
Fungal Pathogens Associated with Crown and Root Rot in Wheat-Growing Areas of Northern Kyrgyzstan

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
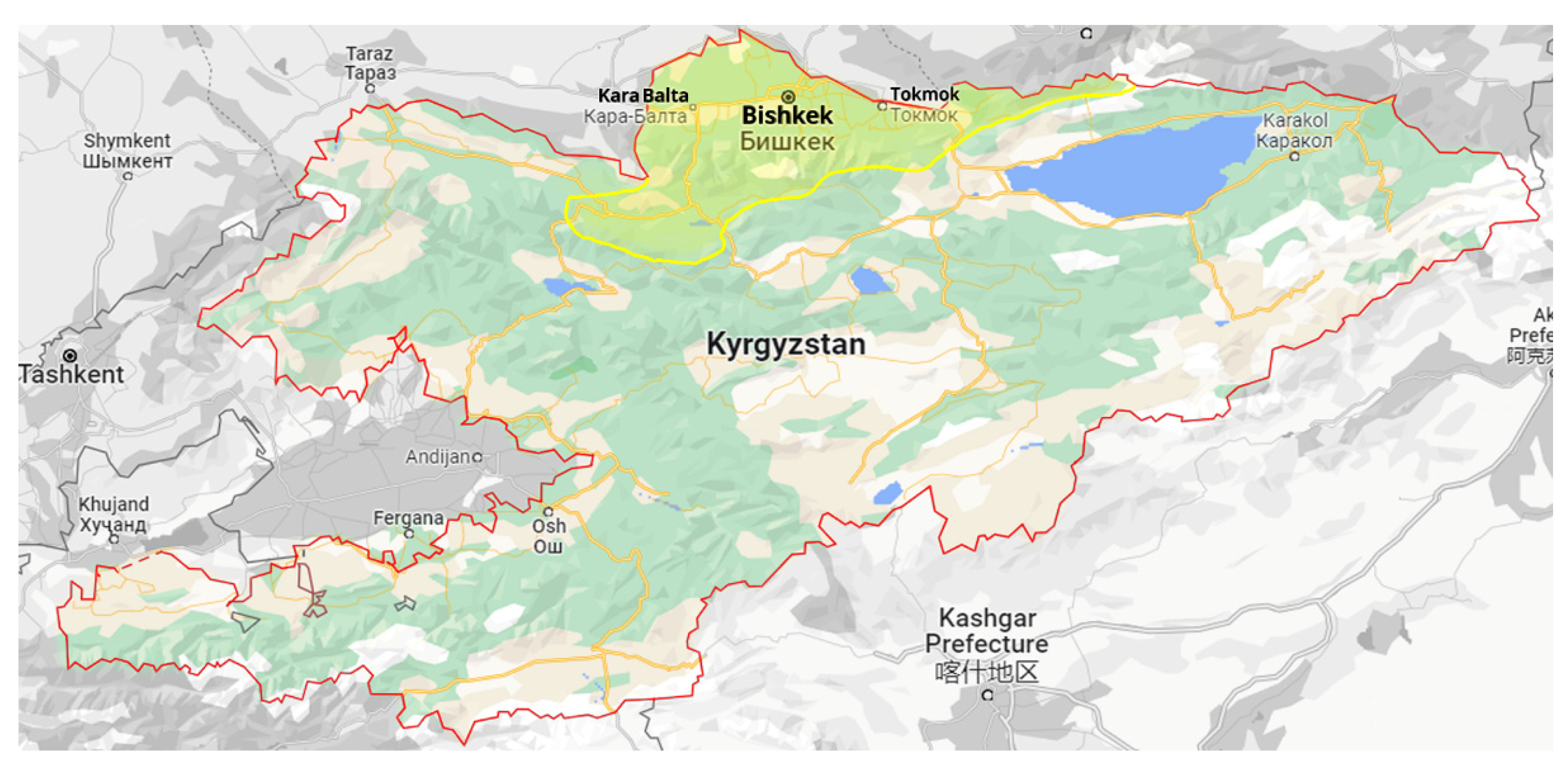
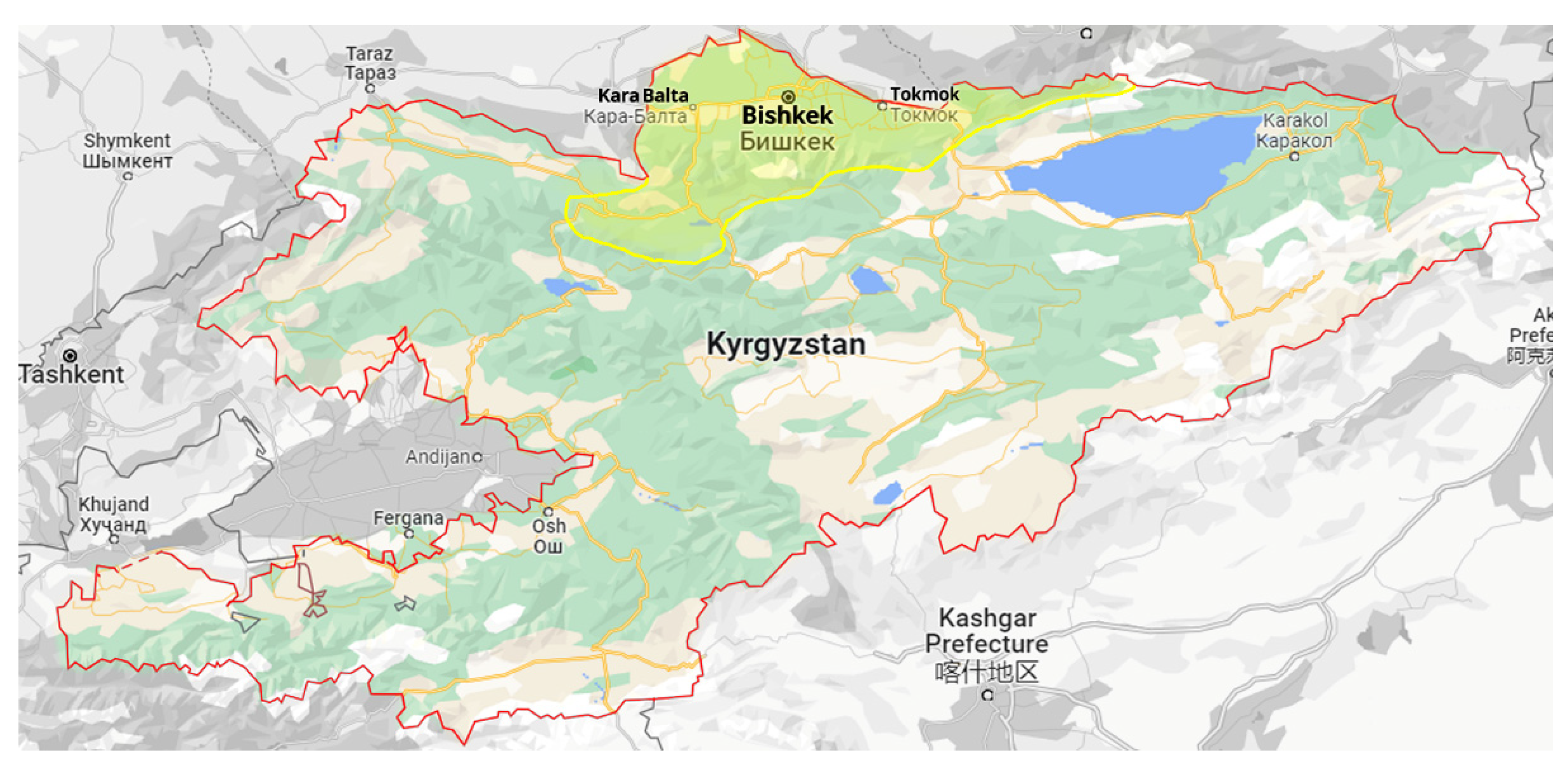
2.1. Protocol for Surveys
2.2. Culture Isolation, Preservation, and Morphological Identification
2.3. Molecular Identification
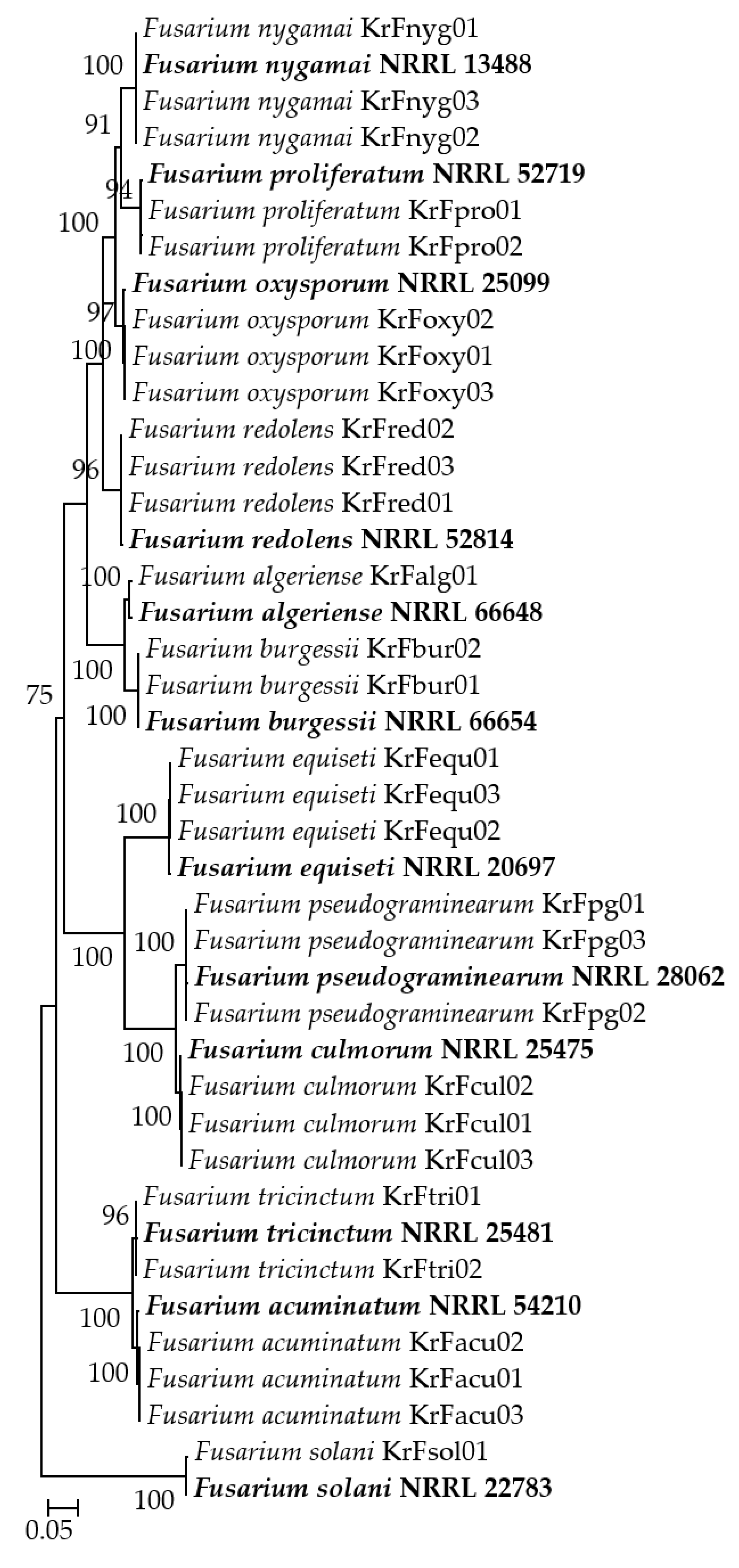
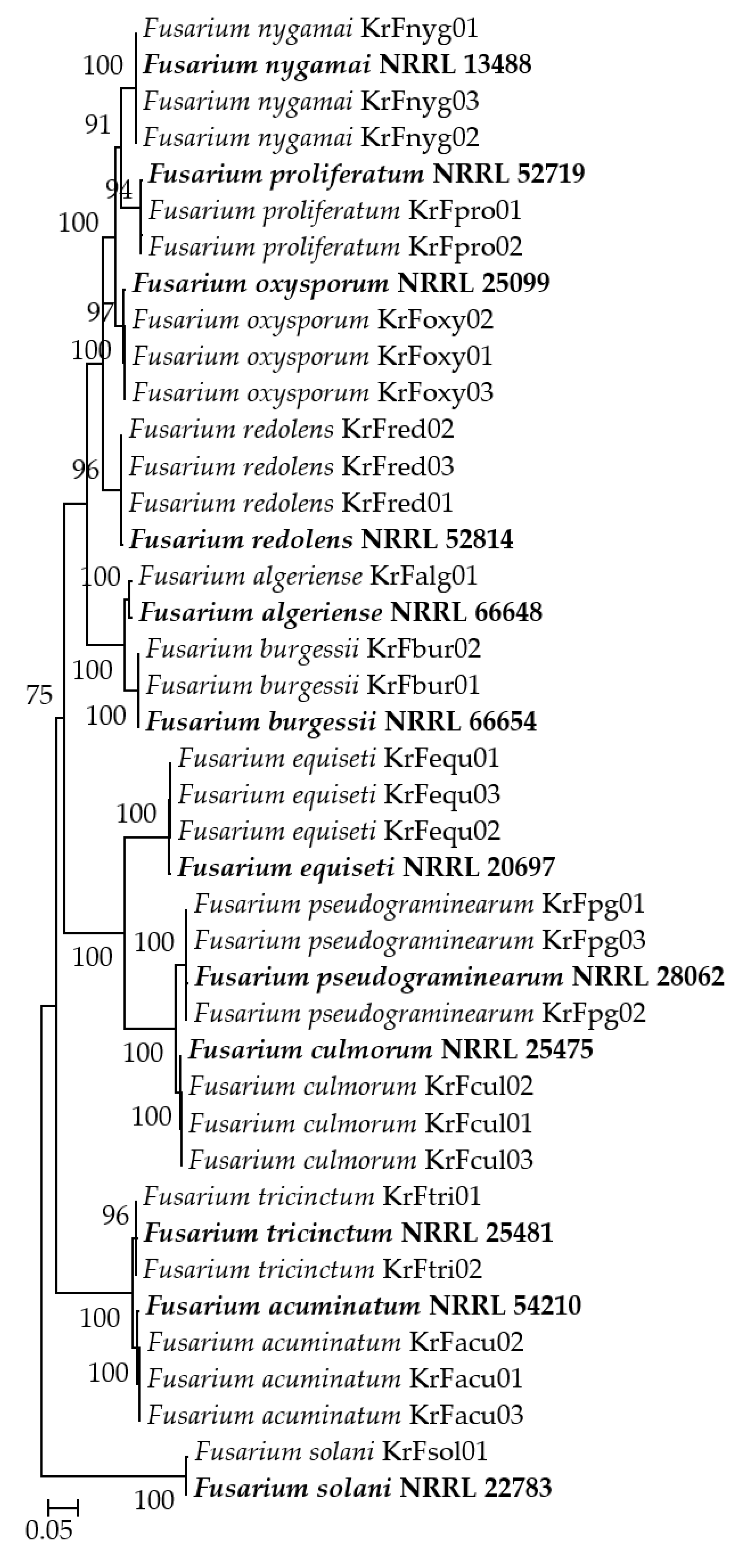
2.4. Phylogenetic Analysis
2.5. Fungal Species Frequency in Isolation and Incidence in the Fields
2.6. Pathogenicity Tests
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- FAOSTAT. Food and Agriculture Organization Statistical Database. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 21 September 2022).
- Shiferaw, B.; Smale, M.; Braun, H.J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Secur. 2013, 5, 291–317. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Fang, G.; Chen, Y.; Duan, W.; Mukanov, Y. Agricultural water demands in Central Asia under 1.5 C and 2.0 C global warming. Agric. Water Manag. 2020, 231, 106020. [Google Scholar] [CrossRef]
- Park, S.; Lim, C.H.; Kim, S.J.; Isaev, E.; Choi, S.E.; Lee, S.D.; Lee, W.K. Assessing climate change impact on cropland suitability in Kyrgyzstan: Where are potential high-quality cropland and the way to the future. Agronomy 2021, 11, 1490. [Google Scholar] [CrossRef]
- Ziyaev, Z.M.; Sharma, R.C.; Nazari, K.; Morgounov, A.I.; Amanov, A.A.; Ziyadullaev, Z.F.; Alikulov, S.M. Improving wheat stripe rust resistance in Central Asia and the Caucasus. Euphytica 2011, 179, 197–207. [Google Scholar] [CrossRef]
- Dzhunusova, M.; Yahyaoui, A.; Morgounov, A.I.; Jyldyz, E. Resistance of international winter wheat germplasm to yellow rust. In Proceedings of the Abstracts of the 3rd Regional Yellow Rust Conference for Central and West Asia and North Africa, Tashkent, Uzbekistan, 8–11 June 2006; ICARDA: Aleppo, Syria, 2006; p. 38. [Google Scholar]
- Kolmer, J.A.; Ordoñez, M.E. Genetic differentiation of Puccinia triticina populations in Central Asia and the Caucasus. Phytopathology 2007, 97, 1141–1149. [Google Scholar] [CrossRef] [Green Version]
- Garadagi, S.M. Resistance to yellow rust in some wheat varieties. Azarb Elmi-Tadg. Akincil Inst. Asar. Temat Macmuasi. 1976, 16, 117–119. (In Russian) [Google Scholar]
- Poole, G.J.; Harries, M.; Hüberli, D.; Miyan, S.; MacLeod, W.J.; Lawes, R.; McKay, A. Predicting cereal root disease in Western Australia using soil DNA and environmental parameters. Phytopathology 2015, 105, 1069–1179. [Google Scholar] [CrossRef] [Green Version]
- Bockus, W.W.; Bowden, R.L.; Hunger, R.M.; Morrill, W.L.; Murray, T.D.; Smiley, R.W. Compendium of Wheat Diseases and Pests, 3rd ed.; American Phytopathological Society: St. Paul, MN, USA, 2010. [Google Scholar]
- Xu, F.; Yang, G.; Wang, J.; Song, Y.; Liu, L.; Zhao, K.; Li, Y.; Han, Z. Spatial distribution of root and crown rot fungi associated with winter wheat in the North China Plain and its relationship with climate variables. Front. Microbiol. 2018, 9, 1054. [Google Scholar] [CrossRef]
- Smiley, R.W.; Gourlie, J.A.; Easley, S.A.; Patterson, L.M. Pathogenicity of fungi associated with the wheat crown rot complex in Oregon and Washington. Plant Dis. 2005, 89, 949–957. [Google Scholar] [CrossRef] [Green Version]
- Özer, G.; Paulitz, T.C.; İmren, M.; Alkan, M.; Muminjanov, H.; Dababat, A.A. Identity and pathogenicity of fungi associated with crown and root rot of dryland winter wheat in Azerbaijan. Plant Dis. 2020, 104, 2149–2157. [Google Scholar] [CrossRef]
- Moya-Elizondo, E.A.; Rew, L.J.; Jacobsen, B.J.; Hogg, A.C.; Dyer, A.T. Distribution and prevalence of Fusarium crown rot and common root rot pathogens of wheat in Montana. Plant Dis. 2011, 95, 1099–1108. [Google Scholar] [CrossRef]
- Moya-Elizondo, E.; Arismendi, N.; Castro, M.P.; Doussoulin, H. Distribution and prevalence of crown rot pathogens affecting wheat crops in southern Chile. Chil. J. Agric. Res. 2015, 75, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Alahmad, S.; Simpfendorfer, S.; Bentley, A.R. Crown rot of wheat in Australia: Fusarium pseudograminearum taxonomy, population biology and disease management. Australas. Plant Pathol. 2018, 47, 285–299. [Google Scholar] [CrossRef]
- Paulitz, T.C.; Smiley, R.W.; Cook, R.J. Insights into the prevalence and management of soilborne cereal pathogens under direct seeding in the Pacific Northwest, USA. Can. J. Plant Pathol. 2002, 24, 416–428. [Google Scholar] [CrossRef]
- Tunali, B.; Nicol, J.M.; Hodson, D.; Uçkun, Z.; Büyük, O.; Erdurmuş, D. Root and crown rot fungi associated with spring, facultative, and winter wheat in Turkey. Plant Dis. 2008, 92, 1299–1306. [Google Scholar] [CrossRef] [Green Version]
- Shikur Gebremariam, E.; Sharma-Poudyal, D.; Paulitz, T.C. Identity and pathogenicity of Fusarium species associated with crown rot on wheat (Triticum spp.) in Turkey. Eur. J. Plant Pathol. 2018, 150, 387–399. [Google Scholar] [CrossRef]
- Zhou, H.; He, X.; Wang, S.; Ma, Q.; Sun, B.; Ding, S.; Chen, L.; Zhang, M.; Li, H. Diversity of the Fusarium pathogens associated with crown rot in the Huanghuai wheat-growing region of China. Environ. Microbiol. 2019, 21, 2740–2754. [Google Scholar] [CrossRef]
- Bozoğlu, T.; Derviş, S.; Imren, M.; Amer, M.; Özdemir, F.; Paulitz, T.C.; Morgounov, A.; Dababat, A.A.; Özer, G. Fungal Pathogens Associated with Crown and Root Rot of Wheat in Central, Eastern, and Southeastern Kazakhstan. J. Fungi 2022, 8, 417. [Google Scholar] [CrossRef]
- Dyer, A.T.; Johnston, R.H.; Hogg, A.C.; Johnston, J.A. Comparison of pathogenicity of the Fusarium crown rot (FCR) complex (F. culmorum, F. pseudograminearum and F. graminearum) on hard red spring and durum wheat. Eur. J. Plant Pathol. 2009, 125, 387–395. [Google Scholar] [CrossRef]
- Cook, R.J. Fusarium root, crown, and foot rots and associated seedling diseases. In Compendium of Wheat Diseases and Pests, 3rd ed.; Bockus, W.W., Bowden, R.L., Hunger, R.M., Morrill, W.L., Murray, T.D., Smiley, R.W., Eds.; APS Press: St. Paul, MN, USA, 2010; pp. 37–39. [Google Scholar]
- Fernandez, M.R.; Conner, R.L. Root and crown rot of wheat. Prairie Soils Crops J. 2011, 4, 151–157. Available online: http://www.prairiesoilsandcrops.ca (accessed on 21 September 2022).
- Kumar, J.; Schäfer, P.; Hückelhoven, R.; Langen, G.; Baltruschat, H.; Stein, E. Bipolaris sorokiniana, a cereal pathogen of global concern: Cytological and molecular approaches towards better control. Mol. Plant Pathol. 2002, 3, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Al-Sadi, A.M. Bipolaris sorokiniana-induced black point, common root rot, and spot blotch diseases of wheat: A review. Front. Cell. Infect. Microbiol. 2021, 11, 584899. [Google Scholar] [CrossRef] [PubMed]
- Acharya, K.; Dutta, A.K.; Pradhan, P. Bipolaris sorokiniana (Sacc.) Shoem.: The most destructive wheat fungal pathogen in the warmer areas. Aust. J. Crop Sci. 2011, 5, 1064–1071. [Google Scholar] [CrossRef]
- Akinsanmi, O.A.; Mitter, V.; Simpfendorfer, S.; Backhouse, D.; Chakraborty, S. Identity and pathogenicity of Fusarium spp. isolated from wheat fields in Queensland and northern New South Wales. Aust. J. Agric. Res. 2004, 55, 97–107. [Google Scholar] [CrossRef]
- Anderson, W.K.; Garlinge, J.R. The Wheat Book: Principles and Practice. Agric. West. Aust. Bull. 2000, 4443, 217–219. [Google Scholar]
- Scherm, B.; Balmas, V.; Spanu, F.; Pani, G.; Delogu, G.; Pasquali, M.; Migheli, Q. Fusarium culmorum: Causal agent of foot and root rot and head blight on wheat. Mol. Plant Pathol. 2013, 14, 323–341. [Google Scholar] [CrossRef]
- Karlsson, I.; Persson, P.; Friberg, H. Fusarium head blight from a microbiome Perspective. Front. Microbiol. 2021, 12, 628373. [Google Scholar] [CrossRef]
- Pérez-Méndez, N.; Miguel-Rojas, C.; Jimenez-Berni, J.A.; Gomez-Candon, D.; Pérez-de-Luque, A.; Fereres, E.; Catala-Forner, M.; Villegas, D.; Sillero, J.C. Plant breeding and management strategies to minimize the impact of water scarcity and biotic stress in cereal crops under Mediterranean conditions. Agronomy 2022, 12, 75. [Google Scholar] [CrossRef]
- Leslie, J.F.; Summerell, B.A. (Eds.) The Fusarium Laboratory Manual; Blackwell: Ames, IO, USA, 2006. [Google Scholar]
- Laurence, M.H.; Summerell, B.A.; Burgess, L.W.; Liew, E.C.Y. Fusarium burgessii sp. nov. representing a novel lineage in the genus Fusarium. Fungal Divers. 2011, 49, 101–112. [Google Scholar] [CrossRef]
- Laraba, I.; Keddad, A.; Boureghda, H.; Abdallah, N.; Vaughan, M.M.; Proctor, R.H.; Busman, M.; O’Donnell, K. Fusarium algeriense, sp. nov., a novel toxigenic crown rot pathogen of durum wheat from Algeria is nested in the Fusarium burgessii species complex. Mycologia 2017, 109, 935–950. [Google Scholar] [CrossRef]
- Sivanesan, A. Graminicolous species of Bipolaris, Curvularia, Drechslera, Exserohilum and their teleomorphs. Mycologia 1987, 158, 1–126. [Google Scholar]
- Holliday, P.; Punithalingam, E. Macrophomina phaseolina. C.M.I. Descr. Pathog. Fungi Bact. 1970, 275, 1–2. [Google Scholar]
- Glynn, N.C.; Hare, M.C.; Parry, D.W.; Edwards, S.G. Phylogenetic analysis of EF-1 alpha gene sequences from isolates of Microdochium nivale leads to elevation of varieties majus and nivale to species status. Mycol. Res. 2005, 109, 872–880. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols, Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- O’Donnell, K.; Cigelnik, E.; Nirenberg, H.I. Molecular systematics and phylogeography of the Gibberella fujikuroi species complex. Mycologia 1998, 90, 465–493. [Google Scholar] [CrossRef]
- Reeb, V.; Lutzoni, F.; Roux, C. Contribution of RPB2 to multilocus phylogenetic studies of the euascomycetes (Pezizomycotina, Fungi) with special emphasis on the lichen-forming Acarosporaceae and evolution of polyspory. Mol. Phylogenet. Evol. 2004, 32, 1036–1060. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQTREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Hoang, D.T.; Chernomor, O.; von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Özer, G.; İmren, M.; Paulitz, T.C.; Bayraktar, H.; Muminjanov, H.; Dababat, A.A. First report of crown rot caused by Fusarium algeriense on wheat in Azerbaijan. Plant Dis. 2020, 104, 2149–2157. [Google Scholar] [CrossRef] [PubMed]
- Duczek, L.J.; Verma, P.R.; Spurr, D.T. Effect of inoculum density of Cochliobolus sativus on common root rot of wheat and barley. Can. J. Plant Pathol. 1985, 7, 382–386. [Google Scholar] [CrossRef]
- Demirci, E. Some hosts of Macrophomina phaseolina (Tassi) Goid. in Erzincan province. Atatürk Üniv. Ziraat Fak. Derg. 1997, 28, 280–284. [Google Scholar]
- Wildermuth, G.B.; McNamara, R.B. Testing wheat seedlings for resistance to crown rot caused by Fusarium graminearum Group 1. Plant Dis. 1994, 78, 949–953. [Google Scholar] [CrossRef]
- Özer, G.; Imren, M.; Alkan, M.; Paulitz, T.C.; Bayraktar, H.; Palacıoğlu, G.; Mehdiyev, I.; Muminjanov, H.; Dababat, A.A. Molecular and pathogenic characterization of Cochliobolus anamorphs associated with common root rot of wheat in Azerbaijan. Phytopathol. Mediterr. 2020, 59, 147–158. [Google Scholar] [CrossRef]
- Hill, J.P.; Fernandez, J.A.; McShane, M.S. Fungi associated with common root rot of winter wheat in Colorado and Wyoming. Plant Dis. 1983, 67, 795–797. [Google Scholar] [CrossRef]
- Gonzales, M.S.; Trevathan, L.E. Identity and pathogenicity of fungi associated with root and crown rot of soft red winter wheat grown on the upper coastal plain land resource area of Mississippi. J. Phytopathol. 2000, 148, 77–85. [Google Scholar] [CrossRef]
- Fernandez, M.R.; Fox, S.L.; Hucl, P.; Singh, A.K.; Stevenson, F.C. Root rot severity and fungal populations in spring common, durum and spelt wheat, and Kamut grown under organic management in western Canada. Can. J. Plant Sci. 2014, 94, 937–946. [Google Scholar] [CrossRef]
- Shrestha, S.; Poudel, R.S.; Zhong, S. Identification of Fungal Species Associated with Crown and Root Rots of Wheat and Evaluation of Plant Reactions to the Pathogens in North Dakota. Plant Dis. 2021, 105, 3564–3572. [Google Scholar] [CrossRef]
- Cook, R.J. Fusarium Diseases of Wheat and Other Small Grains in North America; Nelson, P.E., Tousson, T.A., Cook, R.J., Eds.; Penn State University Press: University Park, PA, USA, 1981; pp. 39–52. [Google Scholar]
- Smiley, R.W.; Patterson, L.M. Pathogenic fungi associated with Fusarium foot rot of winter wheat in the semiarid Pacific Northwest. Plant Dis. 1996, 80, 944–949. [Google Scholar] [CrossRef]
- Pettitt, T.; Xu, X.; Parry, D. Association of Fusarium species in the wheat stem rot complex. Eur. J. Plant Pathol. 2003, 109, 769–774. [Google Scholar] [CrossRef]
- Backhouse, D.; Abubakar, A.A.; Burgess, L.W.; Dennisc, J.I.; Hollaway, G.J.; Wildermuth, G.B.; Henry, F.J. Survey of Fusarium species associated with crown rot of wheat and barley in eastern Australia. Australas. Plant Pathol. 2004, 33, 255–261. [Google Scholar] [CrossRef] [Green Version]
- Abdallah-Nekache, N.; Laraba, I.; Ducos, C. Occurrence of Fusarium head blight and Fusarium crown rot in Algerian wheat: Identification of associated species and assessment of aggressiveness. Eur. J. Plant Pathol. 2019, 154, 499–512. [Google Scholar] [CrossRef]
- Chehri, K.; Salleh, B.; Yli-mattila, T.; Soleimani, M.J.; Yousefi, A.R. Occurrence, pathogenicity and distribution of Fusarium spp. in stored wheat seeds Kermanshah Province, Iran. Pak. J. Biol. Sci. 2010, 13, 1178–1186. [Google Scholar] [CrossRef] [Green Version]
- Chehri, K. Occurrence of Fusarium species associated with economically important agricultural crops in Iran. Afr. J. Microbiol. Res. 2011, 5, 4043–4048. [Google Scholar] [CrossRef]
- Minati, M.H. First record of nine Fusarium spp. causing root rot on wheat (Triticum aestivum L.) in Iraq. AIP Conf. Proc. 2020, 2290, 020009. [Google Scholar] [CrossRef]
- Fernandez, M.R.; Jefferson, P.G. Fungal populations in roots and crowns of common and durum wheat in Saskatchewan. Can. J. Plant Pathol. 2004, 26, 325–334. [Google Scholar] [CrossRef]
- Rossi, V.; Cervi, C.; Chiusa, G.; Languasco, L. Fungi associated with foot rots on winter wheat in northwest Italy. J. Phytopathol. 1995, 143, 115–119. [Google Scholar] [CrossRef]
- Bentley, A.R.; Cromey, M.G.; Farrokhi-Nejad, R.; Leslie, J.F.; Summerell, B.A.; Burgess, L.W. Fusarium crown and root rot pathogens associated with wheat and grass stem bases on the South Island of New Zealand. Australas. Plant Pathol. 2006, 35, 495–502. [Google Scholar] [CrossRef]
- Jevtić, R.; Stošić, N.; Župunski, V.; Lalošević, M.; Orbović, B. Variability of stem-base infestation and coexistence of Fusarium spp. causing crown rot of winter wheat in Serbia. Plant Pathol. J. 2019, 35, 553. [Google Scholar] [CrossRef]
- Taheri Esmaeili, A.; Hamel, C.; Gan, Y.; Vujanovic, V. First report of Fusarium redolens from Saskatchewan and its comparative pathogenicity. Can. J. Plant Pathol. 2011, 33, 559–564. [Google Scholar] [CrossRef]
- Gebremariam, E.S.; Dababat, A.A.; Erginbas-Orakci, G.; Karakaya, A.; Poudyal, D.S.; Paulitz, T.C. First report of Fusarium hostae causing crown rot on wheat (Triticum spp.) in Turkey. Plant Dis. 2016, 100, 216. [Google Scholar] [CrossRef]
- Kane, R.T.; Smiley, R.W.; Sorrells, M.E. Relative pathogenicity of selected Fusarium species and Microdochium bolleyi to winter wheat in New York. Plant Dis. 1987, 71, 177–181. [Google Scholar] [CrossRef]
- Jonavičienė, A.; Supronienė, S.; Semaškienė, R. Microdochium nivale and M. majus as causative agents of seedling blight in spring cereals. Zemdirb.-Agric. 2016, 103, 363–368. [Google Scholar] [CrossRef] [Green Version]
- Bouaicha, O.; Laraba, I.; Boureghda, H. Identification, in vitro growth and pathogenicity of Microdochium spp. associated with wheat crown rot in Algeria. J. Plant Pathol. 2022, 104, 1431–1442. [Google Scholar] [CrossRef]
- Xu, F.; Shi, R.J.; Zhang, J.J.; Song, Y.L.; Liu, L.L.; Han, Z.H.; Wang, J.M.; Li, Y.H.; Feng, C.H.; Li, L.J. First report of Microdochium nivale and M. majus causing brown foot rot of wheat in China. Plant Dis. 2022, 106, 1523. [Google Scholar] [CrossRef]
- Leyva-Mir, S.G.; Vega-Portillo, H.E.; Villaseñor-Mir, H.E.; Tlapal-Bolaños, B.; Vargas-Hernández, M.; Camacho-Tapia, M.; Tovar-Pedraza, J.M. Characterization of Fusarium species causing root rot of wheat in the Bajio, Mexico. Chil. J. Agric. Anim. Sci. Ex Agro-Cienc. 2017, 33, 142–151. [Google Scholar]
- Dehghanpour-Farashah, S.; Taheri, P.; Falahati-Rastegar, M. Identification and pathogenicity of Fusarium spp., the causal agent of wheat crown and root rot in Iran. J. Plant Pathol. 2020, 102, 143–154. [Google Scholar] [CrossRef]
- Tillmann, M.; von Tiedemann, A.; Winter, M. Crop rotation effects on incidence and diversity of Fusarium species colonizing stem bases and grains of winter wheat. J. Plant Dis. Prot. 2017, 124, 121–130. [Google Scholar] [CrossRef]
- Kazan, K.; Gardiner, D.M. Fusarium crown rot caused by Fusarium pseudograminearum in cereal crops: Recent progress and future prospects. Mol. Plant Pathol. 2018, 19, 1547–1562. [Google Scholar] [CrossRef] [Green Version]
- Fernandez, M.R.; Chen, Y. Pathogenicity of Fusarium species on different plant parts of spring wheat under controlled conditions. Plant Dis. 2005, 89, 164–169. [Google Scholar] [CrossRef] [Green Version]
- Özer, G.; Erper, I.; İmren, M.; Bozoglu, T.; Ozdemir, F.; Dababat, A.A. First report of crown rot caused by Fusarium algeriense on wheat in Kyrgyzstan. Plant Dis. 2022, 106, 2998. [Google Scholar] [CrossRef]
- Fouly, H.M.; Pedersen, W.L.; Wilkinson, H.T.; Abd El-Kader, M.M. Wheat root rotting fungi in the “old” and “new” agricultural lands of Egypt. Plant Dis. 1996, 80, 1298–1300. [Google Scholar] [CrossRef]
- Douglas, L.I.; Deacon, J.W. Strain variation in tolerance of water stress by Idriella Microdochium bolleyi, a biocontrol agent of cereal root and stem base pathogens. Biocontrol Sci. 1994, 4, 239–249. [Google Scholar] [CrossRef]
- Matušinsky, P.; Sedláková, B.; Bleša, D. Compatible interaction of Brachypodium distachyon and endophytic fungus Microdochium bolleyi. PLoS ONE 2022, 17, e0265357. [Google Scholar] [CrossRef]
- Demirci, E.; Dane, E. Identification and pathogencity of Fusarium spp. from stem bases of winter wheat in Erzurum, Turkey. Phytoparasitica 2003, 31, 170–173. [Google Scholar] [CrossRef]
- Wagacha, J.M.; Muthomi, J.W. Fusarium culmorum: Infection process, mechanisms of mycotoxin production and their role in pathogenesis in wheat. Crop Prot. 2007, 26, 877–885. [Google Scholar] [CrossRef]
- Temirbekova, S.K.; Kulikov, I.M.; Ashirbekov, M.Z.; Afanasyeva, Y.V.; Beloshapkina, O.O.; Tyryshkin, L.G.; Zuev, E.V.; Kirakosyan, R.N.; Glinushkin, A.P.; Potapova, E.S.; et al. Evaluation of wheat resistance to snow mold caused by Microdochium nivale (Fr) Samuels and I.C. Hallett under abiotic stress influence in the Central Non-Black Earth Region of Russia. Plants 2022, 11, 699. [Google Scholar] [CrossRef]
- O’Donnell, K.; Whitaker, B.K.; Laraba, I.; Proctor, R.H.; Brown, D.W.; Broders, K.; Geiser, D.M. DNA sequence-based identification of Fusarium: A work in progress. Plant Dis. 2022, 106, PDIS-09. [Google Scholar] [CrossRef]
- O’Donnell, K.; Nirenberg, H.I.; Aoki, T.; Cigelnik, E.A. Multigene phylogeny of the Gibberella fujikuroi species complex: Detection of additional phylogenetically distinct species. Mycoscience 2000, 41, 61–78. [Google Scholar] [CrossRef]
- Geiser, D.M.; Al-Hatmi, A.M.; Aoki, T.; Arie, T.; Balmas, V.; Barnes, I.; Viljoen, A. Phylogenomic analysis of a 55.1-kb 19-gene dataset resolves a monophyletic Fusarium that includes the Fusarium solani species complex. Phytopathology 2021, 111, 1064–1079. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Wang, M.M.; Groenewald, M.; Li, A.H.; Guo, Y.T.; Wu, F.; Begerow, D. Proposal of Two New Combinations, Twenty New Species, Four New Genera, One New Family, and One New Order for the Anamorphic Basidiomycetous Yeast Species in Ustilaginomycotina. Front. Microbiol. 2022, 12, 777338. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Pan, M.; Tian, C.; Fan, X. Fungal Richness of Cytospora Species Associated with Willow Canker Disease in China. J. Fungi 2022, 8, 377. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wingfield, M.J.; Duong, T.A.; Wingfield, B.D.; Chen, S. Diversity and Distribution of Calonectria Species from Plantation and Forest Soils in Fujian Province, China. J. Fungi 2022, 8, 811. [Google Scholar] [CrossRef]
- Costa, M.M.; Saleh, A.A.; Melo, M.P.; GuimarŃes, E.A.; Esele, J.P.; Zeller, K.A.; Leslie, J.F. Fusarium mirum sp. nov, intertwining Fusarium madaense and Fusarium andiyazi, pathogens of tropical grasses. Fungal Biol. 2022, 126, 250–266. [Google Scholar] [CrossRef]
- Suga, H.; Kitajima, M.; Nagumo, R.; Tsukiboshi, T.; Uegaki, R.; Nakajima, T.; Kushiro, M.; Nakagawa, H.; Shimizu, M.; Kageyama, K.; et al. A single nucleotide polymorphism in the translation elongation factor 1alpha gene correlates with the ability to produce fumonisin in Japanese Fusarium fujikuroi. Fungal Biol. 2014, 118, 402–412. [Google Scholar] [CrossRef]
- Suga, H.; Arai, M.; Fukasawa, E.; Motohashi, K.; Nakagawa, H.; Tateishi, H.; Hyakumachi, M. Genetic differentiation associated with fumonisin and gibberellin production in Japanese Fusarium fujikuroi. Appl. Environ. Microbiol. 2019, 85, e02414-18. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Species * | Bishkek | Sokuluk | Kara-Balta | Total | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NF | NI | IF | NF | NI | IF | NF | NI | IF | NF | NI | FI | IF | |
| Bipolaris sorokiniana | 9 | 87 | 44.39 | 8 | 77 | 37.75 | 9 | 79 | 39.90 | 26 | 243 | 86.67 | 40.64 |
| Fusarium acuminatum | 5 | 31 | 15.82 | 8 | 51 | 25.00 | 8 | 47 | 23.74 | 21 | 129 | 70.00 | 21.57 |
| Fusarium culmorum | 3 | 11 | 5.61 | 6 | 25 | 12.25 | 5 | 30 | 15.15 | 14 | 66 | 46.67 | 11.04 |
| Fusarium nygamai | 8 | 41 | 20.92 | 1 | 3 | 1.47 | 0 | 0 | 0.00 | 9 | 44 | 30.00 | 7.36 |
| Fusarium equiseti | 3 | 6 | 3.06 | 1 | 5 | 2.45 | 3 | 15 | 7.58 | 7 | 26 | 23.33 | 4.35 |
| Fusarium oxysporum | 2 | 5 | 2.55 | 2 | 6 | 2.94 | 5 | 8 | 4.04 | 9 | 19 | 30.00 | 3.18 |
| Curvularia inaequalis | 0 | 0 | 0.00 | 2 | 13 | 6.37 | 1 | 6 | 3.03 | 3 | 19 | 10.00 | 3.18 |
| Fusarium pseudograminearum | 0 | 0 | 0.00 | 3 | 8 | 3.92 | 2 | 9 | 4.55 | 5 | 17 | 16.67 | 2.84 |
| Macrophomina phaseolina | 1 | 2 | 1.02 | 4 | 6 | 2.94 | 2 | 3 | 1.52 | 7 | 11 | 23.33 | 1.84 |
| Fusarium redolens | 4 | 8 | 4.08 | 0 | 0 | 0.00 | 0 | 0 | 0.00 | 4 | 8 | 13.33 | 1.34 |
| Microdochium bolleyi | 1 | 3 | 1.53 | 1 | 2 | 0.98 | 0 | 0 | 0.00 | 2 | 5 | 6.67 | 0.84 |
| Microdochium nivale | 0 | 0 | 0.00 | 2 | 3 | 1.47 | 0 | 0 | 0.00 | 2 | 3 | 6.67 | 0.50 |
| Fusarium proliferatum | 1 | 2 | 1.02 | 0 | 0 | 0.00 | 0 | 0 | 0.00 | 1 | 2 | 3.33 | 0.33 |
| Fusarium burgessii | 0 | 0 | 0.00 | 2 | 2 | 0.98 | 0 | 0 | 0.00 | 2 | 2 | 6.67 | 0.33 |
| Fusarium tricinctum | 0 | 0 | 0.00 | 1 | 2 | 0.98 | 0 | 0 | 0.00 | 1 | 2 | 3.33 | 0.33 |
| Fusarium solani | 0 | 0 | 0.00 | 0 | 0 | 0.00 | 1 | 1 | 0.51 | 1 | 1 | 3.33 | 0.17 |
| Fusarium algeriense | 0 | 0 | 0.00 | 1 | 1 | 0.49 | 0 | 0 | 0.00 | 1 | 1 | 3.33 | 0.17 |
| Total | 10 | 196 | 100 | 10 | 204 | 100 | 10 | 198 | 100 | 30 | 598 | 100 | 100 |
| Species | Number of Isolates | Average Severity Index * | Average Disease Severity (%) *** | Virulence Category **** |
|---|---|---|---|---|
| Fusarium pseudograminearum | 3 | 3.67 ± 0.49 a ** | 73.33 | HV |
| Fusarium culmorum | 3 | 3.62 ± 0.51 a | 72.33 | HV |
| Bipolaris sorokiniana | 3 | 2.78 ± 0.39 b | 55.50 | MV |
| Fusarium redolens | 3 | 2.23 ± 0.28 c | 44.67 | MV |
| Fusarium algeriense | 1 | 2.22 ± 0.21 bd | 44.50 | MV |
| Curvularia inaequalis | 3 | 1.67 ± 0.26 de | 33.33 | MiV |
| Fusarium nygamai | 3 | 1.43 ± 0.23 e | – | NP |
| Fusarium solani | 1 | 1.42 ± 0.27 e | – | NP |
| Microdochium bolleyi | 3 | 1.42 ± 0.37 e | – | NP |
| Fusarium oxysporum | 3 | 1.40 ± 0.15 e | – | NP |
| Fusarium proliferatum | 2 | 1.39 ± 0.21 e | – | NP |
| Macrophomina phaseolina | 3 | 1.37 ± 0.24 e | – | NP |
| Microdochium nivale | 3 | 1.32 ± 0.26 e | – | NP |
| Fusarium burgessii | 2 | 1.29 ± 0.15 e | – | NP |
| Fusarium tricinctum | 2 | 1.28 ± 0.18 e | – | NP |
| Fusarium acuminatum | 3 | 1.26 ± 0.23 e | – | NP |
| Fusarium equiseti | 3 | 1.25 ± 0.16 e | – | NP |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Özer, G.; Erper, İ.; Yıldız, Ş.; Bozoğlu, T.; Zholdoshbekova, S.; Alkan, M.; Tekin, F.; Uulu, T.E.; İmren, M.; Dababat, A.A.; et al. Fungal Pathogens Associated with Crown and Root Rot in Wheat-Growing Areas of Northern Kyrgyzstan. J. Fungi 2023, 9, 124. https://doi.org/10.3390/jof9010124
Özer G, Erper İ, Yıldız Ş, Bozoğlu T, Zholdoshbekova S, Alkan M, Tekin F, Uulu TE, İmren M, Dababat AA, et al. Fungal Pathogens Associated with Crown and Root Rot in Wheat-Growing Areas of Northern Kyrgyzstan. Journal of Fungi. 2023; 9(1):124. https://doi.org/10.3390/jof9010124
Chicago/Turabian StyleÖzer, Göksel, İsmail Erper, Şenol Yıldız, Tuğba Bozoğlu, Sezim Zholdoshbekova, Mehtap Alkan, Fatih Tekin, Tair Esenali Uulu, Mustafa İmren, Abdelfattah A. Dababat, and et al. 2023. "Fungal Pathogens Associated with Crown and Root Rot in Wheat-Growing Areas of Northern Kyrgyzstan" Journal of Fungi 9, no. 1: 124. https://doi.org/10.3390/jof9010124
APA StyleÖzer, G., Erper, İ., Yıldız, Ş., Bozoğlu, T., Zholdoshbekova, S., Alkan, M., Tekin, F., Uulu, T. E., İmren, M., Dababat, A. A., & Derviş, S. (2023). Fungal Pathogens Associated with Crown and Root Rot in Wheat-Growing Areas of Northern Kyrgyzstan. Journal of Fungi, 9(1), 124. https://doi.org/10.3390/jof9010124