
Adenylsuccinate Synthetase MoADE12 Plays Important Roles in the Development and Pathogenicity of the Rice Blast Fungus

Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains and Culture Conditions
2.2. Identification of the Gene with T-DNA Insertion by Whole-Genome Resequencing
2.3. Targeted Gene Replacement and Complementation for MoADE12
2.4. Pathogenicity Assays
2.5. Conidiation, Sexual Reproduction, and Analysis of Infection-Related Morphogenesis
2.6. Assays for Osmotic Stress, Cell Wall Integrity and Oxidative Stress
2.7. Transcriptomic Analysis
3. Results
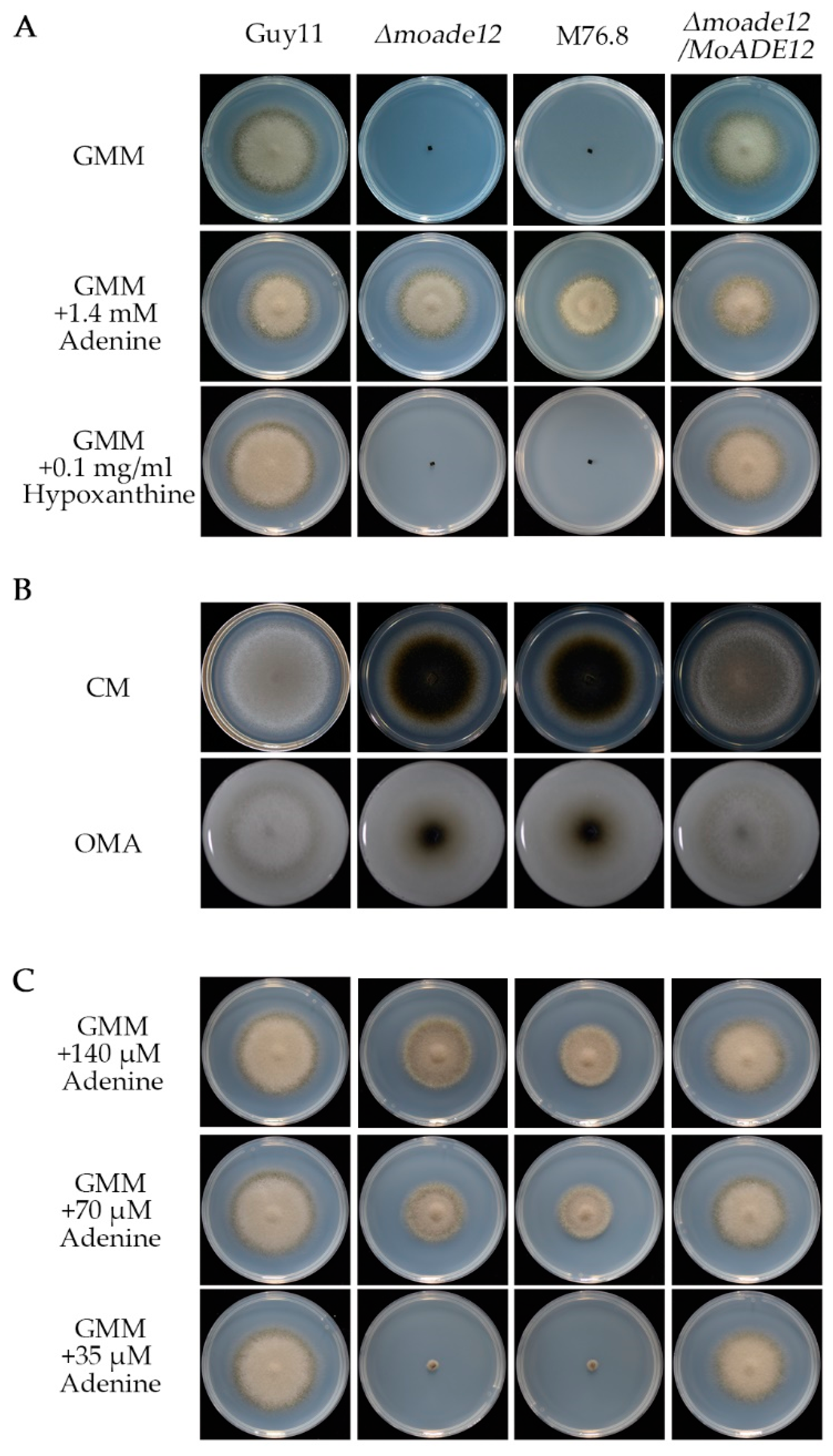
3.1. Identification of MoADE12 in M. oryzae
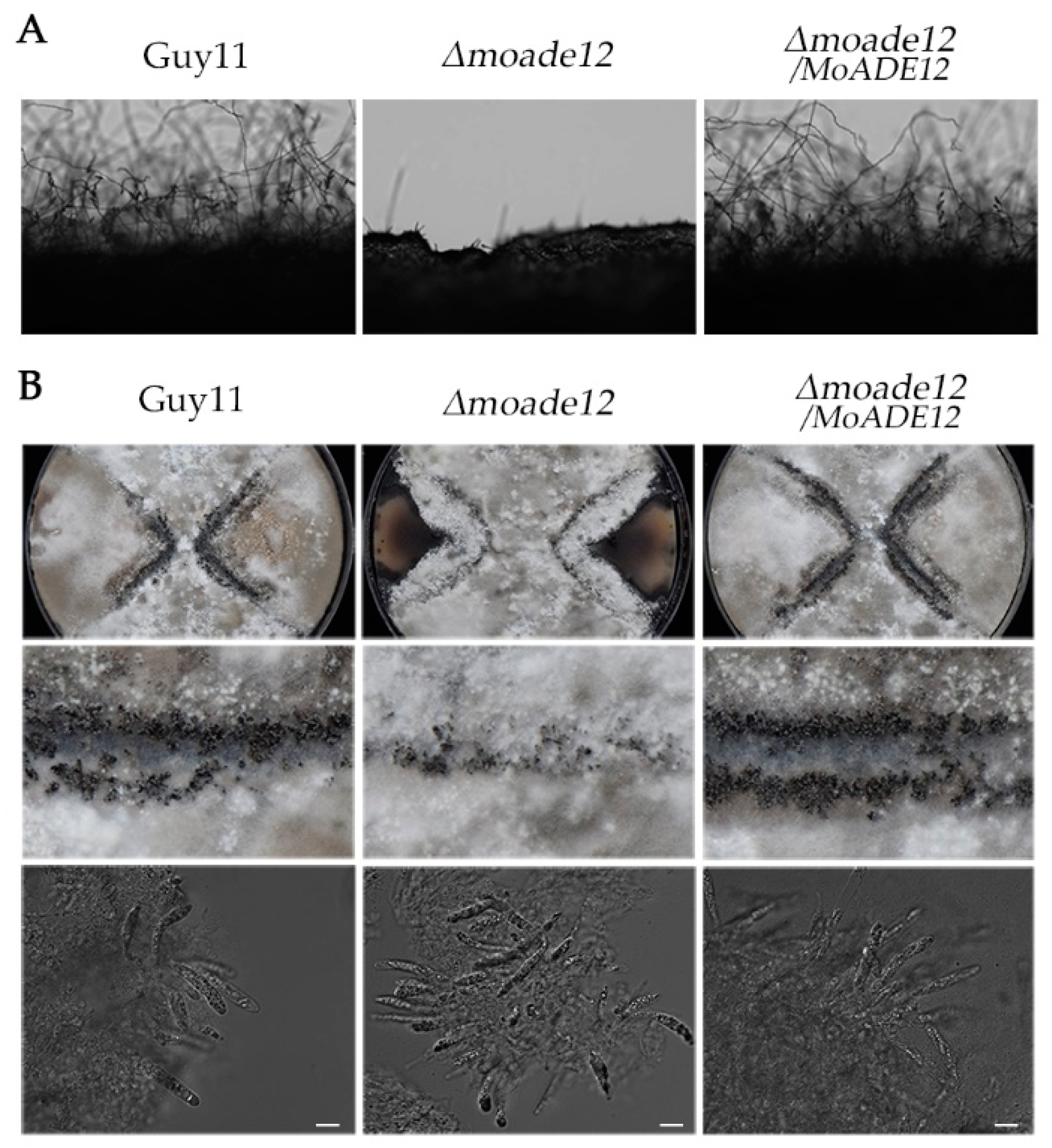
3.2. MoADE12 Is Involved in Asexual Conidiation and Sexual Reproduction
3.3. Deletion of MoADE12 Significantly Attenuates the Virulence of M. oryzae
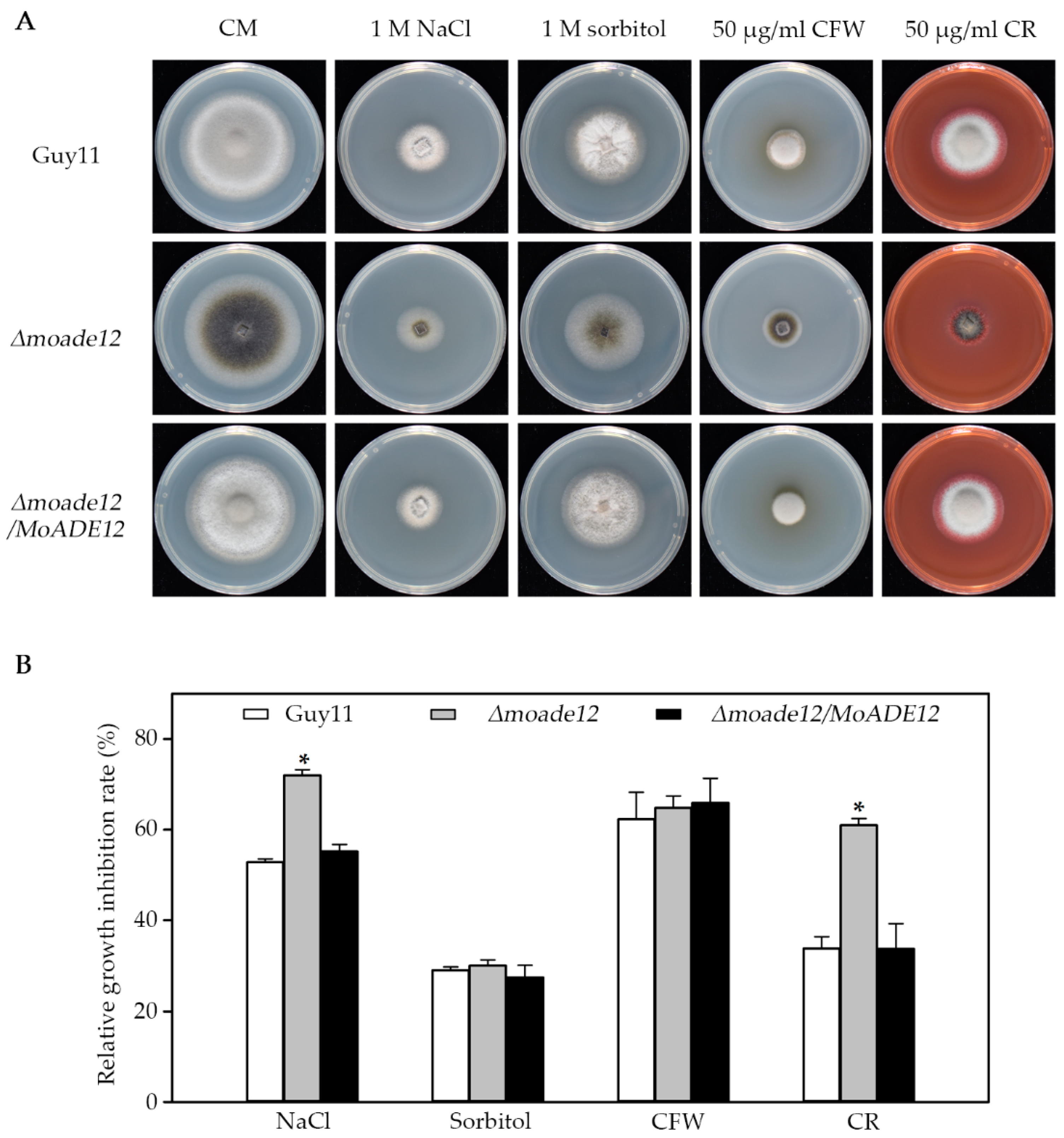
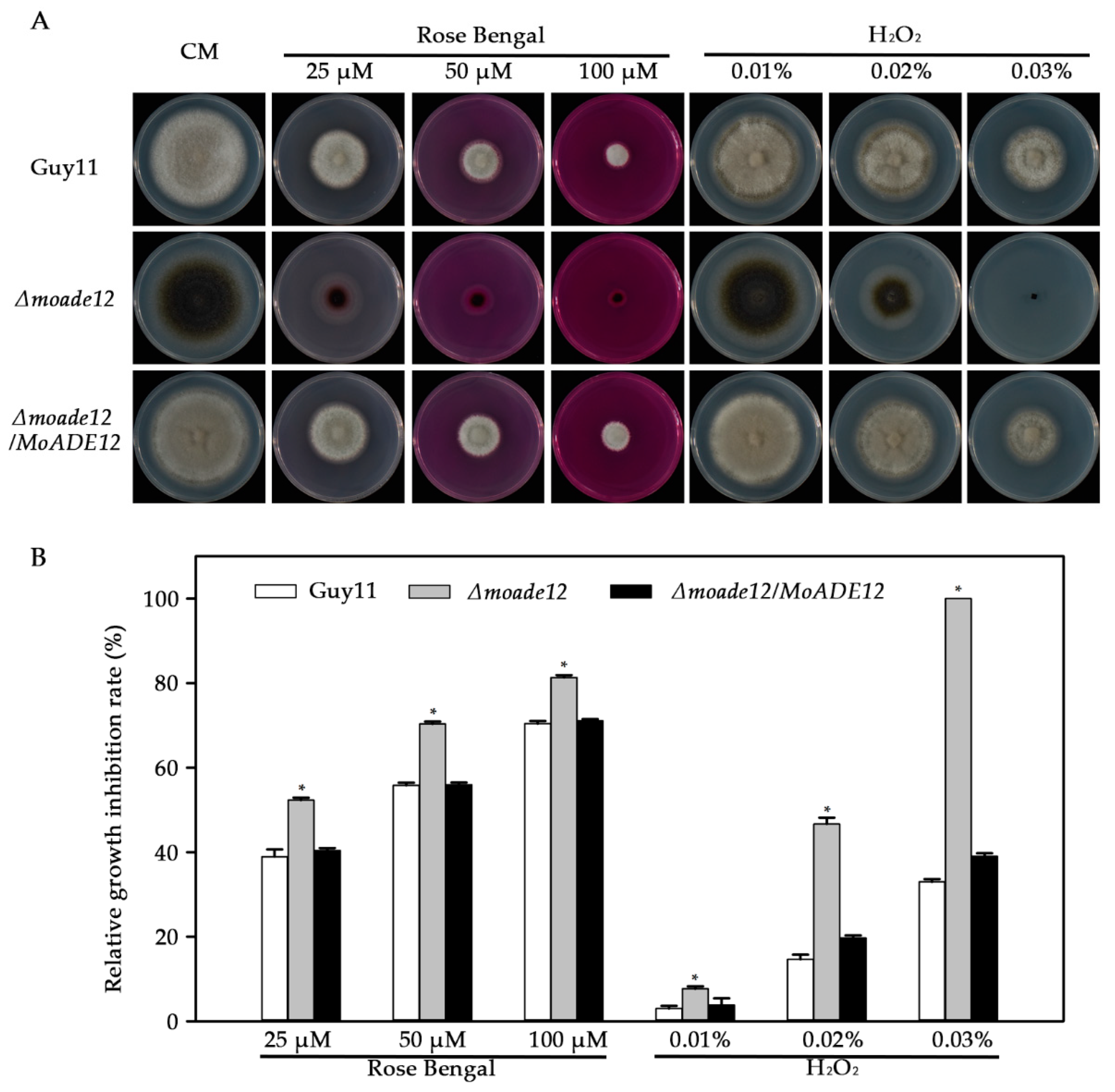
3.4. MoADE12 Is Required for Cell Wall Integrity, Osmotic Stress, and Oxidative Stress Resistance
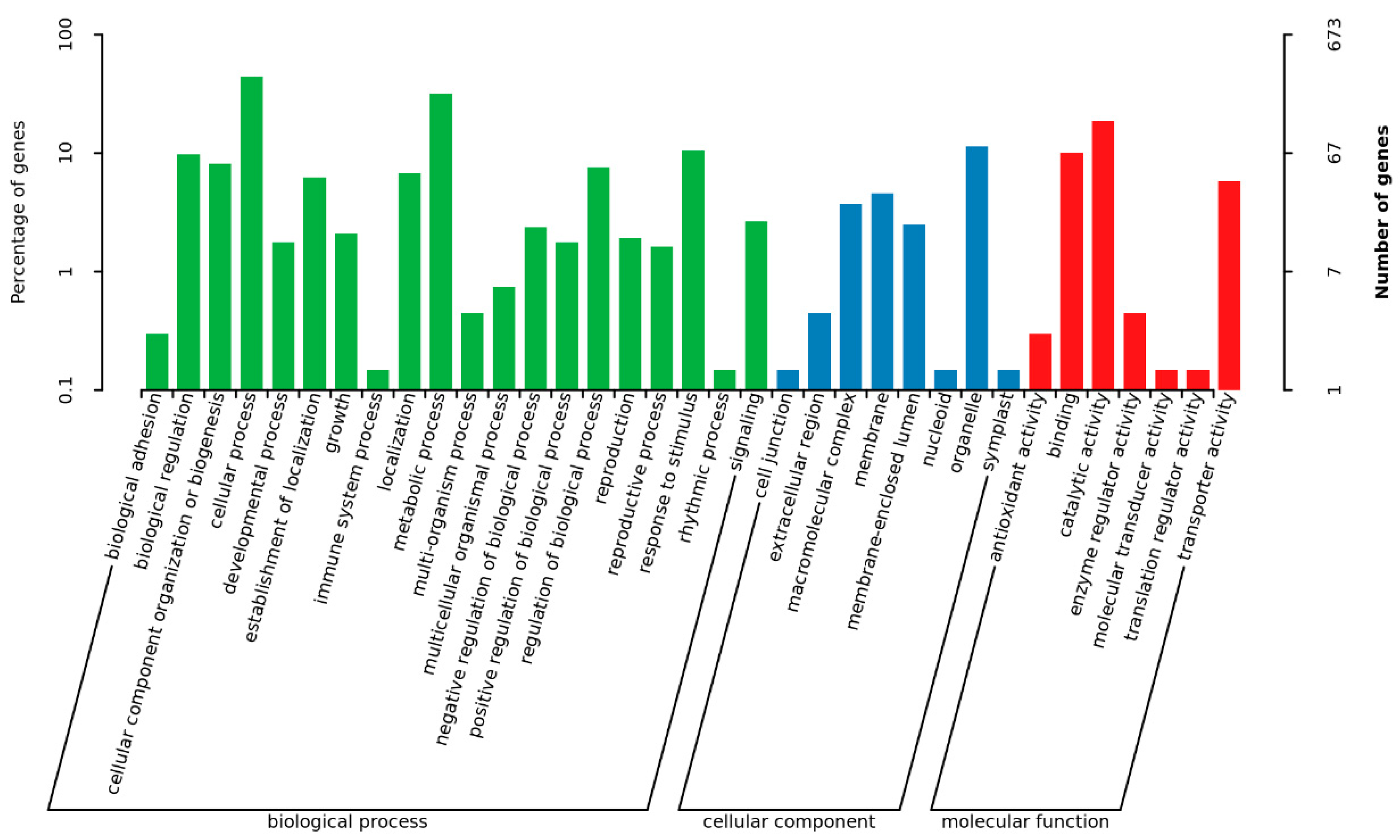
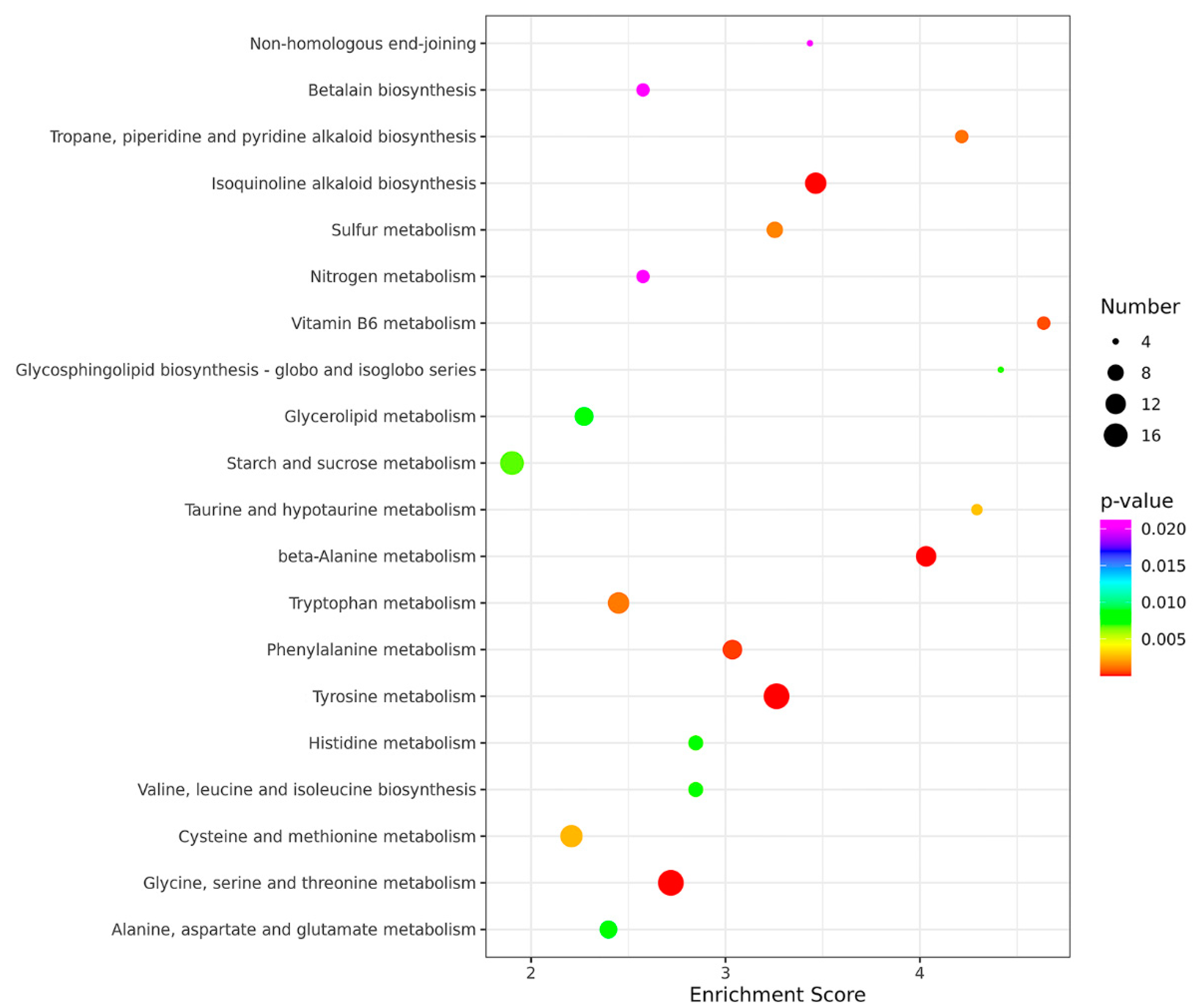
3.5. Transcriptional Analysis for Identification of Differentially Expressed Genes (DEGs) in a Comparison of the ∆moade12 and Wild-Type Strains
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marcel, S.; Sawers, R.; Oakeley, E.; Angliker, H.; Paszkowski, U. Tissue-adapted invasion strategies of the rice blast fungus Magnaporthe oryzae. Plant Cell 2010, 22, 3177–3187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, R.A.; Talbot, N.J. Under pressure: Investigating the biology of plant infection by Magnaporthe oryzae. Nat. Rev. Genet. 2009, 7, 185–195. [Google Scholar]
- Howard, R.J.; Ferrari, M.A.; Roach, D.H.; Money, N.P. Penetration of hard substrates by a fungus employing enormous turgor pressures. Proc. Natl. Acad. Sci. USA 1991, 88, 11281–11284. [Google Scholar] [CrossRef] [Green Version]
- De Jong, J.C.; McCormack, B.J.; Smirnoff, N.; Talbot, N.J. Glycerol generates turgor in rice blast. Nature 1997, 389, 244–245. [Google Scholar]
- Foster, A.J.; Littlejohn, G.R.; Soanes, D.M.; Talbot, N.J. Strategies for Nutrient Acquisition by Magnaporthe Oryzae during the Infection of Rice. In Host—Pathogen Interaction; Unden, G., Thines, E., Schüffler, A., Eds.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2016; pp. 93–108. [Google Scholar]
- Fernandez, J.; Wilson, R.A. Why no feeding frenzy? Mechanisms of nutrient acquisition and utilization during Infection by the rice blast fungus Magnaporthe oryzae. Mol. Plant Microbe Interact. 2012, 25, 1286–1293. [Google Scholar] [PubMed] [Green Version]
- Fernandez, J.; Wright, J.D.; Hartline, D.; Quispe, C.F.; Madayiputhiya, N.; Wilson, R.A. Principles of carbon catabolite repression in the rice blast fungus: Tps1, Nmr1-3, and a MATE-family pump regulate glucose metabolism during infection. PLoS Genet. 2012, 8, e1002673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tudzynski, B. Nitrogen regulation of fungal secondary metabolism in fungi. Front. Microbiol. 2014, 5, 656. [Google Scholar] [CrossRef] [Green Version]
- Wilson, R.A.; Fernandez, J.; Quispe, C.F.; Gradnigo, J.; Seng, A.; Moriyama, E.; Wright, J.D. Towards defining nutrient conditions encountered by the rice blast fungus during host infection. PLoS ONE 2012, 7, e47392. [Google Scholar] [CrossRef]
- Wilson, R.A.; Jenkinson, J.M.; Gibson, R.P.; Littlechild, J.A.; Wang, Z.-Y.; Talbot, N.J. Tps1 regulates the pentose phosphate pathway, nitrogen metabolism and fungal virulence. EMBO J. 2007, 26, 3673–3685. [Google Scholar] [PubMed] [Green Version]
- Fernandez, J.; Marroquin-Guzman, M.; Wilson, R.A. Mechanisms of nutrient acquisition and utilization during fungal Infections of Leaves. Annu. Rev. Phytopathol. 2014, 52, 155–174. [Google Scholar] [CrossRef]
- Solomon, P.S.; Tan, K.-C.; Oliver, R.P. The nutrient supply of pathogenic fungi; a fertile field for study. Mol. Plant Pathol. 2003, 4, 203–210. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, H.; Liang, S.; Ning, G.; Xu, N.; Lu, J.; Liu, X.; Lin, F. MoARG1, MoARG5,6 and MoARG7 involved in arginine biosynthesis are essential for growth, conidiogenesis, sexual reproduction, and pathogenicity in Magnaporthe oryzae. Microbiol. Res. 2015, 180, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zuo, R.; Zhu, Q.; Sun, Y.; Li, M.; Dong, Y.; Ru, Y.; Zhang, H.; Zheng, X.; Zhang, Z. MoLys2 is necessary for growth, conidiogenesis, lysine biosynthesis, and pathogenicity in Magnaporthe oryzae. Fungal Genet. Biol. 2014, 67, 51–57. [Google Scholar] [CrossRef] [PubMed]
- Saint-Macary, M.E.; Barbisan, C.; Gagey, M.J.; Frelin, O.; Beffa, R.; Lebrun, M.H.; Droux, M. Methionine biosynthesis is essential for infection in the rice blast fungus Magnaporthe oryzae. PLoS ONE 2015, 10, e0111108. [Google Scholar] [CrossRef]
- Tang, W.; Jiang, H.; Zheng, Q.; Chen, X.; Wang, R.; Yang, S.; Zhao, G.; Liu, J.; Norvienyeku, J.; Wang, Z. Isopropylmalate isomerase MoLeu1 orchestrates leucine biosynthesis, fungal development, and pathogenicity in Magnaporthe oryzae. Appl. Microbiol. Biotechnol. 2018, 103, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Rolfes, R. Regulation of purine nucleotide biosynthesis: In yeast and beyond. Biochem. Soc. Trans. 2006, 34, 786–790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, X.; Zhang, X.; Li, X.; Liu, M.; Liu, X.; Wang, X.; Zhang, H.; Zheng, X.; Zhang, Z. The atypical guanylate kinase MoGuk2 plays important roles in asexual/sexual development, conidial septation, and pathogenicity in the rice blast fungus. Front. Microbiol. 2017, 8, 2467. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, J.; Yang, K.T.; Cornwell, K.M.; Wright, J.D.; Wilson, R.A. Growth in rice cells requires de novo purine biosynthesis by the blast fungus Magnaporthe oryzae. Sci. Rep. 2013, 3, 2398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipps, G.; Krauss, G. Adenylosuccinate synthase from Saccharomyces cerevisiae: Homologous overexpression, purification and characterization of the recombinant protein. Biochem. J. 1999, 341, 537–543. [Google Scholar] [CrossRef]
- Dorfman, B.Z. The isolation of adenylosuccinate synthetase mutants in yeast by selection for constitutive behavior in pigmented strains. Genetics 1969, 61, 377–389. [Google Scholar] [CrossRef]
- Talbot, N.J.; Salch, Y.P.; Ma, M.; Hamer, J.E. Karyotypic variation within clonal lineages of the rice blast fungus, Magnaporthe grisea. Appl. Environ. Microbiol. 1993, 59, 585–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhang, Z.; Wang, Y.; Li, L.; Chai, R.; Mao, X.; Jiang, H.; Qiu, H.; Du, X.; Lin, F.; et al. PTS1 peroxisomal import pathway plays shared and distinct roles to PTS2 pathway in development and pathogenicity of Magnaporthe oryzae. PLoS ONE 2013, 8, e55554. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Wang, J.; Zhang, Z.; Wang, Y.; Liu, M.; Jiang, H.; Chai, R.; Mao, X.; Qiu, H.; Liu, F.; et al. MoPex19, which is essential for maintenance of peroxisomal structure and woronin bodies, is required for metabolism and development in the rice blast fungus. PLoS ONE 2014, 9, e85252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talbot, N.J.; Ebbole, D.J.; Hamer, J.E. Identification and characterization of MPG1, a gene involved in pathogenicity from the rice blast fungus Magnaporthe grisea. Plant Cell 1993, 5, 1575–1590. [Google Scholar] [PubMed] [Green Version]
- Dixon, K.P.; Xu, J.R.; Smirnoff, N.; Talbot, N.J. Independent signaling pathways regulate cellular turgor during hyperosmotic stress and appressorium-mediated plant infection by Magnaporthe grisea. Plant Cell 1999, 11, 2045–2058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koga, H.; Dohi, K.; Nakayachi, O.; Mori, M. A novel inoculation method of Magnaporthe grisea for cytological observation of the infection process using intact leaf sheaths of rice plants. Physiol. Mol. Plant Pathol. 2004, 64, 67–72. [Google Scholar] [CrossRef]
- Ram, A.F.J.; Klis, F.M. Identification of fungal cell wall mutants using susceptibility assays based on Calcofluor white and Congo red. Nat. Protoc. 2006, 1, 2253–2256. [Google Scholar] [CrossRef]
- Mentlak, T.A.; Kombrink, A.; Shinya, T.; Ryder, L.S.; Otomo, I.; Saitoh, H.; Terauchi, R.; Nishizawa, Y.; Shibuya, N.; Thomma, B.P.; et al. Effector-mediated suppression of chitin-triggered immunity by Magnaporthe oryzae is necessary for rice blast disease. Plant Cell 2012, 24, 322–335. [Google Scholar] [CrossRef] [Green Version]
- Sharpee, W.; Oh, Y.; Yi, M.; Franck, W.; Eyre, A.; Okagaki, L.H.; Valent, B.; Dean, R.A. Identification and characterization of suppressors of plant cell death (SPD) effectors from Magnaporthe oryzae. Mol. Plant Pathol. 2016, 18, 850–863. [Google Scholar] [CrossRef]
- Wu, J.; Kou, Y.; Bao, J.; Li, Y.; Tang, M.; Zhu, X.; Ponaya, A.; Xiao, G.; Li, J.; Li, C.; et al. Comparative genomics identifies the Magnaporthe oryzae avirulence effector AvrPi9 that triggers Pi9-mediated blast resistance in rice. New Phytol. 2015, 206, 1463–1475. [Google Scholar] [CrossRef] [PubMed]
- Beckerman, J.L.; Ebbole, D.J. MPG1, a gene encoding a fungal hydrophobin of Magnaporthe grisea, is involved in surface recognition. Mol. Plant Microbe Interact. 1996, 9, 450–456. [Google Scholar] [CrossRef]
- Skamnioti, P.; Gurr, S.J. Magnaporthe grisea cutinase2 mediates appressorium differentiation and host penetration and is required for full virulence. Plant Cell 2007, 19, 2674–2689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Tang, W.; Liu, K.; Huang, Q.; Zhang, X.; Yan, X.; Chen, Y.; Wang, J.; Qi, Z.; Wang, Z.; et al. Eight RGS and RGS-like proteins orchestrate growth, differentiation, and pathogenicity of Magnaporthe oryzae. PLoS Pathog. 2011, 7, e1002450. [Google Scholar] [CrossRef] [Green Version]
- Jacob, S.; Foster, A.J.; Yemelin, A.; Thines, E. Histidine kinases mediate differentiation, stress response, and pathogenicity in Magnaporthe oryzae. Microbiol. Open 2014, 3, 668–687. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Ahn, I.-P.; Rho, H.-S.; Lee, Y.-H. MHP1, a Magnaporthe grisea hydrophobin gene, is required for fungal development and plant colonization. Mol. Microbiol. 2005, 57, 1224–1237. [Google Scholar] [CrossRef] [PubMed]
- DeZwaan, T.M.; Carroll, A.M.; Valent, B.; Sweigard, J.A. Magnaporthe grisea Pth11p is a novel plasma membrane protein that mediates appressorium differentiation in response to inductive substrate cues. Plant Cell 1999, 11, 2013–2030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, H.; Huang, P.; Zhang, L.; Shi, Y.; Sun, D.; Yan, Y.; Liu, X.; Dong, B.; Chen, G.; Snyder, J.H.; et al. Characterization of 47 Cys2-His2 zinc finger proteins required for the development and pathogenicity of the rice blast fungus Magnaporthe oryzae. New Phytol. 2016, 211, 1035–1051. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Park, S.-Y.; Kim, K.S.; Rho, H.-S.; Chi, M.-H.; Choi, J.; Park, J.; Kong, S.; Park, J.; Goh, J.; et al. Homeobox transcription factors are required for conidiation and appressorium development in the rice blast fungus Magnaporthe oryzae. PLoS Genet. 2009, 5, e1000757. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Li, G.; Lin, C.; He, C. Conidiophore Stalk-less1 Encodes a Putative Zinc-Finger Protein Involved in the Early Stage of Conidiation and Mycelial Infection in Magnaporthe oryzae. Mol. Plant Microbe Interact. 2009, 22, 402–410. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.B.; Suresh, A.; Deng, Y.Z.; Naqvi, N.I. A multidrug resistance transporter in Magnaporthe is required for host penetration and for survival during oxidative stress. Plant Cell 2006, 18, 3686–3705. [Google Scholar] [CrossRef] [Green Version]
- Mosquera, G.; Giraldo, M.C.; Khang, C.H.; Coughlan, S.; Valent, B. Interaction transcriptome analysis identifies Magnaporthe oryzae BAS1-4 as biotrophy-associated secreted proteins in rice blast disease. Plant Cell 2009, 21, 1273–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egan, M.J.; Wang, Z.-Y.; Jones, M.A.; Smirnoff, N.; Talbot, N.J. Generation of reactive oxygen species by fungal NADPH oxidases is required for rice blast disease. Proc. Natl. Acad. Sci. USA 2007, 104, 11772–11777. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.-P.; Cao, H.; Zhang, L.; Huang, P.; Lin, F. Systematic analysis of Zn2Cys6 transcription factors required for development and pathogenicity by high-throughput gene knockout in the rice blast fungus. PLoS Pathog. 2014, 10, e1004432. [Google Scholar] [CrossRef]
- Farman, M.L. Meiotic deletion at the BUF1 locus of the fungus Magnaporthe grisea is controlled by interaction with the homologous chromosome. Genetics 2002, 160, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Turrà, D.; El Ghalid, M.; Rossi, F.; Di Pietro, A. Fungal pathogen uses sex pheromone receptor for chemotropic sensing of host plant signals. Nature 2015, 527, 521–524. [Google Scholar] [CrossRef] [PubMed]
- Sadat, A.; Han, J.-H.; Kim, S.; Lee, Y.-H.; Kim, K.S.; Choi, J. The membrane-bound protein, MoAfo1, is involved in sensing diverse signals from different surfaces in the rice blast fungus. Plant Pathol. J. 2021, 37, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhai, S.; Zhang, H.; Zuo, R.; Wang, J.; Guo, M.; Zheng, X.; Wang, P.; Zhang, Z. Shared and distinct functions of two Gti1/Pac2 family proteins in growth, morphogenesis and pathogenicity of Magnaporthe oryzae. Environ. Microbiol. 2014, 16, 788–801. [Google Scholar] [CrossRef]
- Deng, Y.Z.; Ramos-Pamplona, M.; Naqvi, N.I. Autophagy-assisted glycogen catabolism regulates asexual differentiation in Magnaporthe oryzae. Autophagy 2009, 5, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Liu, X.; Liu, M.; Wang, Y.; Zou, Y.; You, Y.; Yang, L.; Hu, J.; Zhang, H.; Zheng, X.; et al. Magnaporthe oryzae auxiliary activity protein MoAa91 functions as chitin-binding protein to induce appressorium formation on artificial inductive surfaces and suppress plant immunity. mBio 2020, 11, e03304–e03319. [Google Scholar] [CrossRef] [Green Version]
- Raman, V.; Simon, S.A.; Demirci, F.; Nakano, M.; Meyers, B.C.; Donofrio, N.M. Small RNA functions are required for growth and development of Magnaporthe oryzae. Mol. Plant Microbe Interact. 2017, 30, 517–530. [Google Scholar] [CrossRef] [Green Version]
- Stone, T.W.; Taylor, D.A.; Bloom, F.E. Cyclic AMP and cyclic GMP may mediate opposite neuronal responses in the rat cerebral cortex. Science 1975, 187, 845–847. [Google Scholar] [CrossRef] [PubMed]
- Bölker, M. Sex and crime: Heterotrimeric G proteins in fungal mating and pathogenesis. Fungal Genet. Biol. 1998, 25, 143–156. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Wright, S.J.; Krystofova, S.; Park, G.; Borkovich, K.A. Heterotrimeric G protein signaling in filamentous fungi. Annu. Rev. Microbiol. 2007, 61, 423–452. [Google Scholar] [CrossRef]
- Sato, S.; Soga, T.; Nishioka, T.; Tomita, M. Simultaneous determination of the main metabolites in rice leaves using capillary electrophoresis mass spectrometry and capillary electrophoresis diode array detection. Plant J. 2004, 40, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Pantazopoulou, A.; Diallinas, G. Fungal nucleobase transporters. FEMS Microbiol. Rev. 2007, 31, 657–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schuster, S.; Kenanov, D. Adenine and adenosine salvage pathways in erythrocytes and the role of S-adenosylhomocysteine hydrolase. FEBS J. 2005, 272, 5278–5290. [Google Scholar] [CrossRef]
- Mir, A.A.; Park, S.-Y.; Sadat, A.; Kim, S.; Choi, J.; Jeon, J.; Lee, Y.-H. Systematic characterization of the peroxidase gene family provides new insights into fungal pathogenicity in Magnaporthe oryzae. Sci. Rep. 2015, 5, 11831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanabe, S.; Ishii-Minami, N.; Saitoh, K.-I.; Otake, Y.; Kaku, H.; Shibuya, N.; Nishizawa, Y.; Minami, E. The role of catalase-peroxidase secreted by Magnaporthe oryzae during early infection of rice cells. Mol. Plant Microbe Interact. 2011, 24, 163–171. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Description | Log2-fold Change | Name | Reference |
|---|---|---|---|---|
| MGG_09134 | Hypothetical protein | −8.17 | MPG1 | [32] |
| MGG_09100 | Cutinase | −7.97 | CUT2 | [33] |
| MGG_11693 | Hypothetical protein | −4.74 | RGS7 | [34] |
| MGG_12530 | Histidine kinase G7 | −4.61 | HIK3 | [35] |
| MGG_10105 | Hypothetical protein | −4.19 | MHP1 | [36] |
| MGG_05871 | Hypothetical protein | −3.11 | PTH11 | [37] |
| MGG_14931 | Zinc finger protein 32 | −2.60 | VRF1 | [38] |
| MGG_12865 | Hypothetical protein | −2.45 | MoHOX7 | [39] |
| MGG_03977 | Hypothetical protein | −2.40 | COS1 | [40] |
| MGG_13762 | Multidrug resistance protein 3 | −2.38 | ABC3 | [41] |
| MGG_09693 | Hypothetical protein | −2.19 | BAS2 | [42] |
| MGG_00750 | Cytochrome b−245 heavy chain subunit beta | −1.99 | NOX1 | [43] |
| MGG_05659 | Fungal-specific Zn(2)-Cys(6) domain-containing transcription factor | −1.75 | CCA1 | [44] |
| MGG_07219 | Conidial yellow pigment biosynthesis polyketide synthase | −1.71 | ALB1 | [4] |
| MGG_02252 | Tetrahydroxynaphthalene reductase | −1.65 | BUF1 | [45] |
| MGG_10193 | Guanine nucleotide-binding protein subunit gamma | −1.64 | MGG1 | [46] |
| MGG_10422 | C6 transcription factor, putative | −1.64 | MoAFO1 | [47] |
| MGG_08850 | cAMP-independent regulatory protein pac2 | −1.58 | MoGTI1 | [48] |
| MGG_01819 | Glycogen phosphorylase | −1.57 | GPH1 | [49] |
| MGG_06069 | Endoglucanase | −1.53 | MoAA91 | [50] |
| MGG_01062 | Hypothetical protein | 1.67 | ATG8 | [49] |
| MGG_12655 | Hypothetical protein | 2.01 | AVRPi9 | [31] |
| MGG_01294 | Hypothetical protein | 2.09 | MoAGO3 | [51] |
| MGG_03468 | Intracellular hyphae protein 1 | 2.34 | SLP2 | [29] |
| MGG_12942 | Hypothetical protein | 2.50 | SPD2 | [30] |
| MGG_10097 | Intracellular hyphae protein 1 | 7.41 | SLP1 | [29] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Hao, Z.; Chai, R.; Qiu, H.; Wang, Y.; Wang, J.; Sun, G. Adenylsuccinate Synthetase MoADE12 Plays Important Roles in the Development and Pathogenicity of the Rice Blast Fungus. J. Fungi 2022, 8, 780. https://doi.org/10.3390/jof8080780
Zhang Z, Hao Z, Chai R, Qiu H, Wang Y, Wang J, Sun G. Adenylsuccinate Synthetase MoADE12 Plays Important Roles in the Development and Pathogenicity of the Rice Blast Fungus. Journal of Fungi. 2022; 8(8):780. https://doi.org/10.3390/jof8080780
Chicago/Turabian StyleZhang, Zhen, Zhongna Hao, Rongyao Chai, Haiping Qiu, Yanli Wang, Jiaoyu Wang, and Guochang Sun. 2022. "Adenylsuccinate Synthetase MoADE12 Plays Important Roles in the Development and Pathogenicity of the Rice Blast Fungus" Journal of Fungi 8, no. 8: 780. https://doi.org/10.3390/jof8080780
APA StyleZhang, Z., Hao, Z., Chai, R., Qiu, H., Wang, Y., Wang, J., & Sun, G. (2022). Adenylsuccinate Synthetase MoADE12 Plays Important Roles in the Development and Pathogenicity of the Rice Blast Fungus. Journal of Fungi, 8(8), 780. https://doi.org/10.3390/jof8080780