
Peel Diffusion and Antifungal Efficacy of Different Fungicides in Pear Fruit: Structure-Diffusion-Activity Relationships

 ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. General Information and Strains
2.2. Measurement of Residue Distribution
2.3. Computational Studies
2.4. Stability Assay
2.5. In Vitro Antifungal Assay
2.6. In Vivo Antifungal Assay
2.7. Statistical Analysis
3. Results and Discussion
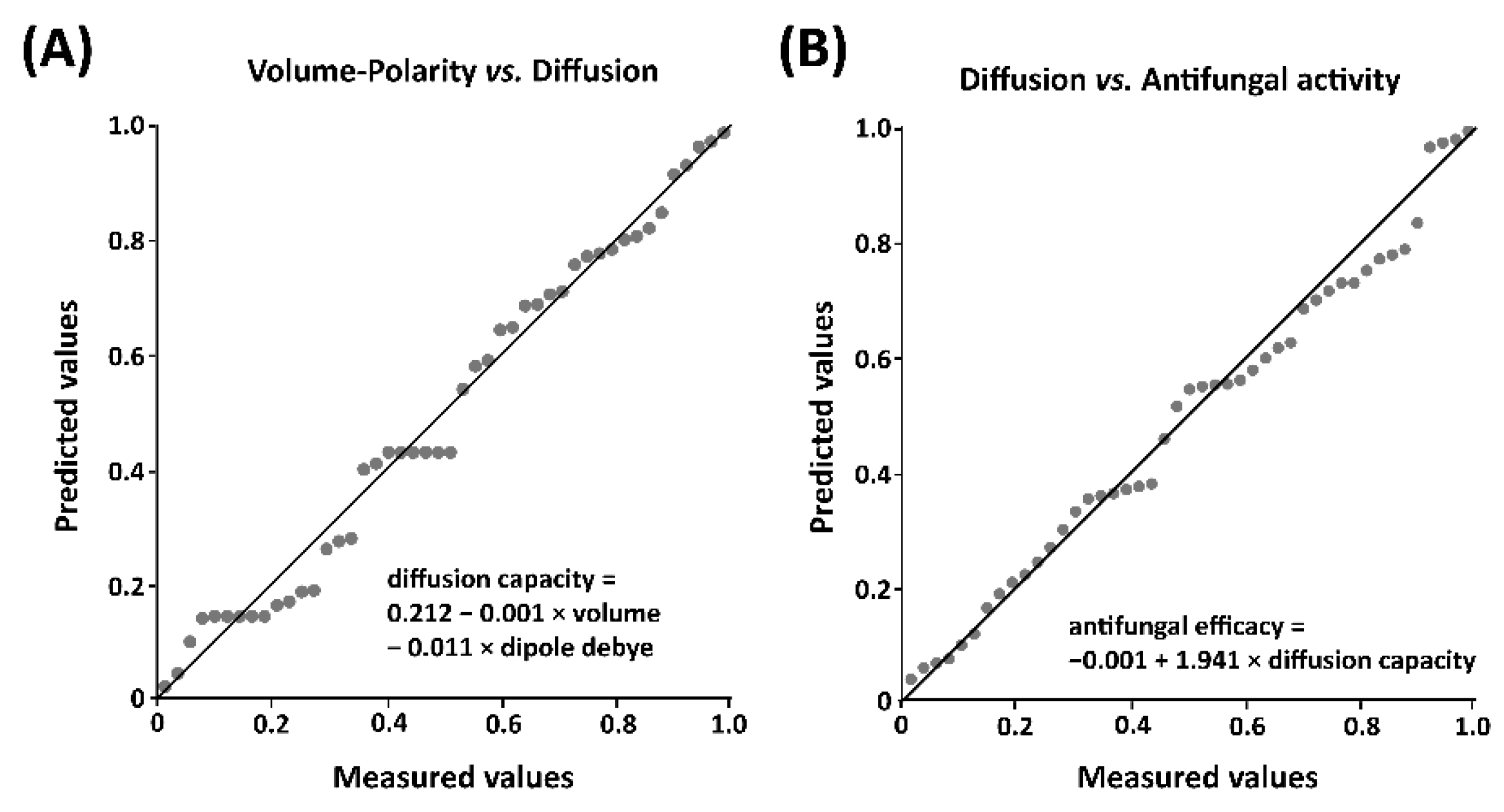
3.1. Correlations between Peel Diffusion and Structural Characteristics of Fungicides
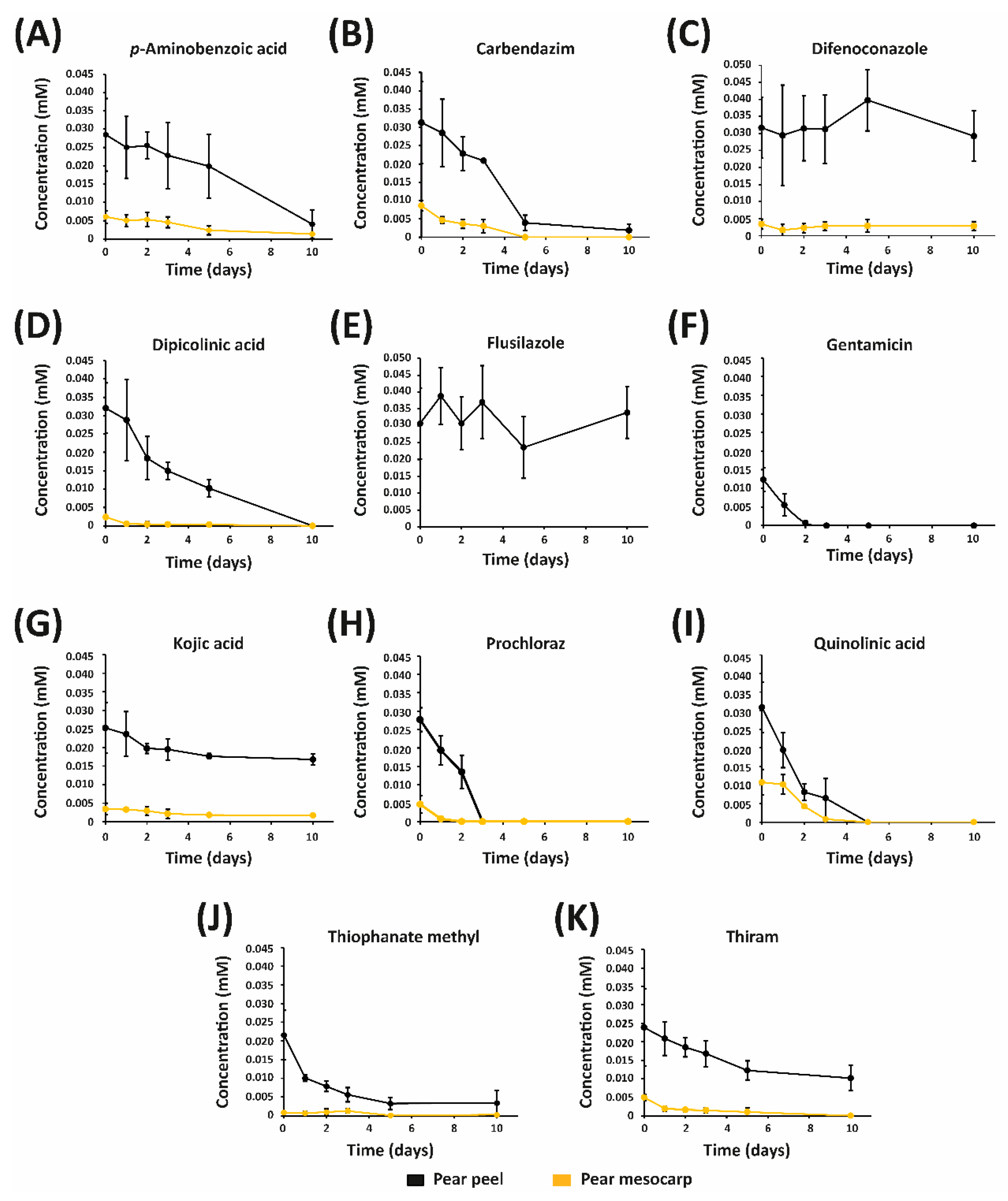
3.2. Stability of Fungicides in Pear Peel and Mesocarp
3.3. Efficacy of Fungicides for the Control of A. alternata in Pear Fruit
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agricultural Organization of the United Nations. 2019. Available online: http://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 13 January 2022).
- Wu, J.; Wang, Y.; Xu, J.; Korban, S.S.; Fei, Z.; Tao, S.; Ming, R.; Tai, S.; Khan, A.M.; Postman, J.D.; et al. Diversification and independent domestication of Asian and European pears. Genome Biol. 2018, 19, 77. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.Y.; Lansky, E.; Kang, S.S.; Yang, M. A review of pears (Pyrus spp.), ancient functional food for modern times. BMC Complement. Med. Ther. 2021, 21, 219. [Google Scholar] [CrossRef] [PubMed]
- Sardella, D.; Muscat, A.; Brincat, J.P.; Gatt, R.; Decelis, S.; Valdramidis, V. A comprehensive review of the pear fungal diseases. Int. J. Fruit Sci. 2016, 16, 351–377. [Google Scholar] [CrossRef]
- Li, C.; Sun, W.; Cao, S.; Hou, R.; Li, X.; Ming, L.; Kan, J.; Zhao, Y.; Liu, F. The CfMK1 gene regulates reproduction, appressorium formation, and pathogenesis in a pear anthracnose-causing fungus. J. Fungi 2022, 8, 77. [Google Scholar] [CrossRef]
- Fisher, M.C.; Hawkins, N.J.; Gurr, S.J. Worldwide emergence of resistance to antifungal drugs challenges human health and food security. Science 2018, 360, 739–742. [Google Scholar] [CrossRef] [Green Version]
- Pan, T.T.; Pu, H.B.; Sun, D.W. Insights into the changes in chemical compositions of the cell wall of pear fruit infected by Alternaria alternata with confocal Raman microspectroscopy. Postharvest Biol. Technol. 2017, 132, 119–129. [Google Scholar] [CrossRef]
- Yang, X.P.; Hu, H.J.; Yu, D.Z.; Sun, Z.H.; He, X.J.; Zhang, J.G.; Chen, Q.L.; Tian, R.; Fan, J.; Liu, J.H. Candidate resistant genes of sand pear (Pyrus pyrifolia nakai) to Alternaria alternata revealed by transcriptome sequencing. PLoS ONE 2015, 10, e0135046. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, Y.; Li, Y.; Bi, Y.; Mao, R.; Yang, Y.; Jiang, Q.; Prusky, D. Cellular responses required for oxidative stress tolerance of the necrotrophic fungus Alternaria alternata, causal agent of pear black spot. Microorganisms 2022, 10, 621. [Google Scholar] [CrossRef]
- Armitage, A.D.; Cockerton, H.M.; Sreenivasaprasad, S.; Woodhall, J.; Lane, C.R.; Harrison, R.J.; Clarkson, J.P. Genomics evolutionary history and diagnostics of the Alternaria alternata species group including apple and Asian pear pathotypes. Front. Microbiol. 2020, 10, 3124. [Google Scholar] [CrossRef] [Green Version]
- Hao, W.; Forster, H.; Elkins, R.B.; Adaskaveg, J.E. First report of Phytophthora lacustris causing a fruit rot of Bartlett pear in North America. Plant Dis. 2020, 104, 2528. [Google Scholar] [CrossRef]
- Kock, S.L.; Holz, G. Blossom-end rot of pears: Systemic infection of flowers and immature fruit by Botrytis cinerea. J. Phytopathol. 1992, 135, 317–327. [Google Scholar] [CrossRef]
- Zhang, M.; Wang, T.; Li, Y.; Bi, Y.; Li, R.; Yuan, J.; Xu, W.; Prusky, D. AaHog1 regulates infective structural differentiation mediated by physicochemical signals from pear fruit cuticular wax, stress response, and Alternaria alternata pathogenicity. J. Fungi 2022, 8, 266. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Li, Y.C.; Bi, Y.; Wang, Y. Role of pear fruit cuticular wax and surface hydrophobicity in regulating the prepenetration phase of Alternaria alternata infection. J. Phytopathol. 2017, 165, 313–322. [Google Scholar] [CrossRef]
- Zhang, M.; Li, Y.C.; Wang, T.L.; Bi, Y.; Li, R.; Huang, Y.; Mao, R.Y.; Jiang, Q.Q.; Liu, Y.X.; Prusky, D.B. AaPKAc regulates differentiation of infection structures induced by physicochemical signals from pear fruit cuticular wax, secondary metabolism, and pathogenicity of Alternaria alternata. Front. Plant Sci. 2021, 12, 642601. [Google Scholar] [CrossRef]
- Lutz, M.C.; Sosa, M.C.; Colodner, A.D. Effect of pre and postharvest application of fungicides on postharvest decay of Bosc pear caused by Alternaria–Cladosporium complex in North Patagonia, Argentina. Sci. Hortic. 2017, 225, 810–817. [Google Scholar] [CrossRef]
- Sugar, D.; Basile, S.R. Orchard calcium and fungicide treatments mitigate effects of delayed postharvest fungicide applications for control of postharvest decay of pear fruit. Postharvest Biol. Technol. 2011, 60, 52–56. [Google Scholar] [CrossRef]
- Lu, M.J.; Yu, C.X.; Cui, X.Y.; Shi, J.Y.; Yuan, L.; Sun, S.J. Gentamicin sinergises with azoles against drug-resistance Candida albicans. Int. J. Antimicrob. Agents 2018, 51, 107–114. [Google Scholar] [CrossRef]
- Song, X.G.; Han, M.H.; He, F.; Wang, S.Y.; Li, C.H.; Wu, G.C.; Huang, Z.G.; Liu, D.; Liu, F.Q.; Laborda, P.; et al. Antifungal mechanism of dipicolinic acid and its efficacy for the control of pear Valsa canker. Front. Microbiol. 2020, 11, 958. [Google Scholar] [CrossRef]
- Chen, Y.; Zhou, Y.D.; Laborda, P.; Wang, H.L.; Wang, R.; Chen, X.; Liu, F.Q.; Yang, D.J.; Wang, S.Y.; Shi, X.C.; et al. Mode of action and efficacy of quinolinic acid for the control of Ceratocystis fimbriata on sweet potato. Pest Manag. Sci. 2021, 77, 4564–4571. [Google Scholar] [CrossRef]
- Laborda, P.; Zhao, Y.; Ling, J.; Hou, R.; Liu, F. Production of antifungal p-aminobenzoic acid in Lysobacter antibioticus OH13. J. Agric. Food Chem. 2018, 66, 630–636. [Google Scholar] [CrossRef]
- Saeedi, M.; Eslamifar, M.; Khezri, K. Kojic acid applications in cosmetic and pharmaceutical preparations. Biomed. Pharmacother. 2019, 110, 582–593. [Google Scholar] [CrossRef] [PubMed]
- Fang, Q.; Wu, R.; Hu, G.; Lai, A.; Wu, K.; Zhang, L.; Feng, J.; Cao, H. Dissipation behavior, residue distribution and risk assessment of three fungicides in pears. J. Sci. Food Agric. 2020, 100, 1757–1763. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Chen, X.; Liu, F.; Pan, C. Residue distribution, dissipation behavior, and removal of four fungicide residues on harvested apple after waxing treatment. J. Agric. Food Chem. 2019, 67, 2307–2312. [Google Scholar] [CrossRef] [PubMed]
- Shimshoni, J.A.; Bommuraj, V.; Chena, Y.; Sperling, R.; Barel, S.; Kaye, Y.; Fallik, E. Residual distribution kinetics of pesticides in cherry tomato peel, pulp, and fruit as a function of irrigation water salinity, household rinsing, and storage regimen. Agronomy 2019, 9, 800. [Google Scholar] [CrossRef] [Green Version]
- Utture, S.C.; Banerjee, K.; Dasgupta, S.; Patil, S.H.; Jadhav, M.R.; Wagh, S.S.; Kolekar, S.S.; Anuse, M.A.; Adsule, P.G. Dissipation and distribution behaviour of azoxystrobin, carbendazim, and difenoconazole in pomegranate fruits. J. Agric. Food Chem. 2011, 59, 7866–7873. [Google Scholar] [CrossRef] [PubMed]
- Laborda, P.; Li, C.; Zhao, Y.; Tang, B.; Ling, J.; He, F.; Liu, F. Antifungal metabolite p-aminobenzoic acid (pABA): Mechanism of action and efficacy for the biocontrol of pear bitter rot disease. J. Agric. Food Chem. 2019, 67, 2157–2165. [Google Scholar] [CrossRef]
- He, F.; Li, B.; Ai, G.; Kange, A.M.; Zhao, Y.; Zhang, X.; Jia, Y.; Dou, D.; Liu, F.; Cao, H. Transcriptomic analysis of the Chinese pear pathotype of Alternaria alternata give insights into novel mechanisms of HSAF antifungal activities. Int. J. Mol. Sci. 2018, 19, 1841. [Google Scholar] [CrossRef] [Green Version]
- Di Muccio, A.; Girolimetti, S.; Barbini, D.A.; Pelosi, P.; Generali, T.; Vergori, L.; De Merulis, G.; Leonelli, A.; Stefanelli, P. Selective clean-up applicable to aqueous acetone extracts for the determination of carbendazim and thiabendazole in fruits and vegetables by high-performance liquid chromatography with UV detection. J. Chromatogr. A 1999, 833, 61–65. [Google Scholar] [CrossRef]
- Laborda, P.; Ling, J.; Chen, X.; Liu, F. ACC deaminase from Lysobacter gummosus OH17 can promote root growth in Oryza sativa Nipponbare plants. J. Agric. Food Chem. 2018, 66, 3675–3682. [Google Scholar] [CrossRef]
- Noore, S.; Ramesh, G.; Vendan, S.E.; Nagaraju, V.D. Persistence and diffusion behavior of chlopyrifos in five different species of vegetables: A comparative analysis. Ecotoxicol. Environ. Saf. 2021, 217, 112208. [Google Scholar] [CrossRef]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Mennucci, B.; Petersson, G.A.; et al. Gaussian 09 Rev. D01; Gaussian, Inc.: Wallingford, CT, USA, 2009. [Google Scholar]
- Zhao, Y.; Truhlar, D.G. The M06 suite of density functionals for main group thermochemistry, thermochemical kinetics, noncovalent interactions, excited states, and transition elements: Two new functionals and systematic testing of four M06-class functionals and 12 other functionals. Theor. Chem. Acc. 2008, 120, 215–241. [Google Scholar]
- Marenich, A.V.; Cramer, C.J.; Truhlar, D.G. Universal solvation model based on solute electron density and a continuum model of the solvent defined by the bulk dielectric constant and atomic surface tensions. J. Phys. Chem. B 2009, 113, 6378–6396. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.Y.; Shi, X.C.; Wang, S.Y.; Wang, B.; Laborda, P. Antifungal mechanism and efficacy of kojic acid for the control of Sclerotinia sclerotiorum in soybean. Front. Plant Sci. 2022, 13, 845698. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.K.; Ediriweera, M.K.; Davaatseren, M.; Hyun, H.B.; Cho, S.K. Antioxidant activity of banana flesh and antiproliferative effect on breast and pancreatic cancer cells. Food Sci. Nutr. 2022, 3, 740–750. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Li, Y.; Liu, Q.; Gao, K. Antimicrobial activities of some thymol derivatives from the roots of Inula hupehensis. Food Chem. 2010, 120, 512–516. [Google Scholar] [CrossRef]
- Wang, K.B.; Jiang, S.S.; Pu, T.; Fan, L.M.; Su, F.W.; Ye, M. Antifungal activity of phenolic monoterpenes and structure-related compounds against plant pathogenic fungi. Nat. Prod. Res. 2019, 33, 1423–1430. [Google Scholar] [CrossRef]
- Ahmadi, A.; Mohammadnejadi, E.; Karami, P.; Razzaghi-Asl, N. Current status and structure activity relationship of privileged azoles as antifungal agents (2016–2020). Int. J. Antimicrob. Agents 2022, 59, 106518. [Google Scholar] [CrossRef]
- Qing, Z.X.; Yang, P.; Tang, Q.; Cheng, P.; Liu, X.B.; Zheng, Y.J.; Liu, Y.S.; Zeng, J.G. Isoquinoline alkaloids and their antiviral, antibacterial, and antifungal activities and structure-activity relationship. Curr. Org. Chem. 2017, 21, 1920–1934. [Google Scholar] [CrossRef]
- Calvaruso, E.; Cammilleri, G.; Pulvirenti, A.; Lo Dico, G.M.; Lo Cascio, G.; Giaccone, V.; Badaco, V.V.; Ciprì, V.; Alessandra, M.M.; Vella, A.; et al. Residues of 165 pesticides in citrus fruits using LC-MS/MS: A study of the pesticides distribution from the peel to the pulp. Nat. Prod. Res. 2020, 34, 34–38. [Google Scholar] [CrossRef]
- Yu, H.; Gu, L.; Chen, L.; Wen, H.; Zhang, D.; Tao, H. Activation of grapefruit derived biochar by its peel extracts and its performance for tetracycline removal. Bioresour. Technol. 2020, 316, 123971. [Google Scholar] [CrossRef]
- Bose, S.; Ghosh, A.; Das, A.; Rahaman, M. Development of mango peel derived activated carbon-nickel nanocomposite as an adsorbent towards removal of heavy metal and organic dye removal from aqueous solution. ChemistrySelect 2020, 5, 14168–14176. [Google Scholar] [CrossRef]
- Ates, A.; Oymak, T. Characterization of persimmon fruit peel and its biochar for removal of methylene blue from aqueous solutions: Thermodynamic, kinetic and isotherm studies. Int. J. Phytoremediation 2020, 22, 607–616. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.B.L.; Priyantha, N.; Latip, S.A.A.; Lu, Y.; Mahadi, A.H. Converting Hylocereus undatus (white dragon fruit) peel waste into a useful potential adsorbent for the removal of toxic Congo red dye. Desalination Water Treat. 2020, 185, 307–317. [Google Scholar] [CrossRef]
- Trivedi, P.; Nguyen, N.; Hykkerud, A.L.; Haggman, H.; Martinussen, I.; Jaakola, L.; Karppinen, K. Developmental and environmental regulation of cuticular wax biosynthesis in fleshy fruits. Front. Plant Sci. 2019, 10, 431. [Google Scholar] [CrossRef] [PubMed]
- Khanal, B.P.; Grimm, E.; Knoche, M. Russeting in apple and pear: A plastic periderm replaces a stiff cuticle. AOB Plants 2013, 5, pls048. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Kamitakahara, H.; Murayama, H.; Ohsako, T.; Itai, A. Analysis of fruit lignin content, composition, and linkage types in pear cultivars and related species. J. Agric. Food Chem. 2020, 68, 2493–2505. [Google Scholar] [CrossRef]
- Kun, D.; Pukanszky, B. Polymer/lignin blends: Interactions, properties, applications. Eur. Polym. J. 2017, 93, 618–641. [Google Scholar] [CrossRef] [Green Version]
- Burghardt, M.; Friedmann, A.; Schreiber, L.; Riederer, M. Modelling the effects of alcohol ethoxylates on diffusion of pesticides in the cuticular wax of Chenopodium album leaves. Pest Manag. Sci. 2006, 62, 137–147. [Google Scholar] [CrossRef]
- Baur, P.; Buchholz, A.; Schonherr, J. Diffusion in plant cuticles as affected by temperature and size of organic solutes: Similarity and diversity among species. Plant Cell Environ. 1997, 20, 982–994. [Google Scholar] [CrossRef]
- Chan, T.C.; Li, H.T.; Li, K.Y. Effects of shapes of solute molecules on diffusion: A study of dependences on solute size, solvent, and temperature. J. Phys. Chem. B 2015, 119, 15718–15728. [Google Scholar] [CrossRef]
- Zhao, H.Y.; Yang, G.L.; Liu, Y.L.; Ye, H.P.; Qi, X.J.; Wang, Q. Residual behaviour and risk assessment of prochloraz in bayberries and bayberry wine for the Chinese population. Environ. Monit. Assess. 2019, 191, 644. [Google Scholar] [CrossRef] [PubMed]
- Dong, F.; Liu, X.; Zheng, Y.; Cao, Q.; Li, C. Stereoselective degradation of fungicide triadimenol in cucumber plants. Chirality 2010, 22, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Pan, X.; Tao, Y.; Jiang, D.; Chen, Z.; Dong, F.; Xu, F.; Liu, X.; Wu, X.; Zheng, Y. Systematic evaluation of chiral fungicide imazalil and its major metabolite R14821 (imazil-M): Stability of enantiomers, enantioselective bioactivity, aquatic toxicity, and dissipation in greenhouse vegetables and soil. J. Agric. Food Chem. 2019, 67, 11331–11339. [Google Scholar] [CrossRef] [PubMed]
- Nallathambi, P.; Umamaheswari, C.; Thakore, B.B.L.; More, T.A. Post-harvest management of ber (Ziziphus mauritiana Lamk) fruit rot (Alternaria alternata Fr. Keissler) using Trichoderma species, fungicides and their combinations. Crop Prot. 2009, 28, 525–532. [Google Scholar] [CrossRef]
- Meena, R.P.; Saran, P.W.L.; Kalariya, K.A.; Manivel, P. Efficacy of fungicides and plant extracts against Alternaria alternata causing leaf blight of chandrasur (Lepidium sativum). Indian J. Agric. Res. 2020, 90, 337–340. [Google Scholar]
- Wang, F.; Saito, S.; Michailides, T.J.; Xiao, C.L. Fungicide resistance in Alternaria alternata from blueberry in California and its impact on control of Alternaria rot. Plant Dis. 2022, 106, 1446–1453. [Google Scholar] [CrossRef]
- Forster, H.; Luo, Y.; Hou, L.; Adaskaveg, J. Mutations in Sdh gene subunits confer different cross resistance patterns to SDHI fungicides in Alternaria alternata causing Alternaria leaf spot of almond in California. Plant Dis. 2022; in press. [Google Scholar] [CrossRef]
- Prusky, D.; Kobiler, I.; Akerman, M.; Miyara, I. Effect of acidic solutions and acidic prochloraz on the control of postharvest decay caused by Alternaria alternata in mango and persimmon fruit. Postharvest Biol. Technol. 2006, 42, 134–141. [Google Scholar] [CrossRef]
- Wang, H.; Huang, Y.; Wang, J.; Chen, X.; Wei, K.; Wang, M.; Shang, S. Activities of azoxystrobin and difenoconazole against Alternaria alternata and their control efficacy. Crop Prot. 2016, 90, 54–58. [Google Scholar] [CrossRef]
- Vujanovic, V.; Hamel, C.; Jabaji-Hare, S.; St-Arnaud, M. Development of a selective myclobutanil agar (MBA) medium for the isolation of Fusarium species from asparagus fields. Can. J. Microbiol. 2002, 48, 841–847. [Google Scholar] [CrossRef]
- Wang, S.Y.; Shi, X.C.; Liu, F.Q.; Laborda, P. Chromatographic methods for the detection and quantification of carbendazim in food. J. Agric. Food Chem. 2020, 68, 11880–11894. [Google Scholar] [CrossRef] [PubMed]
- Lundvist, J.; Hellman, B.; Oskarsson, A. Fungicide prochloraz induces oxidative stress and DNA damage in vitro. Food Chem. Toxicol. 2016, 91, 36–41. [Google Scholar] [CrossRef] [PubMed]
- van Dartel, D.A.M.; Pennings, J.L.A.; de la Fonteyne, L.J.J.; Brauers, K.J.J.; Claessen, S.; van Delft, J.H.; Kleinjans, J.C.S.; Piersma, A.H. Concentration-dependent gene expression responses to flusilazole in embryonic stem cell differentation cultures. Toxicol. Appl. Pharmacol. 2011, 251, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Mu, X.Y.; Chai, T.T.; Wang, K.; Zhu, L.Z.; Huang, Y.; Shen, G.M.; Li, Y.R.; Li, X.F.; Wang, C.J. The developmental effect of difenoconazole on zebrafish embryos: A mechanism research. Environ. Pollut. 2016, 212, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Feki, A.; Ben Saad, H.; Jaballi, I.; Magne, C.; Boudawara, O.; Zeghal, K.M.; Hakim, A.; Ben Ali, Y.; Ben Amara, I. Methyl thiophanate-induced toxicity in liver and kidney of adult rats: A biohemical, molecular and histopathological approach. Cell. Mol. Biol. 2017, 63, 20–28. [Google Scholar] [CrossRef]
- Salam, S.; Iqbal, Z.; Khan, A.A.; Mahmood, R. Oral administration of thiram inhibits brush border membrane enzymes, oxidizes proteins and thiols, impairs redox system and causes histological changes in rat intestine: A dose dependent study. Pestic. Biochem. Physiol. 2021, 178, 104915. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Standard Name | Chemical Name | Molecular Weight (g/mol) | Commercial Source |
|---|---|---|---|
| p-Aminobenzoic acid | p-Aminobenzoic acid | 137.14 | Macklin (Shanghai, China) |
| Carbendazim | Methyl benzimidazole-2-ylcarbamate | 191.19 | Shyuanye (Shanghai, China) |
| Difenoconazole | 1-[2-[2-Chloro-4-(4-chloro-phenoxy)-phenyl]-4-methyl[1,3]dioxolan-2-ylmethyl]-1H-1,2,4-triazole | 406.26 | Aladdin (Shanghai, China) |
| Dipicolinic acid | 2,6-Pyridinedicarboxylic acid | 167.12 | Macklin |
| Flusilazole | Di(4-fluorophenyl)(1,2,4-triazole-2-ylmethyl)methylsilane | 315.39 | Macklin |
| Gentamicin | 2-[4,6-Diamino-3-[5-amino-2-[1-(methylamino)ethyl]oxan-4-yl]oxy-2-hydroxycyclohexyl]oxyoxan-3-ol | 477.60 | Aladdin |
| Kojic acid | 5-Hydroxy-2-hydroxymethylgamma-pyrone | 142.11 | Macklin |
| Prochloraz | N-Propyl-N-[2-(2,4,6-trichlorophenoxy)ethyl]imidazole-1-carboxamid | 376.67 | Aladdin |
| Quinolinic acid | 2,3-Pyridinedicarboxylic acid | 167.12 | Macklin |
| Thiophanate methyl | Dimethyl(1,2-phenylene)bis(iminocarbonothioyl)bis(carbamate) | 342.39 | Aladdin |
| Thiram | Tetramethylthiuram disulfide | 240.43 | Macklin |
| Fungicide | HPLC Method | Retention Time (min) | Absorption Wavelength (nm) |
|---|---|---|---|
| p-Aminobenzoic acid | 1 | 5.6 | 283 |
| Carbendazim | 2 | 20.1 | 254 |
| Difenoconazole | 2 | 20.3 | 254 |
| Dipicolinic acid | 3 | 6.7 | 270 |
| Flusilazole | 2 | 17.3 | 254 |
| Gentamicin | 4 | 35.7 | 350 5 |
| Kojic acid | 3 | 6.1 | 280 |
| Prochloraz | 2 | 19.2 | 254 |
| Quinolinic acid | 3 | 7.5 | 270 |
| Thiophanate methyl | 2 | 9.2 | 254 |
| Thiram | 2 | 12.1 | 254 |
| Fungicide | Diffusion Ability 1 | Volume 2 (cm3/mol) | Dipole 2 (debye) |
|---|---|---|---|
| p-Aminobenzoic acid | 0.205 ± 0.055 a | 91.532 | 2.0528 |
| Carbendazim | 0.138 ± 0.036 ab | 135.058 | 4.3805 |
| Difenoconazole | 0.055 ± 0.032 cd | 285.34 | 6.0014 |
| Dipicolinic acid | 0.029 ± 0.017 cd | 111.23 | 7.7607 |
| Flusilazole | 0 e | 214.571 | 3.6535 |
| Gentamicin | 0 e | 339.397 | 3.1864 |
| Kojic acid | 0.023 ± 0.006 d | 93.169 | 7.233 |
| Prochloraz | 0.057 ± 0.014 c | 240.481 | 6.6457 |
| Quinolinic acid | 0.177 ± 0.020 a | 113.031 | 2.5696 |
| Thiophanate methyl | 0.047 ± 0.019 cd | 248.402 | 6.7427 |
| Thiram | 0.094 ± 0.049 bc | 153.518 | 0.0088 |
| Fungicide | Mycelial Growth 2,3 (mm) | EC50 (mM) | |
|---|---|---|---|
| 1 mM Fungicide | 5 mM Fungicide | ||
| p-Aminobenzoic acid | 32.2 ± 1.6 a | 25.3 ± 1.4 c | 11.4 ± 0.3 c |
| Carbendazim | 34.0 ± 0.7 a | 30.1 ± 0.9 b | 20.1 ± 0.4 b |
| Difenoconazole | 15.3 ± 1.0 c | 9.0 ± 0.9 e | 0.79 ± 0.11 f |
| Dipicolinic acid | 29.8 ± 0.7 | 18.2 ± 1.3 d | 4.5 ± 0.2 e |
| Flusilazole | 6.9 ± 0.2 e | - | 0.51 ± 0.06 f |
| Gentamicin | 34.7 ± 1.9 a | 19.3 ± 0.5 d | 5.3 ± 0.2 d |
| Kojic acid | 30.0 ± 1.0 b | 29.7 ± 0.5 b | 20.3 ± 0.5 b |
| Prochloraz | 9.7 ± 0.5 d | - | 0.13 ± 0.01 g |
| Quinolinic acid | 32.0 ± 1.4 ab | 33.6 ± 0.5 a | 46.9 ± 0.3 a |
| Thiophanate methyl | 33.0 ± 0.9 a | 23.7 ± 1.4 c | 11.1 ± 0.7 c |
| Thiram | 8.8 ± 1.1 d | - | 0.52 ± 0.06 f |
| Control 1 | 33.3 ± 0.5 a | 33.3 ± 0.5 a | - |
| Fungicide | Lesion Length 2 (mm) | Efficacy (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| 3 Days after Inoculation | 4 Days after Inoculation | 5 Days after Inoculation | 6 Days after Inoculation | 3 Days after Inoculation | 4 Days after Inoculation | 5 Days after Inoculation | 6 Days after Inoculation | |
| p-Aminobenzoic acid | 1.7 ± 0.6 c | 4.0 ± 0.7 b | 5.9 ± 1.8 c | 9.7 ± 2.0 c | 43 | 44 | 50 | 46 |
| Carbendazim | 2.2 ± 0.3 bc | 4.0 ± 0.6 b | 6.9 ± 1.5 bc | 11.1 ± 1.5 bc | 28 | 45 | 42 | 38 |
| Difenoconazole | 2.8 ± 0.8 abc | 6.9 ± 0.7 a | 11.6 ± 1.3 a | 18.4 ± 1.5 a | 8 | 4 | 2 | −3 |
| Dipicolinic acid | 2.8 ± 0.1 a | 6.2 ± 0.6 a | 8.4 ± 1.8 abc | 13.6 ± 1.8 b | 7 | 14 | 29 | 24 |
| Flusilazole | 3.2 ± 0.5 a | 6.6 ± 1.1 a | 10.8 ± 1.5 a | 15.4 ± 2.7 ab | −6 | 9 | 8 | 14 |
| Gentamicin | 2.7 ± 1.0 abc | 7.2 ± 1.6 a | 11.0 ± 1.7 a | 16.0 ± 2.6 ab | 11 | 13 | 7 | 11 |
| Kojic acid | 3.0 ± 0.6 ab | 7.1 ± 1.0 a | 11.7 ± 1.0 a | 17.9 ± 0.9 a | 0 | 2 | 1 | 0 |
| Prochloraz | 2.5 ± 0.8 abc | 6.2 ± 0.6 a | 9.9 ± 1.6 ab | 17.6 ± 2.4 a | 17 | 14 | 16 | 2 |
| Quinolinic acid | 2.2 ± 0.7 abc | 5.0 ± 1.1 ab | 8.0 ± 1.8 abc | 12.3 ± 1.7 bc | 26 | 31 | 32 | 31 |
| Thiophanate methyl | 2.7 ± 0.4 abc | 7.8 ± 1.8 a | 12.6 ± 2.2 a | 18.6 ± 2.4 a | 11 | −13 | −7 | −4 |
| Thiram | 2.6 ± 0.6 abc | 6.6 ± 1.7 a | 11.1 ± 2.5 a | 17.2 ± 3.1 a | 13 | 9 | 6 | 4 |
| Control 1 | 3.0 ± 0.4 a | 7.2 ± 1.9 a | 11.8 ± 2.6 a | 17.9 ± 2.4 a | - | - | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, G.-Y.; Chen, Y.; Wang, S.-Y.; Shi, X.-C.; Herrera-Balandrano, D.D.; Polo, V.; Laborda, P. Peel Diffusion and Antifungal Efficacy of Different Fungicides in Pear Fruit: Structure-Diffusion-Activity Relationships. J. Fungi 2022, 8, 547. https://doi.org/10.3390/jof8050547
Zhu G-Y, Chen Y, Wang S-Y, Shi X-C, Herrera-Balandrano DD, Polo V, Laborda P. Peel Diffusion and Antifungal Efficacy of Different Fungicides in Pear Fruit: Structure-Diffusion-Activity Relationships. Journal of Fungi. 2022; 8(5):547. https://doi.org/10.3390/jof8050547
Chicago/Turabian StyleZhu, Gui-Yang, Ying Chen, Su-Yan Wang, Xin-Chi Shi, Daniela D. Herrera-Balandrano, Victor Polo, and Pedro Laborda. 2022. "Peel Diffusion and Antifungal Efficacy of Different Fungicides in Pear Fruit: Structure-Diffusion-Activity Relationships" Journal of Fungi 8, no. 5: 547. https://doi.org/10.3390/jof8050547
APA StyleZhu, G.-Y., Chen, Y., Wang, S.-Y., Shi, X.-C., Herrera-Balandrano, D. D., Polo, V., & Laborda, P. (2022). Peel Diffusion and Antifungal Efficacy of Different Fungicides in Pear Fruit: Structure-Diffusion-Activity Relationships. Journal of Fungi, 8(5), 547. https://doi.org/10.3390/jof8050547