Abstract
Photosynthesis is essential for the growth of all green plants, and the presence of an Epichloë endophyte enhances the photosynthesis of Achnatherum inebrians (drunken horse grass, DHG), including when it is under attack by fungal pathogens. However, few studies have examined the mechanism of the increased photosynthetic activity at the molecular level of A. inebrians when it is under pathogen stress. The present study investigated the effects of the presence of the Epichloë endophyte on the net photosynthetic rate, intercellular CO2 concentration, stomatal conductance, and transpiration rate of DHG plants under a Blumeria graminis infection condition, and we compared the transcriptomes using RNA sequencing. The results showed that the photosynthetic rate of Epichloë endophyte-infected (E+) plants was higher under the B. graminis infection condition, and also without this pathogen, when it was compared with Epichloë endophyte-free (E-) plants. The E+ plants uninfected with B. graminis had 15 up-regulated unigenes that are involved in photosynthesis which were compared to the E- plants that were uninfected with this pathogen. This suggests that the presence of an Epichloë endophyte up-regulates the genes that are involved in the process of photosynthesis.
1. Introduction
Plants living in complex environments interact closely with a wide range of microbes, including pathogens with different lifestyles and infection processes, and this, the plant’s health is challenged [1]. Blumeria graminis is a fungus of the Erysiphaceae, with a wide range of hosts, including 634 Poaceae species. The host grasses include important grain species such as wheat, barley, rye and oats [2]. Blumeria graminis, the cause of powdery mildew, is considered to be the sixth most important fungal phytopathogen in the world [3], infecting the leaves, stems, flowers and fruits of plants. This pathogen causes severe economic losses in food and pasture production [4]. Blumeria graminis is a strictly obligate biotrophic fungus [5,6], whose conidia, which are formed in fragile chains on plant surfaces, are spread by the wind. The germ tubes of germinating conidia form appressoria on the surface of the leaves from which infection pegs penetrate the wall of the epidermal cells and form haustoria that invaginate the plasmalemma within the penetrated cells. The haustoria enable the extraction of water and nutrients, without killing the penetrated epidermal cells. The export of the effector molecules to the host suppresses the immune response of the plant and accelerates the infection cycle [7], rapidly producing large numbers of conidia to efficiently complete multiple infections [7,8]. The infection process of B. graminis is ongoing and may lead to the extensive colonization of the host plants. This process not only absorbs a large amount of nutrients from the host, but it also seriously reduces the photosynthesis rate of the host plant [9,10], enhances respiration and transpiration, but significantly reduces the chlorophyll content, ultimately leading to a reduction in host plant’s carbon assimilation. Therefore, the growth and development of plants are hindered, and the yield is reduced. In severe cases, powdery mildew can lead to the death of the host plant [9,11,12].
Epichloë fungi form symbiotic, typically mutualistic relationships with nearly 30% of cool-season grasses across the globe [13,14,15]. Poaceae and Epichloë endophytes have evolved to form host-fungi-specific relationships that confer protection for the grass against various stresses [16,17,18], and in exchange, the plant provides the fungal endophyte with an ongoing supply of nutrients, a stable protected habitat, and transmission to the next generation through the seed of host grasses. An important characteristic of the relationship between the Epichloë endophytes and host grasses is that the growth of the two symbionts is fully synchronized [19]. E. gansuensis [20] or E. inebrians [21] is present in nearly 100% of Achnatherum inebrians plants (drunken horse grass, DHG), which is a perennial grass growing in natural grasslands in the alpine and sub-alpine regions in Northwestern China [20,22,23,24,25]. The association between DHG and E. gansuensis provides enhanced persistence, in part due to the production of alkaloids that cause livestock poisoning (ergonovine and ergine), and the deterrence of grazing [26,27,28]. It is the effect of these alkaloids that has given this grass the common name of drunken horse grass. An Epichloë endophyte can improve the tolerance of DHG to drought [29,30,31], salt [32], cold [33], heavy metal stress [34], pests [35] and pathogens [36,37,38].
Photosynthesis is the most fundamental metabolic activity in all green plants, and it plays an important role in the growth and development of plants. It is through photosynthesis that plants can produce and store nutrients. Photosynthesis is severely affected in all of its phases by different stresses. Since the mechanism of photosynthesis involves various components, including photosynthetic pigments and photosystems, the electron transport system, and the CO2 reduction pathways, damage at any level caused by a stress may reduce the overall photosynthetic capacity of a green plant [39,40,41]. Both abiotic [31,42] and biotic [37,43,44,45] stresses can influence the photosynthesis of plants. Under drought and flooding stress, the growth of the plants will be hindered, and the photosynthetic capacity of the plants will be reduced [31,42]. Under pathogenic stress, the photosynthesis of the plants is altered. Leaf pathogens reduce the photosynthetic parameters of the diseased leaves, including by reducing the green leaf area of the leaves [37,46,47]. Leaf rust pathogens, obligate biotrophic fungi, colonize the leaves, forming pustules, reducing the green leaf area of the plant and the chlorophyll content, thereby reducing the photosynthetic activity in the infected leaves [43]. The net photosynthetic rate of the light saturation is reduced by the reduced total photosynthesis [44]. The chlorophyll content and net photosynthetic rate of DHG were significantly reduced under the stress of ergot disease [45]. Under the infection of B. graminis, the chlorophyll content, the net photosynthetic rate, and the dry matter content of DHG were significantly reduced, but the intercellular carbon dioxide concentration was enhanced [37]. However, when an Epichloë endophyte is present in stressed plants, this stress is mitigated.
Xia et al. (2016) found that although powdery mildew inhibits the photosynthetic parameters of DHG, the presence of the Epichloë endophyte can reduce the damage caused by B. graminis [37]. Zhang et al. (2022) also found that the Epichloë endophyte significantly increased the chlorophyll content and decreased the intercellular carbon dioxide concentration of DHG under ergot disease stress conditions, thereby increasing the net photosynthetic rate of DHG under ergot stress conditions [45]. Researchers have found that the presence of an Epichloë endophyte enhances the resistance of ryegrass that is inoculated with the pathogens Alternaria alternata, Ascochyta leptospora, Curvularia lunata and Fusarium avenaceum [48]. Under drought stress conditions, the presence of an Epichloë endophyte increased the growth, biomass, and photosynthetic rate of DHG. At the transcriptional level, compared with DHG without an Epichloë endophyte, in DHG plants with an Epichloë endophyte, the participation of the genes involved in photosynthesis are mostly up-regulated, such as PetH, Lhcb1 and PsbQ, and these genes are also up-regulated under drought conditions [31]. Ambrose and Belanger (2012) also noted that 13% of genes up-regulated in the presence of an Epichloë endophyte were involved in the photosynthesis process of red fescue (Festuca rubra) [49]. However, in recent years, the research on the mechanism of the stress that is conferred by pathogens of DHG has mainly focused on the physiological and biochemical aspects, and there is little information at the gene level of the photosynthetic response of DHG under pathogen stress conditions.
Therefore, this study mainly explored the changes of the genes involved in the photosynthetic process of DHG (Epichloë endophyte-infected, E+; Epichloë endophyte-free, E-) under B. graminis infection. To test our predictions, we measured the photosynthetic indicators in the E+ and E- DHG plants and identified specific photosynthesis-related genes that are associated with changes in the photosynthetic rates. The aim of this study was to determine the changes in photosynthetic indexes and the related genes that are involved in photosynthetic processes in Epichloë-mediated DHG plants that are infested with B. graminis.
2. Materials and Methods
2.1. Plant Material and Experimental Design
The seeds of A. inebrians plants host to the E. gansuensis endophyte were generated in 2011 from plants originating in a natural grassland with 100% Epichloë infection rate in the Sunan county of the Gansu province of China. Before sowing, the collected seeds were divided into two parts, and one part was treated with thiophanate-methyl fungicide to ensure that the seeds were free of a viable Epichloë endophyte; the other part was not treated. These two seed types were planted in an experimental field of the College of Pastoral Agriculture Science and Technology, Yuzhong campus of Lanzhou University (104°39′ E, 35°89′ N, Altitude 1653 m) in 2012 [36]. The seed samples harvested from plants of the treated and untreated plants were stained with aniline blue and microscopically examined to determine the Epichloë endophyte status and ensure that the seeds with 100% and 0% infections were used in this study.
From 14 October to 14 December 2021, a pot experiment was conducted in the greenhouse of the College of Pastoral Agriculture Science and Technology, Yuzhong campus of Lanzhou University. On 14 October, 3 plump E+ and E- seeds were sown into each of the 240 pots (120 pots E+ and 120 pots E-, diameter: 24 cm; height: 15 cm), which were filled with vermiculite (75 g) that had been sterilized in an oven at 190 °C for 2 h. The pots were randomly placed in the constant temperature greenhouse (temperature: 26 ± 2 °C, humidity: 42% ± 2%) and watered sufficiently until the surface of the vermiculite was moist. After the emergence of the second fully expanded leaf, Hoagland’s solution was applied every 7 days.
The inoculation of the DHG plants with the pathogen B. graminis took place when the inflorescences had emerged (from the above 240 pots, 200 pots with consistent and healthy growth plants were selected for the treatment). The inoculation process was carried out by smearing 5 mL of a suspension of B. graminis at a concentration of 2 × 106 spores per mL on the 50 E+ and 50 E- plants (pathogen-inoculated plants, P+ plants). Using a sterile brush, the spore suspension of B. graminis that was obtained above, was applied evenly on the surface of each leaf of DHG from tip to base. In the control group, the leaves of the 50 E+ and E- DHG plants were smeared with 5 mL of sterilized water (non-inoculated plants, P- plants). After the inoculation, the pathogen-inoculated and non-inoculated plants were kept in two separate greenhouse compartments under the same growth conditions. The plants were monitored for four weeks after the pathogen inoculation.
2.2. Measurement of Photosynthetic Data
On the morning of 11 January (four weeks after the pathogen inoculation), the photosynthetic indexes (including the net photosynthetic rate, the intercellular carbon dioxide concentration, the transpiration rate and the stomatal conductance) of the plants were measured using a photosynthesis instrument (L1-6400, LI-COR Nebraska, Lincoln, NE, USA). Ten pots with consistent levels of B. graminis infection were selected from each treatment, and three leaves from the same growth position were selected from each pot, with the top of each leaf being measured three times [31,37].
2.3. RNA Extraction and Transcriptome Analysis
At four weeks, three fresh leaves from each of three pots of the E+ and E- DHG plants that had been inoculated with B. graminis, and also, three pots of E+ and E- DHG plants that had not been inoculated were collected and stored in liquid nitrogen at −80 °C for the subsequent RNA extraction and transcriptome sequencing. The total RNA, 3 μg per sample, was extracted using the TRIzol reagent (Invitrogen, Carlsbad, CA, USA). The RNA extraction quality and integrity were assessed by 1% agarose gel electrophoresis and an Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). The mRNA-Seq Library was constructed using Illumina®’s NEBNext® UltraTM RNA Library Prep Kit. The RNA sequencing was performed by the Biomarker Technologies Company (Beijing, China) [38].
High-quality clean reads (=90.54 Gb) were obtained from the raw reads by removing sequences containing adapters, poly-N, and low-quality sequences. The obtained unigene sequences (in FASTQ format) were used to query specific functions in the following databases: NR (NCBI non-redundant protein sequences); Pfam (Protein family); KOG/COG/eggnog (Clusters of Orthologous Groups of proteins); Swiss-Prot (a manually annotated and reviewed protein sequence database); KEGG (Kyoto Encyclopedia of Genes and Genomes); GO (Gene Ontology). The FPKM (Fragments Per Kilobase Million mapped reads) was calculated to estimate the expression level of all of the genes in each sample. The differentially expressed genes (DEGs) (FDR (false discovery rate) < 0.05 and |Log2fold change| ≥ 2) were analyzed using DEseq2_EBseq. Gene ontology (GO), enrichment, and the functional annotation of DEGs were implemented using the GOseq R package. The DEGs were further subjected to a Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis to identify the main biochemical metabolic pathways and signal transduction pathways.
2.4. Meta-Analysis Data Collection
The papers in journals that had been published up to 10 March 2022 were collected from the China National Knowledge Infrastructure (CNKI) and Web of Science databases. The search keywords were “Epichloë/endophyte/pathogens/photosynthesis” and “Grass endophytes, pathogens and photosynthesis” [50]. After multiple screenings, 46 articles were finally screened for the meta-analysis of this study, including 11 English articles and 35 Chinese articles (the references are attached to the supplementary materials).
GetaData Graph Digitizer 2.22 (http://getdata-graph-digitizer.com/, first access: 10 October 2021) was used to obtain the mean, standard deviation (if the standard error could be calculated according to the following formula) and the number of repetitions (n) on the graph of the net photosynthetic rate in the above literature; the data in the table were obtained directly. If the standard deviation could not be obtained directly, it was estimated according to 10% of the mean.
where n represents the number of repetitions of each treatment.
A total of 109 sets of observations were obtained from the selected 46 publications, and the obtained data were analyzed according to the following categories:
1. The net photosynthetic rate under the plant treatment with the Epichloë endophyte was selected and grouped according to different grass species. E+ (Epichloë-infected) was the experimental group; E- (Epichloë-free) was the control group.
2. The net photosynthetic rate under the treatment of plant pathogenic fungi was selected and grouped into E+ and E- groups. Pi+ (pathogen–inoculated, pathogens include Blumeria graminis and ergot) was the experimental group, and Pi- (non-inoculated) was the control group.
MetaWin software (version 2.1) was used to perform a meta-analysis to calculate these effects using the log of the response ratio (lnR) using the following Equation according to another study [51].
where and represent the mean of the experimental group and the control group, respectively.
The variance of the natural logarithm of the response ratio was approximated by the following equation:
and indicate the repetitions of the experimental and control groups, respectively, and and indicate the standard deviations of the experimental and control groups, respectively [52]. The mean of the effect of the Epichloë/pathogen infection on the net photosynthetic rate of host grasses were calculated using the combination of and , where the for each observation was weighted by the inverse variance of the . The 95% bootstrap confidence intervals (95% CI) for the mean were calculated using 9999 iterations of bootstrapping, according to a previous study [52].
2.5. Statistical Analyses
The differences in the photosynthetic indexes of the plants under the Epichloë endophyte and Blumeria graminis infestation conditions were tested using a two-way analysis of variance (ANOVA) using the datarium package of R software. A statistically significant two-way interaction was followed up by simple main effect analyses. All of the values are means ± SE of the mean.
3. Results
3.1. Plant Growth and Biomass
The Epichloë endophytes and Blumeria graminis had significant effects (p < 0.05) on the plant height, dry weight and fresh weight values of A. inebrians. In addition, there was a significant interaction (p < 0.05) between A. inebrians and B. graminis regarding the plant’s height (Table S1). However, there was no significant interaction between Epichloë and B. graminis for the tiller number, dry weight and fresh weight values (Table S1). The presence of the Epichloë endophytes significantly (p < 0.05) increased the plant height (Figure S1a), dry weight (Figure S1c) and fresh weight (Figure S1d) values. However, the inoculation with B. graminis significantly (p < 0.05) reduced the dry weight (Figure S1c) and fresh weight (Figure S1d) values of A. inebrians. Comparing E+ and E-plants that were infected by B. graminis, the presence of the Epichloë endophytes significantly (p < 0.05) increased the plant height of A. inebrians (Figure S1a).
3.2. Photosynthesis Index
The Epichloë endophytes and Blumeria graminis had significant effects (p < 0.05) on net photosynthetic rate, intercellular CO2 concentration, transpiration rate and stomatal conductance of A. inebrians. In addition, there was significant (p < 0.05) interaction between the endophyte and B. graminis regarding the intercellular CO2 concentration (Table 1). However, there were no significant interactions between the endophyte and B. graminis for net photosynthetic rate, transpiration rate and stomatal conductance (Table 1). The presence of the Epichloë endophyte significantly (p < 0.05) enhanced the net photosynthetic rate (Figure 1a), transpiration rate (Figure 1c) and stomatal conductance (Figure 1d) of A. inebrians. However, the inoculation with B. graminis significantly (p < 0.05) reduced the net photosynthetic rate, transpiration rate and stomatal conductance of A. inebrians. Comparing the E+ and E- plants that were infected by B. graminis, the presence of the Epichloë endophyte significantly (p < 0.05) decreased the intercellular CO2 concentration of A. inebrians (Figure 1b).

Table 1.
Two-way ANOVA for the effects of Blumeria graminis (P) and Epichloë endophyte (E) on the photosynthetic rate, intercellular CO2 concentration, stomatal conductance and transpiration rate of Achnatherum inebrians.
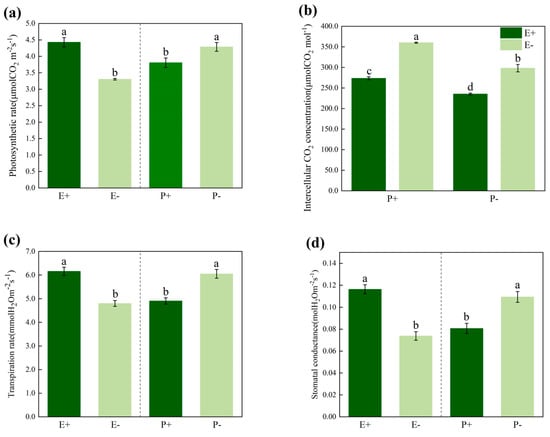
Figure 1.
Net photosynthetic rate (a), intercellular CO2 concentration (b) transpiration rate (c) and stomatal conductance (d) of E+ (Epichloë-infected) and E- (Epichloë-free) Achnatherum inebrians plants under the P+ (pathogen-inoculated) and P- (non-inoculated). Values are mean ± standard error (SE), with bars indicating SE. Columns with non-matching letters indicate a significant difference at p < 0.05.
3.3. Kegg Pathway Enrichment Analysis of the DEGs
A large number of differential genes were detected in this analysis, and these genes were mainly enriched in Glutathione metabolism (ko00480), Photosynthesis-antenna proteins (ko00196), Photosynthesis (ko00195), Prophyrin and chlorophyll metabolism (ko00860) (Figure 2). The photosynthetic genes were enriched in the photosynthetic pathway and in the antenna protein pathway. In this analysis, a total of 22 DEGs were detected in the Photosynthesis and Photosynthesis-antenna proteins.
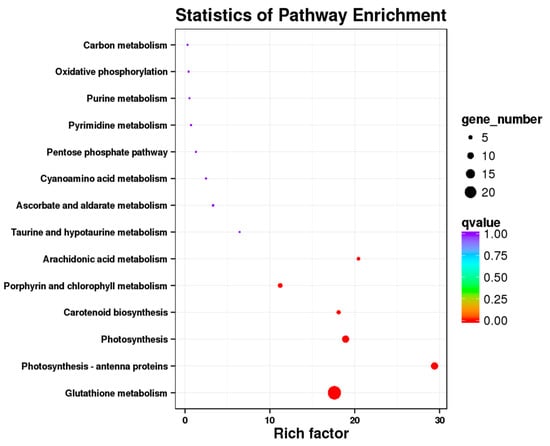
Figure 2.
KEGG pathway enrichment of DEGs associated with Achnatherum inebrians under the E+ (Epichloë-infected), P+ (pathogen-inoculated) and both fungi (E+P+). Only the top 20 most strongly represented pathways are displayed in the diagram. The q-value ranges from 0 to 1, and a q-value that is closer to 0 indicates a greater enrichment.
3.4. DEGs of Regulatory Pathways Related to Photosynthesis
Single genes with fold changes in the gene expression that were greater than or equal to two and FDR values that were below 0.05 were defined as DEGs by a comparison with the E-P- treatment. Based on these stringent criteria, there were seventeen (fifteen up and two down) DEGs between E-P- and E+P-, five (two up and three down) DEGs between E-P- and E+P+, and between E-P- and E-P+, there was one (one up-regulated and zero down-regulated) DEG among the different overlapping differential genes under each treatment (Figure S2).Twenty-two DEGs were identified in the Photosynthesis and Photosynthesis-antenna proteins pathways, and these DEGs were found to be associated with the photosystem, membrane protein complex, photosynthetic electron transport chain, and antenna proteins (Figure 3 and Table S2). The presence of the Epichloë endophyte up-regulated fifteen unigenes and down-regulated two unigenes. Among these, the levels of four genes encoding the PSI protein complex, including PsaD, PsaE, PsaG and PsaH, as well as the PSII protein complex (PsbR) were improved in response to the presence of the Epichloë endophyte. Furthermore, nine unigenes encoding chlorophyII b binding protein Lhcb1, Lhcb2 and Lhcb6 of the light-harvesting complex were also observed to be up-regulated. In addition, one gene encoding the PetE protein involved in photosynthetic electron transport was down-regulated, whereas two unigenes encoding PetH were down-regulated. E- plants in the presence of B. graminis had one up-regulated gene; the unigene encoding the PetF protein that is involved in photosynthetic electron transport was up-regulated. E+ plants in the presence of B. graminis had two up-regulated genes, and three down-regulated genes. One gene encoding subunits of the cytochrome b6/f complex (PetA) and one unigene encoding the beta subunits of the F-type H+-transporting ATP synthase complex were up-regulated, while three genes encoding chlorophyll a/b binding protein Lhca4, Lhca2 and Lhca5 of the light-harvesting complex were also observed to be down-regulated (Figure 3).
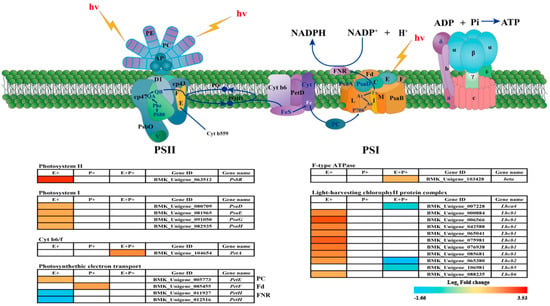
Figure 3.
Photosynthesis-related DEGs in E+ (Epichloë-infected), P+ (pathogen-inoculated) and both fungi (E+P+) on Achnatherum inebrians plants. Red and blue colors represent the up-regulated and down-regulated proteins, respectively. Gene IDs and gene names are shown.
3.5. Go Functional Enrichment Analysis of the DEGs
There are 18 GO categories in the photosynthetic pathway (Figure 4a), and 17 GO categories in the antenna protein pathway (Figure 4b). There are a total of 22 DEGs in the photosynthetic and antenna protein pathways, and the detected genes were mainly divided into three categories: biological process, cellular component and molecular function. In the category of the biological processes, metabolic processes dominated, which was followed by cellular processes. In the category of the cellular components, cells, cell parts, organelles and membranes were predominant, which were followed by the organelle parts. In the molecular functional class, binding is the most important group, which is followed by catalytic activity (Figure 4).
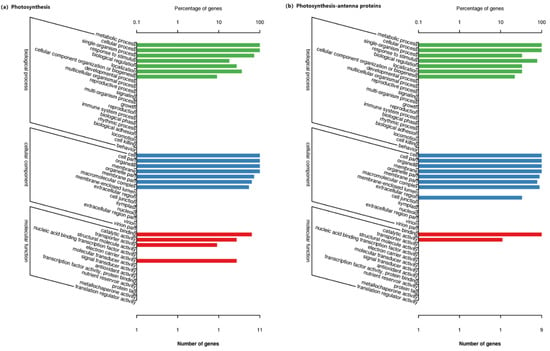
Figure 4.
GO annotations of differentially expressed genes on the photosynthesis (a) and antenna protein (b) pathways of Achnatherum inebrians under each treatment.
3.6. Meta-Analysis
Based on the current research on the effect of the Epichloë endophyte on the net photosynthetic efficiency (NPE) of the host grass, the data of the host grass under normal growth conditions were selected for a meta-analysis, and there was an overall positive effect of the Epichloë endophyte on NPE of E+ plants which was observed when it was compared to E- plants (Qb = 153.3, p = 0.000, df = 9) (Figure 5a). The higher NPE values were observed in the A. inebrians, Elymus tangutorum, Festuca arundinacea, F. sinensis, Calamagrostis epigeios, L. perenne plants hosts of an Epichloë endophyte (Figure 5a and Table S3), and among them, the F. sinensis plants had the highest NPEs (effect size = −0.329, 95% CI = 0.119 to 0.540). However, the presence of an Epichloë endophyte was found to reduce the NPE of the host plants in the A. sibiricum, F. arizonica and Stipa purpurea plants (Figure 5a and Table S3).
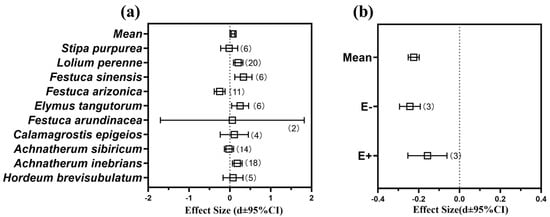
Figure 5.
Effects on net photosynthetic efficiency (NPE) of grasses under the infestation of corresponding pathogens and Epichloë endophyte. (a) Effects of symbiotic Epichloë fungal endophyte on NPE of grasses (Relative effects of Epichloë symbiotic versus non-symbiotic plants in normal growth conditions) (b) Effects of inoculation with pathogens (include Blumeria graminis and ergot) on NPE of E+ (Epichloë-infected) and E- (Epichloë-free) grasses. Note: Error bars represent means ± bootstrap 95% CI. Note: The corresponding effect size and 95% CI are attached to the Supplementary Material Tables S3 and S4.
There was an overall restrictive effect (main effect size = −0.225, 95% CI = from −0.251 to −0.198) of the phytopathogenic fungi on the NPE of the E- plants when they were compared to the E+ plants (Qb = 22.1, p = 0.000, df = 5) (Figure 5b). However, the NPE of the E+ plants were higher than they were in the E- plants under pathogenic fungal infestation condition (Figure 5b and Table S4).
4. Discussion
In the present study, transcriptomics was used to investigate the effect of the interaction between the Epichloë endophyte and the fungal pathogen B. graminis on the photosynthesis of DHG. The results showed that B. graminis weakened the photosynthesis of DHG, while the presence of Epichloë endophyte alleviated the damage of B. graminis on photosynthesis. Our study also indicated that the presence of an Epichloë endophyte upregulates some genes that are involved in the photosynthesis process.
4.1. Effects of Biotic and Abiotic Stresses on Photosynthesis
Our current study, which was conducted in the absence of an abiotic stress, we found that the B. graminis infection significantly reduced the photosynthetic rate, stomatal conductance and transpiration rate of the E+ and E- DHG plants, but the photosynthetic index of E+ plants was higher than it was in the E- plants (Figure 1). This is consistent with the findings of Xia et al. (2016) wherein the photosynthesis of DHG was inhibited both under the conditions of the artificial inoculation with B. graminis and the natural occurrence of powdery mildew [37]. Both biotic and abiotic stresses have been shown to reduce the photosynthetic rate by limiting the stomata or non-stomatal conductance [53,54], reducing plant growth and productivity [36]. For example, mild drought stress affects the photosynthesis, stomatal conductance, and water use efficiency in plant leaves [55,56,57,58]. Photosynthesis has a substantial contribution to the plant’s growth and development, and the chemical energy that is consumed in a wide range of metabolic processes in plants is mainly derived from photosynthesis. All green plants are capable of converting light energy into available chemical energy through photosynthesis [41]. However, the activation of the resistance responses in plants interacting with pathogens results in reduced crop yields and reduced photosynthetic parameters [46,47] when they are compared to uninfected plants. For example, leaves that are infected by B. graminis reduce the photosynthetic parameters of the plant, mainly by changing the stomatal conductance of the leaves and the concentration of carbon dioxide between the cells [59,60], which ultimately reduces the yield of the host plants [11,12].
4.2. Effects of Epichloë endophyte on Photosynthesis
As we expected, the results of the present study indicate that the Epichloë endophyte significantly enhanced the photosynthetic rate of DHG in both pathogen-infected and non-infected plants (Figure 1a). Another important finding of this study was that the Epichloë endophyte is an important indicator of altered photosynthesis in DHG (Table 1 and Figure 1). The largest number of DEGs related to photosynthesis were altered in the E+P- plants (Figure 3), and the E+ DHG plants that were not inoculated with B. graminis had higher growth rates than the non-inoculated E- DHG plants did (Table S1 and Figure S1). The presence of the Epichloë endophyte, growing intercellularly and synchronously with the surrounding tissue, from which it obtains all of its nutrients, should be a cost to the host plant [35,36,61]. However, the plant height, dry weight and fresh weight values were higher in the E+ DHG plants than they were in the E- plants in the heat-sterilized vermiculite, so the E+ DHG plants grew better than the E- plants did (Figure S1). The increased rate of photosynthesis due to the presence of the Epichloë endophyte not only compensated for the cost incurred by the presence of the systemic fungus, but it provided sufficient available nutrients to enhance the plant’s growth. Previous studies have demonstrated that plants that are symbiotic with the Epichloë endophyte have a higher biotic and abiotic tolerance [17,62,63,64]. The effects of biotic and abiotic stress on plant photosynthesis were mainly verified by measuring the photosynthesis indicators including the photosynthetic rate and the stomatal conductance. Meanwhile, the presence of an Epichloë endophyte improved the biomass and photosynthetic rate of DHG under different water treatments [29,31]. The current meta-analysis results indicate that the presence of an Epichloë endophyte can enhance the photosynthesis of the host grasses under pathogen stress conditions (Figure 5b). The results of meta-analysis also validate this point of view: the combination of plants with beneficial symbionts, including endophyte fungi, can alter the plant’s immune responses [65] and induce the plant defenses that enhance plant’s resistance to pathogens [66].
4.3. Presence of Epichloë endophyte Alters Photosynthetic Genes
Consistent with our hypothesis, the Epichloë endophyte can promote the photosynthesis of DHG by upregulating 17 genes encoding PSI, PSII, LHCI, LHCII, Cytochrome b6/f and ATP synthase (Figure 3). In this current study, 15 photosynthetic genes were up-regulated in E+P- DHG (Figure S2a). Unexpectedly, the expression level of the PetH gene encoding FNR in photosynthetic electron transport decreased in E+P- DHG (Figure 3), which blocked the electron transport process and inhibited photosynthesis. FNR is the terminal oxidase of the photosynthetic electron transport chain which receives electrons from Fd and H+ in the substrate and reduces NADP+ to NADPH. Similarly, the gene expression of the PetH protein involved in photoelectron transport was also down-regulated in Dunaliella salina under copper stress conditions [67].
Moreover, the abundance of light-harvesting chlorophyll of LHCI, LHCII protein in the E+ plants is higher than that in the E- plants, and the photosynthetic rate was higher in the E+ ones than it was in the E- plants (Figure 1a, Figure 3). Rozpądek et al. (2015) also noted that the presence of an Epichloë endophyte can improve the photosynthetic rate of Dactylis glomerata, and the abundance of the LHCI, LHCII protein in the E+ plants is higher than that in the E- plants [68]. The chlorophyll protein complex (LHC) plays an important role in the light harvesting of PSI and PSII, affecting the capture and transmission of light energy in the chloroplasts [69].
However, both chlorophyll complex protein I (Lhca4) and chlorophyll complex protein II (Lhcb2, 5) were down-regulated in E+P+ DHG (Figure 3). It may be that the presence of an Epichloë endophyte can only alleviate the pathogen stress to a certain extent, but this may not be a long-term effect. In this current study, the presence of an Epichloë endophyte also up-regulated the unigenes PsbR, PsaD, PsaE, PsaG and PsaH (Figure 3). PsaD, PsaE, PsaG and PsaH are subunits of the PSI-LHCI supercomplex, which are involved in light harvesting and electron transfer from plastocyanin (PC) to ferredoxin (Fd) [70,71].
The presence of the Epichloë endophyte is thought to shift the plant’s metabolism from defense to growth states, altering host transcript levels [33,68], enabling plants to efficiently use available resources for photosynthesis and carbon assimilation, and this enables the plants to obtain a higher biomass [31,37]. Under drought conditions, most of the selected unigenes involved in the photosynthetic process were up-regulated in Epichloë endophyte symbiotic DHG when they were compared with that of the non-symbiotic plants [31]. In addition, the presence of an Epichloë endophyte up- or down-regulates key genes in the biosynthesis and reaction pathways of jasmonic acid and salicylic acid under both biotic and abiotic stresses [30,38], and SA and JA can regulate complex immune responses to protect the plants from damage. In this analysis, 23 DEGs were enriched in the glutathione metabolism pathway (ko00480) (Figure 2). Glutathione has an important role in stress tolerance, and we speculate that the presence of an Epichloë endophyte promotes the production of defensive compounds for the plant’s protection.
In the current study, a total of 18 DEGs involved in photosynthesis were up-regulated under E+P+, E+P- and E-P+ conditions, of which 15 DEGs were in the E+P- plants. The transcriptome analysis showed that the presence of an Epichloë endophyte could significantly upregulate the genes related to photosynthesis.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof8111201/s1, Table S1: Two-way ANOVA for the effects of Blumeria graminis (P) and Epichloë endophyte (E) on plant height, tiller number, dry weight fresh and weight of Achnatherum inebrians; Figure S1: Effects of Epichloë-infected (E+) and Epichloë-free (E-) on plant height (a), tiller number (b), dry weight (c) and fresh weight (d) under the pathogen-inoculated (P+) and non-inoculated (P-). Figure S2: Number of up-regulated (a) and down-regulated (b) plant genes associated with defense responses given the presence of Epichloë-infected (E+), the pathogen Blumeria graminis (P+) and both fungi (E+P+) on Achnatherum inebrians plants. E-: Epichloë-free, P-: non-inoculated; Table S2: Selected unigenes associated with processes of photosynthesis and photosynthesis-antenna proteins identified in the RNA-seq analysis in the present study; Table S3: Effect size of net photosynthetic efficiency (NPE) of different grasses under Epichloë endophyte infestation and Table S4: Effect size of net photosynthetic efficiency (NPE) of Epichloë-infected and Epichloë-free grasses under pathogenic fungi inoculation. References [72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112] are cited in Supplementary Materials.
Author Contributions
Conceptualization, X.Z.; resources, S.Z., F.Z. and Z.N.; data curation, Y.Z.; supervision, Z.Z.; writing—Original draft preparation, Y.Z.; writing—Review and editing, M.J.C., X.Z. and Y.Z.; All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data supporting the findings of this study are available within the paper and within its supplementary materials published online. The RNA-seq used in this study have been deposited in the Squence Read Achieve (SRA) of the NCBI database under the accession number PRJNA748183.
Acknowledgments
We wish to thank the editor and anonymous reviewers for their valuable comments.This work was financially supported by the National Nature Science Foundation of China (31772665), National Basic Research Program of China (2014CB138702) and the Fundamental Research Funds for the Central Universities (lzujbky-2022-ey21), Lanzhou University.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pieterse, C.M.J.; Leon-Reyes, A.; Van der Ent, S.; Van Wees, S.C.M. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef]
- Inuma, T.; Khodaparast, S.A.; Takamatsu, S. Multilocus phylogenetic analyses within Blumeria graminis, a powdery mildew fungus of cereals. Mol. Phylogenetics Evol. 2007, 44, 741–751. [Google Scholar] [CrossRef]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 fungal pathogens in molecular plant pathology: Top 10 fungal pathogens. Mol. Plant Path. 2012, 13, 414–430. [Google Scholar] [CrossRef]
- Glawe, D.A. The powdery mildews: A review of the world’s most familiar (yet poorly known) plant pathogens. Annu. Rev. Phytopathol. 2008, 46, 27–51. [Google Scholar] [CrossRef]
- Edwards, H.H. Development of primary germ tubes by conidia of Blumeria graminis f.sp. hordei on leaf epidermal cells of Hordeum vulgare. Can. J. Bot. 2002, 80, 1121–1125. [Google Scholar] [CrossRef]
- Yamaoka, N.; Matsumoto, I.; Nishiguchi, M. The role of primary germ tubes (PGT) in the life cycle of Blumeria graminis: The stopping of PGT elongation is necessary for the triggering of appressorial germ tube (AGT) emergence. Physiol. Mol. Plant. Pathol. 2006, 69, 153–159. [Google Scholar] [CrossRef]
- Zhu, M.; Riederer, M.; Hildebrandt, U. Very-long-chain aldehydes induce appressorium formation in ascospores of the wheat powdery mildew fungus Blumeria graminis. Fungal Biol. 2017, 121, 716–728. [Google Scholar] [CrossRef]
- Hansjakob, A.; Riederer, M.; Hildebrandt, U. Appressorium morphogenesis and cell cycle progression are linked in the grass powdery mildew fungus Blumeria graminis. Fungal Biol. 2012, 116, 890–901. [Google Scholar] [CrossRef] [PubMed]
- Akhkha, A.; Clarke, D.D.; Dominy, P.J. Relative tolerances of wild and cultivated barley to infection by Blumeria graminis f. sp. hordei (Syn. Erysiphe graminis f. sp. hordei). II-the effects of infection on photosynthesis and respiration. Physiol. Mol. Plant Pathol. 2003, 62, 347–354. [Google Scholar] [CrossRef]
- Cao, A.; Xing, L.; Wang, X.; Wang, W.; Sun, Y.; Qian, C.; Ni, J.; Chen, Y.; Liu, D.; Wang, X.; et al. Serine/threonine kinase gene Stpk-V, a key member of powdery mildew resistance gene Pm21, confers powdery mildew resistance in wheat. Proc. Natl. Acad. Sci. USA 2011, 108, 7727–7732. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.R.; Yao, D.M.; Duan, X.Y.; Liu, W.; Fan, J.R.; Ding, K.J.; Zhou, Y.L. Effects of powdery mildew on 1000-kernel weight, crude protein content and yield of winter wheat in three consecutive growing seasons. J. Integr. Agric. 2014, 13, 1530–1537. [Google Scholar] [CrossRef]
- Liu, N.; Lei, Y.; Gong, G.; Zhang, M.; Wang, X.; Zhou, Y.; Qi, X.; Chen, H.; Yang, J.; Chang, X.; et al. Temporal and spatial dynamics of wheat powdery mildew in Sichuan Province, China. Crop Prot. 2015, 74, 150–157. [Google Scholar] [CrossRef]
- Leuchtmann, A. Systematics, distribution, and host specificity of grass endophytes. Nat. Toxins 1993, 1, 150–162. [Google Scholar] [CrossRef] [PubMed]
- Schardl, C.L.; Craven, K.D.; Speakman, S.; Stromberg, A.; Lindstrom, A.; Yoshida, R. A novel test for host-symbiont codivergence indicates ancient origin of fungal endophytes in grasses. Syst. Biol. 2008, 57, 483–498. [Google Scholar] [CrossRef] [PubMed]
- Leuchtmann, A.; Bacon, C.W.; Schardl, C.L.; White, J.F., Jr.; Tadych, M. Nomenclatural realignment of Neotyphodium species with genus Epichloë. Mycologia 2014, 106, 202–215. [Google Scholar] [CrossRef]
- Vignale, M.V.; Astiz-Gassó, M.M.; Novas, M.V.; Iannone, L.J. Epichloë endophytes confer resistance to the smut Ustilago bullata in the wild grass Bromus auleticus (Trin.). Biol. Control. 2013, 67, 1–7. [Google Scholar] [CrossRef]
- Bastías, D.A.; Alejandra Martínez-Ghersa, M.; Newman, J.A.; Card, S.D.; Mace, W.J.; Gundel, P.E. The plant hormone salicylic acid interacts with the mechanism of anti-herbivory conferred by fungal endophytes in grasses: Effects of salicylic acid on fungal endophytes. Plant Cell Environ. 2018, 41, 395–405. [Google Scholar] [CrossRef] [PubMed]
- Pérez, L.I.; Gundel, P.E.; Zabalgogeazcoa, I.; Omacini, M. An ecological framework for understanding the roles of Epichloë endophytes on plant defenses against fungal diseases. Fungal Biol. Rev. 2020, 34, 115–125. [Google Scholar] [CrossRef]
- Christensen, M.J.; Bennett, R.J.; Ansari, H.A.; Koga, H.; Johnson, R.D.; Bryan, G.T.; Simpson, W.R.; Koolaard, J.P.; Nickless, E.M.; Voisey, C.R. Epichloë endophytes grow by intercalary hyphal extension in elongating grass leaves. Fungal Genet. Biol. 2008, 45, 84–93. [Google Scholar] [CrossRef]
- Li, C.J.; Nan, Z.B.; Paul, V.; Dapprich, P.; Liu, Y. A new Neotyphodium species symbiotic with drunken horse grass (Achnatherum inebrians) in China. Mycotaxon 2004, 90, 141–147. [Google Scholar]
- Chen, L.; Li, X.Z.; Li, C.J.; Swoboda, G.A.; Young, C.A.; Sugawara, K.; Leuchtmann, A.; Schardl, C.L. Two distinct Epichloë species symbiotic with Achnatherum inebrians, drunken horse grass. Mycologia 2015, 107, 863–873. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.Z. Several poisonous weeds in grassland of northwest. J. Gansu Agric. Univ. 1954, 2, 9–16. [Google Scholar]
- Shi, Z.C. Important Poisonous Plants in Chinese Grassland; World Book Publishing Company: Beijing, China, 1997; pp. 166–176. [Google Scholar]
- Nan, Z.B.; Li, C.J. Neotyphodium in native grasses in China and observations on endophyte/host interactions. In Proceedings of the 4th International Neotyphodium/Grass Interactions Symposium, Soest, Germany, 27–29 September 2000. [Google Scholar]
- Liu, Z.P. Animal Toxicology; China Agricultural Press: Beijing, China, 2006; pp. 152–154. [Google Scholar]
- Zhang, X.X.; Li, C.J.; Nan, Z.B. Effects of salt and drought stress on alkaloid production in endophyte-infected drunken horse grass (Achnatherum inebrians). Biochem. Syst. Ecol. 2011, 39, 471–476. [Google Scholar] [CrossRef]
- Zhang, X.X.; Nan, Z.B.; Li, C.J.; Gao, K. Cytotoxic effect of ergot alkaloids in Achnatherum inebrians infected by the Neotyphodium gansuense endophyte. J. Agric. Food. Chem. 2014, 62, 7419–7422. [Google Scholar] [CrossRef]
- Liang, Y.; Wang, H.; Li, C.; Nan, Z.B.; Li, F. Effects of feeding drunken horse grass infected with Epichloë gansuensis endophyte on animal performance, clinical symptoms and physiological parameters in sheep. BMC Vet. Res. 2017, 13, 223. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Li, N.N.; Zhang, Y.W.; Li, C.J.; Zhang, X.X.; Nan, Z.B. Role of Epichloë endophytes in defense responses of cool-season grasses to pathogens: A review. Plant Dis. 2018, 102, 2061–2073. [Google Scholar] [CrossRef]
- Zhao, Z.R.; Kou, M.Z.; Zhong, R.; Xia, C.; Christensen, M.J.; Zhang, X.X. Transcriptome analysis revealed plant hormone biosynthesis and response pathway modification by Epichloë gansuensis in Achnatherum inebrians under different soil moisture avail-ability. J. Fungi 2021, 7, 640. [Google Scholar] [CrossRef]
- Zhong, R.; Bastías, D.A.; Zhang, X.X.; Li, C.J.; Nan, Z.B. Vertically transmitted Epichloë systemic endophyte enhances drought tolerance of Achnatherum inebrians host plants through promoting pbhotosynthesis and biomass accumulation. J. Fungi 2022, 8, 512. [Google Scholar] [CrossRef]
- Wang, J.F.; Tian, P.; Christensen, M.J.; Zhang, X.X.; Li, C.J.; Nan, Z.B. Effect of Epichloë gansuensis endophyte on the activity of enzymes of nitrogen metabolism, nitrogen use efficiency and photosynthetic ability of Achnatherum inebrians under various NaCl concentrations. Plant Soil 2019, 435, 57–68. [Google Scholar] [CrossRef]
- Chen, N.; He, R.; Chai, Q.; Li, C.J.; Nan, Z.B. Transcriptomic analyses giving insights into molecular regulation mechanisms involved in cold tolerance by Epichloë endophyte in seed germination of Achnatherum inebrians. Plant Growth Regul. 2016, 80, 367–375. [Google Scholar] [CrossRef]
- Zhang, X.X.; Fan, X.M.; Li, C.J.; Nan, Z.B. Effects of cadmium stress on seed germination, seedling growth and antioxidative enzymes in Achnatherum inebrians plants infected with a Neotyphodium endophyte. Plant Growth Regul. 2010, 60, 91–97. [Google Scholar] [CrossRef]
- Zhang, X.X.; Li, C.J.; Nan, Z.B. Effects of cadmium stress on seed germination and seedling growth of Elymus dahuricus infected with the Neotyphodium endophyte. Sci. China Life Sci. 2012, 55, 793–799. [Google Scholar] [CrossRef] [PubMed]
- Xia, C.; Zhang, X.X.; Christensen, M.J.; Nan, Z.B.; Li, C.J. Epichloë endophyte affects the ability of powdery mildew (Blumeria graminis) to colonise drunken horse grass (Achnatherum inebrians). Fungal Ecol. 2015, 16, 26–33. [Google Scholar] [CrossRef]
- Xia, C.; Li, N.N.; Zhang, X.X.; Feng, Y.; Christensen, M.J.; Nan, Z.B. An Epichloë endophyte improves photosynthetic ability and dry matter production of its host Achnatherum inebrians infected by Blumeria graminis under various soil water conditions. Fungal Ecol. 2016, 22, 26–34. [Google Scholar] [CrossRef]
- Kou, M.Z.; Bastías, D.A.; Christensen, M.J.; Zhong, R.; Nan, Z.B.; Zhang, X.X. The plant salicylic acid signalling pathway regulates the infection of a biotrophic pathogen in grasses associated with an Epichloë endophyte. J. Fungi 2021, 7, 633. [Google Scholar] [CrossRef]
- Doubnerová, V.; Ryšlavá, H. What can enzymes of C4 photosynthesis do for C3 plants under stress? Plant Sci. 2011, 180, 575–583. [Google Scholar] [CrossRef]
- Freschi, L.; Mercier, H. Connecting environmental stimuli and crassulacean acid metabolism expression: Phytohormones and other signaling molecules. Prog. Bot. 2012, 73, 231–255. [Google Scholar]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Todorova, D.; Aleksandrov, V.; Anev, S.; Sergiev, I. Photosynthesis alterations in wheat plants induced by herbicide, soil drought or flooding. Agronomy 2022, 12, 390. [Google Scholar] [CrossRef]
- Kissana, S.N.; Mujahid, Y.M.; Mustafa, Z.S. A Technical Report to Apprise the Issues and Future Strategies; National Agricultural Research Center, Pakistan Agricultural Research Council: Islamabad, Pakistan, 2003; pp. 2002–2003. [Google Scholar]
- Carretero, R.; Bancal, M.O.; Miralles, D.J. Effect of leaf rust (Puccinia triticina) on photosynthesis and related processes of leaves in wheat crops grown at two contrasting sites and with different nitrogen levels. Eur. J. Agron. 2011, 35, 237–246. [Google Scholar] [CrossRef]
- Zhang, H.J.; Li, X.; White, J.F.; Wei, X.; He, Y.; Li, C.J. Epichloë endophyte improves ergot disease resistance of host (Achnatherum inebrians) by regulating leaf senescence and photosynthetic capacity. J. Plant. Growth. Regul. 2022, 41, 808–817. [Google Scholar] [CrossRef]
- Bassanezi, R.B.; Amorim, L.; Filho, A.B.; Hau, B.; Berger, R.D. Accounting for photosynthetic efficiency of bean leaves with rust, angular leaf spot and anthracnose to assess crop damage: Photosynthesis of diseased bean leaves and crop damage. Plant Path. 2001, 50, 443–452. [Google Scholar] [CrossRef]
- Robert, C.; Bancal, M.; Ney, B.; Lannou, C. Wheat leaf photosynthesis loss due to leaf rust, with respect to lesion development and leaf nitrogen status. New Phytol. 2005, 165, 227–241. [Google Scholar] [CrossRef] [PubMed]
- Ma, M.Z.; Christensen, M.J.; Nan, Z.B. Effects of the endophyte Epichloë festucae var. lolii of perennial ryegrass (Lolium perenne) on indicators of oxidative stress from pathogenic fungi during seed germination and seedling growth. Eur. J. Plant Pathol. 2015, 141, 571–583. [Google Scholar] [CrossRef]
- Ambrose, K.V.; Belanger, F.C. SOLiD-SAGE of endophyte-infected red fescue reveals numerous effects on host transcriptome and an abundance of highly expressed fungal secreted proteins. PLoS ONE 2012, 7, e53214. [Google Scholar] [CrossRef]
- Zhong, R.; Zhang, L.; Zhang, X.X. Allelopathic Effects of Foliar Epichloë Endophytes on Belowground Arbuscular Mycorrhizal Fungi: A Meta-Analysis. Agriculture 2022, 12, 1768. [Google Scholar] [CrossRef]
- Hedges, L.V.; Gurevitch, J.; Curtis, P.S. The meta-analysis of response ratios in experimental ecology. Ecology 1999, 80, 1150–1156. [Google Scholar] [CrossRef]
- Gurevitch, J.; Curtis, P.S.; Jones, M.H. Meta-analysis in ecology. Adv. Ecol. Res. 2001, 32, 199–247. [Google Scholar]
- Saibo, N.J.M.; Lourenço, T.; Oliveira, M.M. Transcription factors and regulation of photosynthetic and related metabolism under environmental stresses. Ann. Bot. 2009, 103, 609–623. [Google Scholar] [CrossRef]
- Rahnama, A.; Poustini, K.; Tavakkol-Afshari, R.; Tavakoli, A. Growth and stomatal responses of bread wheat genotypes in tolerance to salt stress. Int. J. Biol. Sci. 2010, 6, 216–221. [Google Scholar]
- Medrano, H.; Escalona, J.M.; Bota, J.; Gulias, J.; Flexas, J. Regulation of photosynthesis of C3 plants in response to progressive drought: The stomatal conductance as a reference parameter. Ann. Bot. 2002, 89, 895–905. [Google Scholar] [CrossRef] [PubMed]
- Karaba, A.; Dixit, S.; Greco, R.; Aharoni, A.; Trijatmiko, K.R.; Marsch-Martinez, N.; Krishnan, A.; Nataraja, K.N.; Udayakumar, M.; Pereira, A. Improvement of water use efficiency in rice by expression of HARDY, an Arabidopsis drought and salt tolerance gene. Proc. Natl. Acad. Sci. USA 2007, 104, 15270–15275. [Google Scholar] [CrossRef] [PubMed]
- Dias, M.C.; Brüggemann, W. Limitations of photosynthesis in Phaseolus vulgaris under drought stress: Gas exchange, chlorophyll fluorescence and Calvin cycle enzymes. Photosynthetica 2010, 48, 96–102. [Google Scholar] [CrossRef]
- Dias, M.C.; Brüggemann, W. Water-use efficiency in Flaveria species under drought-stress conditions. Photosynthetica 2010, 48, 469–473. [Google Scholar] [CrossRef]
- Ayres, P.G. Patterns of stomatal behaviour, transpiration, and CO2 exchange in pea following infection by powdery mildew (Erysiphe pisi). J. Exp. Bot. 1976, 27, 1196–1205. [Google Scholar] [CrossRef]
- Moriondo, M.; Orlandini, S.; Giuntoli, A.; Bindi, M. The effect of downy and powdery mildew on grapevine (Vitis vinifera L.) leaf gas exchange. J. Phytopathol. 2005, 153, 350–357. [Google Scholar] [CrossRef]
- Xia, C.; Christensen, M.J.; Zhang, X.X.; Nan, Z.B. Effect of Epichloë gansuensis endophyte and transgenerational effects on the water use efficiency, nutrient and biomass accumulation of Achnatherum inebrians under soil water deficit. Plant Soil 2018, 424, 555–571. [Google Scholar] [CrossRef]
- Soleimani, M.; Hajabbasi, M.A.; Afyuni, M.; Mirlohi, A.; Borggaard, O.K.; Holm, P.E. Effect of endophytic fungi on cadmium tolerance and bioaccumulation by Festuca arundinacea and Festuca pratensis. Int. J. Phytoremediation 2010, 12, 535–549. [Google Scholar] [CrossRef]
- Sabzalian, M.R.; Mirlohi, A.; Sharifnabi, B. Reaction to powdery mildew fungus, Blumeria graminis in endophyte-infected and endophyte-free tall and meadow fescues. Australas. Plant Path. 2012, 41, 565–572. [Google Scholar] [CrossRef]
- Gundel, P.E.; Irisarri, J.G.N.; Fazio, L.; Casas, C.; Pérez, L.I. Inferring field performance from drought experiments can be misleading: The case of symbiosis between grasses and Epichloë fungal endophytes. J. Arid. Environ. 2016, 132, 60–62. [Google Scholar] [CrossRef]
- Hamilton, C.E.; Bauerle, T.L. A new currency for mutualism? Fungal endophytes alter antioxidant activity in hosts responding to drought. Fungal Divers. 2012, 54, 39–49. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Zamioudis, C.; Berendsen, R.L.; Weller, D.M.; Van Wees, S.C.M.; Bakker, P.A.H.M. Induced systemic resistance by beneficial microbes. Annu. Rev. Phytopathol. 2014, 52, 347–375. [Google Scholar] [CrossRef]
- Ling, N.; Li, W.; Xu, G.; Qi, Z.; Ji, C.; Liu, X.; Cui, D.; Sun, Y. Transcriptomic sequencing reveals the response of Dunaliella salina to copper stress via the increased photosynthesis and carbon mechanism. Mol. Omics 2021, 17, 769–782. [Google Scholar] [CrossRef]
- Rozpądek, P.; Wężowicz, K.; Nosek, M.; Ważny, R.; Tokarz, K.; Lembicz, M.; Miszalski, Z.; Turnau, K. The fungal endophyte Epichloë typhina improves photosynthesis efficiency of its host orchard grass (Dactylis glomerata). Planta 2015, 242, 1025–1035. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Ma, J.; He, Y.; Yu, S.; Lin, Z.; Xiong, Y.; Rafique, F.; Jiang, F.; Sun, L.; Ma, M.; et al. Comparative transcriptomic and proteomic analyses of the green and white parts of chimeric leaves in Ananas comosus var. bracteatus. PeerJ 2019, 7, e7261. [Google Scholar] [CrossRef] [PubMed]
- Fedorov, V.A.; Kovalenko, I.B.; Khruschev, S.S.; Ustinin, D.M.; Antal, T.K.; Riznichenko, G.Y.; Rubin, A.B. Comparative analysis of plastocyanin-cytochrome f complex formation in higher plants, green algae and cyanobacteria. Physiol. Plant. 2019, 166, 320–335. [Google Scholar] [CrossRef]
- Su, X.; Ma, J.; Pan, X.; Zhao, X.; Chang, W.; Liu, Z.; Zhang, X.; Li, M. Antenna arrangement and energy transfer pathways of a green algal photosystem-I-LHCI supercomplex. Nat. Plants 2019, 5, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Bao, G.S. Effect of the Hemiparasitic Plant Pedicularis Kansuensis on Growth and Photosynthetic Properties of Stipa Purpurea-Epichloë symbiosis; Lanzhou University: Lanzhou, China, 2015. [Google Scholar]
- Bao, G.S.; Song, M.L.; Wang, Y.Q.; Li, C.J. Effects of parasitic Artemisia annua from Gansu on the photosynthetic characteristics of endophytic fungal symbiont in grasses. Acta Microbiol Sin. 2020, 60, 294–305. [Google Scholar]
- Bao, G.S. Study of Epichloë endophyte-grass symbionts on photosynthetic characteristics of hemiparasitic plant. Chin. Qinghai J. Anim. Vet. Sci. 2020, 50, 1–7. [Google Scholar]
- Cao, Y. Effect of Interaction between Jasmonic Acid and Endophytic Fungi on Wild Barely under Salt Stress; Lanzhou University: Lanzhou, China, 2018. [Google Scholar]
- Chen, T.X.; Johnson, R.; Chen, S.H.; Li, C.J. Infection by the fungal endophyte Epichloë bromicola enhances the tolerance of wild barley (Hordeum brevisubulatum) to salt and alkali stresses. Plant Soil 2018, 428, 353–370. [Google Scholar] [CrossRef]
- Cui, X.L.; Xia, C. Effect of exogenous abscisic acid on seedling establishment of Epichloë gansuensis-Achnatherum inebrians symbiont. Acta Prataculture Sin. 2020, 29, 70–80. [Google Scholar]
- Deng, J. Mechanisms of the Effects of Arbuscular Mycorrhizal Fungus and Grass Endophyte on Left Spot of Perennial Ryegrass; Lanzhou University: Lanzhou, China, 2021. [Google Scholar]
- Deng, J.; Li, F.; Duan, T.Y. Effects of AM fungus and grass endophyte on the infection of Lolium perenne by the pathogen Biopolaris sorokinianum in a greenhouse. Acta Prataculture Sin. 2019, 28, 103–113. [Google Scholar]
- Du, Y.J. Stress Resistance Research of Endophyte in Wild Festuca L and Their Association; Beijing Forestry University: Beijing, China, 2010. [Google Scholar]
- Du, Y.J.; Sun, X.B.; Han, L.B. Effect of Neotyphodium starrii infected on photosynthetic and orphological characteristics of tall fescue under high temperature. J. Cent. South Univ. For. Technol. 2010, 30, 41–47. [Google Scholar]
- Du, Y.J.; Wang, Q.; Han, L.B. Endophytic fungus Neotyphodium typhinum infection effects on photosynthetic characteristics of tall fescue. Ecol. Environ. Sci. 2009, 18, 45–47. [Google Scholar]
- Fang, A.G. Effect of Neotyphodium Endophyte and AM Fungi on Growth of Hordeum Brevisubulatum under Salt and Phosphorus Stress Conditions; Lanzhou: Lanzhou University, 2013. [Google Scholar]
- Guo, Y.E. Effect of Phouphorus, Claroideoglomus etunicatum and Grass Endophte on Perennial Ryegrass Left Spot Disease Caused by Bipolaris zeae; Lanzhou University: Lanzhou, China, 2018. [Google Scholar]
- Guo, Y.E.; Gao, P.; Li, F. Effects of AM fungi and grass endophytes on perennial ryegrass Bipolaris sorokiniana leaf spot disease under limited soil nutrients. Eur. J. Plant Path. 2019, 154, 659–671. [Google Scholar] [CrossRef]
- Guo, Y.E.; Li, F.; Wang, X.Y.; Wang, Z.G.; Duan, T.Y. Effects of phosphorus, AM fungi and grass endophytes on leaf spot of perennial ryegrass. Cao Xue 2018, 4, 17–26, 72. [Google Scholar]
- Guo, Y.E.; Li, Y.D.; Gao, P.; Wang, Z.G.; Duan, T.Y. Effects of Claroideoglomus etunicatum and grass endophyte on the growth of Lolium perenne under different phosphorus levels. Acta Prataculture Sin. 2018, 26, 1458–1466. [Google Scholar]
- Han, R.; Li, X.; Ren, A.Z.; Gao, Y.B. Physiological ecological effect of endophyte infection on Achnatherum sibiricum under drought stress. Acta. Ecol. Sin. 2011, 31, 2115–2123. [Google Scholar]
- Jia, T. Effect of Endophytic Species, Host Genotype and Water/Nutrient Supply on the Performance of Grass-Endophyte Symbionts; Nankai Universitg: Tianjing, China, 2014. [Google Scholar]
- Jia, T.; Ren, A.Z.; Gao, Y.B. Host genotype overrides endophyte infection effects on growth, physiology, and nutrient content of a native grass, Achnatherum sibiricum. Plant Ecol. 2014, 215, 875–887. [Google Scholar] [CrossRef]
- Jia, T.; Ren, A.Z.; Wang, S.; Gao, Y.B. Effect of Endophytic fungi on growth and photosynthetic characteristics of Achnatherum sibiricum. Acta. Ecol. Sin. 2011, 31, 4811–4817. [Google Scholar]
- Li, C.; Li, X.; Ren, A.Z.; Gao, Y.B. Effect of endophyte infection on Zn resistance of Achnatherum sibiricum. Acta Sci. Nat. Univ. Nankaiensis 2013, 46, 29–35. [Google Scholar]
- Li, C.; Ren, A.Z.; Gao, Y.B. Effect of endophyte intection on Zn resistance of tall fescue. Acta. Ecol. Sin. 2010, 30, 1684–1690. [Google Scholar]
- Li, F.; Deng, J.; Nzabanita, C. Growth and physiological responses of perennial ryegrass to an AMF and an Epichloë endophyte under different soil water contents. Symbiosis 2019, 79, 151–161. [Google Scholar] [CrossRef]
- Li, F.; Li, Y.Z.; Duan, T.Y. Effects of interactions between a grass endophyte and an arbuscular mycorrhizal fungus on perennial ryegrass growth. Acta Pratculturae Sin. 2017, 26, 132–140. [Google Scholar]
- Li, M.M. Effect of Epichloë Endophyte on Drought Resistance of Different Ecotypes Festuca sinensis; Lanzhou University: Lanzhou, China, 2019. [Google Scholar]
- Li, N.N. Effect of Interaction of Endophyte and Blumeria graminis on the Achnatherum inebrians in Different Water Conditions; Lanzhou University: Lanzhou, China, 2018. [Google Scholar]
- Li, X.; Zhou, Y.; Mace, W.; Qin, J.; Liu, H.; Chen, W.; Ren, A.Z.; Gao, Y.B. Endophyte species influence the biomass production of the native grass Achnatherum sibiricum (L.) Keng under high nitrogen availability. Ecol. Evol. 2016, 6, 8595–8606. [Google Scholar] [CrossRef]
- Liang, Y.; Gao, Y.B.; Chen, S.P.; Ren, A.Z. Effects of endophyte infection on photosynthesis, transpiration and water use efficiency of Lolium perenen L. under drought stress. Acta Phytoecol. Sin. 2001, 5, 537–543. [Google Scholar]
- Lin, F.; Li, C.; Zhang, X.; Bao, X.Y.; Ren, A.Z.; Gao, Y.B. Effect of endophytic fungi on photosynthetic characteristics of three Achnatherum sibiricum populations in Inner Mongolia Steppe. Bull. Botanical. Res. 2009, 29, 61–68. [Google Scholar]
- Liu, B.H. Biological Properties of Endophtes/Calamagrostis Symbiota; Naning Agricultural University: Naning, China, 2012. [Google Scholar]
- Liu, L. Response of Epichloë gansuensis-Achnatherum Inebrians Symbiont on Stresses of Cold and Powdery Mildew Disease Treated by SA and ABA; Lanzhou University: Lanzhou, China, 2016. [Google Scholar]
- Ma, M.Z. Disease Resistance of Perennial Ryegrass (Lolium perenne)-Epichloë festucae var. lolii Endophyte Symbiont and Their Resistance Mechanism to Bipolaris sorokiniana; Lanzhou University: Lanzhou, China, 2015. [Google Scholar]
- Ma, M.Z.; Nan, Z.B. Effect of fungal endophytes against rust disease of perennial ryegrass (Lolium perenne) on growth and physiological indices. Acta Pratculturae Sin. 2011, 20, 150–156. [Google Scholar]
- Morse, L.J.; Day, T.A.; Faeth, S.H. Effect of Neotyphodium endophyte infection on growth and leaf gas exchange of Arizona fescue under contrasting water availability regimes. Environ. Exp. Bot. 2002, 48, 257–268. [Google Scholar] [CrossRef]
- Qin, J.H.; Lu, Y.; Li, Z.; Zhou, Y.; Ren, A.Z.; Gao, Y.B. Effects of methyl jasmonate treatments and endophyte infection on growth of Achnatherum sibiricum. Chin. J. Appl. Ecol. 2015, 26, 1145–1152. [Google Scholar]
- Shi, Z.B.; Zhou, Y.; Li, X.; Ren, A.Z.; Gao, Y.B. Physioecological effects of endophyte infection on the host grass with elevated CO2. Acta. Ecol. Sin. 2013, 33, 6135–6141. [Google Scholar]
- Wang, J.L.; Gao, Y.B.; Ren, A.Z.; Wang, W.; Zhao, N.X. Effects of endophyte infection on photosynthesis, transpiration and biomass of Lolium perenne L. at different nitrogen levels. Chin. Bull. Bot. 2004, 5, 539–546. [Google Scholar]
- Xia, C. Responses of Epichloë gansuensis-Achnatherum inebrians Symbiont to Drought Stress; Lanzhou University: Lanzhou, China, 2018. [Google Scholar]
- Yan, Z.C.; Li, Y.D.; Cheng, W.J.; Gao, P.; Guo, Y.; Duan, T.Y. Effects of AM fungi and grass endophyte on the growth of ryegrass under different salt concentrations. Grassland. Turf. 2018, 38, 63–70. [Google Scholar]
- Zhang, H.J. The Effect of Pathogenic Ergot bacteria on the Epichloë Endophyte Symbionts of Achnatherum sibiricum; Lanzhou University: Lanzhou, China, 2021. [Google Scholar]
- Zhang, J.F. Effect of Fungal Endophyte on Intraspecific Competition of Elymus Nutans under Water Stress; Lanzhou University: Lanzhou, China, 2013. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).