
Isolation and Characterization of a Novel Hydrophobin, Sa-HFB1, with Antifungal Activity from an Alkaliphilic Fungus, Sodiomyces alkalinus

 , , , , and
, , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Strains of Sodiomyces alkalinus
2.2. Cultivation of the Sodiomyces alkalinus and the Extraction of the Hydrophobin
2.3. Purification and Identification of the Hydrophobin
2.3.1. HPLC Analysis
2.3.2. RP-HPLC Analysis of the Hydrophobin Sa-HFB1 Tryptic Hydrolysate
2.3.3. Matrix-Assisted Laser Desorption Ionization Mass Spectrography (MALDI-MS) and Tandem Mass Spectography (MS|MS)
2.3.4. Hydrophobin Sa-HFB1 Peptides Preparation for MALDI-TOF MS Analysis
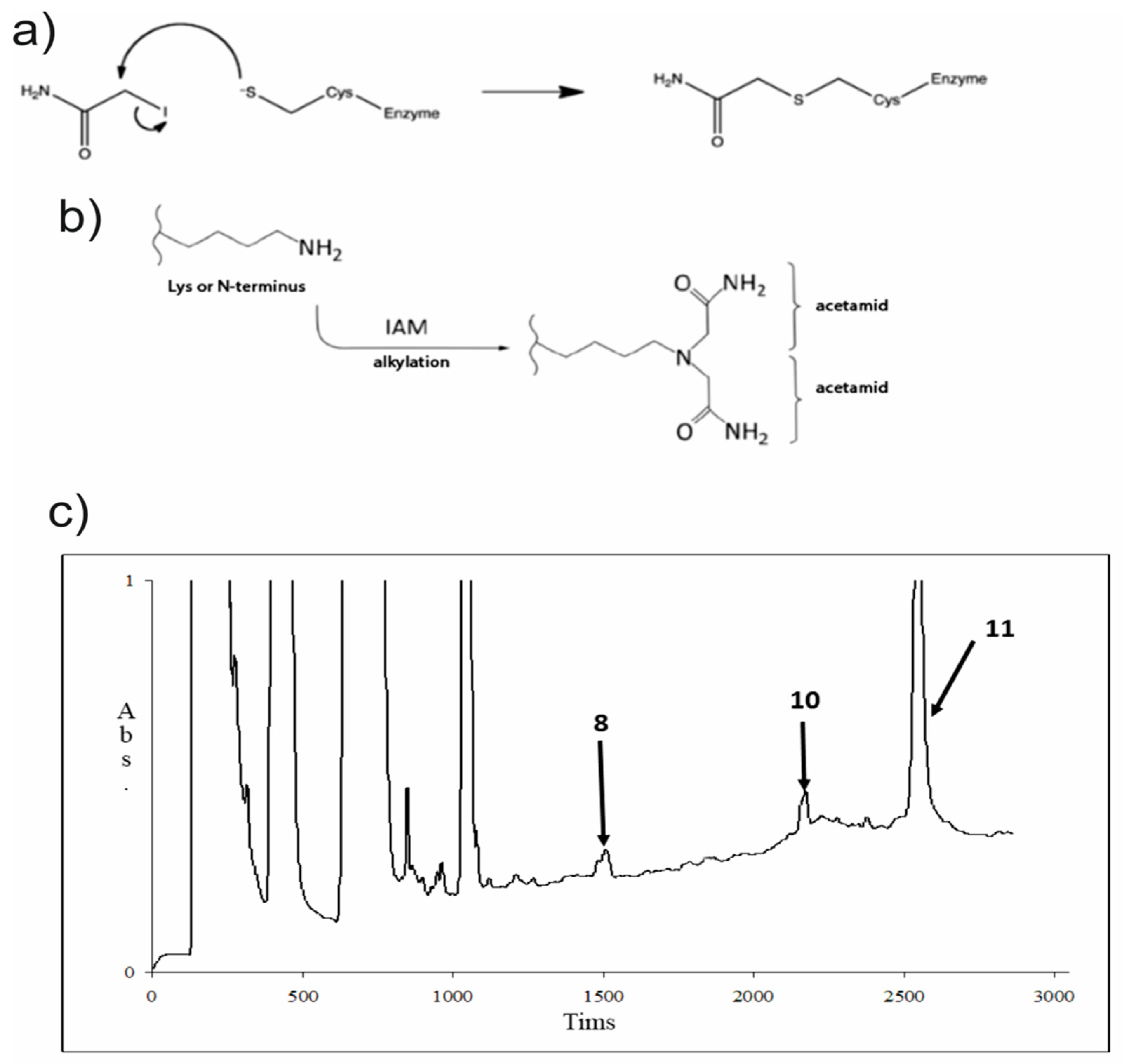
2.3.5. Hydrophobin (Sa-HFB1) Derivate Preparation
2.3.6. Hydrophobin (Sa-HFB1) Proteolysis
2.3.7. Amino Acid Analysis of a Hydrophobin Sa-HFB1 Sample
2.3.8. Edman Sequencing
2.4. Sequences Analysis and Primer Design
2.5. Hydrophobin Gene Sequencing
2.6. Biological Assays
Antifungal Activity
3. Results
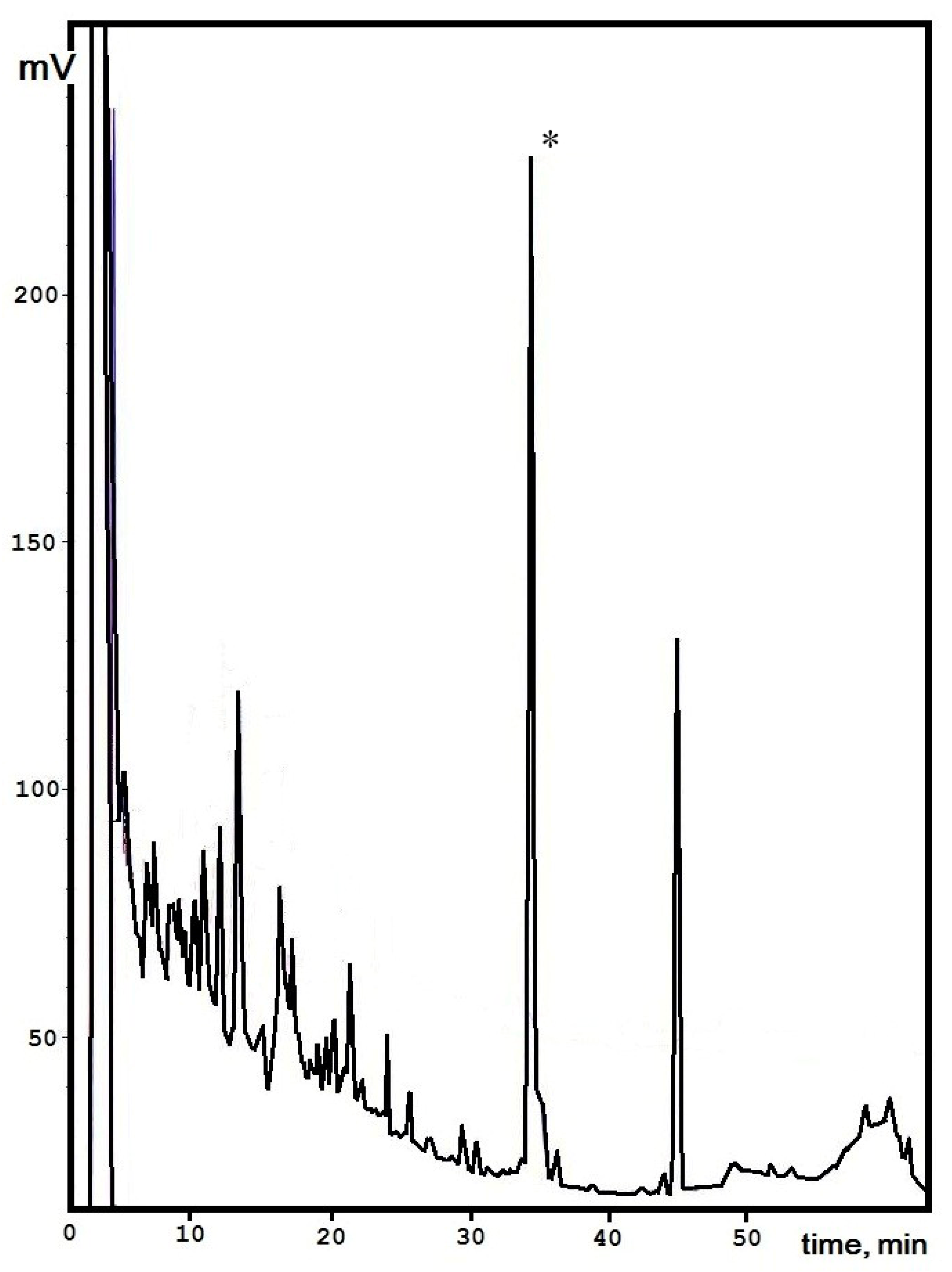
3.1. Isolation of the Active Polypeptide from Culture Broth Extract
3.2. Structural Analysis
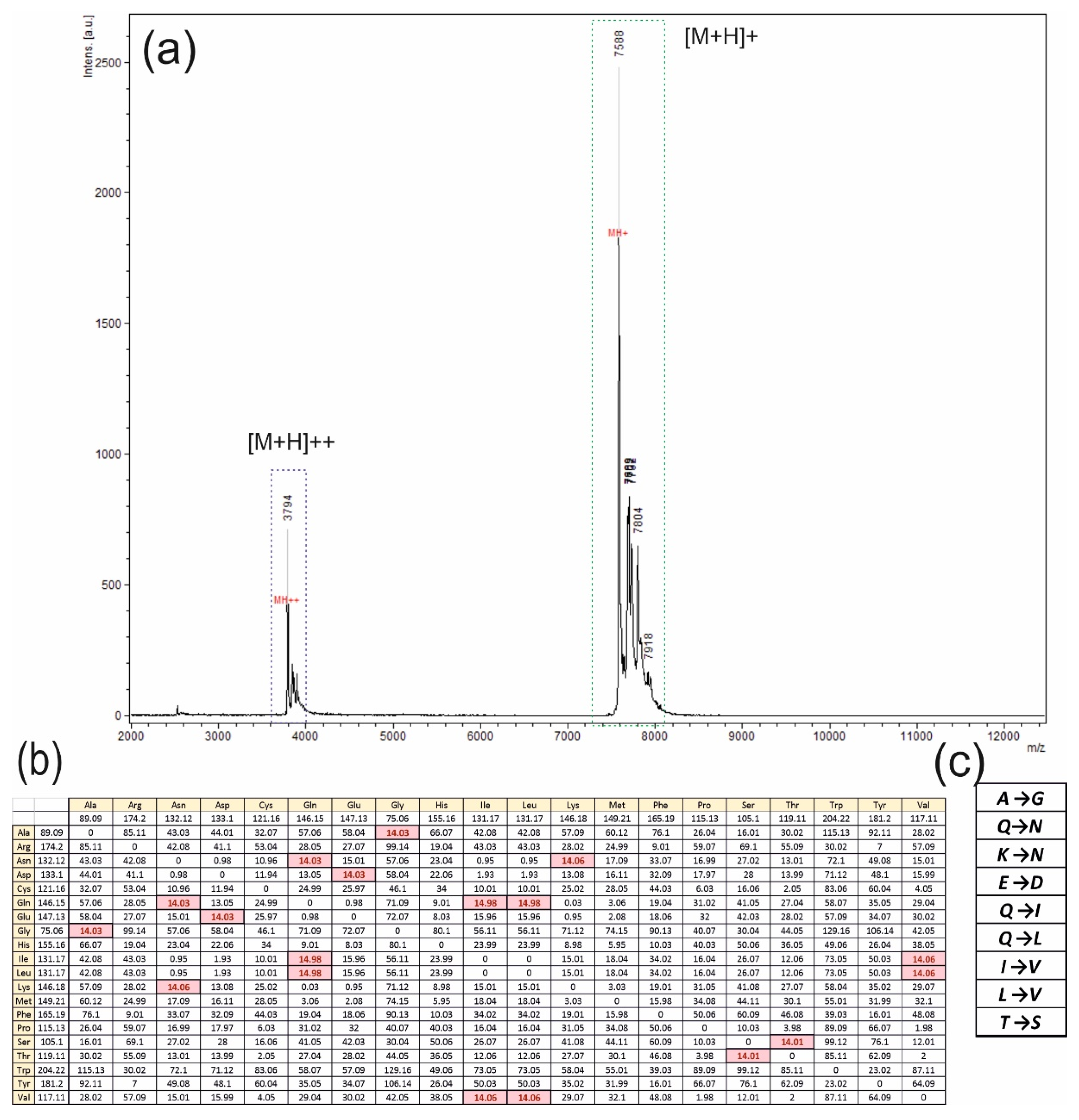
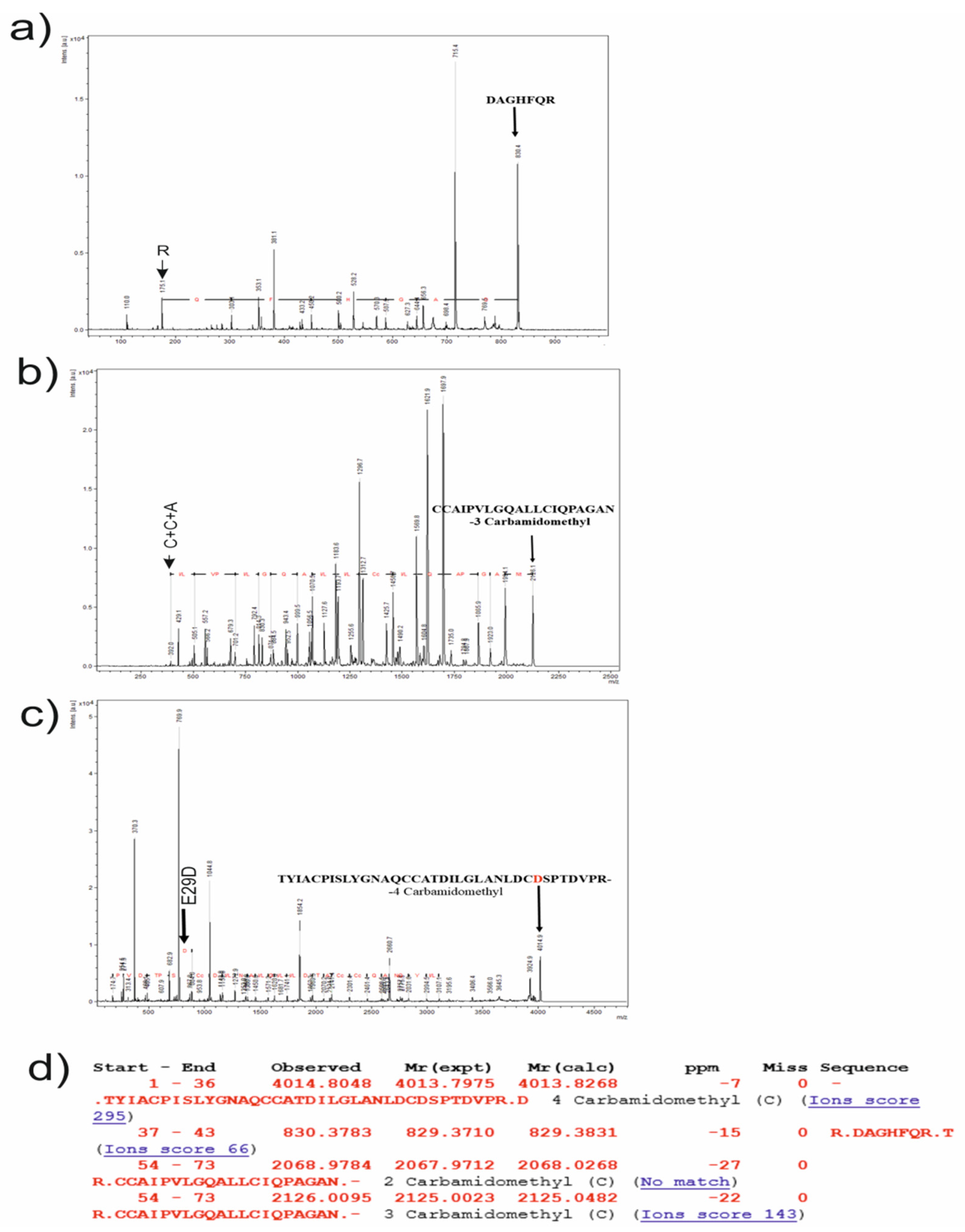
3.3. MALDI-TOF MS
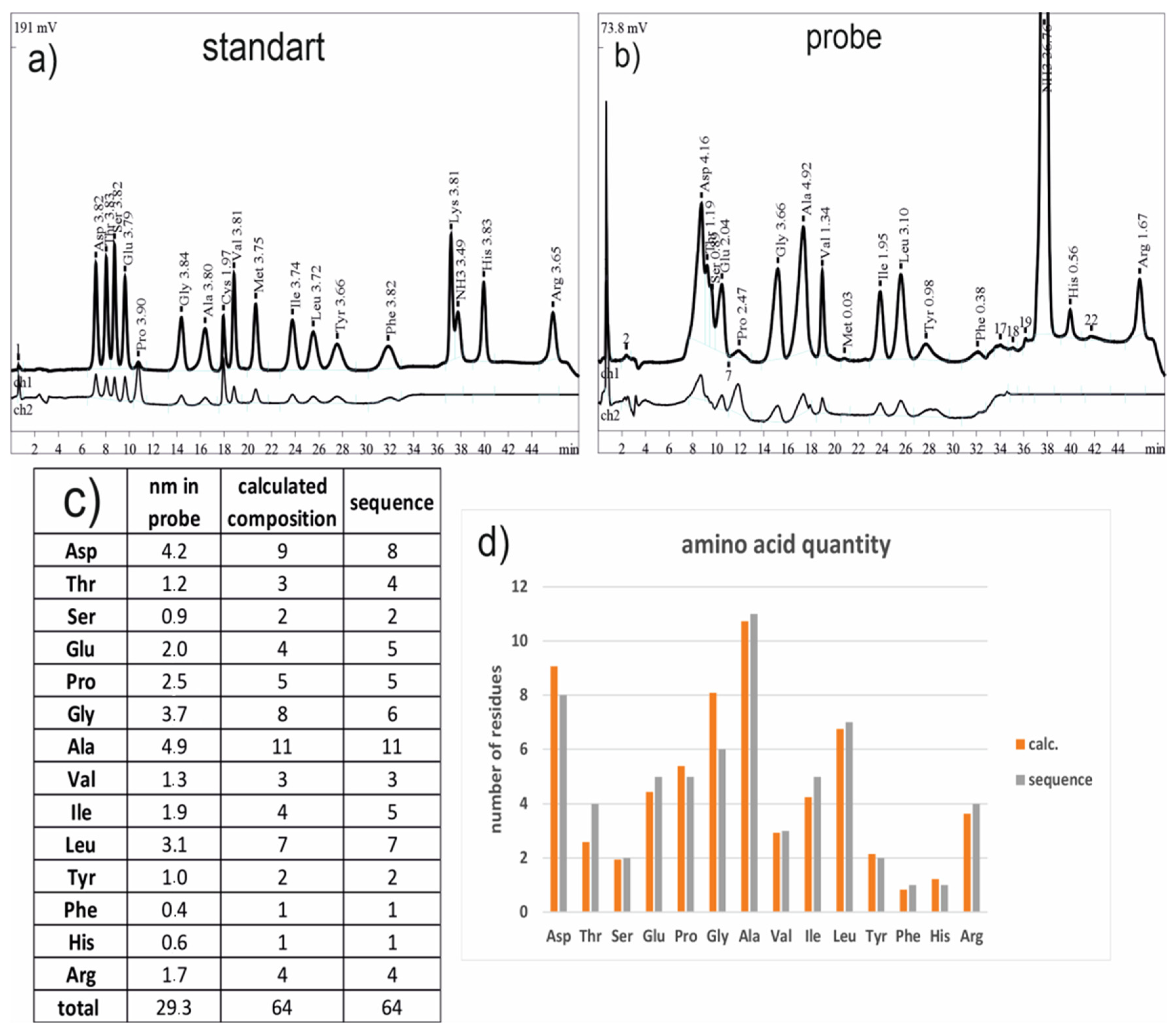
3.4. Amino Acid Analysis
3.5. Proteolysis and Amino Acid Sequencing
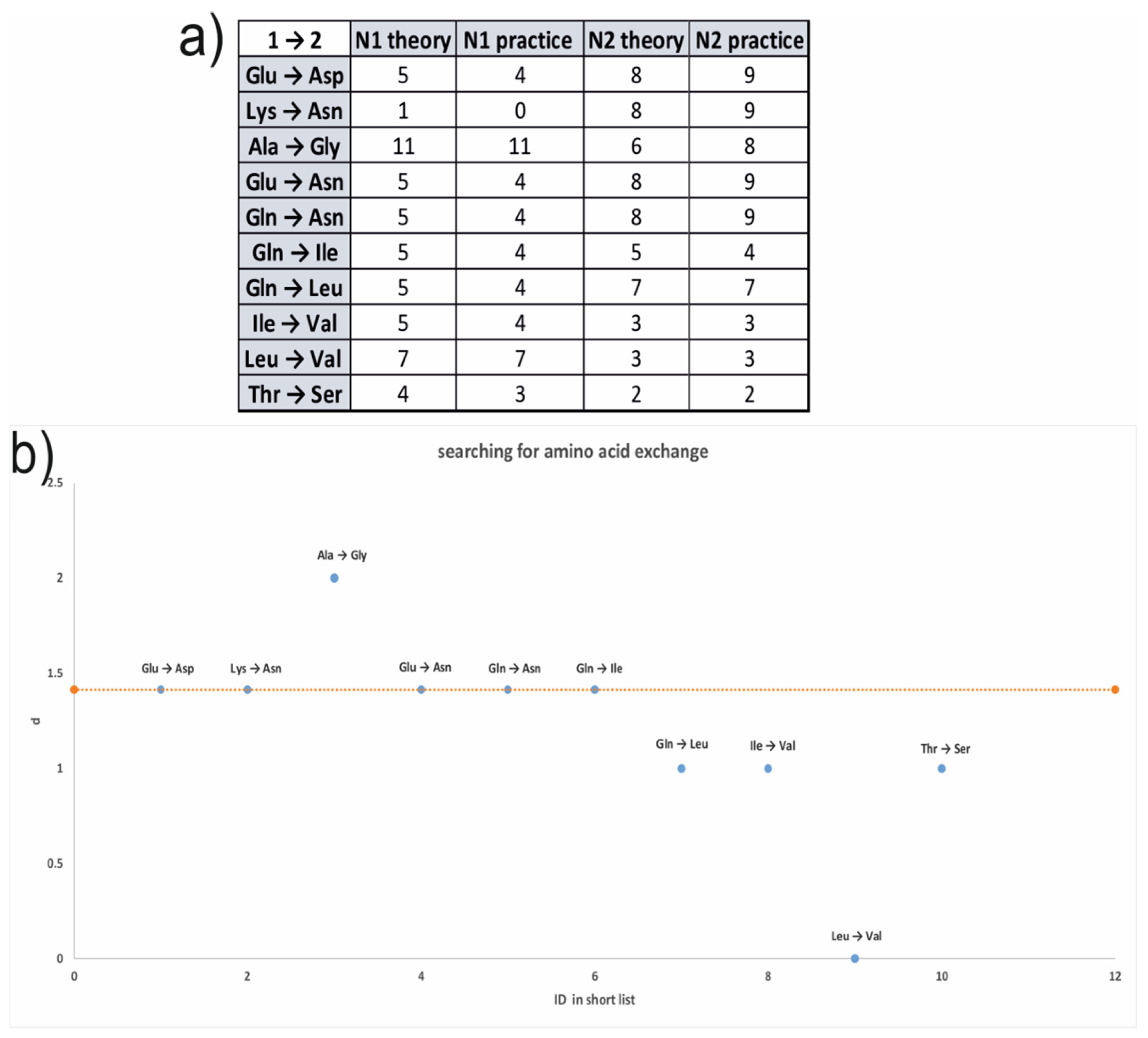
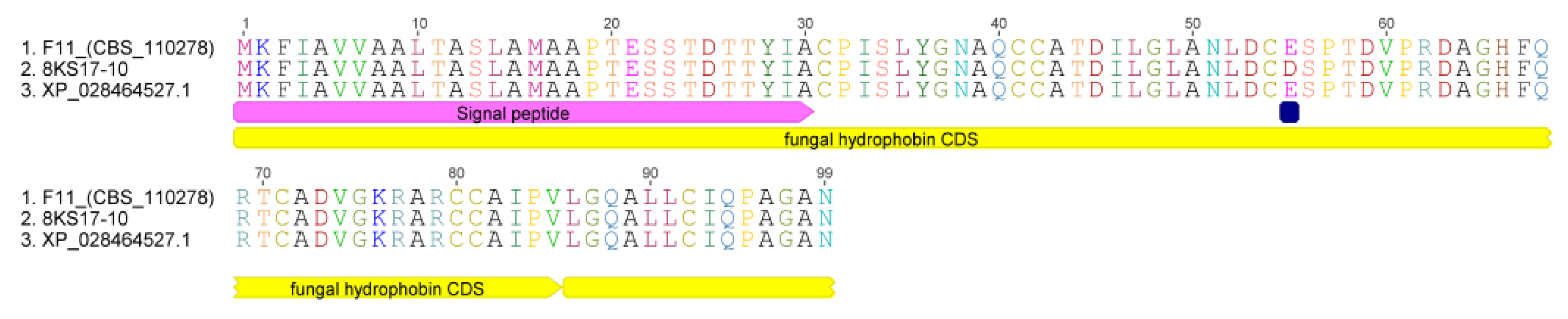
3.6. Sequence Analysis of the S. alkalinus Hydrophobin
3.7. Antifungal Activity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, X.; Li, S.J.; Li, J.J.; Liang, Z.Z.; Zhao, C.Q. Novel natural products from extremophilic fungi. Mar. Drugs 2018, 16, 194. [Google Scholar] [CrossRef] [PubMed]
- Ibrar, M.; Ullah, M.W.; Manan, S.; Farooq, U.; Rafiq, M.; Hasan, F. Fungi from the extremes of life: An untapped treasure for bioactive compounds. Appl. Microbiol. Biotechnol. 2020, 104, 2777–2801. [Google Scholar] [CrossRef] [PubMed]
- Mamo, G.; Mattiasson, B. Alkaliphiles: The versatile tools in biotechnology. In Alkaliphiles in Biotechnology. Advances in Biochemical Engineering/Biotechnology; Mamo, G., Mattiasson, B., Eds.; Springer Nature: Cham, Switzerland, 2020; Volume 172, p. 51. [Google Scholar] [CrossRef]
- Horikoshi, K. Alkaliphiles. In Extremophiles; Springer: Tokyo, Japan, 2016; Volume 4, pp. 53–78. [Google Scholar] [CrossRef]
- Grum-Grzhimaylo, A.A.; Georgieva, M.L.; Bondarenko, S.A.; Debets, A.J.M.; Bilanenko, E.N. On the diversity of fungi from soda soils. Fungal Divers. 2016, 76, 27–74. [Google Scholar] [CrossRef]
- Kevbrin, V.V. Isolation and cultivation of alkaliphiles. In Alkaliphiles in Biotechnology. Advances in Biochemical Engineering/Biotechnology; Mamo, G., Mattiasson, B., Eds.; Springer Nature: Cham, Switzerland, 2020; Volume 172, pp. 53–84. [Google Scholar] [CrossRef]
- Wei, Y.; Zhang, S.H. Abiostress resistance and cellulose degradation abilities of haloalkaliphilic fungi: Applications for saline–alkaline remediation. Extremophiles 2018, 22, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Bilanenko, E.N.; Sorokin, D.; Georgieva, M.L.; Kozlova, M.V. Heleococcum alkalinum a new alkali-tolerant ascomycete from saline soda soils. Mycotaxon 2005, 91, 497–507. [Google Scholar]
- Grum-Grzhimaylo, A.A.; Debets, A.J.M.; van_Diepeningen, A.D.; Georgieva, M.L.; Bilanenko, E.N. Sodiomyces alkalinus, a new holomorphic alkaliphilic ascomycete within the Plectosphaerellaceae. Persoonia 2013, 31, 147–158. [Google Scholar] [CrossRef]
- Deshmukh, S.K.; Dufossé, L.; Chhipa, H.; Saxena, S.; Mahajan, G.B.; Gupta, M.K. Fungal endophytes: A potential source of antibacterial compounds. J. Fungi 2022, 8, 164. [Google Scholar] [CrossRef]
- Kladwang, W.; Bhumirattana, A.; Hywel-Jones, N. Alkaline-tolerant fungi from Thailand. Fungal Divers. 2003, 13, 69–83. [Google Scholar]
- Tiquia-arashiro, S.M.; Grube, M. Fungi in Extreme Environments: Ecological Role and Biotechnological Significance; Grube, M., Ed.; Springer Nature: Cham, Switzerland, 2019; p. 626. [Google Scholar] [CrossRef]
- Alkin, N.; Dunaevsky, Y.; Elpidina, E.; Beljakova, G.; Tereshchenkova, V.; Filippova, I.; Belozersky, M. Proline-specific fungal peptidases: Genomic analysis and identification of secreted DPP4 in alkaliphilic and alkalitolerant fungi. J. Fungi 2021, 7, 744. [Google Scholar] [CrossRef]
- Kuvarina, A.E.; Gavryushina, I.A.; Kulko, A.B.; Ivanov, I.A.; Rogozhin, E.A.; Georgieva, M.L.; Sadykova, V.S. The Emericellipsins A-E from an alkalophilic fungus Emericellopsis alkalina show potent activity against multi-drug-resistant pathogenic fungi. J. Fungi 2021, 7, 153. [Google Scholar] [CrossRef]
- Rogozhin, E.A.; Sadykova, V.S.; Baranova, A.A.; Vasilchenko, A.S.; Lushpa, V.A.; Mineev, K.S.; Georgieva, M.L.; Kul’Ko, A.B.; Krasheninnikov, M.E.; Lyundup, A.V.; et al. A Novel lipopeptaibol Emericellipsin A with antimicrobial and antitumor activity produced by the extremophilic fungus Emericellopsis alkaline. Molecules 2018, 23, 2785. [Google Scholar] [CrossRef] [PubMed]
- Kuvarina, A.E.; Georgieva, M.L.; Rogozhin, E.A.; Kulko, A.B.; Gavryushina, I.A.; Sadykova, V.S. Antimicrobial potential of the alkalophilic fungus Sodiomyces alkalinus and selection of strains–producers of new antimicotic compound. Appl. Biochem. Microbiol. 2021, 57, 86–93. [Google Scholar] [CrossRef]
- Bayry, J.; Aimanianda, V.; Guijarro, J.I.; Sunde, M.; Latgé, J.P. Hydrophobins-unique fungal proteins. PLoS Pathog. 2012, 8, e1002700. [Google Scholar] [CrossRef] [PubMed]
- Galkina, S.I.; Fedorova, N.V.; Serebryakova, M.V.; Arifulin, E.A.; Stadnichuk, V.I.; Gaponov, T.V.; Baratova, L.A.; Sudina, G.F. Inhibition of the GTPase dynamin or actin depolymerisation initiates outward plasma membrane tabulation/vesiculation (cytoneme formation) in neutrophils. Biol. Cell 2015, 107, 144–158. [Google Scholar] [CrossRef]
- Mascot Search Engine. Protein Identification Software for Mass Spectrometry. Available online: https://www.matrixscience.com (accessed on 10 April 2022).
- Tsugita, A.; Scheffler, J.J. A rapid method for acid hydrolysis of protein with a mixture of trifluoroacetic acid and hydrochloric acid. Eur. J. Biochem. 1982, 124, 585–588. [Google Scholar] [CrossRef]
- Trofimova, L.; Ksenofontov, A.; Mkrtchyan, G.; Graf, A.; Baratova, L.; Bunik, V. Quantification of rat brain. Amino acids: Analysis of the data consistency. Curr. Anal. Chem. 2016, 12, 349–356. [Google Scholar] [CrossRef]
- Geneious® 7.1.15. Bioinformatics Software for Sequence Data Analysis. Available online: https://www.geneious.com (accessed on 10 April 2022).
- Primer3 Online Tool (Version 4.1.0) Software. Available online: http://primer3.ut.ee (accessed on 10 April 2022).
- CLSI M27-A3; Reference Method for Broth Dilution Antifungal Susceptibility Testing of Yeasts. Clinical and Laboratory Standards Institute: Pittsburgh, PA, USA, 2013.
- CLSI M38-A2; Reference Method for Broth Dilution Antifungal Susceptibility Testing of Filamentous Fungi; Approved standard. 2nd ed.; Clinical and Laboratory Standards Institute: Pittsburgh, PA, USA, 2008.
- Grum-Grzhimaylo, A.A.; Falkoski, D.L.; van den Heuvel, J.; Valero-Jiménez, C.A.; Min, B.; Choi, I.G.; Lipzen, A.; Daum, C.G.; Aanen, D.K.; Tsang, A.; et al. The obligate alkalophilic soda-lake fungus Sodiomyces alkalinus has shifted to a protein diet. Mol. Ecol. 2018, 27, 4808–4819. [Google Scholar] [CrossRef]
- Fullmer, C.S. Identification of cysteine-containing peptides in protein digests by high-performance liquid chromatography. Anal. Biochem. 1984, 142, 336–339. [Google Scholar] [CrossRef]
- Aitken, A. Analysis of cysteine residues and disulfide bonds. Methods Mol. Biol. 1994, 32, 351–360. [Google Scholar] [CrossRef]
- Shadrivova, O.; Gusev, D.; Vashukova, M.; Lobzin, D.; Gusarov, V.; Zamyatin, M.; Zavrazhnov, A.; Mitichkin, M.; Borzova, Y.; Kozlova, O.; et al. COVID-19-associated pulmonary Aspergillosis in Russia. J. Fungi 2021, 7, 59. [Google Scholar] [CrossRef]
- Revie, N.M.; Iyer, K.R.; Robbins, N.; Cowen, L.E. Antifungal drug resistance: Evolution, mechanisms and impact. Curr. Opin. Microbiol. 2018, 45, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, F.; Liu, M.; Dong, C. Hydrophobin CmHYD1 is involved in conidiation, infection and primordium formation, and regulated by GATA transcription factor CmAreA in edible fungus, Cordyceps militaris. J. Fungi 2021, 7, 674. [Google Scholar] [CrossRef] [PubMed]
- Wösten, H.A. Hydrophobins: Multipurpose proteins. Annu. Rev. Microbiol. 2001, 55, 625–646. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.X.; Chen, R.L.; Yan, J.J.; Long, Y.; Tong, Z.J.; Song, H.B.; Xie, B.G. A hydrophobin gene, Hyd9, plays an important role in the formation of aerial hyphae and primordia in Flammulina filiformis. Gene 2019, 706, 84–90. [Google Scholar] [CrossRef]
- Sammer, D.; Krause, K.; Gube, M.; Wagner, K.; Kothe, E. Hydrophobins in the life cycle of the ectomycorrhizal basidiomycete Tricholoma vaccinum. PLoS ONE 2016, 11, e0167773. [Google Scholar] [CrossRef]
- Linder, M.B.; Szilvay, G.R.; Nakari-Setälä, T.; Penttilä, M.E. Hydrophobins: The protein-amphiphiles of filamentous fungi. FEMS Microbiol. Rev. 2005, 29, 877–896. [Google Scholar] [CrossRef]
- Wang, L.; Lu, C.; Fan, M.; Liao, B. Coriolopsis trogii hydrophobin genes favor a clustering distribution and are widely involved in mycelial growth and primordia formation. Gene 2021, 802, 145863. [Google Scholar] [CrossRef]
- Xu, D.; Wang, Y.; Keerio, A.A.; Ma, A. Identification of hydrophobin genes and their physiological functions related to growth and development in Pleurotus ostreatus. Microbiol. Res. 2021, 247, 126723. [Google Scholar] [CrossRef]
- Khalesi, M.; Zune, Q.; Telek, S.; Riveros-Galan, D.; Verachtert, H.; Toye, D.; Gebruers, K.; Derdelinckx, G.; Delvigne, F. Fungal biofilm reactor improves the productivity of hydrophobin HFBII. Biochem. Eng. J. 2014, 88, 171–178. [Google Scholar] [CrossRef]
- Kulkarni, S.S.; Nene, S.N.; Joshi, K.S. Exploring malted barley waste for fungi producing surface active proteins like hydrophobins. SN Appl. Sci. 2020, 2, 1884. [Google Scholar] [CrossRef]
- Siddiquee, R.; Choi, S.S.C.; Lam, S.S.; Wang, P.; Qi, R.; Otting, G.; Sunde, M.; Kwan, A.H. Cell-free expression of natively folded hydrophobins. Protein Expr. Purif. 2020, 170, 105591. [Google Scholar] [CrossRef] [PubMed]
- Landeta-Salgado, C.; Cicatiello, P.; Lienqueo, M.E. Mycoprotein and hydrophobin like protein produced from marine fungi Paradendryphiella salina in submerged fermentation with green seaweed Ulva spp. Algal Res. 2021, 56, 102314. [Google Scholar] [CrossRef]
- Mosbach, A.; Leroch, M.; Mendgen, K.W.; Hahn, M. Lack of evidence for a role of hydrophobins in conferring surface hydrophobicity to conidia and hyphae of Botrytis cinerea. BMC Microbiol. 2011, 11, 10. [Google Scholar] [CrossRef]
- Sevim, A.; Donzelli, B.G.; Wu, D.; Demirbag, Z.; Gibson, D.M.; Turgeon, B.G. Hydrophobin genes of the entomopathogenic fungus, Metarhizium brunneum, are differentially expressed and corresponding mutants are decreased in virulence. Curr. Genet. 2012, 58, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.Y.; Ligoxygakis, P.; Xia, Y.X. HYD3, a conidial hydrophobin of the fungal entomopathogen Metarhizium acridum induces the immunity of its specialist host locust. Int. J. Biol. Macromol. 2020, 165, 1303–1311. [Google Scholar] [CrossRef] [PubMed]
- Dubey, M.K.; Jensen, D.F.; Karlsson, M. Hydrophobins are required for conidial hydrophobicity and plant root colonization in the fungal biocontrol agent Clonostachys rosea. BMC Microbiol. 2014, 14, 18. [Google Scholar] [CrossRef] [PubMed]
- Brown, N.A.; Ries, L.N.A.; Reis, T.F.; Rajendran, R.; dos Santos, R.A.C.; Ramage, G.; Riano-Pachon, D.M.; Goldman, G.H. RNAseq reveals hydrophobins that are involved in the adaptation of Aspergillus nidulans to lignocellulose. Biotechnol. Biofuels 2016, 9, 145. [Google Scholar] [CrossRef] [PubMed]
- Valsecchi, I.; Dupres, V.; Stephen, V.E.; Guijarro, J.I.; Gibbons, J.; Beau, R.; Bayry, J.; Coppee, J.Y.; Lafont, F.; Latge, J.P.; et al. Role of hydrophobins in Aspergillus fumigatus. J. Fungi 2018, 4, 2. [Google Scholar] [CrossRef]
- Linder, M.B. Hydrophobins: Proteins that self-assemble at interfaces. Curr. Opin. Colloid Interface Sci. 2009, 14, 356–363. [Google Scholar] [CrossRef]
- Casarrubia, S.; Daghino, S.; Kohler, A.; Morin, E.; Khouja, H.R.; Daguerre, Y.; Veneault-Fourrey, C.; Martin, F.M.; Perotto, S.; Martino, E. The hydrophobin-like OmSSP1 may be an effector in the ericoid mycorrhizal symbiosis. Front. Plant Sci. 2018, 9, 546. [Google Scholar] [CrossRef]
- Ball, S.R.; Kwan, A.H.; Sunde, M. Hydrophobin rodlets on the fungal cell wall. Curr. Top. Microbiol. Immunol. 2020, 425, 29–51. [Google Scholar] [CrossRef] [PubMed]
- Salgado, C.L.; Cicatiello, P.; Stanzione, I.; Medino, D.; Mora, I.B.; Gomez, C.; Lienqueo, M.E. The growth of marine fungi on seaweed polysaccharides produces cerato-platanin and hydrophobin self-assembling proteins. Microbiol. Res. 2021, 251, 126835. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, F.; Xu, Y.Y.; Liu, G.J.; Dong, C.H. Cysteine-rich hydrophobin gene family: Genome wide analysis, phylogeny and transcript profiling in Cordyceps militaris. Int. J. Mol. Sci. 2021, 22, 643. [Google Scholar] [CrossRef] [PubMed]
- He, R.; Li, C.; Feng, J.; Zhang, D. A class II hydrophobin gene, Trhfb3, participates in fungal asexual development of Trichoderma reesei. FEMS Microbiol. Lett. 2017, 364, fnw297. [Google Scholar] [CrossRef]
- Khalesi, M.; Jahanbani, R.; Riveros-Galan, D.; Sheikh-Hassani, V.; Sheikh-Zeinoddin, M.; Sahihi, M.; Winterburn, J.; Derdelinckx, G.; Moosavi-Movahedi, A.A. Antioxidant activity and ACE-inhibitory of class II hydrophobin from wild strain Trichoderma reesei. Int. J. Biol. Macromol. 2016, 91, 174–179. [Google Scholar] [CrossRef][Green Version]
- Ruocco, M.; Lanzuise, S.; Lombardi, N.; Woo, S.L.; Vinale, F.; Marra, R.; Varlese, R.; Manganiello, G.; Pascale, A.; Scala, V.; et al. Multiple roles and effects of a novel Trichoderma hydrophobin. Mol. Plant-Microbe Interact. 2015, 28, 167–179. [Google Scholar] [CrossRef]
- Zhang, H.; Ji, S.; Guo, R.; Zhou, C.; Wang, Y.; Fan, H.; Liu, Z. Hydrophobin HFBII-4 from Trichoderma asperellum induces antifungal resistance in poplar. Braz. J. Microbiol. 2019, 50, 603–612. [Google Scholar] [CrossRef]
- Cai, F.; Gao, R.; Zhao, Z.; Ding, M.; Jiang, S.; Yagtu, C.; Zhu, H.; Zhang, J.; Ebner, T.; Mayrhofer-Reinhartshuber, M.; et al. Evolutionary compromises in fungal fitness: Hydrophobins can hinder the adverse dispersal of conidiospores and challenge their survival. ISME J. 2020, 14, 2610–2624. [Google Scholar] [CrossRef]
- Vereman, J.; Thysens, T.; Derdelinckx, G.; Impe, J.V.; de Voorde, I.V. Extraction and spray drying of class II hydrophobin HFBI produced by Trichoderma reesei. Process Biochem. 2019, 77, 159–163. [Google Scholar] [CrossRef]
- Winandy, L.; Hilpert, F.; Schlebusch, O.; Fischer, R. Comparative analysis of surface coating properties of five hydrophobins from Aspergillus nidulans and Trichoderma reseei. Sci. Rep. 2018, 8, 12033. [Google Scholar] [CrossRef]
- Kuvarina, A.E.; Gavryushina, I.A.; Sykonnikov, M.A.; Efimenko, T.A.; Markelova, N.N.; Bilanenko, E.N.; Bondarenko, S.A.; Kokaeva, L.Y.; Timofeeva, A.V.; Serebryakova, M.V.; et al. Exploring peptaibol’s profile, antifungal, and antitumor activity of Emericellipsin A of Emericellopsis species from soda and saline soils. Molecules 2022, 27, 1736. [Google Scholar] [CrossRef] [PubMed]
- Kozlova, M.V.; Ianutsevich, E.A.; Danilova, O.A.; Kamzolkina, O.V.; Tereshina, V.M. Lipids and soluble carbohydrates in the mycelium and ascomata of alkaliphilic fungus Sodiomyces alkalinus. Extremophiles 2019, 23, 487–493. [Google Scholar] [CrossRef] [PubMed]
- Bondarenko, S.A.; Ianutsevich, E.A.; Danilova, O.A.; Grum-Grzhimaylo, A.A.; Kotlova, E.R.; Kamzolkina, O.V.; Bilanenko, E.N.; Tereshina, V.M. Membrane lipids and soluble sugars dynamics of the alkaliphilic fungus Sodiomyces tronii in response to ambient pH. Extremophiles 2017, 21, 743–754. [Google Scholar] [CrossRef] [PubMed]
- Bondarenko, S.A.; Yanutsevich, E.A.; Sinitsyna, N.A.; Georgieva, M.L.; Bilanenko, E.N.; Tereshina, V.M. Dynamics of the cytosol soluble carbohydrates and membrane lipids in response to ambient pH in alkaliphilic and alkalitolerant fungi. Microbiology 2018, 87, 21–32. [Google Scholar] [CrossRef]
- Bondarenko, S.A.; Georgieva, M.L.; Bilanenko, E.N. Fungi inhabiting the coastal zone of lake Magadi. Contemp. Probl. Ecol. 2018, 11, 439–448. [Google Scholar] [CrossRef]
- Sallada, N.D.; Dunn, K.J.; Berger, B.W. A structural and functional role for disulfide bonds in a class II hydrophobin. Biochemistry 2017, 57, 645–653. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zone, mm | ||||
|---|---|---|---|---|
| Compound, 40 µg/disk | ||||
| Strain | Sa-HFB1 | AmpB | FZ | VOR |
| Candida albicans 1582 m | 15 ± 0.1 | 10 ± 0.6 | 0 | 10 ± 0.6 |
| Pichia kudriavzevii 1402 m | 16 ± 0.3 | 15 ± 0.1 | 0 | 10 ± 0.6 |
| Nakaseomyces glabrataa 1447 m | 12.5 ± 0.2 | 0 | 0 | 0 |
| C. tropicalis 156 m | 14 ± 0.1 | 0 | 0 | 0 |
| C. parapsilosis 571 m | 14 ± 0.2 | 18 ± 0.3 | 0 | 0 |
| Cryptococcus neoformans 297 m | 30 ± 0.1 | 18 ± 0.6 | 0 | 0 |
| Aspergillus fumigatus 390 m | 12 ± 0.5 | 9 ± 0.6 | 0 | 0 |
| A. niger 219 | 14 ± 0.1 | 15 ± 0.8 | 0 | 11 ± 0.6 |
| Penicillium brevicompactum VKM F-4481 | 15 ± 0.5 | 17 ± 0.4 | 0 | 10 ± 0.9 |
| Minimal Inhibitory Concentration (MIC, µg/mL) | ||||
|---|---|---|---|---|
| Compound | ||||
| Strain | Sa-HFB1 | AmpB | FZ | IZ |
| Aspergillus niger 219 | 8 | 1 | R | 4 |
| A. fumigatus 390 m | 8 | 1 | R | 0.5 |
| Candida albicans ATCC 2091 | 4 | 1 | >64 | 4 |
| C. albicans 1582 m | 4 | 2 | >64 | 4 |
| Cryptococcus neoformans 297 m | 1 | 0.5 | 16 | 0.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuvarina, A.E.; Rogozhin, E.A.; Sykonnikov, M.A.; Timofeeva, A.V.; Serebryakova, M.V.; Fedorova, N.V.; Kokaeva, L.Y.; Efimenko, T.A.; Georgieva, M.L.; Sadykova, V.S. Isolation and Characterization of a Novel Hydrophobin, Sa-HFB1, with Antifungal Activity from an Alkaliphilic Fungus, Sodiomyces alkalinus. J. Fungi 2022, 8, 659. https://doi.org/10.3390/jof8070659
Kuvarina AE, Rogozhin EA, Sykonnikov MA, Timofeeva AV, Serebryakova MV, Fedorova NV, Kokaeva LY, Efimenko TA, Georgieva ML, Sadykova VS. Isolation and Characterization of a Novel Hydrophobin, Sa-HFB1, with Antifungal Activity from an Alkaliphilic Fungus, Sodiomyces alkalinus. Journal of Fungi. 2022; 8(7):659. https://doi.org/10.3390/jof8070659
Chicago/Turabian StyleKuvarina, Anastasia E., Eugene A. Rogozhin, Maxim A. Sykonnikov, Alla V. Timofeeva, Marina V. Serebryakova, Natalia V. Fedorova, Lyudmila Y. Kokaeva, Tatiana A. Efimenko, Marina L. Georgieva, and Vera S. Sadykova. 2022. "Isolation and Characterization of a Novel Hydrophobin, Sa-HFB1, with Antifungal Activity from an Alkaliphilic Fungus, Sodiomyces alkalinus" Journal of Fungi 8, no. 7: 659. https://doi.org/10.3390/jof8070659
APA StyleKuvarina, A. E., Rogozhin, E. A., Sykonnikov, M. A., Timofeeva, A. V., Serebryakova, M. V., Fedorova, N. V., Kokaeva, L. Y., Efimenko, T. A., Georgieva, M. L., & Sadykova, V. S. (2022). Isolation and Characterization of a Novel Hydrophobin, Sa-HFB1, with Antifungal Activity from an Alkaliphilic Fungus, Sodiomyces alkalinus. Journal of Fungi, 8(7), 659. https://doi.org/10.3390/jof8070659