
Two New Species of Backusella (Mucorales, Mucoromycota) from Soil in an Upland Forest in Northeastern Brazil with an Identification Key of Backusella from the Americas

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Site
2.2. Isolation, Purification, and Identification
2.3. Experiments
2.4. DNA Extraction, Amplification, Cloning, and Sequencing
2.5. Phylogenetic Analyses
3. Results
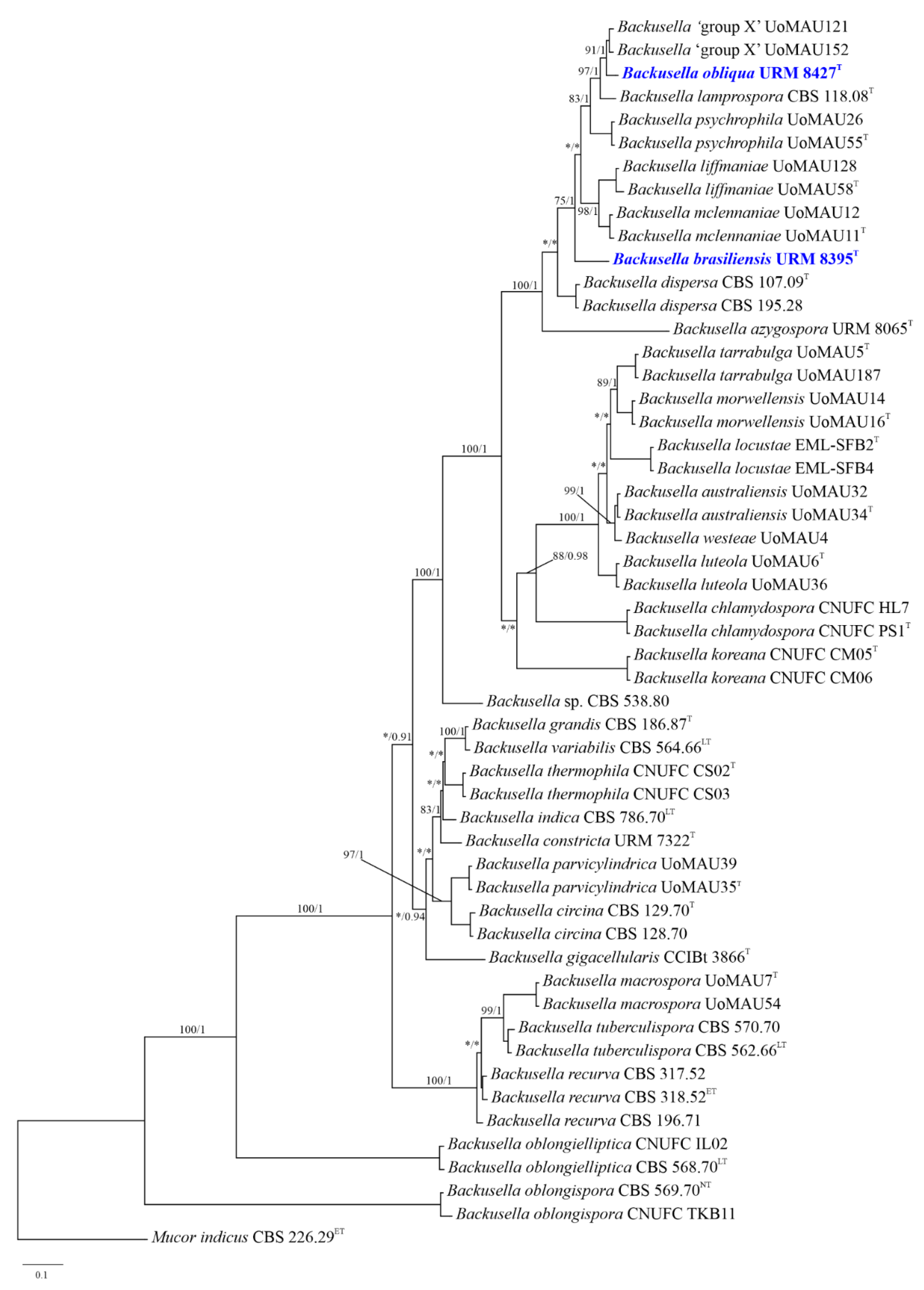
3.1. Phylogenetic Analyses
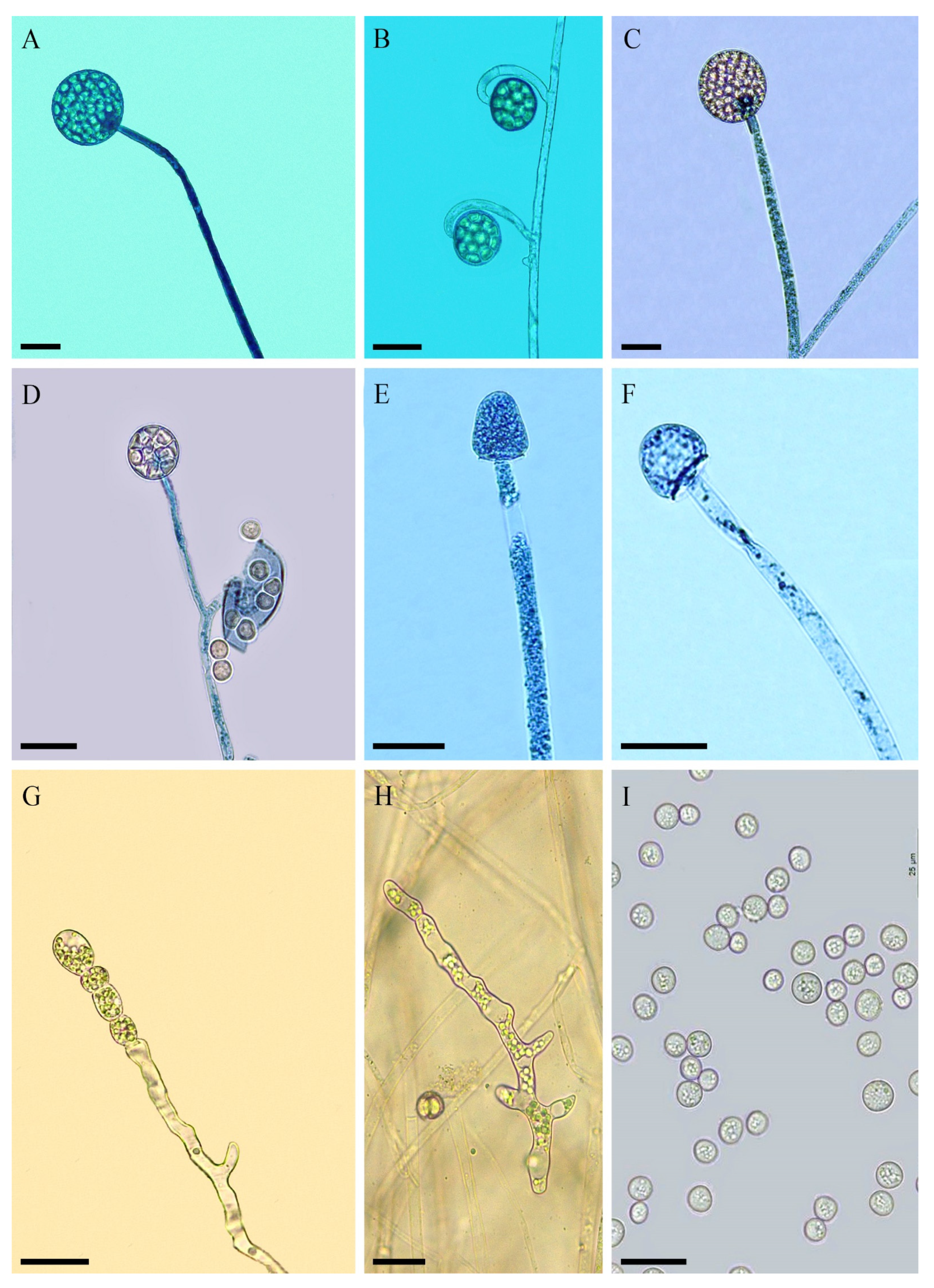
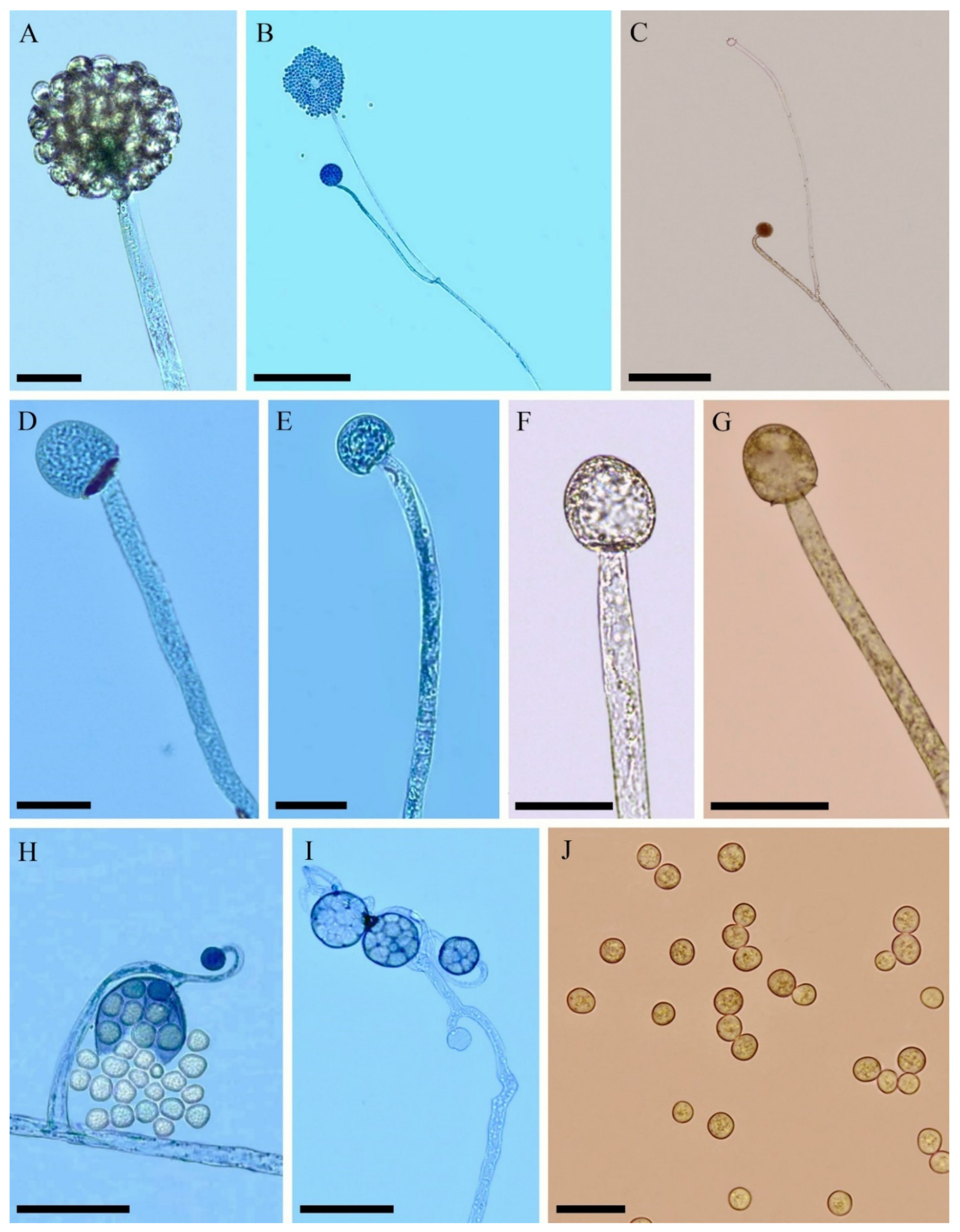
3.2. Taxonomy
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Benny, G.L. Zygomycetes. Available online: http://zygomycetes.org/index.php?id=27 (accessed on 25 May 2022).
- Walther, G.; Pawłowska, J.; Alastruey-Izquierdo, A.; Wrzosek, M.; Rodriguez-Tudela, J.L.; Dolatabadi, S.; Chakrabarti, A.; de Hoog, G.S. DNA barcoding in Mucorales: An inventory of biodiversity. Persoonia 2013, 30, 11–47. [Google Scholar] [CrossRef] [PubMed]
- Wanasinghe, D.N.; Phukhamsakda, C.; Hyde, K.D.; Jeewon, R.; Lee, H.B.; Jones, E.B.G.; Tibpromma, S.; Tennakoon, D.S.; Dissanayake, A.J.; Jayasiri, S.C.; et al. Fungal diversity notes 709–839: Taxonomic and phylogenetic contributions to fungal taxa with an emphasis on fungi on Rosaceae. Fungal Divers. 2018, 89, 1–236. [Google Scholar] [CrossRef]
- Nguyen, T.T.T.; Voigt, K.; Santiago, A.L.C.M.A.; Kirk, P.M.; Lee, H.B. Discovery of novel Backusella (Backusellaceae, Mucorales) isolated from invertebrates and toads in Cheongyang, Korea. J. Fungi 2021, 7, 513. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.J.; Hesseltine, C.W. Two new members of the Mucorales. Mycologia 1969, 61, 863–872. [Google Scholar] [CrossRef]
- Pidoplichko, N.M.; Milko, A.A. Atlas of Mucoralean Fungi; Academy of Sciences of the Ukrainian SSR: Kiev, Ukraine, 1971. [Google Scholar]
- Benny, G.L.; Benjamin, R.K. Observations on Thamnidiaceae (Mucorales). New taxa, new combinations, and notes on selected species. Aliso 1975, 8, 301–351. [Google Scholar] [CrossRef]
- Urquhart, A.S.; Douch, J.K.; Heafield, T.A.; Buddie, A.G.; Idnurm, A. Diversity of Backusella (Mucoromycotina) in south-eastern Australia revealed through polyphasic taxonomy. Persoonia 2021, 46, 1–25. [Google Scholar] [CrossRef]
- de Souza, J.I.; Marano, A.V.; Pires-Zottarelli, C.L.A.; Chambergo, F.S.; Harakava, R. A new species of Backusella (Mucorales) from a Cerrado reserve in Southeast Brazil. Mycol. Progr. 2014, 13, 981. [Google Scholar] [CrossRef]
- Lima, D.X.; Voigt, K.; de Souza, C.A.F.; de Oliveira, R.J.V.; Souza-Motta, C.M.; Santiago, A.L.C.M.A. Description of Backusella constricta sp. nov. (Mucorales, ex Zygomycota) from the Brazilian Atlantic Rainforest, including a key to species of Backusella. Phytotaxa 2016, 289, 059–068. [Google Scholar] [CrossRef]
- Crous, P.W.; Carnegie, A.J.; Wingfield, M.J.; Sharma, R.; Mughini, G.; Noordeloos, M.E.; Santini, A.; Shouche, Y.S.; Bezerra, J.D.P.; Dima, B.; et al. Fungal Planet description sheets: 868–950. Persoonia 2019, 42, 291–473. [Google Scholar] [CrossRef]
- Flora e Funga do Brasil. Jardim Botânico do Rio de Janeiro. Available online: http://floradobrasil.jbrj.gov.br/reflora/listaBrasil/PrincipalUC/PrincipalUC.do;jsessionid=D8E2833E5478AF52DD9209A15B77387F#CondicaoTaxonCP (accessed on 12 March 2022).
- Damasceno, M.L.; Pereira, J.A.S.; Schuler, C.A.B. Análise espaço temporal da cobertura vegetal do município de Arcoverde (Pernambuco). RBSR 2020, 1, 38–50. [Google Scholar]
- IBGE. Instituto Brasileiro de Geografia e Estatística. Available online: http://cod.ibge.gov.br/2AT (accessed on 14 April 2022).
- Departamento de Ciências Atmosféricas—DCA. Available online: http://www.dca.ufcg.edu.br/clima/dadospe.htm (accessed on 14 April 2022).
- Beltrão, B.A.; Mascarenhas, J.C.; Miranda, J.L.F.; Souza Junior, L.C.; Galvão, M.J.T.G.; Pereira, S.N. Projeto Cadastro de Fontes de Abastecimento por Água Subterrânea Estado de Pernambuco. Diagnóstico do Município de Águas Belas. Available online: http://rigeo.cprm.gov.br/xmlui/bitstream/handle/doc/15611 (accessed on 13 July 2022).
- Benny, G.L. The methods used by Dr. R.K. Benjamin, and other Mycologists to isolate Zygomycetes. Aliso 2008, 26, 37–61. [Google Scholar] [CrossRef]
- Kornerup, A.; Wanscher, J.H. Methuen Handbook of Colour; Eyre Methuen: London, UK, 1978. [Google Scholar]
- de Oliveira, R.J.V.; Bezerra, J.L.; Lima, T.E.F.; da Silva, G.A.; Cavalcanti, M.A.Q. Phaeosphaeria nodulispora, a new endophytic coelomycete isolated from tropical palm (Cocos nucifera) in Brazil. Nova Hedwigia 2016, 103, 185–192. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar]
- van Tuinen, D.; Zhao, B.; Gianinazzi-Pearson, V. PCR in studies of AM fungi: From primers to application. In Mycorrhizal Manual; Varma, A.K., Ed.; Springer: Berlin, Germany, 1998; pp. 387–399. [Google Scholar]
- Santiago, A.L.C.M.A.; Hoffmann, K.; Lima, D.X.; de Oliveira, R.J.V.; Malosso, E.; Maia, L.C.; Silva, G.A. A new species of Lichtheimia (Mucoromycotina, Mucorales) isolated from Brazilian soil. Mycol. Progr. 2014, 13, 343–352. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic trees. 2010 Gateway Computing Environments Workshop (GCE). New Orleans 2010, 1–8. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A. MrBayes 3.2: Efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed]
- Schipper, M.A.A. On certain species of Mucor with a key to all accepted species. Studies in Mycology 1978, 17, 1–69. [Google Scholar]
- Arnold, A.E.; Lutzoni, F. Diversity and host range of foliar fungal endophytes: Are tropical leaves biodiversity hotspots? Ecology 2007, 88, 541–549. [Google Scholar] [CrossRef]
- Carvalho, F.; Souza, F.A.; Carrenho, R.; Moreira, F.M.S.; Jesus, E.C.; Fernandes, G.W. The mosaic of habitats in the high-altitude Brazilian rupestrian fields is a hotspot for arbuscular mycorrhizal fungi. Appl. Soil Ecol. 2012, 52, 9–19. [Google Scholar] [CrossRef]
- Maia, L.C.; de Carvalho Júnior, A.A.; de H. Cavalcanti, L.; de M. Gugliotta, A.; Drechsler-Santos, E.R.; de A. Santiago, A.L.M.; da S. Cáceres, M.E.; Gibertoni, T.B.; Aptroot, A.; Giachini, A.J.; et al. Diversity of Brazilian Fungi. Rodriguésia 2015, 66, 1–13. [Google Scholar] [CrossRef]
- Voigt, K.; James, T.Y.; Kirk, P.M.; Santiago, A.L.; Waldman, B.; Griffith, G.W.; Fu, M.; Radek, R.; Strassert, J.F.; Wurzbacher, C.; et al. Early-diverging fungal phyla: Taxonomy, species concept, ecology, distribution, anthropogenic impact, and novel phylogenetic proposals. Fungal Divers. 2021, 109, 59–98. [Google Scholar] [CrossRef]
- de Melo Alves, A.L.S.; de Freitas, L.W.S.; Sartori Gurgel, L.M.; Alves Leitão, J.D.; Leite Cordeiro, T.R.; de Azevedo Santiago, A.L.C.M. First occurrence of Backusella gigacellularis (Mucorales, Mucoromycota) in a fragment of an Upland Forest within the semi-arid region of northeastern Brazil. Rodriguésia 2021, 72, e01262019. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Taxon Name | Collection No. | Country | GenBank Accession Number | |
|---|---|---|---|---|
| ITS | LSU | |||
| Backusella australiensis | UoMAU34 | Australia | MK959062 | MK958800 |
| Backusella australiensis | UoMAU32 | Australia | - | MK958802 |
| Backusella azygospora | URM 8065 | Brazil | MK625216 | MK625222 |
| Backusella brasiliensis | URM 8395 | Brazil | OM458082.1 | OM458083.1 |
| Backusella chlamydospora | CNUFC PS1 | South Korea | MZ171385 | MZ148709 |
| Backusella chlamydospora | CNUFC HL7 | South Korea | MZ171386 | MZ148710 |
| Backusella circina | CBS 128.70 | USA | JN206258 | JN206529 |
| Backusella circina | CBS 129.70 | USA | JN206257 | MH871299 |
| Backusella constricta | URM 7322 | Brazil | KT937158 | KT937156 |
| Backusella dispersa | CBS 107.09 | Norway | JN206269 | MH866118 |
| Backusella dispersa | CBS 195.28 | USA | JN206271 | JN206530 |
| Backusella gigacellularis | CCIBt 3866 | Brazil | KF742415 | - |
| Backusella grandis | CBS 186.87 | India | JN206252 | JN206527 |
| Backusella indica | CBS 786.70 | India | JN206255 | MH871743 |
| Backusella koreana | CNUFC CM05 | Korea | MZ171387 | MZ148711 |
| Backusella koreana | CNUFC CM06 | Korea | MZ171388 | MZ148712 |
| Backusella lamprospora | CBS 118.08 | Switzerland | NR_145291 | NG_058650 |
| Backusella liffmaniae | UoMAU128 | Australia | - | MK958735 |
| Backusella liffmaniae | UoMAU58 | Australia | MK959065 | MK958734 |
| Backusella luteola | UoMAU6 | Australia | MK959058 | MK958795 |
| Backusella luteola | UoMAU36 | Australia | - | MK958794 |
| Backusella locustae | EML-SFB4 | Korea | KY449293 | KY449290 |
| Backusella locustae | EML-SFB2 | Korea | KY449291 | KY449292 |
| Backusella macrospora | UoMAU7 | Australia | MK959107 | MK958628 |
| Backusella macrospora | UoMAU54 | Australia | - | MK958629 |
| Backusella mclennaniae | UoMAU12 | Australia | MK959087 | MK958777 |
| Backusella mclennaniae | UoMAU11 | Australia | MK959077 | MK958776 |
| Backusella morwellensis | UoMAU16 | Australia | MK959059 | MK958808 |
| Backusella morwellensis | UoMAU14 | Australia | - | MK958806 |
| Backusella obliqua | URM 8427 | Brazil | ON858475 | ON858467 |
| Backusella oblongielliptica | CBS 568.70 | Japan | JN206278 | JN206533 |
| Backusella oblongielliptica | CNUFC IL02 | Korea | MZ171391 | MZ148715 |
| Backusella oblongispora | CBS 569.70 | Japan | JN206251 | JN206407 |
| Backusella oblongispora | CNUFC TKB11 | Korea | MZ420786 | MZ148717 |
| Backusella parvicylindrica | UoMAU35 | Australia | MK959109 | MK958727 |
| Backusella parvicylindrica | UoMAU39 | Australia | - | MK958728 |
| Backusella psychrophila | UoMAU26 | Australia | - | MK958748 |
| Backusella psychrophila | UoMAU55 | Australia | MK959093 | MK958749 |
| Backusella recurva | CBS 318.52 | USA | JN206261 | JN206522 |
| Backusella recurva | CBS 317.52 | Macedonia | JN206262 | MH868593 |
| Backusella recurva | CBS 196.71 | - | JN206265 | JN206523 |
| Backusella tarrabulga | UoMAU187 | Australia | - | MK958805 |
| Backusella tarrabulga | UoMAU5 | Australia | MK959060 | MK958804 |
| Backusella thermophila | CNUFC CS02 | Korea | MZ171389 | MZ148713 |
| Backusella thermophila | CNUFC CS03 | Korea | MZ171390 | MZ148714 |
| Backusella tuberculispora | CBS 562.66 | India | JN206267 | JN206525 |
| Backusella tuberculispora | CBS 570.70 | Japan | JN206266 | MH871631 |
| Backusella variabilis | CBS 564.66 | India | JN206254 | JN206528 |
| Backusella westeae | UoMAU4 | Australia | MK959061 | MK958796 |
| Backusella sp. | CBS 538.80 | Egypt | HM999964 | HM849692 |
| Backusella ‘group X’ | UoMAU121 | Australia | MK959103 | MK958792 |
| Backusella ‘group X’ | UoMAU152 | Australia | MK959102 | MK958791 |
| Mucor indicus | CBS 226.29 | Switzerland | MH855050 | HM849690 |
| 1. Sporangiola formed | 2 |
| 1. Sporangiola not formed | B. oblongielliptica |
| 2. Unispored sporangiola abundant | B. circina |
| 2. Unispored sporangiola rare or not formed | 3 |
| 3. Giant cells formed | 4 |
| 3. Giant cells not formed | 5 |
| 4. Columellae of sporangia ellipsoidal, cylindrical, rarely pyriform; chlamydospores absent | B. gigacellularis |
| 4. Columellae of sporangia conical (majority), but also ellipsoidal with a truncate base, globose to subglobose, subglobose to conical, or, rarely, conical or cylindrical with slight constriction at the center; chlamydospores abundant | B. brasiliensis |
| 5. Azygospores formed | B. azygospora |
| 5. Azygospores not formed | 6 |
| 6. Sporangiospores ellipsoidal | 7 |
| 6. Sporangiospores not ellipsoidal | 8 |
| 7. Sporangia up to 150 (–200) μm diam.; sporangiospores 20–26 × 10–12 μm | B. recurva |
| 7. Sporangia up to 100 (–125) μm diam.; sporangiospores 11–15 × 7–9 μm | B. variabilis |
| 8. Sporangiophores forming a terminal sporangium and few lateral pedicellate sporangiola | B. lamprospora |
| 8. Sporangiophores forming a terminal sporangium with no lateral pedicellate sporangiola | 9 |
| 9. Columellae of sporangia of varied shapes, some arranged obliquely on the sporangiophores, some with one side more swollen than the other; sporangiospores globose to subglobose | B. obliqua |
| Columellae of sporangia conical and cylindrical, sometimes constricted at the center, never arranged obliquely on the sporangiophores nor with one side more swollen than the other; sporangiospores subglobose to ellipsoidal, some slightly irregular | B. constricta |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Lima, C.L.F.; Lundgren, J.D.A.L.; Nguyen, T.T.T.; Cordeiro, T.R.L.; Lima, D.X.; Gurgel, L.M.S.; da Costa, D.P.; Lee, H.B.; Santiago, A.L.C.M.d.A. Two New Species of Backusella (Mucorales, Mucoromycota) from Soil in an Upland Forest in Northeastern Brazil with an Identification Key of Backusella from the Americas. J. Fungi 2022, 8, 1038. https://doi.org/10.3390/jof8101038
de Lima CLF, Lundgren JDAL, Nguyen TTT, Cordeiro TRL, Lima DX, Gurgel LMS, da Costa DP, Lee HB, Santiago ALCMdA. Two New Species of Backusella (Mucorales, Mucoromycota) from Soil in an Upland Forest in Northeastern Brazil with an Identification Key of Backusella from the Americas. Journal of Fungi. 2022; 8(10):1038. https://doi.org/10.3390/jof8101038
Chicago/Turabian Stylede Lima, Catarina Letícia Ferreira, Joana D’arc Alves Leitão Lundgren, Thuong Thuong Thi Nguyen, Thalline Rafhaella Leite Cordeiro, Diogo Xavier Lima, Luciana Melo Sartori Gurgel, Diogo Paes da Costa, Hyang Burm Lee, and André Luiz Cabral Monteiro de Azevedo Santiago. 2022. "Two New Species of Backusella (Mucorales, Mucoromycota) from Soil in an Upland Forest in Northeastern Brazil with an Identification Key of Backusella from the Americas" Journal of Fungi 8, no. 10: 1038. https://doi.org/10.3390/jof8101038