
Insight into Biological Control Potential of Hirsutella citriformis against Asian Citrus Psyllid as a Vector of Citrus Huanglongbing Disease in America


Abstract
:1. Introduction
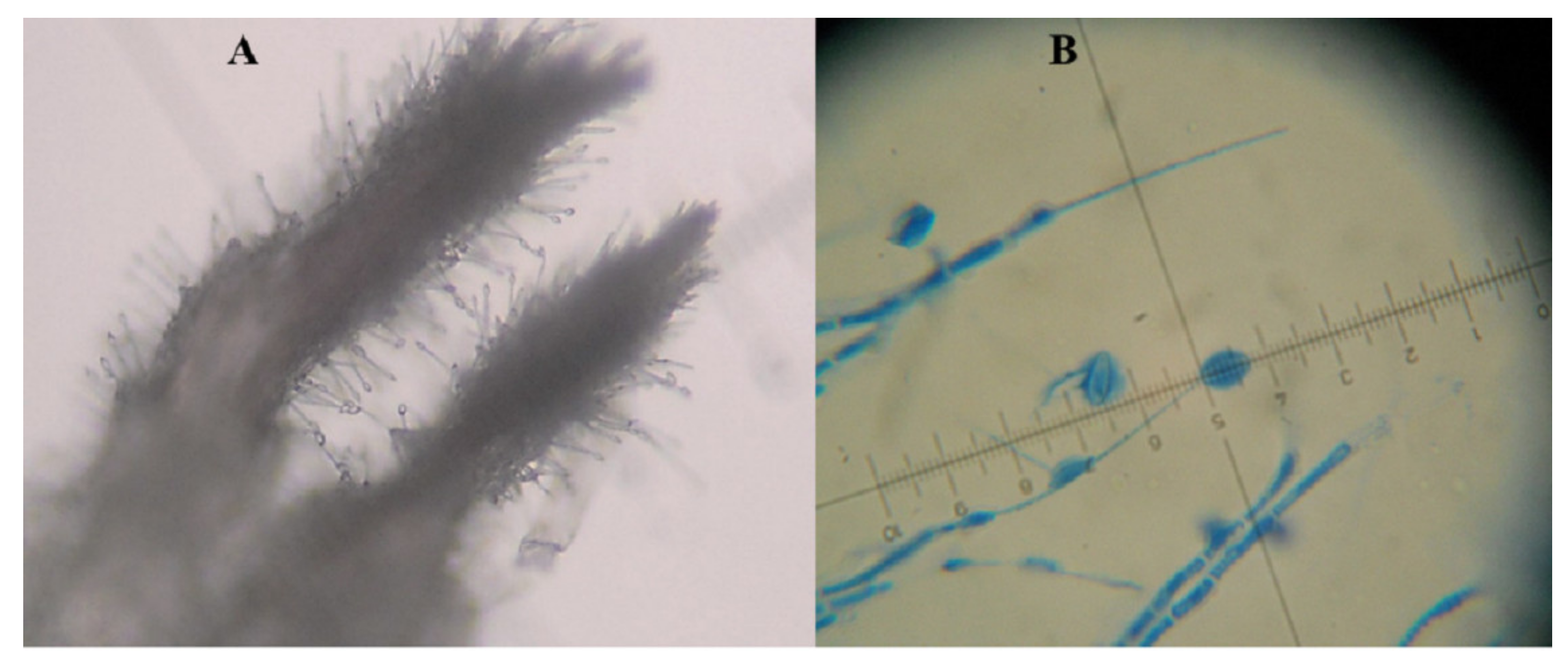
2. Hirsutella citriformis Identification
3. Hirsutella citriformis Origin and Distribution
4. Insects Associated with H. citriformis
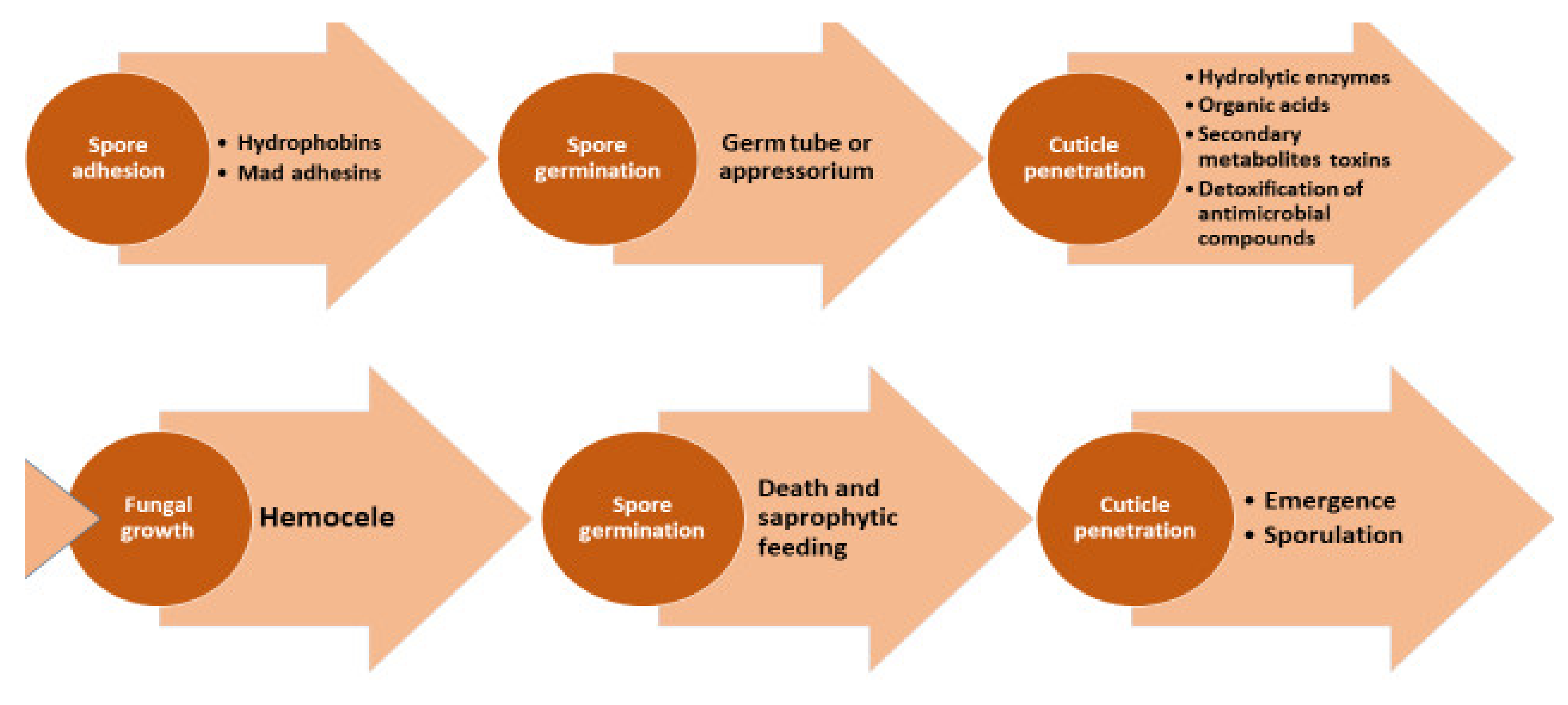
5. H. citriformis Mode of Action
6. Adverse Effects of Environmental Factors on H. citriformis
6.1. Temperature
6.2. Relative Humidity
6.3. Solar Radiation
7. Metabolites Production
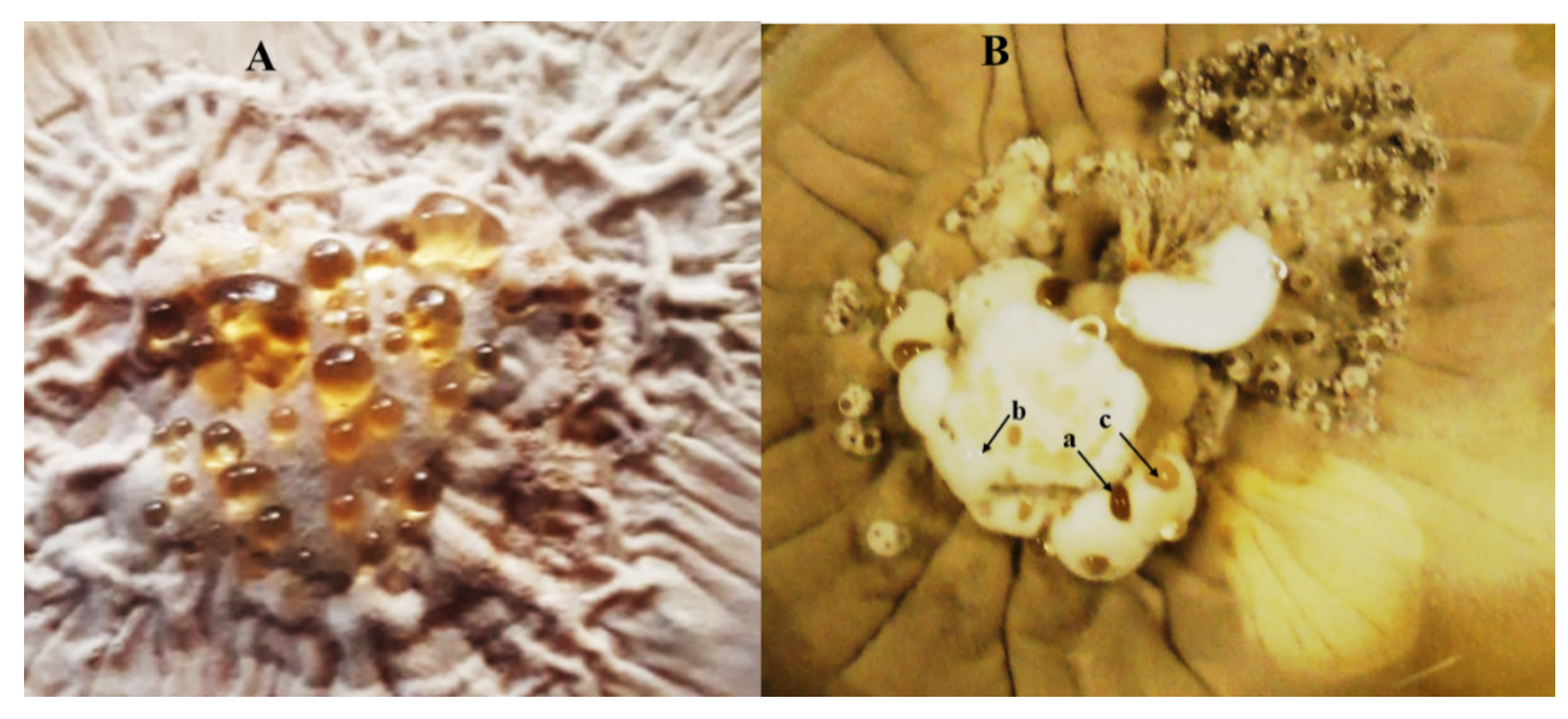
8. Hirsutella citriformis Epizootics on Diaphorina citri
9. Assessments of H. citriformis Biocontrol against D. citri and Bactericera cockerelli
10. Effects on Predators and Other Non-Target Arthropods
11. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ekesi, S.; Maniania, N.K. Use of Entomopathogenic Fungi in Biological Pest Management; Ekesi, S., Maniania, N.K., Eds.; Research Signpost Editorial: Trivandrum, India, 2007; pp. 57–90. Available online: https://www.researchgate.net/publication/297758806 (accessed on 15 April 2022).
- Al-Ani, L.K.T. Entomopathogenic fungi in IP landscape. In Intellectual Property Issues in Microbiology; Singh, H., Keswani, C., Singh, S., Eds.; Springer: Singapore, 2019. [Google Scholar] [CrossRef]
- Moorhouse, E.R.; Gillespie, A.T.; Sellers, E.K.; Charnley, A.K. Influence of fungicides and insecticides on the entomogenous fungus Metarhizium anisopliae a pathogen of the vine weevil, Otiorhynchus sulcatus. Biocontrol Sci. Technol. 1992, 2, 49–58. [Google Scholar] [CrossRef]
- De Faria, M.R.; Wraight, S.P. Mycoinsecticides and Mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control 2007, 43, 237–256. [Google Scholar] [CrossRef]
- Maina, U.M.; Galadima, I.B.; Gamba, F.M.; Zakaria, D. A review on the use of entomopathogenic fungus in the management of insect pest of field crops. J. Entomol. Zool. Stud. 2018, 6, 27–32. [Google Scholar]
- Subandiyah, S.; Nikon, N.; Sato, H.; Wagiman, F.; Tsuyumu, S.; Fakatsu, T. Isolation and characterization of two entomopathogenic fungi attacking Diaphorina citri (Homoptera, Psilloidea) in Indonesia. Mycoscience 2000, 41, 509–513. [Google Scholar] [CrossRef]
- Meyer, J.M.; Hoy, M.A.; Boucias, D.G. Morphological and molecular characterization of a Hirsutella species infecting the Asian citrus psyllid, Diaphorina citri Kuwayama (Hemiptera: Psyllidae), in Florida. J. Invertebr. Pathol. 2007, 95, 101–109. [Google Scholar] [CrossRef]
- Hall, D.G.; Hentz, M.G.; Meyer, J.M.; Kriss, A.B.; Gottwald, T.R.; Boucias, D.G. Observations on the entomopathogenic fungus Hirsutella citriformis attacking adult Diaphorina citri (Hemiptera: Psyllidae) in a managed citrus grove. BioControl 2012, 57, 663–675. [Google Scholar] [CrossRef]
- Petch, T. The genus Hirsutella Petch. Trans. Br. Mycol. Soc. 1923, 9, 93–94. [Google Scholar] [CrossRef]
- Mains, E.B. Entomogenous species of Hirsutella, Tilachlidium and Synnematium. Mycologia 1951, 43, 691–718. [Google Scholar] [CrossRef]
- Speare, A.T. On certain entomogenous fungi. Slycologia 1920, 12, 62–76. [Google Scholar] [CrossRef]
- Casique-Valdes, R.; Reyes-Martinez, A.Y.; Sanchez-Peña, S.R.; Bidochka, M.J.; Lopez-Arroyo, J.I. Pathogenicity of Hirsutella citriformis (Ascomycota: Cordycipitaceae) to Diaphorina citri (Hemiptera: Psyllidae) and Bactericera cockerelli (Hemiptera: Triozidae). Fla. Entomol. 2011, 94, 703–705. [Google Scholar] [CrossRef]
- Toledo, A.V.; Simurro, M.E.; Balatti, P.A. Morphological and molecular characterization of a fungus, Hirsutella sp., isolated from planthoppers and psocids in Argentina. J. Insect Sci. 2013, 13, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Gonzalez, O.; Rodriguez-Guerra, R.; Lopez-Arroyo, J.I.; Sandoval-Coronado, C.F.; Maldonado-Blanco, M.G. Radial growth, sporulation, and virulence of Mexican isolates of Hirsutella citriformis against Diaphorina citri. Southwest Entomol. 2015, 40, 111–120. [Google Scholar] [CrossRef]
- Tanada, Y.; Kaya, H.K. Fungal Infections. In Insect Pathology; Tanada, Y., Kaya, H.K., Eds.; Academic Press Inc.: Cambridge, MA, USA, 1993; pp. 318–387. [Google Scholar]
- Hywel-Jones, N.L. Hirsutella species associated with hoppers (Homoptera) in Thailand. Mycol. Res. 1997, 101, 1202–1206. [Google Scholar] [CrossRef] [Green Version]
- Álvarez, J.F.; Montes de Oca, F.N.; Grillo-Ravelo, H. Hongos entomopatógenos de Diaphorina citri Kirk. (Homoptera: Psyllidae) en Jovellanos, Matanzas. Centro Agrícola 2003, 30, 87. (In Spanish) [Google Scholar]
- Perez-Gonzalez, O.; Rodriguez-Villarreal, R.A.; Lopez-Arroyo, J.I.; Maldonado-Blanco, M.G.; Rodríguez-Guerra, R. Mexican strains of Hirsutella isolated from Diaphorina citri (Hemiptera: Liviidae): Morphologic and molecular characterization. Fla. Entomol. 2015, 98, 290–297. [Google Scholar] [CrossRef]
- Kondo, T.; González, G.; Tauber, C.; Guzmán-Sarmiento, Y.C.; Vinasco-Mondragon, A.F.; Forero, D. A checklist of natural enemies of Diaphorina citri Kuwayama (Hemiptera: Liviidae) in the department of Valle del Cauca, Colombia, and the world. Insecta Mundi 2015, 0457, 1–14. Available online: http://centerforsystematicentomology.org/ (accessed on 20 May 2022).
- Reddy, N.; Mahesh, G.; Priya, M.; Singh, R.S.; Manjunatha, L. Hirsutella. In Beneficial Microbes in Agro-Ecology; Academic Press: Cambridge, MA, USA; pp. 817–831. [CrossRef]
- Étienne, J.; Quilici, S.; Marival, D.; Franck, A. Biological control of Diaphorina citri (Hemiptera: Psyllidae) in Guadeloupe by imported Tamarixia radiata (Hymenoptera: Eulophidae). Fruits 2001, 56, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Sajap, A.S. Prevalence of an entomopathogenic fungus, Hirsutella citriformis on Leucaena Psyllid, Heterapsylla cubana, in Malaysia. Pertanika J. Trop. Agric. Sci. 1993, 16, 95–99. [Google Scholar]
- Aubert, B. Trioza erytreae Del Guercio and Diaphorina citri Kuwayama (Homoptera: Psylloidea), the two vectors of citrus greening disease: Biological aspects and possible control strategies. Fruits 1987, 42, 149–162. [Google Scholar]
- Reyes-Rosas, M.A.; Loera-Gallardo, J.; López-Arroyo, J.I. Comparison of the chemical and natural control of Asian citrus psyllid Diaphorina citri. Rev. Mex. Cienc. Agric. 2013, 4, 95–501. Available online: http://www.redalyc.org/articulo.oa?id=263127562001 (accessed on 20 May 2022).
- Henríquez-Liria, D.J.; Navarro-Morales, S.; Bastardo, R.H.; Reyes-Valentín, M. Aspectos ecológicos de Diaphorina citri Kuwayama, 1908, en el cultivo de cítricos en Villa Altagracia y El Seibo, República Dominicana. Novit. Caribaea 2016, 10, 52–62. [Google Scholar] [CrossRef] [Green Version]
- Godoy-Ceja, C.A.; Cortez-Madrigal, H. Potential of Aclepias curassavica L. (Apocynaceae) in the biological control of pests. Rev. Mex. Cienc. Agric. 2018, 9, 303–315. [Google Scholar] [CrossRef] [Green Version]
- Ayala-Zermeño, M.A.; Gallou, A.; Berlanga-Padilla, A.M.; Serna-Domínguez, M.G.; Arredondo-Bernal, H.C.; Montesinos-Matías, R. Characterisation of entomopathogenic fungi used in the biological control programme of Diaphorina citri in Mexico. Biocontrol Sci. Technol. 2015, 25, 1192–1207. [Google Scholar] [CrossRef]
- Suaste-Dzul, A.; Gallou, A.; Félix-Portillo, M.; Moreno-Carrillo, G.; Sánchez-González, J.; Palomares-Pérez, M.; Arredondo-Bernal, H. Seasonal incidence of ‘Candidatus Liberibacter asiaticus’ (Rhizobiales: Rhizobiaceae) in Diaphorina citri (Hemiptera: Liviidae) in Colima, Mexico. Trop. Plant Pathol. 2017, 42, 410–415. [Google Scholar] [CrossRef]
- Boucias, D.G.; Scharf, D.W.; Breaux, S.E.; Purcell, D.H.; Mizell, R.E. Studies on the fungi associated with the glassy-winged sharpshooter Homalodisca coagulata with emphasis on a new species Hirsutella homalodiscae nom. prov. BioControl 2007, 52, 231–258. [Google Scholar] [CrossRef]
- Pérez-González, O.; Gomez-Flores, R.; Tamez-Guerra, P. Mycelial compatibility, anastomosis, and nuclear characterization of Mexican Hirsutella citriformis strains isolated from Diaphorina citri. PeerJ 2021, 9, e11080. [Google Scholar] [CrossRef]
- Qu, J.; Zou, X.; Yu, J.; Zhou, Y. The conidial mucilage, natural film coatings, is involved in environmental adaptability and pathogenicity of Hirsutella satumaensis Aoki. Sci. Rep. 2017, 7, 1301. [Google Scholar] [CrossRef]
- Jansson, H.B.; Friman, E. Infection-related surface proteins on conidia of the nematophagous fungus Drechmeria coniospora. Mycol. Res. 1999, 103, 249–256. [Google Scholar] [CrossRef]
- Vidal, C.; Fargues, J. Climatic constraints for fungal biopesticides. In Use of Entomopathogenic Fungi in Biological Pest Management; Ekesi, S., Maniania, N.K., Eds.; Research Signpost Editorial: Trivandrum, India, 2007; pp. 39–55. [Google Scholar]
- Orduño-Cruz, N.; Guzmán-Franco, A.W.; Rodríguez-Leyva, E.; Alatorre-Rosas, R.; González-Hernández, H.; Mora-Aguilera, G.; Rodríguez-Maciel, J.C. In vitro selection of a fungal pathogen for use against Diaphorina citri. Biol. Control 2015, 90, 6–15. [Google Scholar] [CrossRef]
- Hall, D.G.; Richardson, M.L.; Ammar, E.D.; Halbert, S.E. Asian citrus psyllid, Diaphorina citri, vector of citrus huanglongbing disease. Entomol. Exp. Appl. 2013, 146, 207–223. [Google Scholar] [CrossRef]
- Gardner, W.A.; Sutton, R.M.; Noblet, R. Persistence of Beauveria bassiana, Nomuraea rileyi, and Nosema necatrix on soybean foliage. Environ. Entomol. 1977, 6, 616–618. [Google Scholar] [CrossRef]
- Liu, W.Z.; Boucias, D.G.; McCoy, C.W. Extraction and characterization of the insecticidal toxin Hirsutellin A produced by Hirsutella thompsonii var. thompsonii. Exp. Mycol. 1995, 19, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Boucias, D.G.; Farmerie, W.G.; Pendland, J.C. Cloning and sequencing of cDNA of the insecticidal toxin Hirsutellin A. J. Invertebr. Pathol. 1998, 72, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Quesada-Sojo, K.A.; Rivera-Méndez, W. Hirsutella, agente biocontrolador de ácaros e insectos de importancia agronómica. Revista Tecnología en Marcha 2016, 29, 85–93. [Google Scholar] [CrossRef]
- Rosas-Acevedo, J.L.; Boucias, D.G.; Lezama, R.; Sims, K.; Pescador, A. Exudate from sporulating cultures of Hirsutella thompsonii Fisher inhibit oviposition by the two-spotted spider mite Tetranychus urticae Koch. Exp. Appl. Acarol. 2003, 29, 213–222. [Google Scholar] [CrossRef]
- Wang, B.; Wu, W.; Liu, X. Purification and characterization of a neutral serine protease with nematicidal activity from Hirsutella rhossiliensis. Mycopathologia 2007, 163, 169–176. [Google Scholar] [CrossRef]
- Mazet, I.; Vey, A. Hirsutellin A, a toxic protein produced in vitro by Hirsutella thompsonii. Microbiology 1995, 141, 1343–1348. [Google Scholar] [CrossRef] [Green Version]
- Herrero-Galan, E.; Lacadena, J.; Martinez-del-Pozo, A.; Boucias, D.G.; Olmo, N.; Oñaderra, M.; Gavilanes, J. The insecticidal protein Hirsutellin A from the mite fungal pathogen Hirsutella thompsonii is a ribotoxin. Proteins 2008, 72, 217–228. [Google Scholar] [CrossRef]
- Maimala, S.; Tartar, A.; Boucias, D.; Chandrapaty, A. Detection of the toxin Hirsutellin A from Hirsutella thompsonii. J. Invertebr. Pathol. 2002, 80, 112–126. [Google Scholar] [CrossRef]
- Madla, S.; Isaka, M.; Wongsa, P. Modification of culture conditions for production of the anti-tubercular hirsutellones by the insect pathogenic fungus Hirsutella nivea BCC2594. Lett. Appl. Microbiol. 2008, 47, 74–78. [Google Scholar] [CrossRef]
- Ramachandran, A.; Thangappan, B.S.; Ponmurugan Ponnusamy, P. Evaluation of secondary metabolites of Hirsutella citriformis against Udaspes folus infecting Curcuma longa L. J. Pharm. Res. 2013, 7, 7–14. [Google Scholar] [CrossRef]
- Meng, L.; Sun, S.; Shen, Z.; Wang, P.; Jiang, X. Antioxidant activity of polysaccharides produced by Hirsutella sp. and relation with their chemical characteristics. Carbohydr. Polym. 2015, 117, 452–457. [Google Scholar] [CrossRef] [PubMed]
- Leung, P.; Zhao, P.; Wu, J.Y. Chemical properties and antioxidant activity of exopolysaccharides from mycelial culture of Cordyceps sinensis fungus Cs-HK1. Food Chem. 2009, 114, 1251–1256. [Google Scholar] [CrossRef]
- Li, R.; Jiang, X.; Guan, H. Optimization of mycelium biomass and exopolysaccharides production by Hirsutella sp. In submerged fermentation and evaluation of exopolysaccharides antibacterial activity. Afr. J. Biotechnol. 2010, 9, 195–202. [Google Scholar] [CrossRef]
- Samson, R.A.; McCoy, C.W.; O’Donnell, K.L. Taxonomy of the acarine parasite Hirsutella thompsonii. Mycologia 1980, 72, 359–377. [Google Scholar] [CrossRef]
- Loughheed, T.C.; MacLeod, D.M. Extracellular metabolic products of a Hirsutella species. Nature 1958, 182, 114–115. [Google Scholar] [CrossRef]
- Blango, M.G.; Kniemeyer, O.; Brakhage, A.A. Conidial surface proteins at the interface of fungal infections. PLoS Pathog. 2019, 15, e1007939. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Palma-Ramos, A.; Moreno-Aparicio, A.M.; Castrillón-Rivera, L.E.; Castañeda-Sánchez, J.I.; Muñoz-Duarte, A.R.; Mendoza-Pérez, F.; Padilla-Desgarennes, C. Detecting enzymatic activity of β-glucuronidase polymorphonuclear neutrophil in human grains Nocardia brasiliensis actinomycetoma, in vitro. Dermatol. Rev. Mex. 2016, 60, 488–498. [Google Scholar]
- Pérez-Gonzalez, O.; Rodriguez-Guerra, R.; Lopez-Arroyo, J.I.; Sandoval-Coronado, C.F.; Maldonado-Blanco, M.G. Effect of Mexican Hirsutella citriformis (Hypocreales: Ophiocordycipitaceae) strains on Diaphorina citri (Hemiptera: Liviidae) and the predators Chrysoperla rufilabris (Neuroptera: Chrysopidae) and Hippodamia convergens (Coleoptera: Coccinellidae). Fla. Entomol. 2016, 99, 509–515. [Google Scholar] [CrossRef] [Green Version]
- Ibarra-Cortés, K.H.; González-Hernández, H.; Guzmán-Franco, A.W. Susceptibility of nymphs and adults of Diaphorina citri to the entomophathogenic fungus Hirsutella citriformis. Biocontrol Sci. Technol. 2017, 27, 433–438. [Google Scholar] [CrossRef]
- Halbert, S.E.; Manjunath, K.L. Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: A literature review and assessment of risk in Florida. Fla. Entomol. 2004, 87, 330–353. [Google Scholar] [CrossRef]
- Pérez-González, O.; Sandoval-Coronado, C.F.; Maldonado-Blanco, M.G. Evaluation of Mexican strains of Hirsutella citriformis against Diaphorina citri in a semifield bioassay. Southwest Entomol. 2016, 41, 361–372. [Google Scholar] [CrossRef]
- Pérez-González, O.; Arredondo-Bernal, H.C.; Montesinos-Matías, R.; Mellín-Rosas, M.A.; Maldonado-Blanco, M.G. Effect of native strains of Hirsutella citriformis1 on Diaphorina citri2 adults under field conditions. Southwest Entomol. 2020, 45, 435–446. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Source 1 | Phialide Length | Conidium | Mucus Diameter | |||
|---|---|---|---|---|---|---|---|
| Total | Base | Sterigma | Length | Diameter | |||
| [11] | H | 5.5–8.5 | 1.5–1.8 | ||||
| [10] | H | 36.0–54.0 | 6.0–14.0 | 30.0–40.0 | 5.0–8.0 | 2.0–2.5 | |
| [16] | H | 18.5–52.0 | 3.5–5.0 | 1.0–1.5 | |||
| [6] | CM | 27.5–62.3 | 5.1–9.4 | 22.4–52.9 | 6.4–7.6 | 2.1–2.8 | |
| [17] | H | 16.8–23.6 | 6.8–9.1 | 1.5–2.3 | |||
| [7] | H | 17.5 ± 1.9 | 5.9 ± 0.8 | 2.6 ± 0.3 | |||
| [12] | H | 6.8–7.0 | 1.5–2.0 | ||||
| [13] | H | 35.6–55.4 | 28.7–47.5 | 5.9–7.9 | 2.0–3.0 | ||
| [13] | H | 22.4–34.7 | 16.8–28.0 | 5.6–7.8 | 2.2–2.8 | ||
| [14] | CM | 26.0–42.0 | 4.0–8.5 | 20.0–38.0 | 5.4–6.3 | 1.6–2.0 | |
| Country | Region | Reference |
|---|---|---|
| Cuba | Jovellanos, Matanzas | [17] |
| Carmelina in Cienfuegos; Morón and Ceballos in Ciego de Ávila; San Antonio de los Baños in La Habana | Cabrera et al., 2004, cited by [20] | |
| Jaguey El Grande, Matanzas | [19] | |
| Argentina | Los Hornos and La Plata, Buenos Aires | [13] |
| United States | Hawaii | [11] |
| Polk, Marion, and Hendry, Florida | [7] | |
| Indian River, Florida | [8] |
| Parasitized Insect (Family) | Location | Reference |
|---|---|---|
| Fulgoridea family Latreille | New Zeeland | [11] |
| Ricania discalis Germar (Ricaniidae) | New Zeeland | |
| Perkinsiella sacharicida Kirkaldy (Hemíptera: Cicadellidae) | Hawaii | |
| Siphanta acuta Walker (Hemiptera: Flatidae) | Hawaii | |
| Pentatomidae family Leach | India | [9] |
| Bothriocera venosa Fowler (Cixiidae) | Puerto Rico | Gregory & Martorell 1940, cited by [10] |
| Leptopharsa constricta Champion (Tingidae) | United States | [10] |
| Corythuca ulmi Osborn & Drake (Tingidae) | United States | [10] |
| Heteropsylla cubana Crawford (Psyllidae) | Malaysia | [22] |
| Oliarus dimidiatus Berg (Cixiidae) | Argentina | [23] |
| [13] | ||
| Diaphorina citri Kuwayama (Liviidae) | Rénion island, France | [17] |
| Cuba | [6] | |
| [19] | ||
| Indonesia | [21] | |
| Isla Guadalupe | [7] | |
| Florida | [8] | |
| Veracruz, MX | [24] | |
| Tamaulipas, MX | [12] | |
| Tamaulipas, MX | [25] | |
| Veracruz & Puebla, MX | [26] | |
| Veracrux, MX | [27] | |
| Campeche, MX | [14] | |
| Colima, MX | ||
| Chiapas, MX | ||
| Quintana Roo, MX | ||
| San Luis Potosí, MX | ||
| Tabasco, MX | ||
| Veracruz, MX | ||
| Yucatán, MX | ||
| Hidalgo, MX | Pérez-González et al., (unpublished data) | |
| Oaxaca, MX |
| State | Municipality | Reference |
|---|---|---|
| Campeche | Edzna | [18] |
| Chiapas | Tapachula | [18] |
| Colima | Tecomán | [18] |
| Hidalgo | Santa Ana | Pérez-González et al., (unpublished data) |
| Oaxaca | Melchor Ocampo | Pérez-González et al., (unpublished data) |
| Puebla | La Legua and La Garita | [26] |
| Quintana Roo | Nuevo Israel | [18] |
| San Luis Potosí | Xolol | [18] |
| Tabasco | Huimanguillo | [18] |
| Tamaulipas | Rio Bravo | [24] |
| Gómez Farías and Llera | [12] | |
| Veracruz | Tlapacoyan | [18] |
| Troncones, Ixtacuaco and El Lindero | [26] | |
| Tuxpan | [25] | |
| Yucatán | Mococha | [18] |
| Hirsutella Species | Toxin | Type of Toxin | Toxin Size/Yield | Activity | Reference |
|---|---|---|---|---|---|
| H. thompsonii strain JAB-04 | Hirsutellin A | Dot-blot anti HtA | 16.3 kDa/0.12 mg/mL | Insecticidal vs. Galleria mellonella | [37,38] |
| thompsonii | Hirsutellin A Hirsutellin B | Ribotoxin | 130 amino-acids | Insecticidal vs. Galleria mellonella Insecticidal vs. Drosophila melanogaster | [39] |
| H. thompsonii strain HtM120I | Hirsutellin A | Dot-blot anti HtA | 16 kDa (200–400 ng/mL) | Insecticidal vs. Tetranychus urticae | [40] |
| H. thompsonii | Culture exudates | Phomalactones? | Undetermined | Reduces eggs production in mites | [40] |
| H. rhossilensis | Hnsp | SAAPF-pNA protease activity & others | 30 kDa | Nematicide | [41] |
| H. thompsonii | Hirsutellin A | W Blot NH2 terminal | 16 kDa | Insecticidal vs. Aedes aegypti, Galleria mellonella | [42] |
| H. thompsonii | Hirsutellin A | Ribotoxin | 130 aa | Unidentified | [43] |
| H. thompsonii | Hirsutellin A | Dot-blot anti HtA | 481–936 ng/mL | Insecticidal vs. Galleria mellonella | [44] |
| H. nivea strain BCC2594 | Hirsutellones A, B, C and D | Alkaloids | 29.9, 1696, 22.6 and 15.7 mg/L | Antibacterial against Mycobacterium tuberculosis | [45] |
| Unidentified sp. | Exo-polysaccharide (EPS) | Mannose (man), galactose (gal), glucose (glc) | 23 kDa | Gram (+) bacteria, Bacillus subtilis, Micrococcus tetragenus | [46] |
| H. beakdumountain | EPS | EPS 1 y 2: gal, glc, man EPS3: man, glc | EPS1 = 43 kDa EPS2 = 19.5 kDa EPS3 = 4.7 kDa | Potential to eliminate hydroxyl radicals | [47] |
| Intracellular polysaccharide | IPS 1 y 2: gal, glc, man IPS 3: man, glc | IPS1 = 23.1 kDa IPS2 = 21.5 kDa IPS3 = 10.4 kDa | Potential to eliminate hydroxyl radicals | [47] | |
| H. sinensis | EPS | Polysaccharides (65–70%) + protein (25%) | 5 kDa–200 kDa | Antioxidant activity | [48] |
| H. satumaensis Aoki | Mucilage from aerial conidia | Protein content, oligo-saccharides + man | 0.12 mg/mL | Insecticidal vs. Galleria mellonella Plutella xylostella | [30] |
| H. citriformis | Culture supernatant | Benzene and phthalic acid | Unidentified | Insecticidal activity | [49] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-González, O.; Gomez-Flores, R.; Tamez-Guerra, P. Insight into Biological Control Potential of Hirsutella citriformis against Asian Citrus Psyllid as a Vector of Citrus Huanglongbing Disease in America. J. Fungi 2022, 8, 573. https://doi.org/10.3390/jof8060573
Pérez-González O, Gomez-Flores R, Tamez-Guerra P. Insight into Biological Control Potential of Hirsutella citriformis against Asian Citrus Psyllid as a Vector of Citrus Huanglongbing Disease in America. Journal of Fungi. 2022; 8(6):573. https://doi.org/10.3390/jof8060573
Chicago/Turabian StylePérez-González, Orquídea, Ricardo Gomez-Flores, and Patricia Tamez-Guerra. 2022. "Insight into Biological Control Potential of Hirsutella citriformis against Asian Citrus Psyllid as a Vector of Citrus Huanglongbing Disease in America" Journal of Fungi 8, no. 6: 573. https://doi.org/10.3390/jof8060573