1. Introduction
The higher fungi of
Agaricomycetes class (
Basidiomycota division) are a huge source of a wide range of natural structurally diverse bioactive compounds with promising nutritious and therapeutic properties and have great potential as nutraceuticals [
1,
2] The basidiomycetes have a remarkable history and have been used in traditional Asian medicine for thousands of years because of their beneficial effects on human health, with no negative side effects [
3,
4]. Despite the fact that species from
Basidiomycota were much less studied than
Ascomycota representatives, approximately 700 basidiomycete species have been shown to have pharmacological activity due to the bioactive metabolites that they synthesize [
5]. The main bioactive substances produced by basidiomycetes are homo- and hetero-polysaccharides, lectins, phenolic and flavonoid compounds, terpenoids, sterols and volatile organic compounds [
2,
6,
7]. The diversity of bioactivities could be explained by the fact that mushrooms’ natural habitats are dark places and highly competitive environments, combined with constant attack by numerous bacterial species. Due to this, fungal strains create their own barrier systems and synthesize numerous bioactive metabolites that enable them to survive in their natural environment [
8]. This is the reason why research related to the discovery of new components produced by higher fungi, as well as the determination of their in vitro and in vivo biological activity, has increased exponentially in the last ten years [
4,
9,
10]. Beyond their nutritional value, mushroom metabolites have been shown to exhibit a plethora of bioactive properties such as antioxidant properties, anticarcinogenic activity, immunomodulatory action, prebiotic activity, antibacterial and antiviral activity, anti-inflammatory actions, antihypoglycemic activities, neuroprotective and anti-aging properties, etc. [
2,
6]. There is evidence that β-glucan polysaccharides, obtained from mushrooms, possess antiviral activity and could fight against coronavirus SARS-CoV-2 disease (COVID-19) due to different mechanisms of immunomodulation [
11,
12].
The mushroom species which are most often used as food for thousands of years and which demonstrate potential to synthesize bioactive components with therapeutic properties are
Ganoderma lucidum,
Grifola frondosa,
Lentinus edodes,
Schizophyllum commune,
Trametes versicolor,
Hericium erinaceus,
Inonotus obliquus,
Phellinus linteus and Pleuortus ostreatus [
13,
14,
15,
16,
17]. Most of them are referred to as edible and the basidiocarps of
G. lucidum,
T. versicolor and
I. obliquus are referred to as non-edible medicinal mushrooms because of their bitter taste and toughness [
18,
19,
20].
The widespread cultivation techniques for macromycetes are solid-state ones which lead to fruiting body formation, a very long and laborious process. The proper controlled submerged cultivation in an appropriate nutrient medium is a promising approach leading to the increased production of macrofungal biomass and bioactive substances with consistent quality, decreased production time, reduced downstream processing cost, decreased contamination risks and assured sustainability possibility [
2,
21,
22,
23,
24]. Serious efforts are being made to increase the yield of bioactive metabolites by the optimization of the cultivation medium composition and conditions [
23,
25,
26,
27,
28], together with exploring the genetic mechanisms of the biosynthetic pathways and their regulation [
29].
One of the most studied medicinal mushrooms in the past decade is
T. versicolor, commonly known as Turkey tail. The species belongs to
Basidiomycota phylum, order
Polyporales, family
Polyporaceae. Due to its high lignin degrading enzyme activities, the application of
T. versicolor in mycoremediation processes is well known [
30]. On the other hand,
T. versicolor has been used in traditional medicine for centuries and lately is a part of modern cancer treatment because of its diversity of bioactive compounds with an enormous variety of chemical structure and physiological activities [
31]. The best investigated carriers of biological activity are the polysaccharopeptides (PSPs) obtained from submerged batch cultivation of
T. versicolor [
32,
33]. They exhibited antitumor, hepatoprotective and analgesic activities [
34,
35,
36,
37]. Additionally, it was reported that these compounds demonstrate significant immunomodulatory activity and have promising anti-diabetic properties as α-glucosidase inhibitors [
7,
38]. It was reported that the protein-bound polysaccharides (PSPs) were nontoxic during prolonged use in the treatment of cancers by suppressing DNA/RNA synthesis and enhancing immune function [
32,
39,
40]. In recent years,
T. versicolor has attracted the attention of scientists not only as a source of pharmacologically active substances, but also as adjuvants in conventional chemo-or radiation therapy to reduce their side effects or to enhance their potency [
41].
The aim of our study was to investigate the composition of mycelial biomass and crude EPS obtained by submerged cultivation of a local macrofungal T. versicolor NBIMCC 8939 strain, which had previously been studied for its application in mycoremediation processes. The evaluation of the antioxidant activity of different biomass extracts together with the prebiotic and in vitro anti-inflammatory activity of the crude EPS is a prerequisite to reveal the nutraceutical potential of the mycelium obtained by submerged cultivation and the synthesized extracellular exopolysaccharides, in addition to the established mycoremediation capacity of T. versicolor NBIMCC 8939.
2. Materials and Methods
2.1. Mushroom
The macrofungal strain T. versicolor NBIMCC 8939 is part of the microbial collection of the Department of Biotechnology, University of Food Technologies, Plovdiv, Bulgaria. The strain was previously isolated, molecularly identified and deposited in the National Bank for Industrial Microorganisms and Cell Cultures. The strain was maintained at 4 °C on Mushroom Complete medium (MCM) containing 20.0 g/L of glucose, 0.5 g/L of KH2PO4, 1.0 g/L of K2HPO4, 0.5 g/L of MgSO4 × H2O, 2.0 g/L of peptone, 2.0 g/L of yeast extract, 2.0 g/L of agar, pH 4.8–5.2, and was subcultured every 30 days onto fresh medium.
2.2. Submerged Cultivation of T. versicolor NBIMCC 8939
Previously optimized by Angelova et al. [
42], liquid Yeast Malts Extract Medium (YM) with the following content: 40.32 g/L of glucose, 3.51 g/L of yeast extract, 3.51 g/L of peptone, 3.00 g/L of malt extract, 7.09 g/L of (NH
4)
2SO
4, 0.5 g/L of KCl, 0.5 g/L of MgSO
4 × 7H
2O and 0.01 g/L of FeSO
4 × 7H
2O, pH 6.0, was used for submerged cultivation of
T. versicolor for biomass and EPS production. The vegetative inoculum was prepared from a 7-day old culture of
T. versicolor grown on MCM agar slants. Each Erlenmeyer flask, containing 100 mL nutrient medium, was inoculated with vegetative biomass from a single MCM-slant culture. The submerged cultivation was carried out at 28 °C on a rotary shaker at 220 rpm for 9 days. Then the mycelium biomass was separated by filtration, washed with distilled water and lyophilized. The biomass was presented as grams dry weight per liter (g DW/L). The culture liquid was used for EPS precipitation.
2.3. Isolation of Exopolysaccharides
EPS were isolated from the culture liquid by overnight freezing at −18 °C followed by thawing at room temperature. The precipitated EPS were recovered by centrifugation (6000×
g rpm, 20 min, 5 °C). The recovered crude EPS were dried in a laboratory dryer (PolEco, Poznań, Poland) at 30 °C for 12 h and the mass in grams was determined. The total carbohydrate content of the crude EPS was determined using the phenol-sulfuric method [
43] with glucose as a standard and the protein content was assessed spectrophotometrically [
44].
2.4. Preparation of Mycelial Biomass Extracts
Distilled water, ethanol (80%, v/v) and methanol were used as extracting solvents for obtaining biomass extract. The lyophilized biomass (1.5 ± 0.05 g) was ground, precisely weighed and mixed with 30 mL distilled water, 80% ethanol or methanol, respectively, then left in a laboratory shaker at 25 °C for 24 h. After that, the extracts were separated from the mycelium biomass by centrifugation. The biomass was treated with an additional 15 mL solvent at the same conditions. After the second centrifugation, the extracts were combined and kept at −8 °C.
2.5. Characterization of Mycelium Biomass
2.5.1. Dietary Fibers Content Analysis
The total (TDF), soluble (SDF) and insoluble (IDF) dietary fibers were determined with K-TDFR-100A (Megazyme Int., Dublin, Ireland), according to AOAC method 991.43 “Total, soluble and insoluble dietary fibers in foods” (First action 1991) and AACC method 32–07.01 “Determination of soluble, insoluble and total dietary fibers in foods and food products” (Final approval 10-16-91) [
45]. All values for TDF, SDF and IDF were expressed as g/100 g of a DW biomass
2.5.2. Glucans Content Analysis
Contents of total, α-and β-glucans were determined in the biomass and EPS using the Mushroom and Yeast β-glucan Assay Kit (Megazyme Int., Dublin, Ireland), following the instruction of the manufacturer. Briefly, to estimate the total glucans content in the samples, 2 mL ice-cold 12 M sulfuric acid was used for hydrolysis of the polysaccharides in the samples for 2 h at 100 °C. After neutralization, hydrolysis proceeded to glucose using a mixture of exo-β-(1,3)-D-glucanase plus β-glucosidase in sodium acetate buffer (pH 4.5) for 1 h at 40 °C. Enzymatic hydrolysis with amyloglucosidase and invertase was conducted for α-glucan content estimation. To estimate total glucan and α-glucan content, glucose oxidase/peroxidase reagent was added and the absorbance of the samples was measured at 510 nm. The β-glucan content was calculated by subtracting the α-glucan from the total glucan content. All values of total, α- and β-glucans in biomass were expressed as g/100 g of a DW biomass.
2.5.3. Total Polyphenol (TPC) and Flavonoid Content Analysis
The total polyphenol content of the fungal biomass extracts was analyzed using the Folin-Ciocalteu method of Kujala et al. [
46] with some modifications. Each sample (1 mL) was mixed with 0.5 mL of Folin-Ciocalteu’s phenol reagent and 0.4 mL of 7.5% Na
2CO
3. The mixture was vortexed well and left for 5 min at 50 °C. After incubation, the absorbance was measured at 765 nm. The TPC in the extracts was expressed as mg gallic acid equivalent (GAE) per g dry weight (mg GAE/g DW).
The total flavonoid content of the fungal biomass extracts was evaluated according to the method described by Kivrak et al. [
47]. An aliquot of 0.5 mL of the sample was added to 0.1 mL of 10% Al(NO
3)
3, 0.1 mL of 1 M CH
3COOK and 3.8 mL ethanol. After incubation for 40 min at room temperature, the absorbance was measured (415 nm). Quercetin (QE) was used as a standard and the results were expressed as mg QE/g DW.
2.5.4. Total Protein and Amino Acid Determination
The total nitrogen of the
T. versicolor biomass was determined by the Kjeldahl method [
45]. The total protein content was calculated by multiplying the total nitrogen by 4.38. The result was expressed as a percentage.
The amino acid composition was determined by the method described by Tumbarski et al. [
48]. The lyophilized biomass was subjected to acid hydrolysis using 6N HCl for 24 h at 105 °C. An aliquot of the hydrolysate was derivatised using AccQ-Fluor reagent Kit (Waters). The derivative was separated on RP AccQ-Tag™ silica-bonded amino acid column C18, 3.9 mm × 150 mm (Waters, Etten-Leur, The Netherlands), and conditioned at 37 °C using an ELITE LaChrom HPLC system (VWR™ Hitachi, Tokyo, Japan). A sample of 20 μL was injected and the elution of the amino acids was performed by the following gradient system: eluent A, buffer WAT052890 (Waters, Etten-Leur, The Netherlands) and eluent B, 60% acetonitrile (Merck KGaA, Darmstadt, Germany) with a constant flow rate of 1.0 mL/min. The amino acids were detected using a diode array detector (DAD) at 254 nm. The amino acid peaks were then analyzed using EZChrom Elite™ software [
49] and the amino acid content was calculated based on the amino acid standard calibration curve (amino acid standard H, Thermo Fisher Scientific, Waltham, MA, USA). The results were expressed as mg AA/g sample and as a percentage [
50].
2.6. Chemical and Structural Characterization of the Crude Exopolysaccharides
2.6.1. Analysis of Monosaccharide Composition
The determination of individual neutral sugars, galacturonic acid and glucuronic acid was performed as follows: 10 mg crude EPS was hydrolyzed with 15 mL 2 M trifluoroacetic acid (Merck KGaA, Darmstadt, Germany) for 3 h at 120 °C. In order to remove the trifluoroacetic acid, the hydrolysate was evaporated to dryness under a vacuum and dissolved in 10 mL deionized water; this procedure was repeated three times. The residue from the last evaporation was dissolved in 1 mL deionized water. The quantities of galactose, rhamnose, fucose, galacturonic and glucuronic acid were determined by chromatographic system ELITE LaChrome (Hitachi High-Tech Corporation, Ibaraki, Japan) HPLC with a VWR Hitachi Chromaster 5450 with refractive index detector using an Aminex HPX-85H column. The samples and standards were eluted with 5 mM H2SO4 (Merck KGaA, Darmstadt, Germany) at an elution rate of 0.5 mL/min, column temperature of 50 °C and detector temperature of 35 °C. The amounts of xylose and mannose were determined separately with the same chromatographic system using a Sugar SP0810 (Shodex®) column. The samples and standards were eluted with ultrapure water at an elution rate of 1.0 mL/min, column temperature of 85 °C and detector temperature of 35 °C.
2.6.2. Molecular Weight Measurement
The molecular weight of the crude EPS was determined by size exclusion chromatography using an ELITE LaChrome (Hitachi High-Tech Corporation, Ibaraki, Japan) HPLC system with a VWR Hitachi Chromaster 5450 with refractive index detector and an OHpak SB-806M (Shodex®) column. The samples and standards were eluted with 0.1M NaNO3 at an elution rate of 0.8 mL/min, column, temperature 30 °C and detector temperature 35 °C. The column was equilibrated with Shodex pullulan (Showa DENKO, Tokio, Japan) standards (2 mg/mL) with molecular weights of 0.62 × 104, 1.00 × 104, 2.17 × 104, 4.88 × 104, 11.3 × 104, 20.0 × 104, 36.6 × 104, and 80.5 × 104 Da.
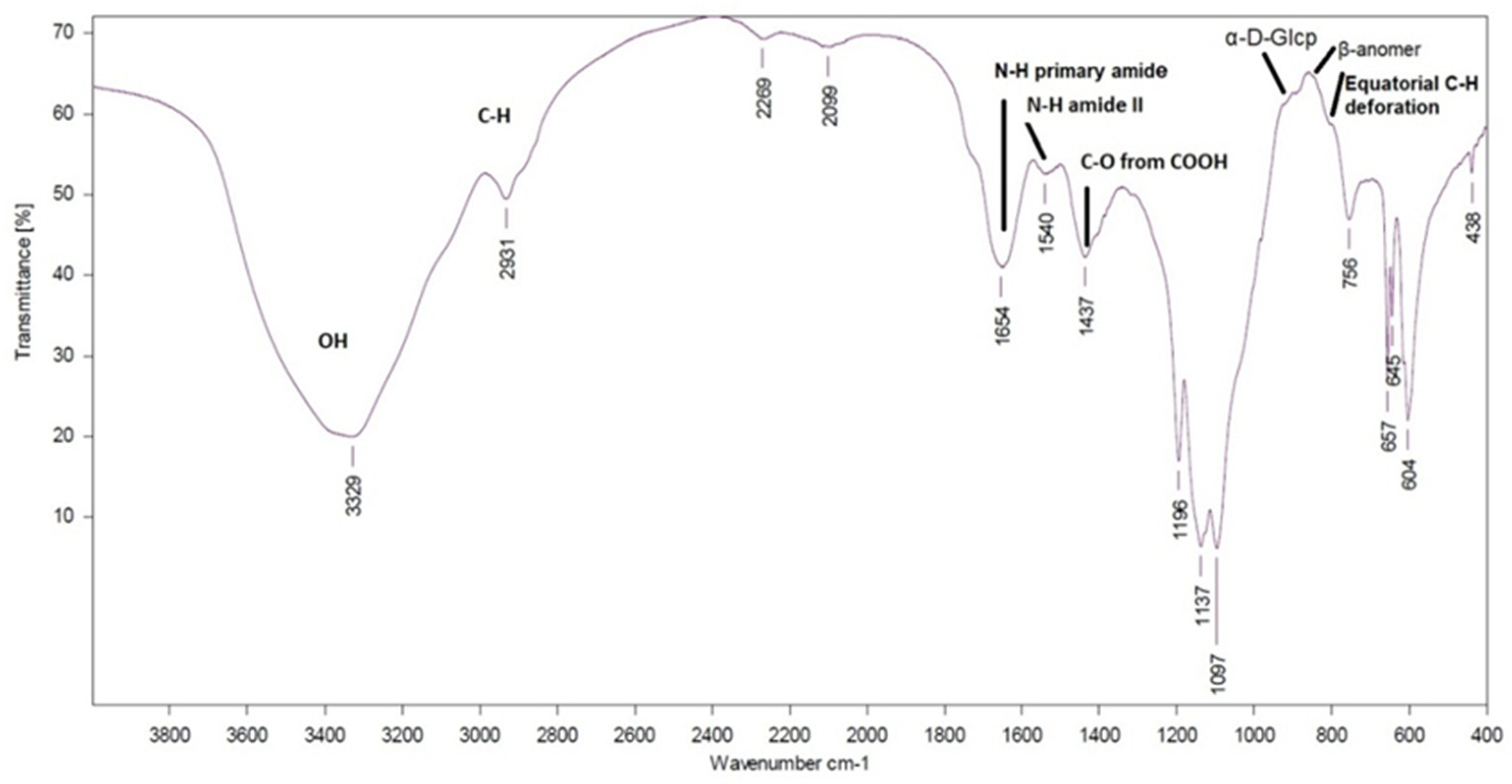
2.6.3. Infrared (IR) Spectra
The IR spectra of the EPS (2 mg) were collected on a Fourier transform infrared (FTIR) spectrophotometer VERTEX 70v (Bruker, Bremen, Germany) in KBr pellets. The spectra were recorded in the 4000–400 cm−1 range at 132 scans with a spectral resolution of 2 cm−1.
2.7. Bioactivity Assays
2.7.1. Determination of the In Vitro Antioxidant Activity (AOA) of Biomass Extracts
The ability of the mycelial biomass extracts to donate an electron and scavenge DPPH radicals was determined by the slightly modified method of Brand-Williams et al. [
51]. Freshly prepared 4 × 10
−4 M methanol solution of DPPH was mixed with the mycelium biomass extract at a ratio of 2:0.5 (
v/
v). The light absorption was measured at 517 nm at room temperature after 30 min incubation. The DPPH radical scavenging activity was presented as a function of the concentration of Trolox-Trolox equivalent antioxidant capacity (TEAC), and was defined as the concentration of Trolox having equivalent AOA expressed as the μM Trolox per g DW (μM TE/g DW).
The radical scavenging activity of the extracts against radical cation (ABTS•+) was estimated according to a previously reported procedure with some modifications [
52]. The results were expressed as TEAC values (μM TE/g DW).
The FRAP assay was carried out according to the procedure of Benzie and Strain [
53]. The FRAP reagent was prepared fresh daily and was warmed to 37 °C prior to use. The absorbance of the reaction mixture was recorded at 593 nm after incubation at 37 °C for 4 min. The results were expressed as μM TE/g DW.
CUPRAC assay was performed according to the method of Apak et al. [
54]. Amounts of 1 mL of CuCl
2 solution (1.0 × 10
−2 M), 1 mL of neocuproine methanolic solution (7.5 × 10
−3 M) and 1 mL NH
4OOCCH
3 buffer solution (pH 7.0) were added to a test tube and mixed; 0.1 mL of the mycelium biomass extract followed by 1 mL of water were added (total volume of 4.1 mL) and mixed well. Absorbance against a reagent blank was measured at 450 nm after 30 min. Trolox was used as standard and total antioxidant capacity of fungal biomass extracts was determined as μM TE/g DW.
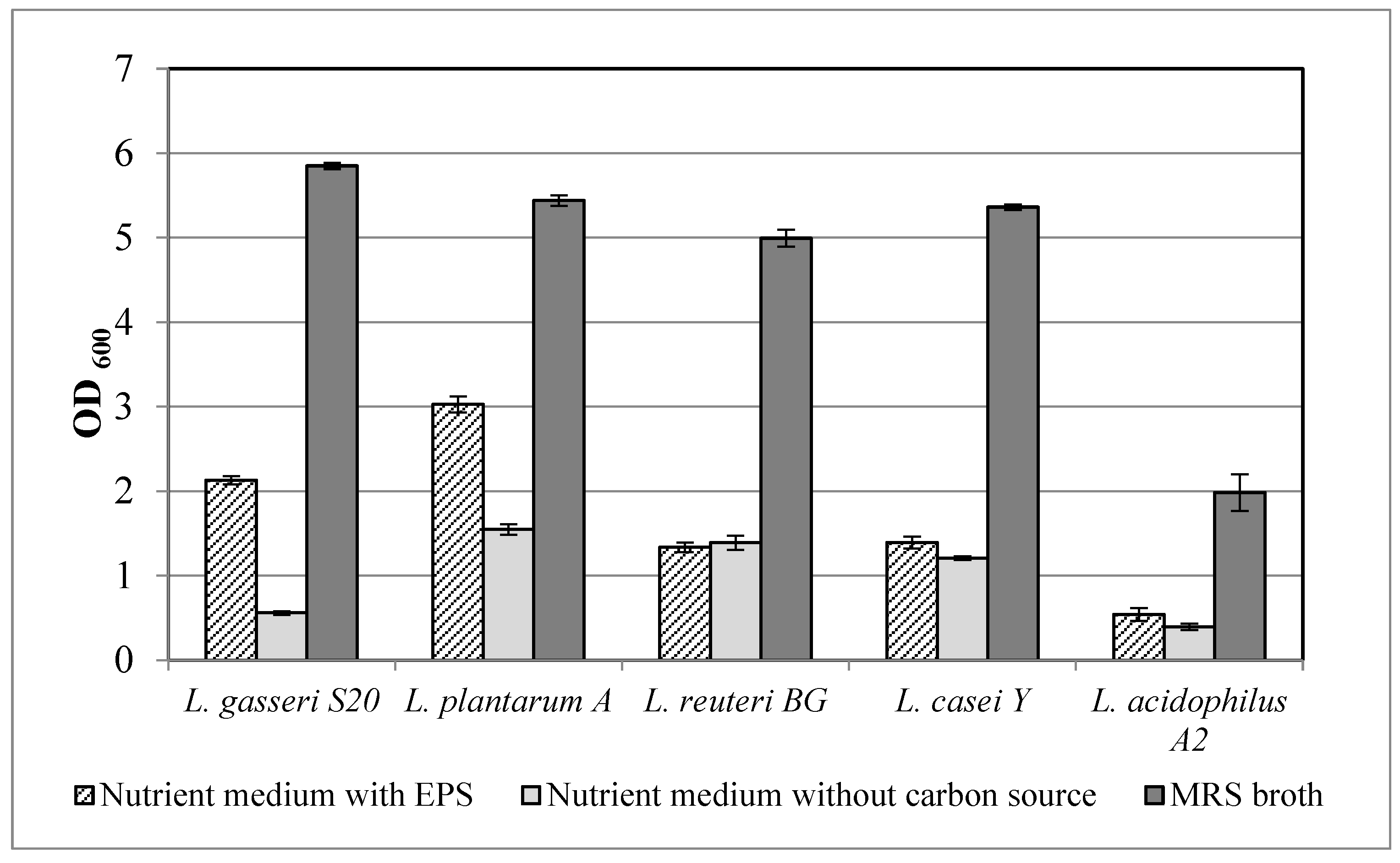
2.7.2. Prebiotic Activity of Crude Exopolysaccharides
The prebiotic activity of the EPS was determined using the following probiotic bacteria: Lactobacillus gasseri S20, Lactiplantibacillus plantarum A, Limosilactobacillus reuteri BG, Lacticaseibacillus casei Y and Lactobacillus acidophilus A2. The effect of EPS was investigated through the cultivation of the lactic acid bacteria in 96-deep-well plates containing 2 mL nutrient medium (in g/L: proteose peptone–10.0, meat extract–8.0, yeast extract–4.0, sodium acetate–5.0, triamonium citrate–2.0, magnesium sulfate–0.2, manganese sulfate–0.05 and dipotassium sulfate–2.0; Tween, 80–1.0 mL; pH 6.2) with 1.0 g/L EPS. MRS broth (Merck KGaA, Darmstadt, Germany) was used as a positive control and the above-mentioned medium without carbon source was used as negative control. Every well was inoculated with 1% inoculum with OD600 = 0.5 ÷ 0.6. The plates were incubated at 37 °C and the optical density was determined after 24 h.
2.7.3. In Vitro Determination of Anti-Inflammatory Potential of the Crude Exopolysaccharides
The HT-29 cells were cultured to monolayer in DMEM (Dulbecco’s modified Eagle’s medium, Gibco, UK), supplemented with 10% fetal bovine serum at 37 °C and 5% CO2. In about 90% of the cell monolayer, cells were passaged by incubation with 0.25% trypsin and 10 mM EDTA solution for 10 min at 37 °C. To determine the anti-inflammatory effect, eukaryotic cells were cultured in 48-well plates at a concentration of 2 × 105 cells/mL. The medium was changed every 2 days for a total of 14 days, supporting not only a monolayer formation (3–4 days), but also the maturation of cellular receptors. The resulting monolayer was washed twice with phosphate buffered saline (PBS) buffer. In each well, 500 µL DMEM containing 2 mg/mL crude EPS were added and incubated for 20 h at 37 °C. After that, the cell-free supernatant was used for determination of cytokine expression. IL-1β, IL-8, TGF-β were assessed using enzyme-linked immunosorbent assays according to the manufacturer’s instruction (Diaclone, Ann Arbor, MI, USA).
2.8. Statistical Analysis
All the experiments were conducted in triplicate and the values were expressed as mean ± SD. Statistical significance was detected by analysis of variance (ANOVA, Tukey’s test; value of p < 0.05 indicated statistical difference.


 ,
, 






{kind=link}
{kind=link}