
Discovery of Novel Backusella (Backusellaceae, Mucorales) Isolated from Invertebrates and Toads in Cheongyang, Korea

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Fungal Isolation
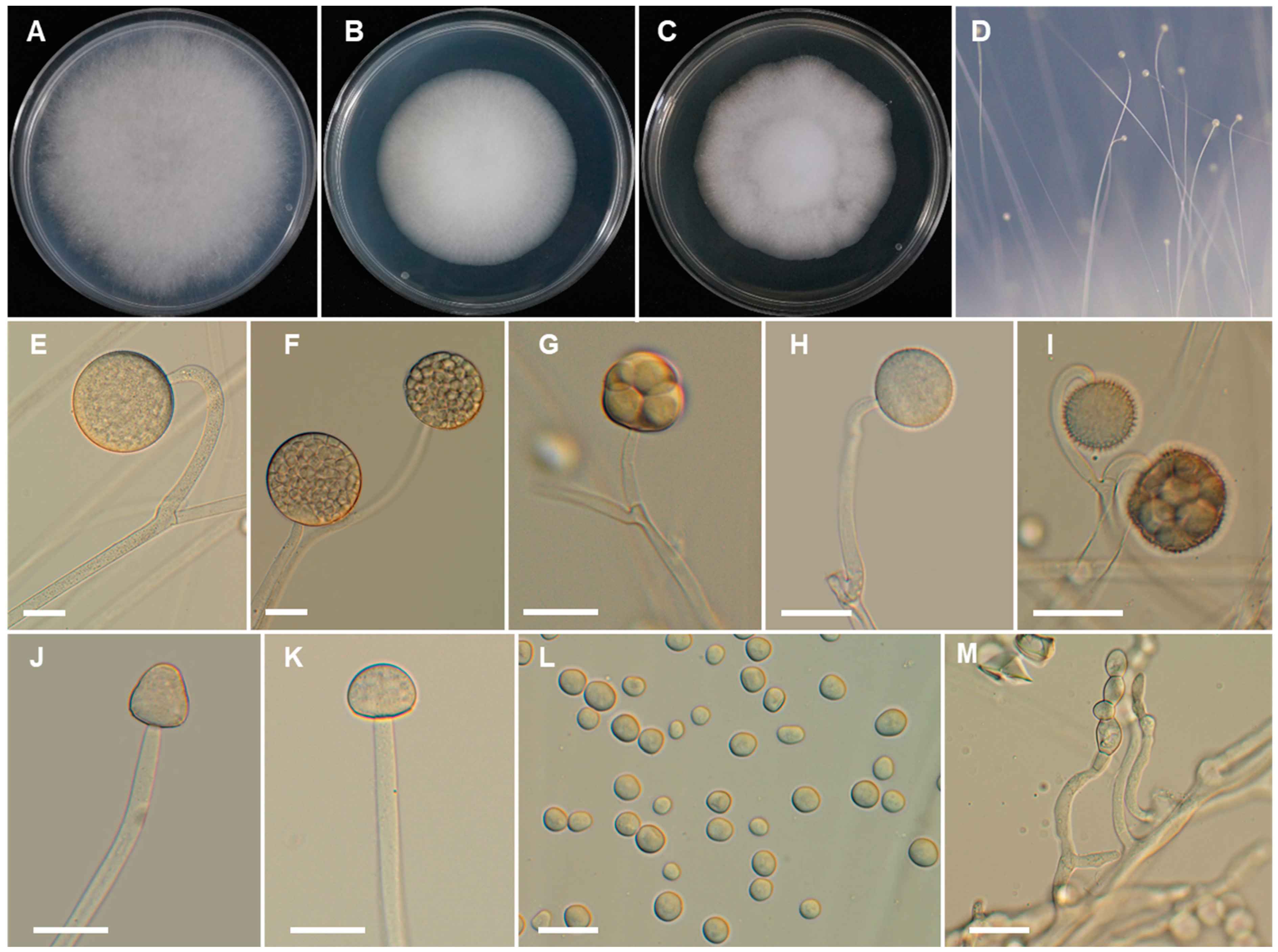
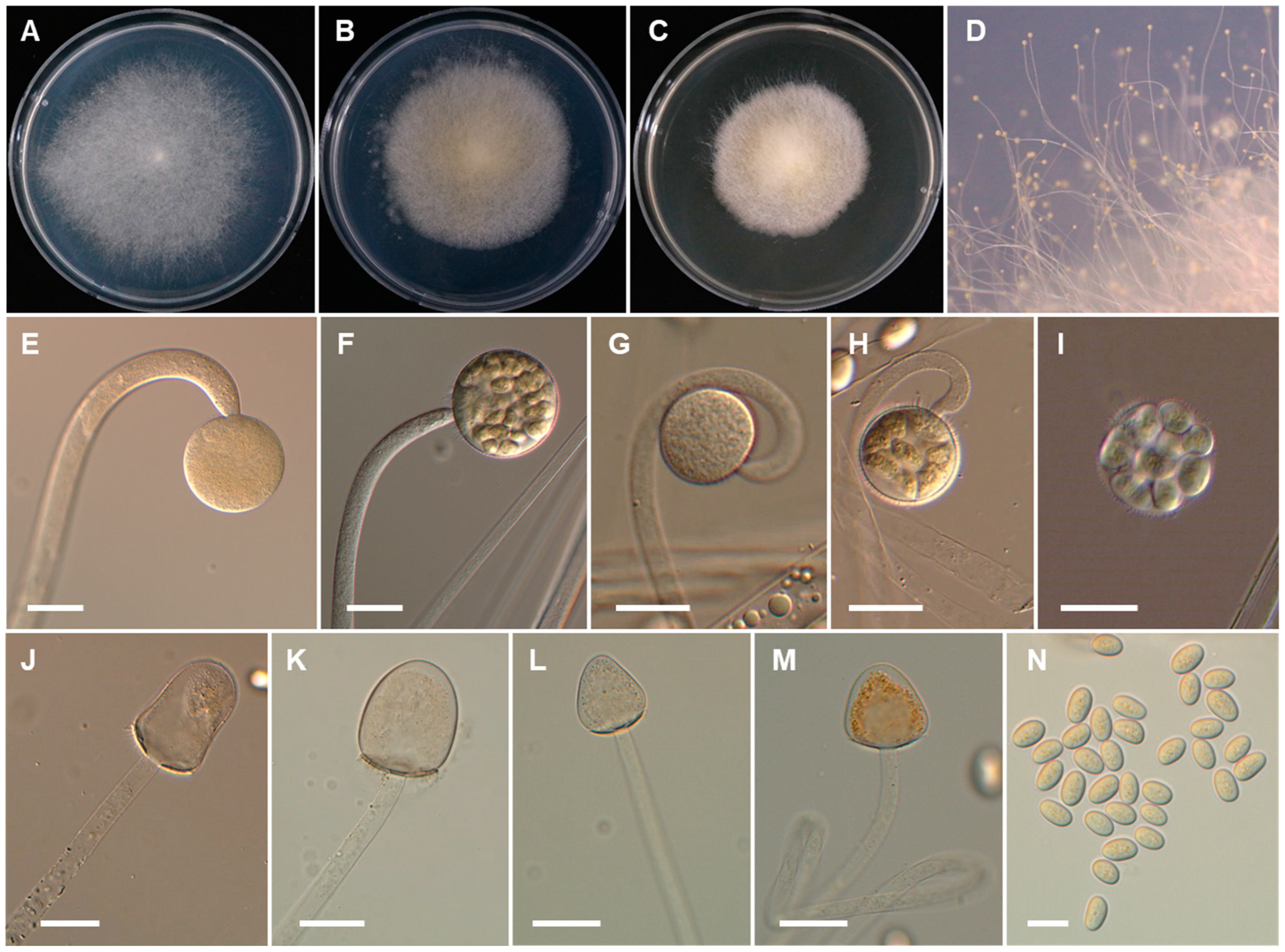
2.2. Morphological Studies
2.3. DNA Extraction, PCR, Cloning, and Sequencing
2.4. Sequence Alignment and Phylogenetic Analyses
3. Results
3.1. Phylogenetic Analysis
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ellis, J.J.; Hesseltine, C.W. Two new members of the Mucorales. Mycologia 1969, 61, 863–872. [Google Scholar] [CrossRef]
- Pidoplichko, N.M.; Mil’ko, A.A. Atlas of Mucoralean Fungi; Academy of Sciences of the Ukrainian SSR: Ukrainian SSR, 1971; p. 188. (In Russian) [Google Scholar]
- Benny, G.L.; Benjamin, R.K. Observations on Thamnidiaceae (Mucorales). New taxa, new combinations, and notes on selected species. Aliso 1975, 8, 301–351. [Google Scholar] [CrossRef] [Green Version]
- Walther, G.; Pawłowska, J.; Alastruey-Izquierdo, A.; Wrzosek, M.; Rodriguez-Tudela, J.L.; Dolatabadi, S.; Chakrabarti, A.; de Hoog, G.S. DNA barcoding in Mucorales: An inventory of biodiversity. Persoonia 2013, 30, 11–47. [Google Scholar] [CrossRef] [Green Version]
- Crous, P.; Carnegie, A.J.; Wingfield, M.J.; Sharma, R.; Mughini, G.; Noordeloos, M.E.; Santini, A.; Shouche, Y.S.; Bezerra, J.D.P.; Dima, B.; et al. Fungal Planet description sheets: 868–950. Persoonia 2019, 42, 291–473. [Google Scholar] [CrossRef] [PubMed]
- Lima, D.X.; Voigt, K.; de Souza, C.A.; de Oliveira, R.J.; Souza-Motta, C.M.; Santiago, A.L.C.M.d.A. Backusella constricta sp. nov. (Mucorales, ex Zygomycota) from the Brazilian Atlantic Rainforest, including a key to species of Backusella. Phytotaxa 2016, 289, 59–68. [Google Scholar] [CrossRef]
- de Souza, J.I.; Marano, A.V.; Pires-Zottarelli, C.L.A.; Chambergo, F.S.; Harakava, R. A new species of Backusella (Mucorales) from a Cerrado reserve in Southeast Brazil. Mycol. Prog. 2014, 13, 975–980. [Google Scholar] [CrossRef]
- Wanasinghe, D.N.; Phukhamsakda, C.; Hyde, K.D.; Jeewon, R.; Lee, H.B.; Jones, E.B.G.; Tibpromma, S.; Tennakoon, D.S.; Dissanayake, A.J.; Jayasiri, S.C.; et al. Fungal diversity notes 709–839: Taxonomic and phylogenetic contributions to fungal taxa with an emphasis on fungi on Rosaceae. Fungal Divers. 2018, 89, 1–236. [Google Scholar] [CrossRef]
- Urquhart, A.S.; Douch, J.K.; Heafield, T.A.; Buddie, A.G.; Idnurm, A. Diversity of Backusella (Mucoromycotina) in south-eastern Australia revealed through polyphasic taxonomy. Persoonia 2021, 46, 1–25. [Google Scholar] [CrossRef]
- Kirk, P.M. Nomenclatural novelties. Index Fungorum. 2012, 11, 1. [Google Scholar]
- Nguyen, T.T.T.; Lee, H.B. Isolation and characterization of three Zygomycetous fungi in Korea: Backusella circina, Circinella muscae, and Mucor ramosissimus. Mycobiology 2018, 46, 317–327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benny, G.L. Current systematics of Zygomycota with a brief review of their biology. In Systematics and Evolution of Fungi; Misra, J.K., Tewari, J.P., Deshmukh, S.K., Eds.; Science: Enfield, UK, 2012; pp. 55–105. [Google Scholar]
- Santiago, A.L.C.M.d.A.; Trufem, S.F.B.; Malosso, E.; Santos, P.J.P.; Cavalcanti, M.A.de.Q. Zygomycetes from herbivore dung in the ecological reserve of Dois Irmãos, Northeast Brazil. Braz. J. Microbiol. 2011, 42, 89–95. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.W.; Hyde, K.D.; Ho, W.H. Single spore isolation of fungi. Fungal Divers. 1999, 3, 29–38. [Google Scholar]
- de Hoog, G.S.; Gerrits van den Ende, A.H.G. Molecular diagnostics of clinical strains of filamentous Basidiomycetes. Mycoses 1998, 41, 183–189. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- Bunyard, B.A.; Nicholson, M.S.; Royse, D.J. A systematic assessment of Morchella using RFLP analysis of the 28S ribosomal RNA gene. Mycologia 1994, 86, 762–772. [Google Scholar] [CrossRef]
- Vilgalys, R.; Hester, M. Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several Cryptococcus species. J. Bacteriol. 1990, 172, 4238–4246. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Glez-Peña, D.; Gómez-Blanco, D.; Reboiro-Jato, M.; Fdez-Riverola, F.; Posada, D. ALTER: Program-oriented format conversion of DNA and protein alignments. Nucleic Acids Res. 2010, 38, 14–18. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods. 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.T.T.; Lee, H.B. Mucor cheongyangensis, a new species isolated from the surface of Lycorma delicatula in Korea. Phytotaxa 2020, 446, 33–42. [Google Scholar] [CrossRef]
- Schipper, M.A.A.; Samson, R.A. Miscellaneous notes on Mucoraceae. Mycotaxon 1994, 50, 475–491. [Google Scholar]
- Sarbhoy, A.K. Mucor variabilis sp. nov. from India. Trans. Brit. Mycol. Soc. 1965, 48, 559–560. [Google Scholar] [CrossRef]
- Walther, G.; Wagner, L.; Kurzai, O. Updates on the taxonomy of Mucorales with an emphasis on clinically important taxa. J. Fungi 2019, 5, 106. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon Name | Strain Number | Host/Substrate | Country | GenBank Accession Number | |
|---|---|---|---|---|---|
| ITS | LSU | ||||
| B. australiensis | UoMAU34 T | Leaf litter | Australia | MK959062 | MK958800 |
| B. australiensis | UoMAU33 | – | Australia | – | MK958801 |
| B. azygospora | URM 8065 T | Soil | Brazil | MK625216 | MK625222 |
| B. chlamydospora | CNUFC PS1 T | Porcellio scaber | South Korea | MZ171385 | MZ148709 |
| B. chlamydospora | CNUFC HL7 | Theuronema hilgendorfi hilgendorfi | South Korea | MZ171386 | MZ148710 |
| B. circina | CBS 128.70 T | Soil with lichens | USA | JN206258 | JN206529 |
| B. constricta | URM 7322 T | Soil | Brazil | KT937158 | KT937156 |
| B. dispersa | CBS 195.28 | Fallen leaf | USA | JN206271 | JN206530 |
| B. dispersa | UoMAU119 | – | Australia | – | MK958770 |
| B. dispersa | UoMAU116 | – | Australia | – | MK958769 |
| B. gigacellularis | CCIBt 3866 T | Leaf litter canopy-plates | Brazil | KF742415 | – |
| B. grandis | CBS 186.87 T | Dung of mouse | India | JN206252 | JN206527 |
| B. indica | CBS 786.70 LT | – | India | JN206255 | JN206526 |
| B. koreana | CNUFC CM05 T | Scolopendra morsitans | South Korea | MZ171387 | MZ148711 |
| B. koreana | CNUFC CM06 | Scolopendra morsitans | South Korea | MZ171388 | MZ148712 |
| B. lamprospora | CBS 118.08 T | – | Switzerland | JN206268 | JN206531 |
| B. liffmaniae | UoMAU58 T | Leaf litter | Australia | MK959065 | MK958734 |
| B. liffmaniae | UoMAU128 | – | Australia | – | MK958735 |
| B. locustae | EML-SFB4 | Grasshopper feces | South Korea | KY449293 | KY449290 |
| B. locustae | EML-SFB2 T | Grasshopper feces | South Korea | KY449291 | KY449292 |
| B. luteola | UoMAU6 T | Leaf litter | Australia | MK959058 | MK958795 |
| B. luteola | UoMAU36 | – | Australia | – | MK958794 |
| B. macrospora | UoMAU7 T | Leaf litter | Australia | MK959107 | MK958628 |
| B. mclennaniae | UoMAU11 T | Leaf litter | Australia | MK958776 | MK958776 |
| B. mclennaniae | UoMAU12 | – | Australia | MK959087 | MK958777 |
| B. morwellensis | UoMAU16 T | Leaf litter | Australia | MK959059 | MK958808 |
| B. morwellensis | UoMAU14 | – | Australia | – | MK958806 |
| B. oblongielliptica | CBS 568.70 LT | Agaric | Japan | JN206278 | JN206533 |
| B. oblongielliptica | CNUFC IL02 | Timomenus komarovi | South Korea | MZ171391 | MZ148715 |
| B. oblongispora | CBS 569.70 NT | Soil | Japan | JN206251 | JN206407 |
| B. oblongispora | CNUFC TKB11 | Skin of Bufo gargarizans | South Korea | MZ420786 | MZ148717 |
| B. parvicylindrica | UoMAU35 T | Leaf litter | Australia | MK959109 | MK958727 |
| B. parvicylindrica | UoMAU213 | – | Australia | – | MK958732 |
| B. psychrophila | UoMAU55 T | Leaf litter | Australia | MK959093 | MK958749 |
| B. psychrophila | UoMAU26 | – | Australia | – | MK958748 |
| B. recurva | CBS 318.52 ET | Fragaria; diseased root | USA | JN206261 | JN206522 |
| B. recurva | CBS 196.71 | – | n.a | JN206265 | JN206523 |
| Backusella sp. | CBS 538.80 | Medicago sativa | Egypt | HM999964 | HM849692 |
| B. tarrabulga | UoMAU5 T | Leaf litter | Australia | MK959060 | MK958804 |
| B. tarrabulga | UoMAU187 | – | Australia | – | MK958805 |
| B. thermophila | CNUFC CS02 T | Gryllus bimaculatus | South Korea | MZ171389 | MZ148713 |
| B. thermophila | CNUFC CS03 | Gryllus bimaculatus | South Korea | MZ171390 | MZ148714 |
| B. tuberculispora | CBS 562.66 LT | – | India | JN206267 | JN206525 |
| B. tuberculispora | CBS 570.70 | – | Japan | MH859852 | MH871631 |
| B. variabilis | CBS 564.66 LT | Excrements of human | India | JN206254 | JN206528 |
| B. westeae | UoMAU4 T | Contaminant during attempts to culture Laccaria species from freshly collected sporocarps | Australia | MK959061 | MK958796 |
| Backusella ‘group X’ | UoMAU9 | – | Australia | MK959098 | MK958787 |
| Backusella ‘group X’ | UoMAU98 | – | Australia | MK959099 | MK958789 |
| Mucor indicus | CBS 226.29 ET | – | Switzerland | HM999956 | HM849690 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, T.T.T.; Voigt, K.; Santiago, A.L.C.M.d.A.; Kirk, P.M.; Lee, H.B. Discovery of Novel Backusella (Backusellaceae, Mucorales) Isolated from Invertebrates and Toads in Cheongyang, Korea. J. Fungi 2021, 7, 513. https://doi.org/10.3390/jof7070513
Nguyen TTT, Voigt K, Santiago ALCMdA, Kirk PM, Lee HB. Discovery of Novel Backusella (Backusellaceae, Mucorales) Isolated from Invertebrates and Toads in Cheongyang, Korea. Journal of Fungi. 2021; 7(7):513. https://doi.org/10.3390/jof7070513
Chicago/Turabian StyleNguyen, Thuong T.T., Kerstin Voigt, André Luiz Cabral Monteiro de Azevedo Santiago, Paul M. Kirk, and Hyang Burm Lee. 2021. "Discovery of Novel Backusella (Backusellaceae, Mucorales) Isolated from Invertebrates and Toads in Cheongyang, Korea" Journal of Fungi 7, no. 7: 513. https://doi.org/10.3390/jof7070513