scholar] [CrossRef]
- Hibbett, D.S.; Binder, M.; Bischoff, J.F.; Blackwell, M.; Cannon, P.F.; Eriksson, O.E.; Huhndorf, S.; James, T.; Kirk, P.M.; Lücking, R.; et al. A higher-level phylogenetic classification of the Fungi. Mycol. Res. 2007, 111, 509–547. [Google Scholar] [CrossRef]
- Fell, J.W.; Boekhout, T.; Fonseca, A.; Scorzetti, G.; Statzell-Tallman, A. Biodiversity and systematics of basidiomycetous yeasts as determined by large-subunit rDNA D1/D2 domain sequence analysis. Int. J. Syst. Evol. Microbiol. 2000, 50, 1351–1371. [Google Scholar] [CrossRef]
- Kwon-Chung, K.J.; Bennett, J.E.; Wickes, B.L.; Meyer, W.; Cuomo, C.A.; Wollenburg, K.R.; Bicanic, T.A.; Castañeda, E.; Chang, Y.C.; Chen, J.; et al. The Case for Adopting the “Species Complex” Nomenclature for the Etiologic Agents of Cryptococcosis. mSphere 2017, 2, e00357-16. [Google Scholar] [CrossRef]
- Hagen, F.; Lumbsch, H.T.; Arsenijevic, V.A.; Badali, H.; Bertout, S.; Billmyre, R.B.; Bragulat, M.R.; Cabañes, F.J.; Carbia, M.; Chakrabarti, A.; et al. Importance of Resolving Fungal Nomenclature: The Case of Multiple Pathogenic Species in the Cryptococcus Genus. mSphere 2017, 2, 1–13. [Google Scholar] [CrossRef]
- Baker, M.R.D. The Primary Pulmonary Lymph Node Complex of Cryptococcosis. Am. J. Clin. Pathol. 1976, 65, 83–92. [Google Scholar] [CrossRef]
- May, R.C.; Stone, N.; Wiesner, D.L.; Bicanic, T.; Nielsen, K.V. Cryptococcus: From environmental saprophyte to global pathogen. Nat. Rev. Genet. 2016, 14, 106–117. [Google Scholar] [CrossRef]
- Iii, E.J.B.; Bildfell, R.J.; Frank, S.A.; Mitchell, T.G.; Marr, K.A.; Heitman, J. Molecular Evidence That the Range of the Vancouver Island Outbreak of Cryptococcus gattii Infection Has Expanded into the Pacific Northwest in the United States. J. Infect. Dis. 2009, 199, 1081–1086. [Google Scholar] [CrossRef]
- Springer, D.J.; Billmyre, R.B.; Filler, E.E.; Voelz, K.; Pursall, R.; Mieczkowski, P.; Larsen, R.A.; Dietrich, F.S.; May, R.C.; Filler, S.G.; et al. Cryptococcus gattii VGIII Isolates Causing Infections in HIV/AIDS Patients in Southern California: Identification of the Local Environmental Source as Arboreal. PLoS Pathog. 2014, 10, e1004285. [Google Scholar] [CrossRef]
- Da Costa, M.M.; Teixeira, F.M.; Schalcher, T.R.; De Brito, M.T.F.M.; Valerio, E.S.; Monteiro, M.C. Cryptococcosis, A Risk for Immunocompromised and Immunocompetent Individuals. Open Epidemiol. J. 2013, 6, 9–17. [Google Scholar] [CrossRef]
- Tintelnot, K.; Lemmer, K.; Losert, H.; Schar, G.; Polak, A. Follow-up of epidemiological data of cryptococcosis in Austria, Germany and Switzerland with special focus on the characterization of clinical isolates. Fortfuhrung der Datenerhebung zur Cryptococcose in Osterreich, Deutschland und der Schweiz unter besonderer Berucksichtigung der Charakterisierung der klinischen Isolate. Mycoses 2004, 47, 455–464. [Google Scholar] [CrossRef]
- Kang, R.; Quah, J.; Low, T.; Chong, C. Cryptococcus gattii infection with pulmonary and CNS involvement in an immu-nocompetent patient. Chest 2016, 149, A117. [Google Scholar] [CrossRef]
- Tello, M.; Gutiérrez, E.; Béjar, V.; Galarza, C.; Ramos, W.; Ortega, A. Criptococosis. Rev. Méd. Risaralda 2013, 19, 147–153. [Google Scholar]
- Galanis, E. Epidemiology of Cryptococcus gattii, British Columbia, Canada, 1999–2007. Emerg. Infect. Dis. 2010, 16, 251–257. [Google Scholar] [CrossRef]
- Li, Y.; Zou, M.; Yin, J.; Liu, Z.; Lu, B. Microbiological, Epidemiological, and Clinical Characteristics of Patients with Cryptococcal Meningitis at a Tertiary Hospital in China: A 6-Year Retrospective Analysis. Front. Microbiol. 2020, 11, 1837. [Google Scholar] [CrossRef]
- Anacona, C.; Vásquez, A.L.R.; Escandón, P. First isolation and molecular characterization of Cryptococcus neoformans var. grubii in excreta of birds in the urban perimeter of the Municipality of Popayán, Colombia. Rev. Iberoam. De Micol. 2018, 35, 123–129. [Google Scholar] [CrossRef]
- Martínez, O.I.C.; Morinelli, M.P.A.; Furnieles, J.L.A.; Ramos, A.M.J. Presumptive identification of Cryptococcus gattii isolated from Terminalia catappa in Montería, Córdoba, Colombia. [Identificación presuntiva de Cryptococcus gattii aislado de Terminalia catappa en Montería, Córdoba, Colombia. Rev. Cubana Med. Trop. 2011, 63, 117–122. [Google Scholar]
- Castañeda, A.; Castañeda, E. Isolation of Cryptococcus species associated with Eucalyptus in a park in Bogotá. Biomédica 2011, 21, 75–78. [Google Scholar] [CrossRef]
- Duarte, A.; Ordoñez, N.; Castañeda, E. Association of yeasts of the genus Cryptococcus with Eucalyptus species in Santafe de Bogota. [Asociacion de leveduras del genero Cryptococcus con especies de Eucalyptus en Santafe de Bogota]. Rev. Inst. Med. Trop. São Paulo 1994, 36, 125–130. [Google Scholar] [CrossRef]
- Virviescas, C.; Aragón, M.; González, F.; Escandón, P.; Castro, H.; Vasquez, L.R. Molecular characterization of Cryptococcus neoformans recovered from pigeon droppings in Rivera and Neiva, Colombia. Rev. MVZ Córdoba 2018, 23, 6991–6997. [Google Scholar] [CrossRef]
- Timarán, D.A.V.; Melo, C.J.B.; Caicedo, M.I.M.; Ceballos, A.M.C.; Vallejo, D.; Velásquez, C.A.C. Aislamiento de Cryptococcus neoformans en heces de palomas (Columba livia) en el casco urbano del municipio de Pasto, Colombia. Biosalud 2016, 15, 62–71. [Google Scholar] [CrossRef]
- Castañeda, A.; Huérfano, S.; Rodríguez, M.C.; Castañeda, E. Recovery of Cryptococcus neoformans var. gattii serotype C from almond tree debris. [Recuperación de Cryptococcus neoformans var. gattii serotipo C a partir de detritos de almendros]. Biomédica 2001, 21, 70–74. [Google Scholar] [CrossRef]
- Callejas, A.; Ordoñez, N.; Rodriguez, M.C.; Castañeda, E. First isolation of Cryptococcus neoformans var gattii, serotype C, from the environment in Colombia. Med. Mycol. 1998, 36, 341–344. [Google Scholar] [CrossRef]
- Firacative, C.; Torres, G.; Rodríguez, M.C.; Escandón, P. First environmental isolation of Cryptococcus gattii serotype B, from Cúcuta, Colombia. Biomédica 2011, 31, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Angarita-Sánchez, A.; Cárdenas-Sierra, D.; Parra-Giraldo, C.; Diaz-Carvajal, C.; Escandon-Hernandez, P. Recovery of environmental Cryptococcus neoformans and Cryptococcus gattii and their association with clinical isolates in Cúcuta, Colom-bia. [Recuperación de Cryptococcus neoformans y Cryptococcus gattii ambientales y su asociación con aislados clínicos en Cú-cuta, Colombia]. MVZ Córdoba 2019, 24, 7137–7144. [Google Scholar]
- Caicedo, B.L.D.; Alvarez, V.M.I.; Llanos, C.E.; Molina, D. Cryptococcus neoformans in pigeon excreta from the urban perimeter of Cali. [Cryptococcus neoformans en excretas de palomas del perímetro urbano de Cali]. Colomb. Med. 1996, 27, 106–109. [Google Scholar]
- Caicedo, L.D.; Alvarez, M.I.; Delgado, M.; Cárdenas, A. Cryptococcus neoformans in bird excreta in the city zoo of Cali, Colombia. Mycopathol. 1999, 147, 121–124. [Google Scholar] [CrossRef]
- Archibald, L.K.; Tuohy, M.J.; Wilson, D.A.; Nwanyanwu, O.; Kazembe, P.N.; Tansuphasawadikul, S.; Eampokalap, B.; Chaovavanich, A.; Reller, L.; Jarvis, W.R.; et al. Antifungal Susceptibilities of Cryptococcus neoformans. Emerg. Infect. Dis. 2004, 10, 143–145. [Google Scholar] [CrossRef]
- Nnadi, N.; Enweani, I.; Cogliati, M.; Ayanbimpe, G.; Okolo, M.; Kim, E.; Sabitu, M.; Criseo, G.; Romeo, O.; Scordino, F. Molecular characterization of environmental Cryptococcus neoformans VNII isolates in Jos, Plateau State, Nigeria. J. Mycol. Méd. 2016, 26, 306–311. [Google Scholar] [CrossRef]
- Irokanulo, E.O.A.; Makinde, A.A.; Akuesgi, C.O.; Ekwonu, M. Cryptococcus neoformans var neoformans Isolated from Droppings of Captive Birds in Nigeria. J. Wildl. Dis. 1997, 33, 343–345. [Google Scholar] [CrossRef]
- Dou, H.; Wang, H.; Xie, S.; Chen, X.; Xu, Z.; Xu, Y. Molecular characterization of Cryptococcus neoformans isolated from the environment in Beijing, China. Med. Mycol. 2017, 55, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Mseddi, F.; Sellami, A.; Jarboui, M.A.; Makni, F.; Ayadi, A. First Environmental Isolations of Cryptococcus neoformans and Cryptococcus gattii in Tunisia and Review of Published Studies on Environmental Isolations in Africa. Mycopathologia 2011, 171, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, M.M.; Lazera, M.S.; Barbosa, G.G.; Trilles, L.; Balassiano, B.R.; Macedo, R.C.L.; Bezerra, C.C.F.; Pérez, M.A.; Cardarelli, P.; Wanke, B. Serotyping of 467 Cryptococcus neoformans Isolates from Clinical and Environmental Sources in Brazil: Analysis of Host and Regional Patterns. J. Clin. Microbiol. 2003, 41, 73–77. [Google Scholar] [CrossRef]
- Kobayashi, C.C.B.A.; Souza, L.K.H.E.; Fernandes, O.D.F.L.; De Brito, S.C.A.; Silva, A.C.; De Sousa, E.D.; Silva, M.D.R.R. Characterization of Cryptococcus neoformans isolated from urban environmental sources in Goiânia, Goiás State, Brazil. Rev. Inst. Med. Trop. São Paulo 2005, 47, 203–207. [Google Scholar] [CrossRef]
- Kidd, S.E.; Chow, Y.; Mak, S.; Bach, P.; Chen, H.; Hingston, A.O.; Kronstad, J.W.; Bartlett, K.H. Characterization of Environmental Sources of the Human and Animal Pathogen Cryptococcus gattii in British Columbia, Canada, and the Pacific Northwest of the United States. Appl. Environ. Microbiol. 2006, 73, 1433–1443. [Google Scholar] [CrossRef]
- Montagna, M.T.; De Donno, A.; Caggiano, G.; Serio, F.; De Giglio, O.; Bagordo, F.; D’Amicis, R.; Lockhart, S.R.; Cogliati, M. Molecular characterization of Cryptococcus neoformans and Cryptococcus gattii from environmental sources and genetic comparison with clinical isolates in Apulia, Italy. Environ. Res. 2018, 160, 347–352. [Google Scholar] [CrossRef] [PubMed]
- Laurenson, I.F.; Lalloo, D.G.; Naraqi, S.; Seaton, R.A.; Trevett, A.J.; Matuka, A.; Kevau, I.H. Cryptococcus neoformans in Papua New Guinea: A common pathogen but an elusive source. [Cryptococcus neoformans en Papua Nueva Guinea: Un patógeno común pero una fuente evasiva]. J. Med. Vet. Mycol. 1997, 35, 437–440. [Google Scholar] [CrossRef]
- Refojo, N.; Perrotta, D.; Brudny, R.; Abrantes, A.; Hevia, I.; Davel, G. Isolation of Cryptococcus neoformans and Cryptococ-cus gattii from trunk hollows of living trees in Buenos Aires City, Argentina. Med. Mycol. 2009, 47, 177–184. [Google Scholar] [CrossRef]
- Cattana, M.E.; de los Ángeles Sosa, M.; Fernández, M.; Rojas, F.; Mangiaterra, M.; Giusiano, G. Native trees of the Northeast Argentine: Natural hosts of the Cryptococcus neoformans-Cryptococcus gattii species complex. Rev. Iberoam. Micol. 2014, 31, 188–192. [Google Scholar] [CrossRef]
- Pedroso, R.S.; Lavrador, M.A.; Ferreira, J.C.; Candido, R.C.; Maffei, C.M. Cryptococcus neoformans var.grubii—Pathogenicity of environmental isolates correlated to virulence factors, susceptibility to fluconazole and molecular profile. Memórias Inst. Oswaldo Cruz 2010, 105, 993–1000. [Google Scholar] [CrossRef]
- Meyer, W.; Castañeda, A.; Jackson, S.; Huynh, M.; Castañeda, E.; The IberoAmerican Cryptococcal Study Group Molecular Typing of IberoAmerican. Cryptococcus neoformans Isolates. Emerg. Infect. Dis. 2003, 9, 189–195. [Google Scholar] [CrossRef]
- Rajasingham, R.; Smith, R.M.; Park, B.J.; Jarvis, J.N.; Govender, N.P.; Chiller, T.M.; Denning, D.W.; Loyse, A.; Boulware, D. Global burden of disease of HIV-associated cryptococcal meningitis: An updated analysis. Lancet Infect. Dis. 2017, 17, 873–881. [Google Scholar] [CrossRef]
- Kumar, C.P.G.; Prabu, D.; Mitani, H.; Mikami, Y.; Menon, T. Environmental isolation of Cryptococcus neoformans and Cryptococcus gattii from living trees in Guindy National Park, Chennai, South India. Mycoses 2010, 53, 262–264. [Google Scholar] [CrossRef]
- Toro Zúñiga, V. Presumptive isolation and characterization of Cryptococcus neoformans and Cryptococcus gattii from trees in the O’Higgins and Maule region, Chile. [Aislamiento presuntivo y caracterización de Cryptococcus neoformans y Cryptococcus gattii desde árboles en]. Boletín Micológico 2015, 30, 6–15. [Google Scholar] [CrossRef][Green Version]
- Kwon-Chung, K.J.; Bennett, J.E. Epidemiologic differences between the two varieties of Cryptococcus neoformans. Am. J. Epidemiol. 1984, 120, 123–130. [Google Scholar] [CrossRef]
- Bauwens, L.; Swinne, D.; De Vroey, C.; De Meurichy, W. Isolation of Cryptococcus neoformans var. neoformans in the aviaries of the Antwerp Zoological Gardens. Mykosen 1986, 29, 291–294. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, C.; Salim, R.; Runco, R. Presence of Cryptococcus neoformans in urban pigeons excreta from San Miguel de Tu-cumán—Argentina. [Presencia de Cryptococcus neoformans en excretas de palomas urbanas en San Miguel de Tucumán—Ar-gentina]. Boletín Micológico 2010, 25, 29–35. [Google Scholar] [CrossRef][Green Version]
- Rosario, I.; Acosta, B.; Colom, F. The pigeon and other birds as reservoirs for Cryptococcus spp. [La paloma y otras aves como reservorio de Cryptococcus spp]. Rev. Iberoam. Micol. 2008, 25, S13–S18. [Google Scholar] [CrossRef]
- Afshari, S.A.K.; Shokohi, T.; Aghili, S.R.; Badali, H. Epidemiology and molecular characterization of Cryptococcus neoformans isolated from pigeon excreta in Mazandaran province, northern Iran. J. Mycol. Méd. 2012, 22, 160–166. [Google Scholar] [CrossRef]
- Abulreesh, H.H.; Organji, S.R.; Elbanna, K.; Osman, G.E.H.; Almalki, M.H.K.; Abdel-Mallek, A.Y. First report of en-vironmental isolation of Cryptococcus neoformans and other fungi from pigeon droppings in Makkah, Saudi Arabia and in vitro susceptibility testing. Asian Pac. J. Trop. Dis. 2015, 5, 622–626. [Google Scholar] [CrossRef]
- Huamán, A.; Béjar, V.; Sáez, G.; Guevara, J.; Sevilla, R.; Tapia, M.; Castillo, E.; Valencia, E.; Marocho, L.; Pareja, E.; et al. Cryptococcus neoformans en heces de palomas (Columba livia) en Lima Metropolitana. Rev. Med. Hered. 2018, 29, 85. [Google Scholar] [CrossRef]
- Cabañes, F.J. Mycoses and zoonoses: Cryptococcus spp. Rev. Iberoam. Micol. 2008, 25, S1–S3. [Google Scholar] [CrossRef]
- Cogliati, M.; D’Amicis, R.; Zani, A.; Montagna, M.T.; Caggiano, G.; De Giglio, O.; Balbino, S.; De Donno, A.; Serio, F.; Susever, S.; et al. Environmental distribution of Cryptococcus neoformans and C. gattii around the Mediterranean basin. FEMS Yeast Res. 2016, 16, 1–12. [Google Scholar] [CrossRef]
- Quintero, E.; Castañeda, E.; Ruiz, A. Environmental distribution of Cryptococcus neoformans in the department of Cun-dinamarca-Colombia. [Distribución ambiental de Cryptococcus neoformans en el departamento de Cundinamarca-Colombia]. Rev. Iberoam. Micol. 2005, 22, 93–98. [Google Scholar] [CrossRef]
- Ergin, Ç.; Şengül, M.; Aksoy, L.; Döğen, A.; Sun, S.; Averette, A.F.; Cuomo, C.A.; Seyedmousavi, S.; Heitman, J.; Ilkit, M. Cryptococcus neoformans Recovered from Olive Trees (Olea europaea) in Turkey Reveal Allopatry With African and South American Lineages. Front. Cell. Infect. Microbiol. 2019, 9, 384. [Google Scholar] [CrossRef]
- Springer, D.J.; Chaturvedi, V. Projecting Global Occurrence of Cryptococcus gattii. Emerg. Infect. Dis. 2010, 16, 14–20. [Google Scholar] [CrossRef]
- Escandón, P.; Sánchez, A.; Firacative, C.; Castañeda, E. Isolation of Cryptococcus gattii molecular type VGIII, from Corymbia ficifolia detritus in Colombia. Med. Mycol. 2010, 48, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Escandón, P.; Quintero, E.; Granados, D.; Huérfano, S.; Ruiz, A.; Castañeda, E. Aislamiento de Cryptococcus gattii serotipo B a partir de detritos de Eucalyptus spp. en Colombia. Biomédica 2005, 25, 390. [Google Scholar] [CrossRef] [PubMed]
- Campbell, L.T.; Fraser, J.A.; Nichols, C.B.; Dietrich, F.S.; Carter, D.; Heitman, J. Clinical and Environmental Isolates of Cryptococcus gattii from Australia That Retain Sexual Fecundity. Eukaryot. Cell 2005, 4, 1410–1419. [Google Scholar] [CrossRef]
- Chowdhary, A.; Randhawa, H.S.; Prakash, A.; Meis, J.F. Environmental prevalence of Cryptococcus neoformans and Cryptococcus gattii in India: An update. Crit. Rev. Microbiol. 2012, 38, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Datta, K.; Bartlett, K.H.; Baer, R.; Byrnes, E.; Galanis, E.; Heitman, J.; Hoang, L.; Leslie, M.J.; MacDougall, L.; Magill, S.S.; et al. Spread of Cryptococcus gattii into Pacific Northwest Region of the United States. Emerg. Infect. Dis. 2009, 15, 1185–1191. [Google Scholar] [CrossRef]
- Hurst, S.; Lysen, C.; Cooksey, G.; Vugia, D.J.; Litvintseva, A.P.; Lockhart, S.R. Molecular typing of clinical and environ-mental isolates of Cryptococcus gattii species complex from southern California, United States. Mycoses 2019, 62, 1029–1034. [Google Scholar] [CrossRef]
- Linares, C.; Colom, M.F.; Torreblanca, M.; Esteban, V.; Álvaro, R.; Hagen, F. Environmental sampling of Ceratonia siliqua (carob) trees in Spain reveals the presence of the rare Cryptococcus gattii genotype AFLP7/VGIV. Rev. Iberoam. Micol. 2015, 32, 269–272. [Google Scholar] [CrossRef]
- Sorrell, T.C. Cryptococcus neoformans variety gattii. Med Mycol. 2001, 39, 155–168. [Google Scholar] [CrossRef]
- Davel, G.; Abrantes, R.; Brudny, M.; Cordoba, S.; Rodero, L.; Canteros, C.E.; Perrotta, D. First environmental isolation of Cryptococcus neoformans var. gattii in Argentina. [Primer aislamiento ambi-ental de Cryptococcus neoformans var. gattii en Argentina]. Rev. Argent. Microbiol. 2003, 35, 110–112. [Google Scholar]
- Andrade-Silva, L. Susceptibility profile of clinical and environmental isolates of Cryptococcus neoformans and Crypto-coccus gattii in Uberaba, Minas Gerais, Brazil. Med. Mycol. 2013, 51, 635–640. [Google Scholar] [CrossRef]
- Brito-Santos, F.; Barbosa, G.G.; Trilles, L.; Nishikawa, M.M.; Wanke, B.; Meyer, W.; Carvalho-Costa, F.A.; Lazéra, M.D.S. Environmental Isolation of Cryptococcus gattii VGII from Indoor Dust from Typical Wooden Houses in the Deep Amazonas of the Rio Negro Basin. PLoS ONE 2015, 10, e0115866. [Google Scholar] [CrossRef] [PubMed]
- Kassi, F.K.; Bellet, V.; Drakulovski, P.; Krasteva, D.; Roger, F.; Valérie, B.T.A.; Aboubakar, T.; Doumbia, A.; Kouakou, G.A.; Delaporte, E.; et al. Comparative typing analyses of clinical and environmental strains of the Cryptococcus neofor-mans/Cryptococcus gattii species complex from ivory coast. J. Med. Microbiol. 2018, 67, 87–96. [Google Scholar] [CrossRef]
- Randhawa, H.S.; Kowshik, T.; Preeti Sinha, K.; Chowdhary, A.; Khan, Z.U.; Yan, Z.; Xu, J.; Kumar, A. Distribution of Cryptococcus gattii and Cryptococcus neoformans in decayed trunk wood of Syzygium cumini trees in north-western India. Med. Mycol. 2006, 44, 623–630. [Google Scholar] [CrossRef]
- Casadevall, A.; Perfect, J.R. Cryptococcus Neoformans; ASM Press: Washington, DC, USA, 1998; Volume 595. [Google Scholar] [CrossRef]
- Ellis, D.H.; Pfeiffer, T.J. Natural habitat of Cryptococcus neoformans var. gattii. J. Clin. Microbiol. 1990, 28, 1642–1644. [Google Scholar] [CrossRef]
- Byrnes, E.J.; Bartlett, K.H.; Perfect, J.R.; Heitman, J. Cryptococcus gattii: An emerging fungal pathogen infecting humans and animals. Microbes Infect. 2011, 13, 895–907. [Google Scholar] [CrossRef]
- Firacative, C.; Lizarazo, J.; Illnait-Zaragozí, M.T.; Castañeda, E. The status of cryptococcosis in Latin America. Memórias Inst. Oswaldo Cruz 2018, 113, e170554. [Google Scholar] [CrossRef]
- Bartlett, K.H.; Kidd, S.E.; Kronstad, J.W. The emergence of Cryptococcus gattii in British Columbia and the Pacific Northwest. Curr. Infect. Dis. Rep. 2008, 10, 58–65. [Google Scholar] [CrossRef]
- Upton, A.; Fraser, J.A.; Kidd, S.E.; Bretz, C.; Bartlett, K.H.; Heitman, J.; Marr, K.A. First Contemporary Case of Human Infection with Cryptococcus gattii in Puget Sound: Evidence for Spread of the Vancouver Island Outbreak. J. Clin. Microbiol. 2007, 45, 3086–3088. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cogliati, M.; Medica, L.M.; Biomediche, S.; Pascal, V. Global molecular epidemiology of Cryptococcus neoformans and Cryptococcus gattii: An atlas of molecular types. Hindawi Publ. Corp. Sci. 2013, 2013, 1–23. [Google Scholar]
- Kidd, S.E.; Hagen, F.; Tscharke, R.L.; Huynh, M.; Bartlett, K.H.; Fyfe, M.; Macdougall, L.; Boekhout, T.; Kwon-Chung, K.J.; Meyer, W. A rare genotype of Cryptococcus gattii caused the cryptococcosis outbreak on Vancouver Island (British Co-lumbia, Canada). Proc. Natl. Acad. Sci. USA 2004, 101, 17258–17263. [Google Scholar] [CrossRef]
- Escandón, P.; Lizarazo, J.; Agudelo, C.I.; Castañeda, E. Cryptococcosis in Colombia: Compilation and analysis of data from laboratory based surveillance. J. Fungi (Basel) 2018, 4, 32. [Google Scholar] [CrossRef]
- Granados, D.P.; Castañeda, E. Isolation and characterization of Cryptococcus neoformans varieties recovered from natu-ral sources in Bogotá, Colombia, and study of ecological conditions in the área. Microb. Ecol. 2005, 49, 282–290. [Google Scholar] [CrossRef] [PubMed]
- Valiente, M.F.; Alberdi, M.; Meseguer, I.; Torres Rodriguez, J.M. Isolation of Cryptococcus neoformans in environmental samples from Alicante. [Aislamiento de Cryptococcus neoformans en muestras de medio ambiente de Alicante]. Rev. Iberoam. Micol. 1997, 14, 63–64. [Google Scholar]
- Noguera, M.C.; Escandón, P.; Castañeda, E. Cryptococcosis in Atlántico, Colombia: An approximation of the preva-lence of this mycosis and the distribution of the etiological agent in the environment. Rev. Soc. Bras. Med. Trop. 2015, 48, 580–586. [Google Scholar] [CrossRef] [PubMed]
- Instituto de Hidrología Meteorología y Estudios Ambientales—IDEAM. Study of the Climatic Characterization of Bogotá and the Upper Tunjuelo River Basin; Instituto de Hidrología Meteorología y Estudios Ambientales: Bogotá, Colombia, 1999; pp. 19–21. [Google Scholar]
- Gonzalo Hurtado, M. Analysis of the Average Behavior and Long-Term Trends of the Mean Maximum Temperature for the Hydroclimatic Regions of Colombia; Instituto de Hidrología Meteorología y Estudios Ambientales: Bogotá, Colombia, 2012; p. 61. [Google Scholar]
- IDEAM. Map: Multi-Year Average Annual Total Mean Precipitation 1981–2010; Instituto de Hidrología Meteorología y Estudios Ambientales: Bogotá, Colombia, 2014; p. 1. [Google Scholar]
- Bennett, J.; Kwon-Chung, K.; Theodore, T.Y. Biochemical differences between Cryptococcus neoformans serotypes. [Diferencias bioquímicas entre los serotipos de Cryptococcus neoformans]. Med. Mycol. 1978, 16, 167–174. [Google Scholar] [CrossRef]
- Cadena, M.; López, N.; Carlos, R.; Vega, A.; González, O.; Cubillos, M.I. Climatological Yearbook 2016; Instituto de Hidrología Meteorología y Estudios Ambientales: Bogotá, Colombia, 2016; p. 355. [Google Scholar]
- Mak, S.; Vélez, N.; Castañeda, E.; Escandón, P.; Colombian Group. The Fungus among Us: Cryptococcus neoformans and Cryptococcus gattii Ecological Modeling for Colombia. J. Fungi 2015, 1, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Granados, D.P.; Castañeda, E. Influence of climatic conditions on the isolation of members of the Cryptococcus neoformans species complex from trees in Colombia from 1992–2004. FEMS Yeast Res. 2006, 6, 636–644. [Google Scholar] [CrossRef]
- Watkins, R.; King, J.; Johnston, S. Nutritional Requirements and Their Importance for Virulence of Pathogenic Crypto-coccus Species. Microorganisms 2017, 5, 65. [Google Scholar] [CrossRef]
- Mazuelos, E.M.; Ana Isabel, A.G. Microbiological aspects of cryptococcosis in the post-HAART era. [Aspectos microbi-ológicos de la criptococosis en la era post-TARGA]. Enferm. Infecc. Microbiol. Clin. 2010, 28, 40–45. [Google Scholar] [CrossRef]
- Cogliati, M.; Puccianti, E.; Montagna, M.T.; De Donno, A.; Susever, S.; Ergin, C.; Velegraki, A.; Ellabib, M.S.; Nardoni, S.; Macci, C.; et al. Fundamental niche prediction of the pathogenic yeasts Cryptococcus neoformans and Cryptococcus gattii in Europe. Environ. Microbiol. 2017, 19, 4318–4325. [Google Scholar] [CrossRef]
- Sorrell, T.C.; David, E.H. Ecology of Cryptococcus neoformans in central Africa. Rev. Iberoam. Micol. 1997, 14, 42–43. [Google Scholar] [PubMed]
- Wang, Y.; Aisen, P.; Casadevall, A. Cryptococcus neoformans melanin and virulence: Mechanism of action. Infect. Immun. 1995, 63, 3131–3136. [Google Scholar] [CrossRef]
- Lazéra, M.S.; Pires, F.D.; Camillo-Coura, L.; Nishikawa, M.M.; Bezerra, C.C.; Trilles, L.; Wanke, B. Natural habitat of Cryptococcus neoformans var. neoformans in decaying wood forming hollows in living trees. J. Med. Vet. Mycol. 1996, 34, 127–131. [Google Scholar] [CrossRef] [PubMed]
- Escandón, P.; Huérfano, S.; Castañeda, E. Experimental inoculation of Terminalia catappa with an environmental isolate of Cryptococcus neoformans var. gattii serotype C. Inoculación experimental de Terminalia catappa con un aislamiento ambien-tal de Cryptococcus neoformans var. gattii serotipo C]. Biomédica 2002, 22, 524. [Google Scholar] [CrossRef][Green Version]
- Randhawa, H.S.; Kowshik, T.; Khan, Z.U. Decayed wood of Syzygium cumini and Ficus religiosa living trees in Del-hi/New Delhi metropolitan area as natural habitat of Cryptococcus neoformans. Med. Mycol. 2003, 41, 199–209. [Google Scholar] [CrossRef] [PubMed]
- Levitz, S.M. The Ecology of Cryptococcus neoformans and the Epidemiology of Cryptococcosis. Clin. Infect. Dis. 1991, 13, 1163–1169. [Google Scholar] [CrossRef]
- Ruiz, A.; Fromtling, R.A.; Bulmer, G.S. Distribution of Cryptococcus neoformans in a natural site. Infect. Immun. 1981, 31, 560–563. [Google Scholar] [CrossRef] [PubMed]
- Ordóñéz, N.; Castaneda, E. Serotipificación de aislamientos clínicos y del medio ambiente de Cryptococcus neoformans en Colombia. Biomédica 1994, 14, 131–139. [Google Scholar] [CrossRef][Green Version]
- Litvintseva, A.P.; Thakur, R.; Reller, L.B.; Mitchell, T.G. Prevalence of Clinical Isolates of Cryptococcus gattii Serotype C among Patients with AIDS in Sub-Saharan Africa. J. Infect. Dis. 2005, 192, 888–892. [Google Scholar] [CrossRef]
- Escandón, P.; Sánchez, A.; Martínez, M.; Meyer, W.; Castañeda, E. Molecular epidemiology of clinical and environmental isolates of the Cryptococcus neoformans species complex reveals a high genetic diversity and the presence of the molecular type VGII mating type a in Colombia. FEMS Yeast Res. 2006, 6, 625–635. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).