
Synergistic Antifungal Activity of Chito-Oligosaccharides and Commercial Antifungals on Biofilms of Clinical Candida Isolates


Abstract
:1. Introduction
2. Materials and Methods
2.1. Enzymatic Production of Chito-Oligosaccharides
2.2. Commercial Antifungals (CA)
2.3. Candida Strains
2.4. Candida Inocula
2.5. Biofilm Formation
2.6. Candida Cell Enumeration in Biofilms
2.7. Metabolic Activity Determination
2.8. Effect of C32 and Flu on Preformed Biofilms
2.9. Effect of C32 and CA on Biofilm Formation
2.10. Synergy
2.11. Statistical Analysis of Data
3. Results
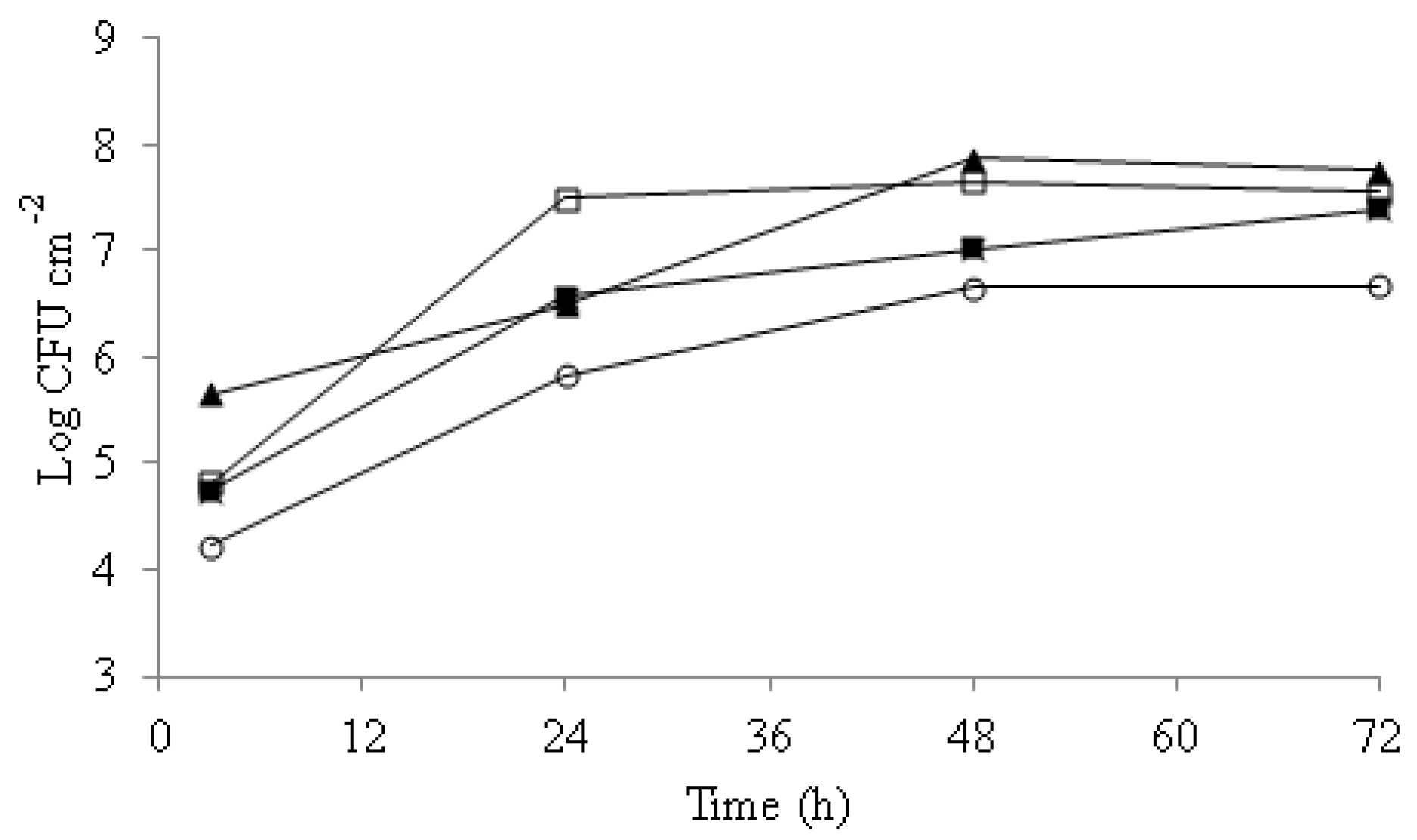
3.1. Candida Biofilm Formation
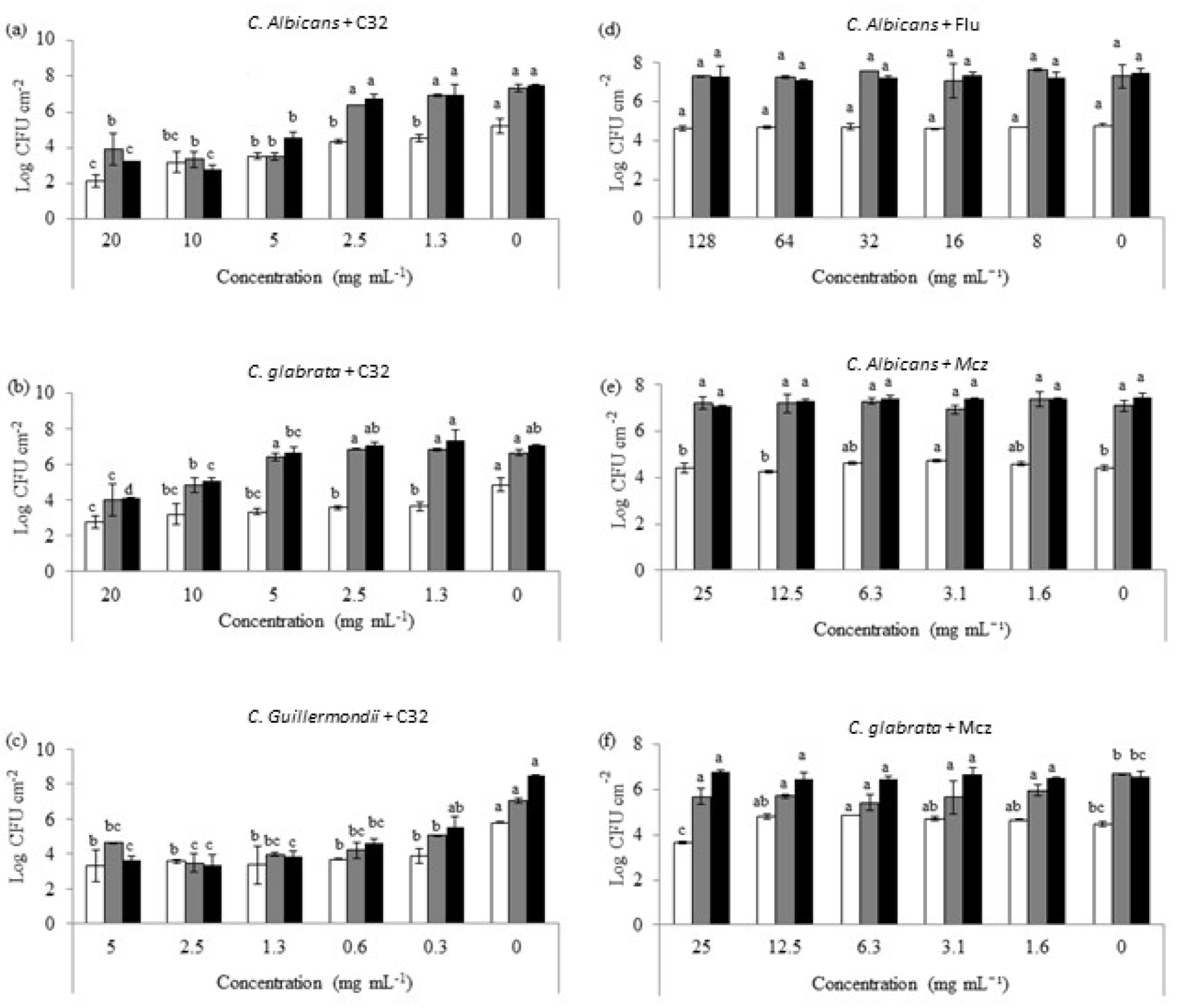
3.2. Effect of C32 and CA on Preformed Candida Biofilms
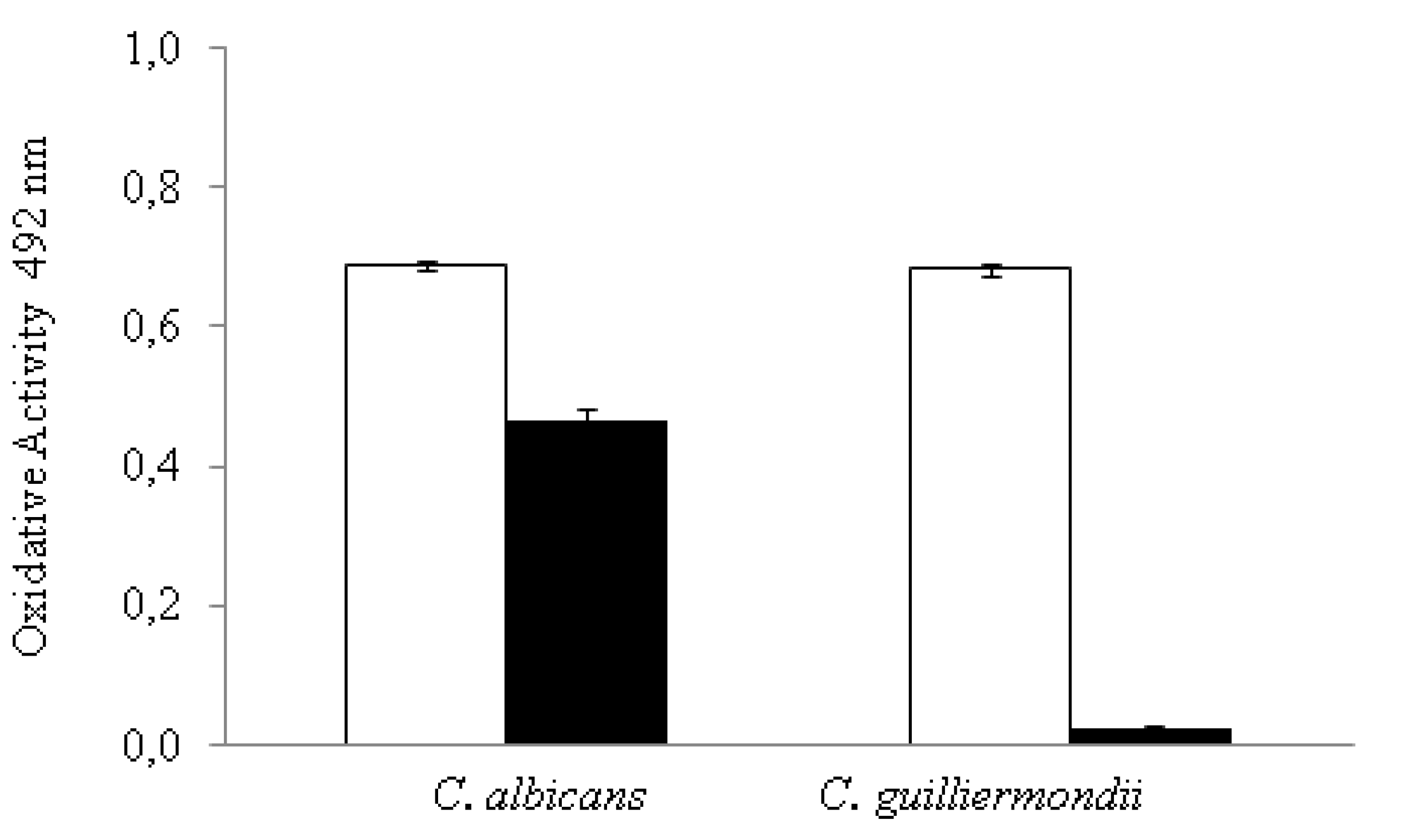
3.3. Effect of C32 on the Metabolic Activity of Candida Biofilms
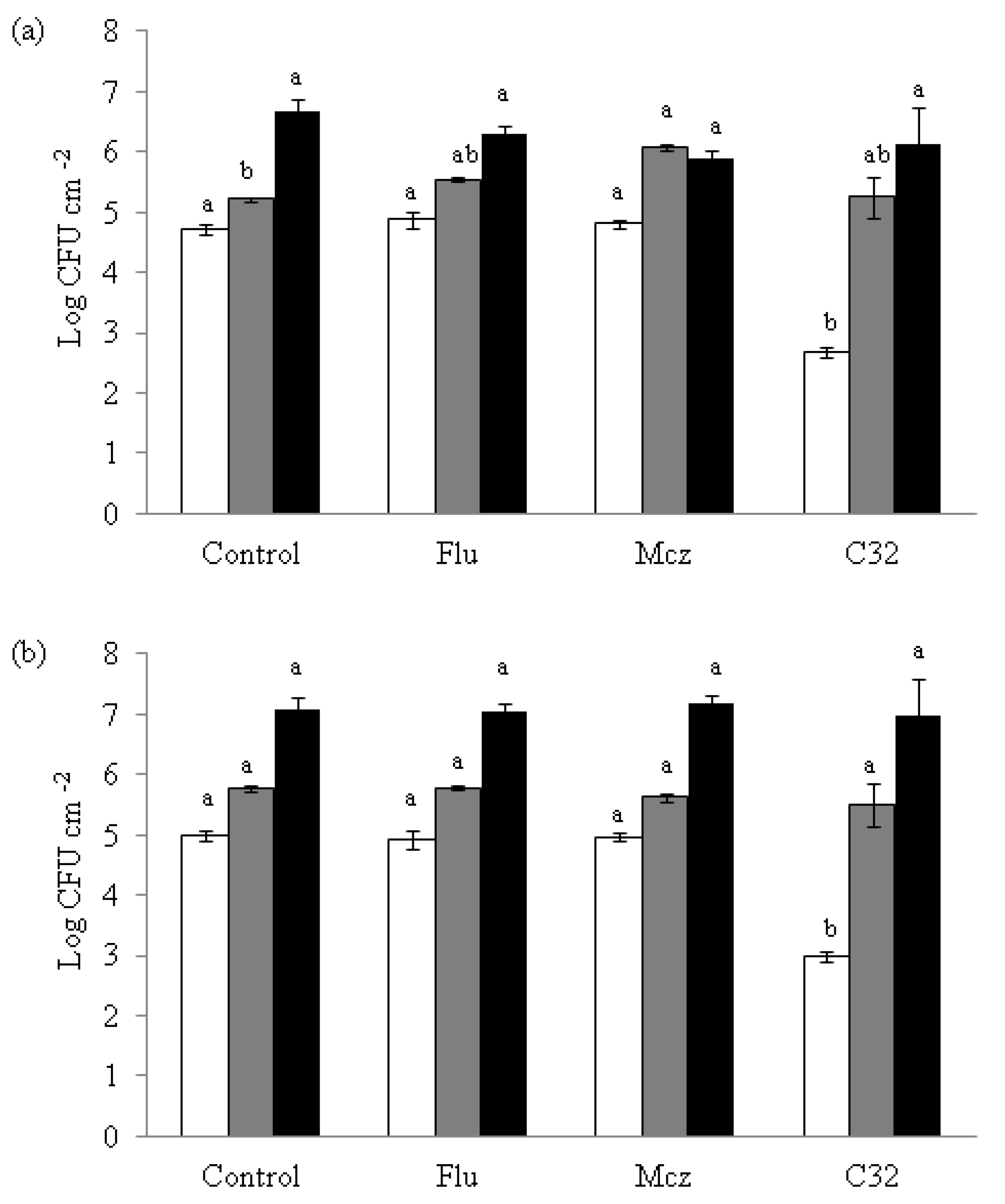
3.4. Effect of C32 on the Formation of Candida Biofilms
3.5. Combined Antifungal Effect of C32 and CA on Preformed Candida Biofilms
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Colombo, A.L.; Júnior, J.N.D.A.; Guinea, J. Emerging multidrug-resistant Candida species. Curr. Opin. Infect. Dis. 2017, 30, 528–538. [Google Scholar] [CrossRef] [PubMed]
- Ganan, M.; Lorentzen, S.B.; Agger, J.W.; Heyward, C.A.; Bakke, O.; Knutsen, S.H.; Aam, B.B.; Eijsink, V.G.H.; Gaustad, P.; Sørlie, M. Antifungal activity of well-defined chito-oligosaccharide preparations against medically relevant yeasts. PLoS ONE 2019, 14, e0210208. [Google Scholar] [CrossRef] [PubMed]
- Ganan, M.; Lorentzen, S.B.; Aam, B.B.; Eijsink, V.G.H.; Gaustad, P.; Sørlie, M. Antibiotic saving effect of combination therapy through synergistic interactions between well-characterized chito-oligosaccharides and commercial antifungals against medically relevant yeasts. PLoS ONE 2019, 14, e0227098. [Google Scholar] [CrossRef]
- Rahman, H.; Shovan, L.R.; Hjeljord, L.G.; Aam, B.B.; Eijsink, V.G.H.; Sørlie, M.; Tronsmo, A. Inhibition of Fungal Plant Pathogens by Synergistic Action of Chito-Oligosaccharides and Commercially Available Fungicides. PLoS ONE 2014, 9, e93192. [Google Scholar] [CrossRef]
- Chandra, J.; Kuhn, D.M.; Mukherjee, P.K.; Hoyer, L.L.; McCormick, T.; Ghannoum, M.A. Biofilm Formation by the Fungal Pathogen Candida Albicans: Development, Architecture, and Drug Resistance. J. Bacteriol. 2001, 183, 5385–5394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raad, I.; Hanna, H.; Maki, D. Intravascular Catheter-Related Infections: Advances in Diagnosis, Prevention, and Management. Lancet Infect. Dis. 2007, 7, 645–657. [Google Scholar] [CrossRef]
- Mermel, L.; Allon, M.; Bouza, E.; Craven, D.; Flynn, P.; O’Grady, N.; Raad, I.; Rijnders, B.; Sherertz, R.; Warren, D. Clinical Practice Guidelines for the Diagnosis and Management of Intravascular Catheter-Related Infection: 2009 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 49, 1–45. [Google Scholar] [CrossRef]
- Harriott, M.M.; Lilly, E.A.; Rodriguez, T.E.; Fidel, P.L.; Noverr, M. Candida albicans forms biofilms on the vaginal mucosa. Microbiology 2010, 156, 3635–3644. [Google Scholar] [CrossRef] [Green Version]
- GGanguly, S.; Mitchell, A.P. Mucosal biofilms of Candida albicans. Curr. Opin. Microbiol. 2011, 14, 380–385. [Google Scholar] [CrossRef] [Green Version]
- Ramage, G.; Tomsett, K.; Wickes, B.L.; López-Ribot, J.L.; Redding, S.W. Denture Stomatitis: A Role for Candida Biofilms. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endodont. 2004, 98, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Douglas, L. Candida biofilms and their role in infection. Trends Microbiol. 2003, 11, 30–36. [Google Scholar] [CrossRef]
- Kean, T.; Thanou, M. Biodegradation, biodistribution and toxicity of chitosan. Adv. Drug Deliv. Rev. 2010, 62, 3–11. [Google Scholar] [CrossRef]
- Kasaai, M.R. Determination of the degree of N-acetylation for chitin and chitosan by various NMR spectroscopy techniques: A review. Carbohydr. Polym. 2010, 79, 801–810. [Google Scholar] [CrossRef]
- Knaul, J.Z.; Kasaai, M.R.; Bui, V.T.; Creber, K.A. Characterization of deacetylated chitosan and chitosan molecular weight review. Can. J. Chem. 1998, 76, 1699–1706. [Google Scholar] [CrossRef]
- Wang, Q.Z.; Chen, X.G.; Liu, N.; Wang, S.X.; Liu, C.; Meng, X.H. Protonation constants of chitosan with different molecular weight and degree of deacetylation. Carbohydr. Polym. 2006, 65, 194–201. [Google Scholar] [CrossRef]
- Atanase, L. Micellar Drug Delivery Systems Based on Natural Biopolymers. Polymers 2021, 13, 477. [Google Scholar] [CrossRef] [PubMed]
- Iurciuc-Tincu, C.E.; Atanase, L.I.; Ochiuz, L.; Jérôme, C.; Sol, V.; Martin, P.; Popa, M. Curcumin-Loaded Polysaccharides-Based Complex Particles Obtained by Polyelectrolyte Complexation and Ionic Gelation. I-Particles Obtaining and Characterization. Int. J. Biol. Macromol. 2020, 147, 629–642. [Google Scholar] [CrossRef] [PubMed]
- Rata, D.M.; Cadinoiu, A.N.; Atanase, L.I.; Bacaita, S.E.; Mihalache, C.; Daraba, O.-M.; Gherghel, D.; Popa, M. “In vitro” behaviour of aptamer-functionalized polymeric nanocapsules loaded with 5-fluorouracil for targeted therapy. Mater. Sci. Eng. C 2019, 103, 109828. [Google Scholar] [CrossRef]
- Aam, B.B.; Heggset, E.B.; Norberg, A.L.; Sørlie, M.; Vårum, K.M.; Eijsink, V.G. Production of Chitooligosaccharides and Their Potential Applications in Medicine. Marr. Drugs 2010, 8, 1482–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keong, L.C.; Halim, A.S. In Vitro Models in Biocompatibility Assessment for Biomedical-Grade Chitosan Derivatives in Wound Management. Int. J. Mol. Sci. 2009, 10, 1300–1313. [Google Scholar] [CrossRef] [Green Version]
- Kong, M.; Chen, X.G.; Xing, K.; Park, H.J. Antimicrobial properties of chitosan and mode of action: A state of the art review. Int. J. Food Microbiol. 2010, 144, 51–63. [Google Scholar] [CrossRef]
- Cassone, M.; Otvos, L., Jr. Synergy among Antibacterial Peptides and between Peptides and Small-Molecule Antibiotics. Expert. Rev. Anti Infect. Ther. 2010, 8, 703–716. [Google Scholar] [CrossRef] [PubMed]
- Duong, L.; Gross, S.P.; Siryaporn, A. Developing Antimicrobial Synergy with AMPs. Front. Med Technol. 2021, 3, 9. [Google Scholar] [CrossRef]
- Rahman, H.; Hjeljord, L.G.; Aam, B.B.; Sørlie, M.; Tronsmo, A. Antifungal effect of chito-oligosaccharides with different degrees of polymerization. Eur. J. Plant Pathol. 2015, 141, 147–158. [Google Scholar] [CrossRef]
- Hawser, S.P.; Norris, H.; Jessup, C.J.; Ghannoum, M.A. Comparison of a 2,3-Bis(2-Methoxy-4-Nitro-5-Sulfophenyl)-5-[(Phenylamino)Carbonyl]-2h-T Etrazolium Hydroxide (Xtt) Colorimetric Method with the Standardized National Committee for Clinical Laboratory Standards Method of Testing Clinical Yeast Isolates for Susceptibility to Antifungal Agents. J. Clin. Microbiol. 1998, 36, 1450–1452. [Google Scholar] [PubMed]
- White, R.L.; Burgess, D.S.; Manduru, M.; Bosso, J.A. Comparison of Three Different in Vitro Methods of Detecting Synergy: Time-Kill, Checkerboard, and E Test. Antimicrob. Agents Chemother. 1996, 40, 1914–1918. [Google Scholar] [CrossRef] [Green Version]
- Silva-Dias, A.; Palmeira-De-Oliveira, A.; Miranda, I.; Branco, J.; Cobrado, L.; Monteiro-Soares, M.; Queiroz, J.; Pina-Vaz, C.; Rodrigues, A.G. Anti-biofilm activity of low-molecular weight chitosan hydrogel against Candida species. Med Microbiol. Immunol. 2014, 203, 25–33. [Google Scholar] [CrossRef]
- Martinez, L.R.; Mihu, M.R.; Tar, M.; Cordero, R.J.; Han, G.; Friedman, A.J.; Friedman, J.M.; Nosanchuk, J.D. Demonstration of Antibiofilm and Antifungal Efficacy of Chitosan against Candidal Biofilms, Using an in Vivo Central Venous Catheter Model. J. Infect. Dis. 2010, 201, 1436–1440. [Google Scholar] [CrossRef] [Green Version]
- Carlson, R.P.; Taffs, R.; Davison, W.M.; Stewart, P.S. Anti-biofilm properties of chitosan-coated surfaces. J. Biomater. Sci. Polym. Ed. 2008, 19, 1035–1046. [Google Scholar] [CrossRef]
- Tan, H.; Peng, Z.; Li, Q.; Xu, X.; Guo, S.; Tang, T. The use of quaternised chitosan-loaded PMMA to inhibit biofilm formation and downregulate the virulence-associated gene expression of antibiotic-resistant staphylococcus. Biomaterials 2012, 33, 365–377. [Google Scholar] [CrossRef]
- Uppuluri, P.; Nett, J.; Heitman, J.; Andes, D. Synergistic Effect of Calcineurin Inhibitors and Fluconazole against Candida albicans Biofilms. Antimicrob. Agents Chemother. 2008, 52, 1127–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chauhan, A.; Lebeaux, D.; Ghigo, J.-M.; Beloin, C. Full and Broad-SpectrumIn VivoEradication of Catheter-Associated Biofilms Using Gentamicin-EDTA Antibiotic Lock Therapy. Antimicrob. Agents Chemother. 2012, 56, 6310–6318. [Google Scholar] [CrossRef] [Green Version]
- I Raad, I.; Fang, X.; Keutgen, X.M.; Jiang, Y.; Sherertz, R.; Hachem, R. The role of chelators in preventing biofilm formation and catheter-related bloodstream infections. Curr. Opin. Infect. Dis. 2008, 21, 385–392. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y. The chelating properties of chitosan fibers. J. Appl. Polym. Sci. 1993, 49, 727–731. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CA | Time (h) | CA MDC Alone | CA MDC in Presence of C32 | C32 MDC Alone | C32 MDC in Presence of CA | Detection of S or I | |
|---|---|---|---|---|---|---|---|
| C. albicans | Flu | 3 | >32.00 | 8.00 | 5000.00 | 625.00 | S |
| 24 | >32.00 | 8.00 | 2500.00 | 625.00 | S | ||
| 48 | >32.00 | 8.00 | 2500.00 | 625.00 | S | ||
| Mcz | 3 | >25.00 | 6.25 | 5000.00 | 625.00 | S | |
| 24 | >25.00 | 6.25 | 2500.00 | 625.00 | S | ||
| 48 | >25.00 | 6.25 | 2500.00 | 625.00 | S | ||
| C. glabrata | Mcz | 3 | >25.00 | 6.25 | >10,000.00 | 625.00 | S |
| 24 | >25.00 | 6.25 | 10,000.00 | 625.00 | S | ||
| 48 | >25.00 | 12.50 | 10,000.00 | 1250.00 | I |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ganan, M.; Lorentzen, S.B.; Gaustad, P.; Sørlie, M. Synergistic Antifungal Activity of Chito-Oligosaccharides and Commercial Antifungals on Biofilms of Clinical Candida Isolates. J. Fungi 2021, 7, 718. https://doi.org/10.3390/jof7090718
Ganan M, Lorentzen SB, Gaustad P, Sørlie M. Synergistic Antifungal Activity of Chito-Oligosaccharides and Commercial Antifungals on Biofilms of Clinical Candida Isolates. Journal of Fungi. 2021; 7(9):718. https://doi.org/10.3390/jof7090718
Chicago/Turabian StyleGanan, Monica, Silje B. Lorentzen, Peter Gaustad, and Morten Sørlie. 2021. "Synergistic Antifungal Activity of Chito-Oligosaccharides and Commercial Antifungals on Biofilms of Clinical Candida Isolates" Journal of Fungi 7, no. 9: 718. https://doi.org/10.3390/jof7090718
APA StyleGanan, M., Lorentzen, S. B., Gaustad, P., & Sørlie, M. (2021). Synergistic Antifungal Activity of Chito-Oligosaccharides and Commercial Antifungals on Biofilms of Clinical Candida Isolates. Journal of Fungi, 7(9), 718. https://doi.org/10.3390/jof7090718