Abstract
Calcineurin (CN) is an attractive antifungal target as it is critical for growth, stress response, drug resistance, and virulence in fungal pathogens. The immunosuppressive drugs, tacrolimus (FK506) and cyclosporin A (CsA), are fungistatic and specifically inhibit CN through binding to their respective immunophilins, FK506-binding protein (FKBP12), and cyclophilin (CypA). We are focused on CN structure-based approaches for the development of non-immunosuppressive FK506 analogs as antifungal therapeutics. Here, we examined the effect of the novel CN inhibitor, CN585, on the growth of the human pathogen Aspergillus fumigatus, the most common cause of invasive aspergillosis. Unexpectedly, in contrast to FK506, CN585 exhibited off-target effect on A. fumigatus wild-type and the azole- and echinocandin-resistant strains. Unlike with FK506 and CsA, the A. fumigatus CN, FKBP12, CypA mutants (ΔcnaA, Δfkbp12, ΔcypA) and various FK506-resistant mutants were all sensitive to CN585. Furthermore, in contrast to FK506 the cytosolic to nuclear translocation of the CN-dependent transcription factor (CrzA-GFP) was not inhibited by CN585. Molecular docking of CN585 onto human and A. fumigatus CN complexes revealed differential potential binding sites between human CN versus A. fumigatus CN. Our results indicate CN585 may be a non-specific inhibitor of CN with a yet undefined antifungal mechanism of activity.
Keywords:
Aspergillus; calcineurin; CN585; cyclosporin A; cyclophilin; FK506; FKBP12; molecular dynamics; molecular docking 1. Introduction
Calcineurin (CN) is a highly conserved Ca2+-calmodulin-dependent serine/threonine phosphatase in mammalian and fungal systems [1,2]. Apart from its multi-functional cellular roles in mammalian systems [3,4,5], its importance for regulating growth, stress responses, and virulence in different fungal pathogens is well established in species of Aspergillus, Candida, Cryptococcus, and Mucor [1,6].
CN functions as a complex of the catalytic subunit (CnA) and regulatory subunit (CnB) and is activated by increased intracellular Ca2+ concentration and the binding of the Ca2+-binding protein calmodulin [7]. Members of the NFAT transcription factor family in their hyperphosphorylated form are the major substrates dephosphorylated by CN in immune cells [8,9]. Following dephosphorylation, nuclear translocation of NFAT and its binding onto regulatory promoter elements of genes stimulates the expression of cytokines and chemokines, which play vital role in the initiation of immune responses [10]. The well-established mode of CN-complex inhibition is through the immunosuppressive drugs, tacrolimus (FK506), and cyclosporin A (CsA), that are widely used in transplantation to prevent graft rejection [9,11,12,13]. FK506 and CsA bind to their respective immunophilins, FKBP12 (12-kDa FK506 binding protein), and CypA (cyclophilin A). This immunophilin-immunosuppressant complex then binds CN complex to block substrate binding and inhibit CN function [14,15].
Although FK506 and CsA exhibit potent antifungal activity in vitro, their host immunosuppressive effects make it difficult to exploit them as antifungal therapeutics. In an effort to develop CN inhibitors that could be used as antifungals, FK506 analogs that permeated mammalian cells but not fungal cells were synthesized and dosed along with FK506 to antagonize the immunosuppressive effect of FK506 without lowering its antifungal activity [16]. Non-immunosuppressive FK506 analogs with antifungal activity have also been generated through combinatorial biosynthetic approaches with structural modifications at CN-binding and FKBP-binding regions [17,18]. Our recent studies have focused on structural analysis of fungal CN-inhibitor complexes to identify differences between mammalian and fungal CN ternary complexes to enable the rational design of fungal-specific non-immunosuppressive FK506 analogs [19,20,21]. Here, we tested the inhibitory activity of one novel mammalian CN inhibitor, CN585 [22], on the fungal pathogen A. fumigatus. CN585 is a cell permeable 2,6-diaryl-substituted pyrimidine derivative belonging to a new class of CN inhibitors and characterized as an immunosuppressive and non-competitive reversible inhibitor of CN in mammalian cells. It has been shown to specifically inhibit mammalian CN activity without affecting other serine/threonine phosphatases [22].
2. Materials and Methods
2.1. Strains, Culture Conditions and In Vitro Drug Susceptibility Assays
The A. fumigatus wild-type strain (akuBKU80), azole-resistant clinical isolates (F14946 and F16216), echinocandin-resistant strain (EMFR-S678P), CN pathway deletion strains including CN catalytic subunit deletion (ΔcnaA), immunophilin FKBP12 and cyclophilin A deletion strains (Δfkbp12; ΔcypA) and various cnaA mutated strains were utilized for antifungal susceptibility assays. Strains were cultured on GMM (glucose minimal medium) agar for 5 days at 37 °C for collection of conidia for the assays. Broth dilution antifungal susceptibility testing was performed according to the Clinical Laboratory Standards Institute (CLSI) guidelines in RPMI-1640 medium with slight modifications. Conidia were harvested in 0.05% Tween 80 and counted to obtain a spore concentration of 104/mL and subsequently diluted to obtain 5 × 102/mL in each well (200 µL) of a 96-well plate. Effect of CN585 (Sigma-Aldrich, St. Louis, MO, USA) or FK506 (Astellas Pharma, Tokyo, Japan) on growth was tested at concentrations ranging from 0 to 10 µg/mL. Interpretation of results on growth inhibition was performed after 48 h of growth at 37 °C by microscopic observation. Bright-field photomicrographs were acquired on a Nikon Diaphot phase contrast inverted microscope equipped with a Canon T5i digital camera.
2.2. Microscopy
To assess the effect of CN585 or FK506 on CrzA localization, conidia from the A. fumigatus akuBKU80 strain expressing CrzA-GFP fusion protein [23] were cultured for 18 h at 37 °C in 35 mm cover glass bottom Petri dish (Matek) in a total volume of 1000 µL GMM broth. After 18 h of growth, cultures were treated with FK506 (0.5 µg/mL) or CN585 (6 µg/mL) for 30 min and 120 min. Following CN inhibitor treatments, the respective cultures were exposed to CaCl2 (50 mM) for time points ranging between 0–60 min and visualized by fluorescence microscopy using an Axio Observer 3 scope. Differential interference contrast (DIC) and GFP-fluorescent images were captured at 10 min intervals over a period of 1 h using the 100x/1.4 oil Plan Apochromat DIC objective and Chroma GFP filter with Zeiss Axiocam 506 mono high resolution digital camera.
2.3. Molecular Modeling of Aspergillus fumigatus eIF4E
A. fumigatus eIF4E has not been structurally characterized, therefore, the protein sequence of A. fumigatus eIF4E (XP_748025.1) was submitted to the SWISS-MODEL web server to develop a structural model [24]. The sequence aligned well with that of the “Complex crystal structure of Ascaris suum eIF4E-3 with m7G cap” (PDB code: 3M93) and this was used as a template for the construction of the A. fumigatus eIF4E model [25].
2.4. Molecular Dynamic Simulations
All molecular dynamic (MD) simulations were performed with the GROMACS 2020 software package utilizing six CPU cores and one NVIDIA Tesla K80 GPU to provide a solvated representation (Abraham MJ, van der Spoel D, Lindahl E, Hess B. Gromacs Development Team. GROMACS User Manual, ver. 5.0, 2019). The single starting conformations used for all MD simulations were the X-ray crystal structures 6TZ7 (crystal structure of A. fumigatus CnA, CnB, FKBP12 and FK506 with FKBP12 and FK506 removed), 1MF8 (crystal structure of human CN complexed with CsA and human cyclophilin) with CsA and cyclophilin removed, and a structural model of A. fumigatus eIF4E (see Section 2.3) [26,27]. MD simulations were performed with the AMBER 99sb-ildn force field using the flexible simple point charge water model. The initial structures were immersed in a periodic water box with a dodecahedral shape that extended 1 nm beyond the protein in any dimension and neutralized with counter ions. Energy minimization was accomplished using the steepest descent algorithm with a final maximum force of <100 kJ mol−1 min−1 (0.01 nm step size, cutoff of 1.0 nm for the neighbor list, Coulomb interactions, and van der Waals interactions). After energy minimization, the system was subjected to equilibration at 300 K and normal pressure for 1 ns. All bonds were constrained with the LINCS algorithm, and virtual sites were used to allow a 4 fs time step (cutoff of 1.0 nm for the neighbor list, Coulomb interactions, and van der Waals interactions). After temperature stabilization, pressure stabilization was achieved by utilizing the v-rescale thermostat to hold the temperature at 300 K and the Berendsen barostat was used to bring the system to 1 bar pressure. Production MD calculations (100 ns) were performed under the same conditions, except that the positional restraints were removed (cutoff of 1.0 nm for the neighbor list, Coulomb interactions, and van der Waals interactions). GROMACS built-in and homemade scripts were used to analyze the MD simulation results. The α-carbon root-mean-square deviation (Cα-RMSD), radius of gyration (Rg), and center of mass (COM) were analyzed to confirm the stability and accuracy of the MD simulations. All images were produced in part through PyMOL (The PyMOL Molecular Graphics System, ver. 2.0; Schrödinger, LLC., New York, NY, USA).
2.5. Molecular Docking
Default High Ambiguity Drive Docking (HADDOCK) parameters were used throughout the docking procedure on the webserver with the following exceptions: auto_passive_radius = 6.5, rotate180_0 = False, crossdock = True, and calcdesolv = True [28,29]. Every 9 ns frame after equilibration (10 ns) from the MD simulation of the protein was extracted to form an ensemble of structures prior to docking over the last 90 ns. This helps to ensure the docking procedure examines multiple conformational states within the potential binding site prior to each docking event and the docked structures, providing a more accurate representation of the docked structures and their conformational flexibility in solution. Active residues for the proteins were defined as residues with a ≥60% solvent exposure for the protein and 100% of the CN585 small molecule was set to as actively involved in the interaction. The HADDOCK webserver was used for docking, therefore, the CN585 small molecule topology and parameter file was automatically generated through PRODRG [30]. Through the HADDOCK webserver, one thousand structures were generated for the first iteration (rigid docking), and 400 were generated for each subsequent iteration (semiflexible docking and water refinement). The Cα-RMSD values of the complexes were calculated using ProFit version 3.1 (SciTech Software, Chico, CA, USA). The structures were clustered based on Cα-RMSD and scored using HADDOCK’s internal scoring mechanism. The resulting Z-score analysis provided by HADDOCK indicates how many standard deviations from the average this cluster is located in terms of score (the more negative the better). The top cluster is the most reliable according to HADDOCK. The structures that represented the median score for the most reliable cluster were then visualized with PyMOL to ensure the integrity of the complex formed (The PyMOL Molecular Graphics System, Version 2.0 Schrödinger, LLC.).
3. Results
3.1. CN585 Significantly Inhibits Growth of A. fumigatus Wild-Type and the Azole- and Echinocandin-Resistant Strains
Both FK506 and CsA inhibit A. fumigatus growth by inducing the formation of hyperbranched and stunted hyphae with aberrant septation mimicking a CN deletion phenotype [31,32]. As a first step to analyze the effect of CN585 on A. fumigatus growth, the wild-type (akuBKU80) strain was cultured in RPMI liquid medium at increasing concentrations of CN585 ranging from 0–10 µg/mL based on the reported IC50 value for mammalian CN inhibition [22]. In contrast to our previously observed growth phenotypes following FK506/CsA treatment, CN585 treatment did not result in any hyperbranching, hyperseptation, or a stunted growth phenotype (Figure 1). CN585 caused a concentration-dependent growth inhibition of hyphal growth and, unlike FK506 or CsA, resulted in complete growth inhibition between 6–8 µg/mL. Azoles and echinocandins are currently used drugs for the treatment of invasive aspergillosis and target the fungal cell membrane (ergosterol) and cell wall (β-glucan and chitin), respectively. Strains with mutations in the cyp51A gene (F14946 and F16216) and fksA gene (EMFR-S678P) exhibit resistance to azoles and echinocandins, respectively. To further identify any strain-dependent variations in susceptibility to CN585 we next tested the A. fumigatus azole- (F14946 and F16216) [33] and echinocandin-resistant (EMFR-S678P) [34] strains which also showed a similar trend in susceptibility.

Figure 1.
Growth of the wild-type A. fumigatus (KU80) and the echinocandin-resistant (EMFR-S678P) and the azole-resistant (F14946, F16216 and F16134) strains in the presence of increasing concentrations of CN585. DMSO control growth is shown. Growth was monitored for 2 days in RPMI at 37 °C. Light microscopy photographs (×10 magnification) after 2 days of growth are shown. Note the complete inhibition in growth at >8 µg/mL CN585. Growth susceptibility assays were performed three times in triplicate.
3.2. CN585-Mediated Growth Inhibition Is Independent of Key Residues in the CN-Effector Binding Regions
Previous analysis of human CN inhibition by CN585 indicated that CsA-Cyp18 (cyclosporin A-cyclophilin) complex and CN585 may share similar binding site on the CN catalytic subunit (CnA) [22]. Earlier studies have also shown that the loop 7 region in CnA that is in close proximity to the CsA-CypA and FKBP12-FK506 complex binding region influences CN activity [35]. Based on our A. fumigatus CN ternary complex structure, the CnA loop 7 region (residues 328–339) is proximal to the FK506-FKBP12 complex binding region. In order to verify if the mode of CN inhibition by CN585 is mediated through interaction with any of these key residues near the catalytic domain including the loop 7 region or in the CnB-binding helix, which includes the CsA-CypA binding and the FK506-FKBP12 binding domains, we next tested the susceptibility of various CnA mutant strains to CN585. However, none of the loop 7 residue (Leu334; Asp335; Val336) mutants (cnaAmt-L334A; cnaAmt-D335A; cnaAmt-V336R) induced any resistance to CN585 (Figure 2) but exhibited variable resistance to CsA at 10 µg/mL (Figure S1). Mutations in residues Tyr363 (cnaAmt-Y363F) and Trp364 (cnaAmt-W364A) which are present in the CsA-CypA binding region also did not have any impact on CN585 sensitivity but exhibited resistance to CsA at 10 µg/mL (Figure S1). Mutations in the critical residues Asn367, Trp374 and Ser375 (cnaAmt-N367D; cnaAmt-W374L; cnaAmt-S375T), known to influence FK506 sensitivity [36], also did not reveal any altered sensitivity to CN585.

Figure 2.
Growth of the wild-type A. fumigatus (KU80), FK506-binding protein deletion (Δfkbp12), cyclophilin A deletion (ΔcypA) and the various cnaA mutant strains in the presence of increasing concentrations of CN585. DMSO control growth is shown. Growth was monitored for 2 days in RPMI at 37 °C. Light microscopy photographs (×10 magnification) after 2 days of growth are shown. Note the complete inhibition in growth for all the strains at >8 µg/mL CN585. Growth susceptibility assays were performed twice in triplicate.
To further investigate the direct role of immunophilins in binding to CN585 and inhibiting CN, we next generated a cyclophilin A deletion strain (ΔcypA) (Figure S2) and also utilized the FKBP12 (Δfkbp12) deletion strain [37] for susceptibility testing. While the ΔcypA and the Δfkbp12 strains were resistant to CsA and FK506, respectively, no induction in resistance to CN585 was noted in the ΔcypA or the Δfkbp12 strains.
3.3. Hypersensitivity of the A. fumigatus CN Deletion Mutant to CN585 Indicates Off-Target Effects of CN585
In order to test if CN585 specifically inhibited CN function in A. fumigatus, we next compared the susceptibility of the A. fumigatus CN deletion strain (ΔcnaA) [38] to both FK506 and CN585 in RPMI liquid medium for 48 h. While FK506 treatment of the wild-type strain mimicked the CN deletion phenotype, the CN deletion strain was not susceptible to inhibition by FK506 treatment in a range of concentration between 0.1–5 µg/mL (Figure 3A) indicating the absence of target for FK506. Treatment with >5 µg/mL of FK506 also yielded a similar pattern, further confirming the specific action of FK506 on CN. However, the CN deletion strain exhibited a concentration-dependent growth inhibition with CN585 (Figure 3B). The CN deletion strain exhibited drastic reduction in growth at 2 µg/mL of CN585 with almost complete growth inhibition at ≥4 µg/mL CN585. To further examine if CN585 is causing a fungicidal effect, we cultured the wild-type and CN deletion strains on GMM agar medium for 5 days. While CN585 at 10 µg/mL could not mimic the CN inhibition phenotype observed with FK506 at 1 µg/mL in the wild-type strain, there was a drastic growth inhibition of the CN deletion strain in the presence of CN585 (10 µg/mL) as opposed to no inhibition in growth in the presence of FK506 (1 µg/mL) (Figure 3C). These results indicated the possibility of CN585 having off-target antifungal effects in A. fumigatus.
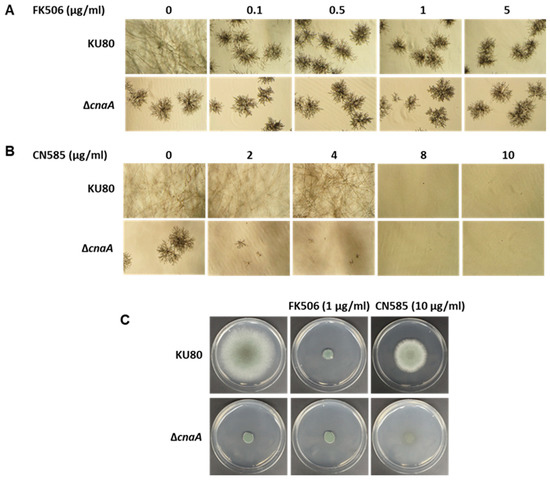
Figure 3.
(A) Growth of the wild-type A. fumigatus (KU80) and the CnA deletion (ΔcnaA) strains in the presence of increasing concentrations of FK506. Note that the ΔcnaA strain is not susceptible to growth inhibition by FK506. (B) Growth of the wild-type A. fumigatus (KU80) and the CnA deletion (ΔcnaA) strains in the presence of increasing concentrations of CN585. Note that the ΔcnaA strain is sensitive to CN585 with complete inhibition in growth at >4 µg/mL CN585. Growth was monitored for 2 days in RPMI at 37 °C. Light microscopy photographs (×10 magnification) after 2 days of growth are shown. (C) Comparative growth of the wild-type A. fumigatus (KU80) and the CnA deletion (ΔcnaA) strains in the presence of FK506 and CN585 on GMM agar for 5 days at 37 °C. Note that the ΔcnaA strain is sensitive to CN585 with complete inhibition in growth at 10 µg/mL CN585 in contrast to FK506. Growth susceptibility assays were performed in triplicate.
3.4. CN585-Mediated Growth Inhibition Is Independent of the CN-Dependent Transcription Factor CrzA
CN585 was shown to inhibit nuclear translocation of GFP-NFAT in HeLa cells [22]. As Crz1 is a well-known NFAT ortholog in fungi [39,40], we utilized an A. fumigatus strain expressing CrzA-GFP fusion protein to verify the effect of CN585 on CrzA’s localization in vivo. Cultures grown for 20 h under control conditions were incubated with 6 µg/mL CN585 for various time periods (10, 30 and 120 min, respectively) and stimulated by the addition of 50 mM CaCl2 for 5–60 min. Simultaneously, FK506 treatment (100–500 ng/mL) was also performed as a comparison. While the addition of CaCl2 under control conditions resulted in nuclear translocation of CrzA-GFP within 5 min (Figure 4A), the addition of FK506 blocked CrzA’s nuclear translocation indicating the inhibition of CN signaling by FK506 (Figure 4B). However, CN585 did not have any impact on the nuclear localization of CrzA (Figure 4C), further indicating CN and CrzA-independent growth inhibition by CN585.
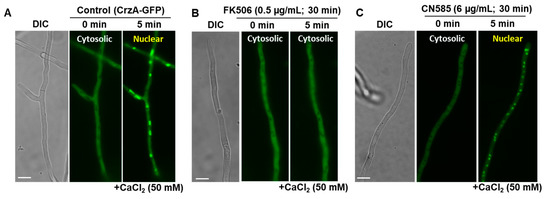
Figure 4.
Microscopic localization CrzA-GFP in A. fumigatus in the absence or presence of FK506 and CN585. (A) Nuclear localization of CrzA-GFP under control conditions. (B) Inhibition of nuclear localization of CrzA by FK506. (C) Nuclear localization of CrzA-GFP under CN585 treated conditions. DIC-Differential Interference Contrast; Scale bar, 10 µm.
3.5. Molecular Docking of CN585 Reveals Differential Potential Binding Sites between A. fumigatus and Human CN
Next, to structurally characterize a possible molecular interaction of CN585 with A. fumigatus and human CN we utilized a complex usage of MD simulations and molecular docking protocols. This method ensured a more accurate description of the molecular dynamics a protein and small molecule undergo prior to and in complex when in solution compared to the more traditional static docking models. The 100 ns MD simulation showed equilibration after 10 ns of the 100 ns simulation indicative of a well performed MD simulation (Figure 5A). For the most part, the Cα-RMSD, radius of gyration (Rg), and center of mass between (COM) CnA and CnB stabilized after ~10 ns. Interestingly the results show that HADDOCK blind molecular docking predicted two different binding sites for CN585 when docked against A. fumigatus and human CN (Figure 5B). When docked to human CN, CN585 overlays with the binding site of CsA and solely interacts with CnA (Figure 5B right), whereas when docked to A. fumigatus CN585 bridges an interface between CnA and CnB at a site allosteric to the known CsA binding site (Figure 5B left).
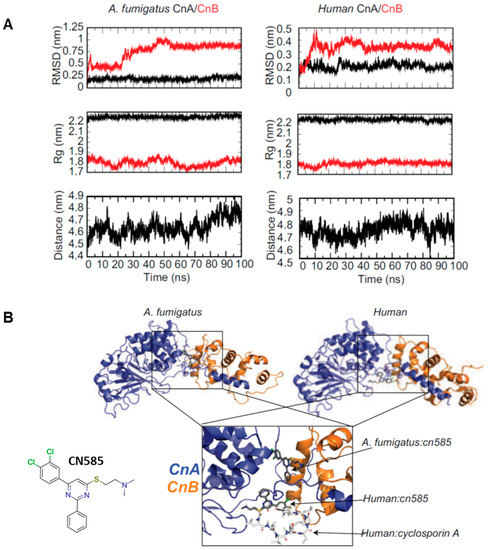
Figure 5.
Molecular dynamics simulation and docking of A. fumigatus and human CnA/CnB to CN585. (A) MD simulation analysis showing the RMSD (top), Rg (middle), and COM of CnA to CnB (bottom) of CnA/CnB during the simulation. (B) Docking model of A. fumigatus CnA/CnB:CN585 (left—CnA (blue) and CnB (orange) compared to human CnA/CnB:CN585 (right—CnA (blue) and CnB (orange). Overlay of the docking poses of CN585 and CsA is shown at the bottom. Labels provided for clarity.
To supplement the docking analysis, target prediction for CN585 was performed using chemoinformatics resources including the Similarity Ensemble Approach (SEA) [41] and Swiss Target Prediction (STP) [42,43]. These methods yielded predictions based on Tanimoto coefficient similarity to known ligands of a biological target. The results of FK506 and CN585 were analyzed for prediction of interactions with CN or FKBPs. As expected, FK506 did show predicted interaction with FKBPs (Figures S3 and S4), whereas no molecules with high Tanimoto coefficient similarity to CN585 were found to interact with CN or FKBPs (Figure S5). The STP of CN585 also does not suggest any interactions with any phosphatase or any FKBP (Figure S6). Interestingly CN585 showed molecular similarity to ligands of a eukaryotic translation initiation factor 4E-binding protein (eIF4E) and other proteins.
3.6. Molecular Modeling and Docking of CN585 Reveals Similar Potential Binding Sites between A. fumigatus and Human eIF4E:m7GTP
Next to explore a possible interaction of CN585 with eIF4E we first performed the molecular modeling of A. fumigatus eIF4E which resulted in a model that overlaid with Ascaris suum eIF4E:m7GTP (3M93) with a Cα-RMSD of 1.7 Å. To structurally characterize the molecular interaction of CN585 with A. fumigatus eIF4E we utilized the same MD simulation and molecular docking protocol as above. The 100 ns MD simulation showed equilibration after 10 ns of the 100 ns simulation indicative of a well performed MD simulation (Figure 6A)—the Cα-RMSD and Rg stabilized after 10 ns. The HADDOCK blind docking results predicted an identical binding site as m7GTP with A. suum eIF4E (Figure 6B). When docked to A. fumigatus eIF4E, CN585 overlays with the binding site of m7GTP; however, while in slightly different orientation the aromatic moieties of both CN585 and m7GTP overlay in nearly identical orientations.
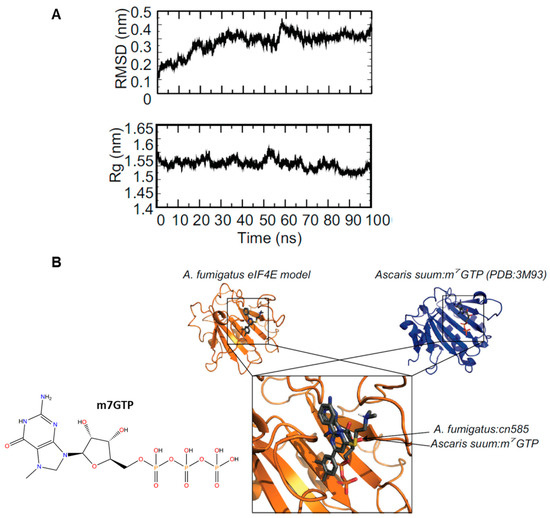
Figure 6.
Molecular dynamics simulation and docking of A. fumigatus eIF4E to CN585. (A) MD simulation analysis showing the RMSD (top) and Rg (middle) of A. fumigatus eIF4E during the simulation. (B) Docking model of A. fumigatus eIF4E: CN585 (orange—left) compared to Ascaris suum eIF4E:m7GTP (blue—right). Overlay of the docking poses of CN585 and m7GTP is shown at the bottom. Labels provided for clarity.
4. Discussion
Due to high structural and conformational similarity between the active site of CnA and other serine/threonine protein phosphatases, other low-molecular weight inhibitors of these phosphatases are also known to inhibit CN [44]. To better understand fungal CN inhibitory mechanisms, here we analyzed the activity of a novel mammalian CN inhibitor, CN585, on A. fumigatus. CN585 belongs to a new class of CN inhibitors and characterized as an immunosuppressive and non-competitive reversible inhibitor of CN in mammalian cells [22].
FK506 and CsA are known to inhibit CN by binding to their respective immunophilins FKBP12 and CypA [14,15]. Unexpectedly, we found that in contrast to FK506 the fungal growth inhibition pattern by CN585 was completely different from the usual stunted and hyperbranching phenotype observed upon CN inhibition by FK506 or CsA (Figure 1). For generalizability, we found that the A. fumigatus wild-type, azole- and echinocandin-resistant strains, and the various CN mutants showing resistance to FK506/CsA, were equally sensitive to CN585 (Figure 2). As in vitro susceptibility assays with the various CN pathway mutants indicated an unexpected pattern of growth inhibition by CN585, we speculated if CN585 was indeed specifically inhibiting CN function in A. fumigatus. These initial findings indicated that CN585 may probably inhibit CN and also other targets in the cell to bring about complete growth inhibition not normally observed with FK506/CsA treatment. While the CN catalytic subunit deletion strain (ΔcnaA) exhibited hypersensitivity to CN585 in comparison to FK506 (Figure 3), the nuclear localization of the CN-dependent transcription factor CrzA was also not blocked by CN585 (Figure 4), indicating the possibility of it being non-specific and having off-target effects in A. fumigatus leading to antifungal activity. Furthermore, as we have established the requirement of the calcineurin regulatory subunit, CnB, for the function of CnA and shown the phenotypic similarity between the CnA deletion and the CnB deletion strains [38], we expect that the CnB deletion strain strain would also exhibit similar susceptibility to CN585. Taken together, these findings revealed that CN585 also does not bind to the immunophilins to inhibit CN, but may inhibit CN through another alternative mechanism, interaction with other yet unknown residues in CnA, or interaction with other yet unknown target(s).
Based on the molecular docking assessments of A. fumigatus and human CN, CN585 binds to different sites when comparing docking to A. fumigatus and human; however, the docking scores have similar values to one another and compared to binding of FK506 or CsA. Structurally, when CN585 is docked to human CN it binds in a manner almost identical to that of CsA and has many of the same interactions with CnA residues as CsA binding (Figure 5B). Both CN585 and CsA interact with residues L312, Y341, W342, L343, P344 and W352 on human CnA. While the docked CN585 did not interact with CnB, CsA is known to interact with CnB in a limited manner. The list of residues that interact with CN585 in the docked model are remarkably similar to residues shown as essential to confer resistance to CsA (but not FK506) [45]. Specifically, V314R and Y341F mutations have shown to confer resistance to CsA, while modification of CsA has been proposed to take advantage to include a hydrogen bond donor for the carbonyl of W342 and/or L312 [46]. Interestingly though, the molecular docking of CN585 suggested a completely different site of binding for CN585 when docked to CN from A. fumigatus (Figure 5A-left). This docking suggested the A. fumigatus binding site is in the interface of CnA and CnB allosteric to the known FK506 or CsA binding site, approximately 16 Å away. There is only one conserved contact with CN585 docking to A. fumigatus CN (W364) observed in the docking with human CN, W342. However, we did not observe any CN585 resistance in our W364A mutant (Figure 2). The rest of the contacts in human CN were with residues A21-R53 and D346 of CnA and residues M118, E138 and N158 of CnB. As the docked CN585 structure does not have many similar CsA contacts, it is possible the binding site observed in the A. fumigatus:CN585 model would not sterically inhibit the binding of FK506 and/or CsA, therefore in this conformation cannot be directly attributed to inhibitory phenotypes and/or the allosteric binding site. Based on this we hypothesize that CN585 does not in actuality interact with A. fumigatus CN.
Interestingly, we also did not find FKBPs as potential interactors of CN585 using the chemoinformatics resources SEA or STP. While FK506 was predicted to interact with FKBPs, no molecules with high Tanimoto coefficient similarity to CN585 were found to interact with CN or FKBPs. Ultimately, neither the structure-based nor ligand-base techniques support a hypothesis of CN585 directly interacting with A. fumigatus CN. The computational method utilized here still leaves the possibility that CN585’s antifungal properties may be due to its potential interaction with other proteins that are important for fungal growth.
As such, CN585 was molecularly docked to a model of the eIF4E sequence from A. fumigatus (the top hit from STP) to compare its predicted binding site in comparison with known X-ray characterized structure of eIF4E in complex with m7GTP in an identical manner to CN585 docking to A. fumigatus and Human CN. The docking showed a near identical binding site sharing many of the similar/identical residues between the A. fumigatus and Ascaris suum sequences (Figure 6B). Despite the differences in the chemical composition between m7GTP and CN585, these two structures share similar contacts (Figure 6B). A. fumigatus eIF4E:CN585 had contacts with Y157, P201, V202, W203 and R254 which are similar to contacts between Ascaris suum:m7GTP W69, P113, M114, W115 and R170 when the sequences are aligned (Figure S7).
While CN585 has been shown to inhibit mammalian CN [22], our results described here suggested that CN585 does not appear to specifically inhibit A. fumigatus CN and caution should be exercised in utilizing CN585 as a CN-specific inhibitor in fungi. Further studies are required to understand the exact mode of inhibition of fungal growth by CN585.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/jof8121281/s1. Figure S1: CnaA mutants showing CsA resistance. Figure S2: Generation of cypA deletion verified by Southern analysis and susceptibility to CsA. Figure S3: Swiss Target Prediction for FK506. Figure S4: Similarity Ensemble Approach prediction of FK506. Figure S5: Swiss Target Prediction for CN585. Figure S6: Similarity Ensemble Approach prediction of CN585. Figure S7: Alignment of A. fumigatus and Ascaris suum eIF4.
Author Contributions
Conceptualization, P.R.J. and B.G.B.; Methodology, P.R.J., B.G.B., D.C.C. and M.A.; Validation, P.R.J. and B.G.B.; Formal Analysis, P.R.J., B.G.B., D.C.C. and M.A.; Investigation, P.R.J., B.G.B., D.C.C. and W.J.S.; Resources, P.R.J., B.G.B. and W.J.S.; Data Curation, P.R.J., B.G.B. and D.C.C.; Writing—Original Draft Preparation, P.R.J. and B.G.B.; Writing—Review and Editing, P.R.J., B.G.B., M.A. and W.J.S.; Visualization, P.R.J., B.G.B. and D.C.C.; Supervision, P.R.J.; Project Administration, P.R.J. and W.J.S.; Funding Acquisition, P.R.J. and W.J.S. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by funding from the National Institutes of Health—National Institute of Allergy and Infectious Diseases, grant R21AI163301 to P.R.J. and W.J.S.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Acknowledgments
We would like to the thank David Denning and David Perlin for providing the A. fumigatus azole-resistant and echinocandin-resistant strains.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Park, H.S.; Lee, S.C.; Cardenas, M.E.; Heitman, J. Calcium-Calmodulin-Calcineurin Signaling: A Globally Conserved Virulence Cascade in Eukaryotic Microbial Pathogens. Cell Host Microbe 2019, 26, 453–462. [Google Scholar] [CrossRef] [PubMed]
- Hemenway, C.; Heitman, J. Calcineurin. Cell Biochem. Biophys. 1999, 30, 115–151. [Google Scholar] [CrossRef] [PubMed]
- Creamer, T.P. Calcineurin. Cell Commun. Signal. 2020, 18, 137. [Google Scholar] [CrossRef] [PubMed]
- Shibasaki, F.; Hallin, U.; Uchino, H. Calcineurin as a multifunctional regulator. J. Biochem. 2002, 131, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Pallen, C.J.; Wang, J.H. A multifunctional calmodulin-stimulated phosphatase. Arch. Biochem. Biophys. 1985, 237, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Juvvadi, P.R.; Lee, S.C.; Heitman, J.; Steinbach, W.J. Calcineurin in fungal virulence and drug resistance: Prospects for harnessing targeted inhibition of calcineurin for an antifungal therapeutic approach. Virulence 2017, 8, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Rusnak, F.; Mertz, P. Calcineurin: Form and Function. Physiol. Rev. 2000, 80, 1483–1521. [Google Scholar] [CrossRef]
- Hogan, P.G. Calcium-NFAT transcriptional signalling in T cell activation and T cell exhaustion. Cell Calcium 2017, 63, 66–69. [Google Scholar] [CrossRef]
- Grigoriu, S.; Bond, R.; Cossio, P.; Chen, J.A.; Ly, N.; Hummer, G.; Page, R.; Cyert, M.S.; Peti, W. The Molecular Mechanism of Substrate Engagement and Immunosuppressant Inhibition of Calcineurin. PLoS Biol. 2013, 11, e1001492. [Google Scholar] [CrossRef]
- Gwack, Y.; Feske, S.; Srikanth, S.; Hogan, P.G.; Rao, A. Signalling to transcription: Store-operated Ca2+ entry and NFAT activation in lymphocytes. Cell Calcium 2007, 42, 145–156. [Google Scholar] [CrossRef]
- Ho, S.; Clipstone, N.; Timmermann, L.; Northrop, J.; Graef, I.; Fiorentino, D.; Nourse, J.; Crabtree, G.R. The Mechanism of Action of Cyclosporin A and FK506. Clin. Immunol. Immunopathol. 1996, 80, S40–S45. [Google Scholar] [CrossRef] [PubMed]
- Kawamura, A.; Su, M.S.-S. Interaction of FKBP12-FK506 with Calcineurin A at the B Subunit-binding Domain. J. Biol. Chem. 1995, 270, 15463–15466. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Farmer, J.D., Jr.; Lane, W.S.; Friedman, J.; Weissman, I.; Schreiber, S.L. Calcineurin is a common target of cyclophilin-cyclosporin A and FKBP-FK506 complexes. Cell 1991, 66, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Griffith, J.P.; Kim, J.L.; Kim, E.E.; Sintchak, M.D.; Thomson, J.A.; Fitzgibbon, M.J.; Fleming, M.A.; Caron, P.R.; Hsiao, K.; Navia, M.A. X-ray structure of calcineurin inhibited by the immunophilin-immunosuppressant FKBP12-FK506 complex. Cell 1995, 82, 507–522. [Google Scholar] [CrossRef]
- Huai, Q.; Kim, H.Y.; Liu, Y.; Zhao, Y.; Mondragon, A.; Liu, J.O.; Ke, H. Crystal structure of calcineurin-cyclophilin-cyclosporin shows common but distinct recognition of immunophilin-drug complexes. Proc. Natl. Acad. Sci. USA 2002, 99, 12037–12042. [Google Scholar] [CrossRef]
- Nambu, M.; Covel, J.A.; Kapoor, M.; Li, X.; Moloney, M.K.; Numa, M.M.; Soltow, Q.A.; Trzoss, M.; Webb, P.; Webb, R.R., 2nd; et al. A calcineurin antifungal strategy with analogs of FK506. Bioorg. Med. Chem. Lett. 2017, 27, 2465–2471. [Google Scholar] [CrossRef]
- Lee, Y.; Lee, K.T.; Lee, S.J.; Beom, J.Y.; Hwangbo, A.; Jung, J.A.; Song, M.C.; Yoo, Y.J.; Kang, S.H.; Averette, A.F.; et al. In Vitro and In Vivo Assessment of FK506 Analogs as Novel Antifungal Drug Candidates. Antimicrob. Agents Chemother. 2018, 62, e01627-18. [Google Scholar] [CrossRef]
- Jung, J.A.; Yoon, Y.J. Development of Non-Immunosuppressive FK506 Derivatives as Antifungal and Neurotrophic Agents. J. Microbiol. Biotechnol. 2020, 30, 1–10. [Google Scholar] [CrossRef]
- Juvvadi, P.R.; Fox, D., 3rd; Bobay, B.G.; Hoy, M.J.; Gobeil, S.M.C.; Venters, R.A.; Chang, Z.; Lin, J.J.; Averette, A.F.; Cole, D.C.; et al. Harnessing calcineurin-FK506-FKBP12 crystal structures from invasive fungal pathogens to develop antifungal agents. Nat. Commun. 2019, 10, 4275. [Google Scholar] [CrossRef]
- Gobeil, S.M.C.; Bobay, B.G.; Juvvadi, P.R.; Cole, D.C.; Heitman, J.; Steinbach, W.J.; Venters, R.A.; Spicer, L.D. Leveraging Fungal and Human Calcineurin-Inhibitor Structures, Biophysical Data and Dynamics to Design Selective and Non-Immunosuppressive FK506 Analogs. mBio 2021, 12, e03000-21. [Google Scholar] [CrossRef]
- Hoy, M.J.; Park, E.; Lee, H.; Lim, W.Y.; Cole, D.C.; DeBouver, N.D.; Bobay, B.G.; Pierce, P.G.; Fox, D., 3rd; Ciofani, M.; et al. Structure-Guided Synthesis of FK506 and FK520 Analogs with Increased Selectivity Exhibit In Vivo Therapeutic Efficacy against Cryptococcus. mBio 2022, 13, e0104922. [Google Scholar] [CrossRef] [PubMed]
- Erdmann, F.; Weiwad, M.; Kilka, S.; Karanik, M.; Pätzel, M.; Baumgrass, R.; Liebscher, J.; Fischer, G. The novel calcineurin inhibitor CN585 has potent immunosuppressive properties in stimulated human T cells. J. Biol. Chem. 2010, 285, 1888–1898. [Google Scholar] [CrossRef]
- Shwab, E.K.; Juvvadi, P.R.; Waitt, G.; Soderblom, E.J.; Barrington, B.C.; Asfaw, Y.G.; Moseley, M.A.; Steinbach, W.J. Calcineurin-dependent dephosphorylation of the transcription factor CrzA at specific sites controls conidiation, stress tolerance, and virulence of Aspergillus fumigatus. Mol. Microbiol. 2019, 112, 62–80. [Google Scholar] [CrossRef]
- Waterhouse, A.; Bertoni, M.; Bienert, S.; Studer, G.; Tauriello, G.; Gumienny, R.; Heer, F.T.; de Beer, T.A.P.; Rempfer, C.; Bordoli, L.; et al. SWISS-MODEL: Homology modelling of protein structures and complexes. Nucleic Acids Res. 2018, 46, W296–W303. [Google Scholar] [CrossRef]
- Liu, W.; Jankowska-Anyszka, M.; Piecyk, K.; Dickson, L.; Wallace, A.; Niedzwiecka, A.; Stepinski, J.; Stolarski, R.; Darzynkiewicz, E.; Kieft, J.; et al. Structural basis for nematode eIF4E binding an m(2,2,7)G-Cap and its implications for translation initiation. Nucleic Acids Res. 2011, 39, 8820–8832. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Caselli, E.; Fini, F.; Introvigne, M.L.; Stucchi, M.; Taracila, M.A.; Fish, E.R.; Smolen, K.A.; Rather, P.N.; Powers, R.A.; Wallar, B.J.; et al. 1,2,3-Triazolylmethaneboronate: A Structure Activity Relationship Study of a Class of β-Lactamase Inhibitors against Acinetobacter baumannii Cephalosporinase. ACS Infect. Dis. 2020, 6, 1965–1975. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Harrison, S.C. Crystal structure of human calcineurin complexed with cyclosporin A and human cyclophilin. Proc. Natl. Acad. Sci. USA 2002, 99, 13522–13526. [Google Scholar] [CrossRef]
- van Dijk, M.; Wassenaar, T.A.; Bonvin, A.M. A Flexible, Grid-Enabled Web Portal for GROMACS Molecular Dynamics Simulations. J. Chem. Theory Comput. 2012, 8, 3463–3472. [Google Scholar] [CrossRef]
- van Zundert, G.C.P.; Rodrigues, J.; Trellet, M.; Schmitz, C.; Kastritis, P.L.; Karaca, E.; Melquiond, A.S.J.; van Dijk, M.; de Vries, S.J.; Bonvin, A. The HADDOCK2.2 Web Server: User-Friendly Integrative Modeling of Biomolecular Complexes. J. Mol. Biol. 2016, 428, 720–725. [Google Scholar] [CrossRef]
- Schüttelkopf, A.W.; van Aalten, D.M. PRODRG: A tool for high-throughput crystallography of protein-ligand complexes. Acta Crystallogr. D Biol. Crystallogr. 2004, 60, 1355–1363. [Google Scholar] [CrossRef]
- Steinbach, W.J.; Cramer, R.A., Jr.; Perfect, B.Z.; Henn, C.; Nielsen, K.; Heitman, J.; Perfect, J.R. Calcineurin inhibition or mutation enhances cell wall inhibitors against Aspergillus fumigatus. Antimicrob. Agents Chemother. 2007, 51, 2979–2981. [Google Scholar] [CrossRef] [PubMed]
- Steinbach, W.J.; Schell, W.A.; Blankenship, J.R.; Onyewu, C.; Heitman, J.; Perfect, J.R. In Vitro Interactions between Antifungals and Immunosuppressants against Aspergillus fumigatus. Antimicrob. Agents Chemother. 2004, 48, 1664–1669. [Google Scholar] [CrossRef] [PubMed]
- Howard, S.J.; Cerar, D.; Anderson, M.J.; Albarrag, A.; Fisher, M.C.; Pasqualotto, A.C.; Laverdiere, M.; Arendrup, M.C.; Perlin, D.S.; Denning, D.W. Frequency and evolution of Azole resistance in Aspergillus fumigatus associated with treatment failure. Emerg. Infect. Dis. 2009, 15, 1068–1076. [Google Scholar] [CrossRef] [PubMed]
- Rocha, E.M.; Garcia-Effron, G.; Park, S.; Perlin, D.S. A Ser678Pro substitution in Fks1p confers resistance to echinocandin drugs in Aspergillus fumigatus. Antimicrob. Agents Chemother. 2007, 51, 4174–4176. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Huang, C.; Wang, H.L.; Zhou, K.; Xiao, F.X.; Qun, W. The importance of Loop 7 for the activity of calcineurin. FEBS Lett. 2004, 577, 205–208. [Google Scholar] [CrossRef] [PubMed]
- Juvvadi, P.R.; Pemble, C.W.t.; Ma, Y.; Steinbach, W.J. Novel motif in calcineurin catalytic subunit is required for septal localization of calcineurin in Aspergillus fumigatus. FEBS Lett. 2016, 590, 501–508. [Google Scholar] [CrossRef]
- Falloon, K.; Juvvadi, P.R.; Richards, A.D.; Vargas-Muñiz, J.M.; Renshaw, H.; Steinbach, W.J. Characterization of the FKBP12-Encoding Genes in Aspergillus fumigatus. PLoS ONE 2015, 10, e0137869. [Google Scholar] [CrossRef]
- Juvvadi, P.R.; Fortwendel, J.R.; Rogg, L.E.; Burns, K.A.; Randell, S.H.; Steinbach, W.J. Localization and activity of the calcineurin catalytic and regulatory subunit complex at the septum is essential for hyphal elongation and proper septation in Aspergillus fumigatus. Mol. Microbiol. 2011, 82, 1235–1259. [Google Scholar]
- Stathopoulos, A.M.; Cyert, M.S. Calcineurin acts through the CRZ1/TCN1-encoded transcription factor to regulate gene expression in yeast. Genes Dev. 1997, 11, 3432–3444. [Google Scholar] [CrossRef]
- Soriani, F.M.; Malavazi, I.; da Silva Ferreira, M.E.; Savoldi, M.; Von Zeska Kress, M.R.; de Souza Goldman, M.H.; Loss, O.; Bignell, E.; Goldman, G.H. Functional characterization of the Aspergillus fumigatus CRZ1 homologue, CrzA. Mol. Microbiol. 2008, 67, 1274–1291. [Google Scholar] [CrossRef]
- Keiser, M.J.; Roth, B.L.; Armbruster, B.N.; Ernsberger, P.; Irwin, J.J.; Shoichet, B.K. Relating protein pharmacology by ligand chemistry. Nat. Biotechnol. 2007, 25, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Gfeller, D.; Grosdidier, A.; Wirth, M.; Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: A web server for target prediction of bioactive small molecules. Nucleic Acids Res. 2014, 42, W32–W38. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissTargetPrediction: Updated data and new features for efficient prediction of protein targets of small molecules. Nucleic Acids Res. 2019, 47, W357–W364. [Google Scholar] [CrossRef] [PubMed]
- Erdmann, F.; Weiwad, M. Calcineurin inhibitors: Status quo and perspectives. Biomol. Concepts 2011, 2, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Cardenas, M.E.; Heitman, J. Calcineurin mutants render T lymphocytes resistant to cyclosporin A. Mol. Pharmacol. 1996, 50, 506–511. [Google Scholar] [PubMed]
- Alberg, D.G.; Schreiber, S.L. Structure-based design of a cyclophilin-calcineurin bridging ligand. Science 1993, 262, 248–250. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).