
Virulence Traits and Azole Resistance in Korean Candida auris Isolates

 , , , , , ,
, , , , , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Fungal Isolates and Antifungal Susceptibility Testing
2.2. MLST
2.3. Thermotolerance Assay
2.4. Biofilm Formation and Aggregate-Forming Capacity
2.5. In Vivo Virulence Analysis Using Galleria Mellonella
2.6. In Vitro Competitive Growth Assay
2.7. Sequence Analysis of ERG11, TAC1A, and TAC1B
2.8. Statistical Analyses
3. Results
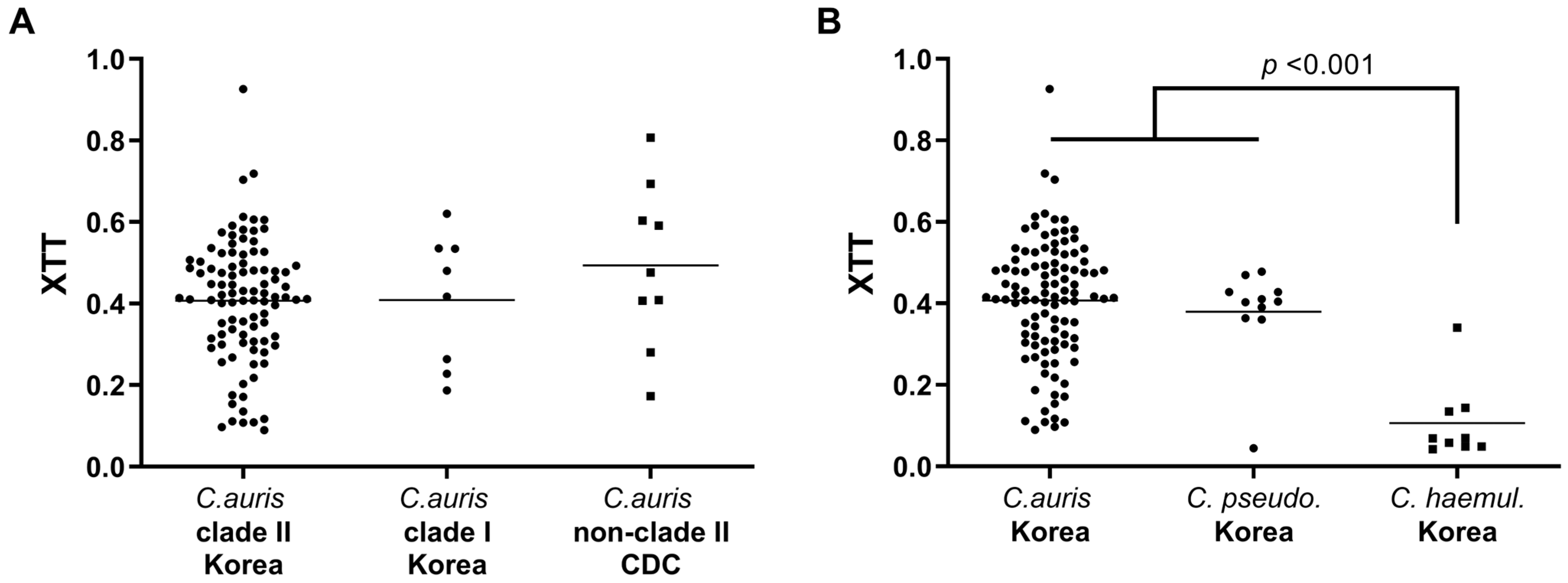
3.1. MLST Analysis and Antifungal Susceptibility Testing
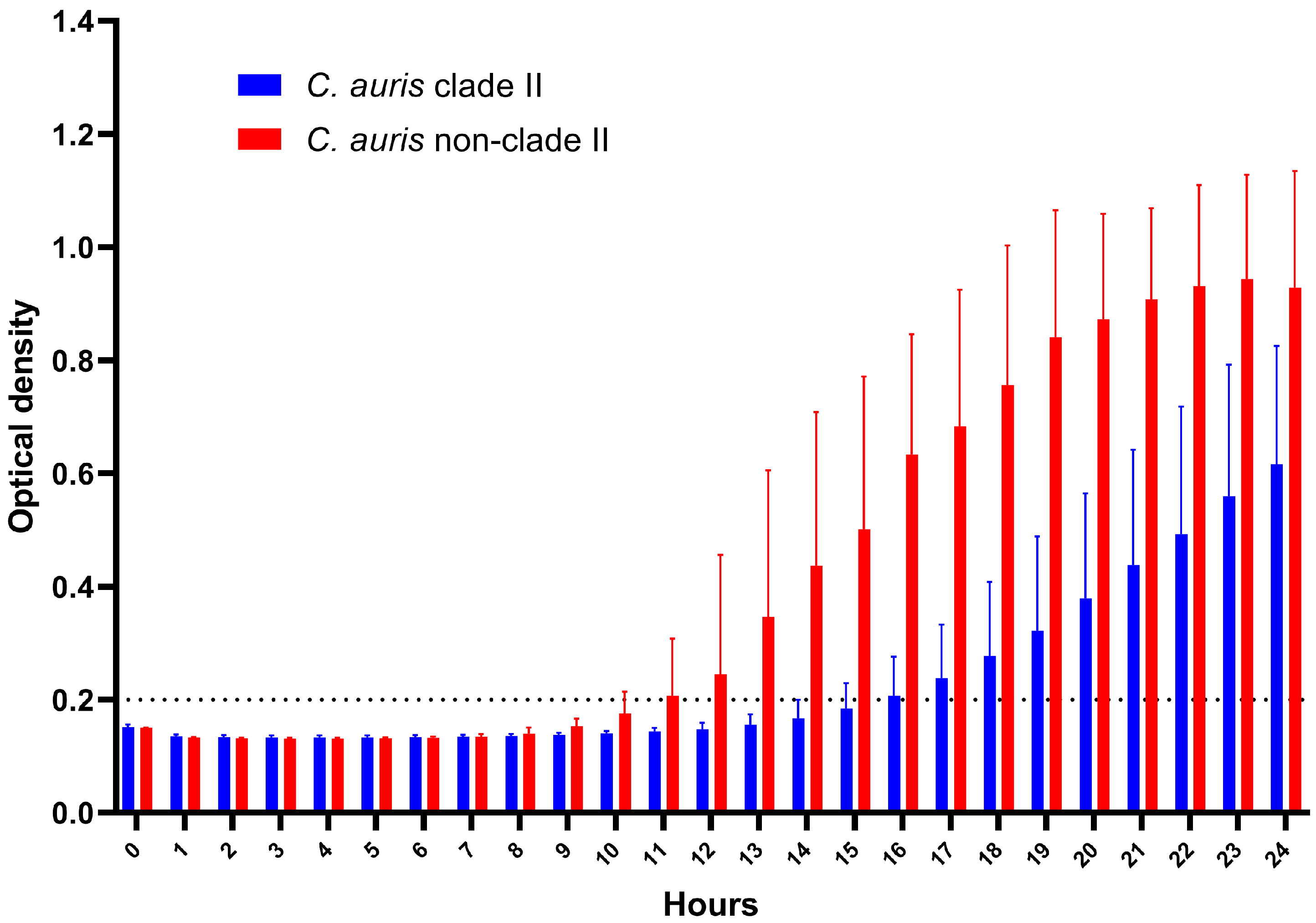
3.2. Thermotolerance and Biofilm Formation Assay
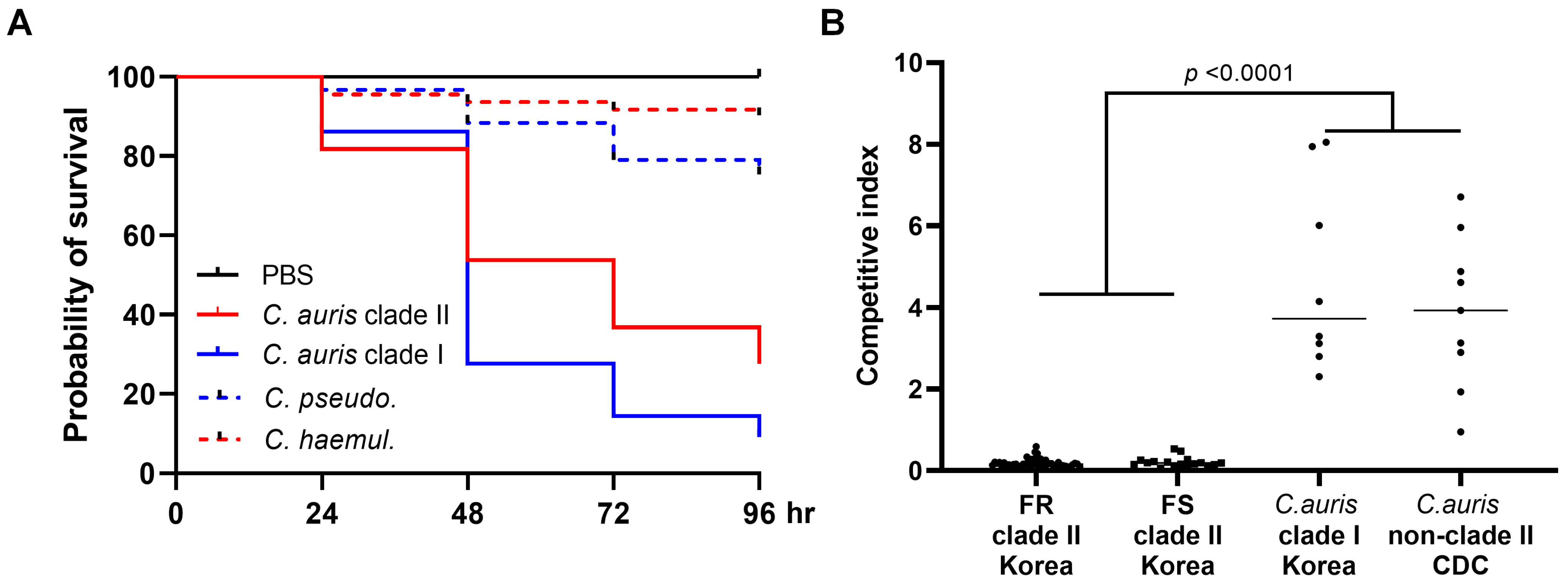
3.3. In Vivo Virulence Analysis Using G. mellonella and In Vitro Competitive Growth Assay
3.4. Sequence Analysis of ERG11, TAC1A, and TAC1B
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Satoh, K.; Makimura, K.; Hasumi, Y.; Nishiyama, Y.; Uchida, K.; Yamaguchi, H. Candida auris sp. nov., a novel ascomycetous yeast isolated from the external ear canal of an inpatient in a Japanese hospital. Microbiol. Immunol. 2009, 53, 41–44. [Google Scholar] [CrossRef] [PubMed]
- Spivak, E.S.; Hanson, K.E. Candida auris: An Emerging Fungal Pathogen. J. Clin. Microbiol. 2018, 56. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.N.; Shin, J.H.; Sung, H.; Lee, K.; Kim, E.C.; Ryoo, N.; Lee, J.S.; Jung, S.I.; Park, K.H.; Kee, S.J.; et al. Candida haemulonii and closely related species at 5 university hospitals in Korea: Identification, antifungal susceptibility, and clinical features. Clin. Infect. Dis. 2009, 48, e57–e61. [Google Scholar] [CrossRef] [PubMed]
- Lee, W.G.; Shin, J.H.; Uh, Y.; Kang, M.G.; Kim, S.H.; Park, K.H.; Jang, H.C. First three reported cases of nosocomial fungemia caused by Candida auris. J. Clin. Microbiol. 2011, 49, 3139–3142. [Google Scholar] [CrossRef] [PubMed]
- CDC. Centers for Disease Control and Prevention Tracking Candida auris. Available online: https://www.cdc.gov/fungal/candida-auris/tracking-c-auris.html (accessed on 10 September 2023).
- Du, H.; Bing, J.; Hu, T.; Ennis, C.L.; Nobile, C.J.; Huang, G. Candida auris: Epidemiology, biology, antifungal resistance, and virulence. PLoS Pathog. 2020, 16, e1008921. [Google Scholar] [CrossRef] [PubMed]
- Kean, R.; Ramage, G. Combined Antifungal Resistance and Biofilm Tolerance: The Global Threat of Candida auris. mSphere 2019, 4, e00458-19. [Google Scholar] [CrossRef] [PubMed]
- Lockhart, S.R.; Etienne, K.A.; Vallabhaneni, S.; Farooqi, J.; Chowdhary, A.; Govender, N.P.; Colombo, A.L.; Calvo, B.; Cuomo, C.A.; Desjardins, C.A.; et al. Simultaneous Emergence of Multidrug-Resistant Candida auris on 3 Continents Confirmed by Whole-Genome Sequencing and Epidemiological Analyses. Clin. Infect. Dis. 2017, 64, 134–140. [Google Scholar] [CrossRef]
- Escandon, P.; Chow, N.A.; Caceres, D.H.; Gade, L.; Berkow, E.L.; Armstrong, P.; Rivera, S.; Misas, E.; Duarte, C.; Moulton-Meissner, H.; et al. Molecular Epidemiology of Candida auris in Colombia Reveals a Highly Related, Countrywide Colonization With Regional Patterns in Amphotericin B Resistance. Clin. Infect. Dis. 2019, 68, 15–21. [Google Scholar] [CrossRef]
- Rhodes, J.; Abdolrasouli, A.; Farrer, R.A.; Cuomo, C.A.; Aanensen, D.M.; Armstrong-James, D.; Fisher, M.C.; Schelenz, S. Author Correction: Genomic epidemiology of the UK outbreak of the emerging human fungal pathogen Candida auris. Emerg. Microbes Infect. 2018, 7, 104. [Google Scholar] [CrossRef]
- Kwon, Y.J.; Shin, J.H.; Byun, S.A.; Choi, M.J.; Won, E.J.; Lee, D.; Lee, S.Y.; Chun, S.; Lee, J.H.; Choi, H.J.; et al. Candida auris Clinical Isolates from South Korea: Identification, Antifungal Susceptibility, and Genotyping. J. Clin. Microbiol. 2019, 57, 01624-18. [Google Scholar] [CrossRef]
- Chow, N.A.; de Groot, T.; Badali, H.; Abastabar, M.; Chiller, T.M.; Meis, J.F. Potential Fifth Clade of Candida auris, Iran, 2018. Emerg. Infect. Dis. 2019, 25, 1780–1781. [Google Scholar] [CrossRef] [PubMed]
- Sathi, F.A.; Aung, M.S.; Paul, S.K.; Nasreen, S.A.; Haque, N.; Roy, S.; Ahmed, S.; Alam, M.M.; Khan, S.; Rabbany, M.A.; et al. Clonal Diversity of Candida auris, Candida blankii, and Kodamaea ohmeri Isolated from Septicemia and Otomycosis in Bangladesh as Determined by Multilocus Sequence Typing. J. Fungi 2023, 9, 658. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Messer, S.A.; Deshpande, L.M.; Rhomberg, P.R.; Utt, E.A.; Castanheira, M. Evaluation of Synergistic Activity of Isavuconazole or Voriconazole plus Anidulafungin and the Occurrence and Genetic Characterization of Candida auris Detected in a Surveillance Program. Antimicrob. Agents Chemother. 2021, 65, e02031-20. [Google Scholar] [CrossRef] [PubMed]
- Magobo, R.; Mhlanga, M.; Corcoran, C.; Govender, N.P. Multilocus sequence typing of azole-resistant Candida auris strains, South Africa. S Afr. J. Infect. Dis. 2020, 35, 116. [Google Scholar] [CrossRef] [PubMed]
- Sharma, C.; Kadosh, D. Perspective on the origin, resistance, and spread of the emerging human fungal pathogen Candida auris. PLoS Pathog. 2023, 19, e1011190. [Google Scholar] [CrossRef]
- Forgacs, L.; Borman, A.M.; Prepost, E.; Toth, Z.; Kardos, G.; Kovacs, R.; Szekely, A.; Nagy, F.; Kovacs, I.; Majoros, L. Comparison of in vivo pathogenicity of four Candida auris clades in a neutropenic bloodstream infection murine model. Emerg. Microbes Infect. 2020, 9, 1160–1169. [Google Scholar] [CrossRef]
- Spettel, K.; Kriz, R.; Wu, C.; Achter, L.; Schmid, S.; Galazka, S.; Selitsch, B.; Camp, I.; Makristathis, A.; Lagler, H.; et al. Candida auris in Austria-What Is New and What Is Different. J. Fungi 2023, 9, 129. [Google Scholar] [CrossRef]
- Narayanan, A.; Vadnala, R.N.; Ganguly, P.; Selvakumar, P.; Rudramurthy, S.M.; Prasad, R.; Chakrabarti, A.; Siddharthan, R.; Sanyal, K. Functional and Comparative Analysis of Centromeres Reveals Clade-Specific Genome Rearrangements in Candida auris and a Chromosome Number Change in Related Species. mBio 2021, 12, e00905-21. [Google Scholar] [CrossRef]
- Jackson, B.R.; Chow, N.; Forsberg, K.; Litvintseva, A.P.; Lockhart, S.R.; Welsh, R.; Vallabhaneni, S.; Chiller, T. On the Origins of a Species: What Might Explain the Rise of Candida auris? J. Fungi 2019, 5, 58. [Google Scholar] [CrossRef]
- Jangir, P.; Kalra, S.; Tanwar, S.; Bari, V.K. Azole resistance in Candida auris: Mechanisms and combinatorial therapy. APMIS 2023, 131, 442–462. [Google Scholar] [CrossRef]
- Morschhäuser, J.; Barker, K.S.; Liu, T.T.; Blaß-Warmuth, J.; Homayouni, R.; Rogers, P.D. The transcription factor Mrr1p controls expression of the MDR1 efflux pump and mediates multidrug resistance in Candida albicans. PLoS Pathog. 2007, 3, e164. [Google Scholar] [CrossRef]
- Rybak, J.M.; Munoz, J.F.; Barker, K.S.; Parker, J.E.; Esquivel, B.D.; Berkow, E.L.; Lockhart, S.R.; Gade, L.; Palmer, G.E.; White, T.C.; et al. Mutations in TAC1B: A Novel Genetic Determinant of Clinical Fluconazole Resistance in Candida auris. mBio 2020, 11, e00365-20. [Google Scholar] [CrossRef]
- Mayr, E.M.; Ramirez-Zavala, B.; Krüger, I.; Morschhäuser, J. A zinc cluster transcription factor contributes to the intrinsic fluconazole resistance of Candida auris. mSphere 2020, 5, e00279-20. [Google Scholar] [CrossRef] [PubMed]
- Bing, J.; Hu, T.; Zheng, Q.; Muñoz, J.F.; Cuomo, C.A.; Huang, G. Experimental Evolution Identifies Adaptive Aneuploidy as a Mechanism of Fluconazole Resistance in Candida auris. Antimicrob. Agents Chemother. 2020, 65, e01466-20. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Holowka, T.; Orner, E.P.; Fries, B.C. Gene Duplication Associated with Increased Fluconazole Tolerance in Candida auris cells of Advanced Generational Age. Sci. Rep. 2019, 9, 5052. [Google Scholar] [CrossRef] [PubMed]
- Lima, S.L.; Francisco, E.C.; de Almeida Junior, J.N.; Santos, D.; Carlesse, F.; Queiroz-Telles, F.; Melo, A.S.A.; Colombo, A.L. Increasing Prevalence of Multidrug-Resistant Candida haemulonii Species Complex among All Yeast Cultures Collected by a Reference Laboratory over the Past 11 Years. J. Fungi 2020, 6, 110. [Google Scholar] [CrossRef] [PubMed]
- Won, E.J.; Choi, M.J.; Jeong, S.H.; Kim, D.; Shin, K.S.; Shin, J.H.; Kim, Y.R.; Kim, H.S.; Kim, Y.A.; Uh, Y.; et al. Nationwide Surveillance of Antifungal Resistance of Candida Bloodstream Isolates in South Korean Hospitals: Two Year Report from Kor-GLASS. J. Fungi 2022, 8, 996. [Google Scholar] [CrossRef]
- Prakash, A.; Sharma, C.; Singh, A.; Kumar Singh, P.; Kumar, A.; Hagen, F.; Govender, N.P.; Colombo, A.L.; Meis, J.F.; Chowdhary, A. Evidence of genotypic diversity among Candida auris isolates by multilocus sequence typing, matrix-assisted laser desorption ionization time-of-flight mass spectrometry and amplified fragment length polymorphism. Clin. Microbiol. Infect. 2016, 22, 277.e1–277.e9. [Google Scholar] [CrossRef]
- Ben-Ami, R.; Berman, J.; Novikov, A.; Bash, E.; Shachor-Meyouhas, Y.; Zakin, S.; Maor, Y.; Tarabia, J.; Schechner, V.; Adler, A.; et al. Multidrug-Resistant Candida haemulonii and C. auris, Tel Aviv, Israel. Emerg. Infect. Dis. 2017, 23, 195–203. [Google Scholar] [CrossRef]
- Bordallo-Cardona, M.Á.; Sánchez-Carrillo, C.; Muñoz, P.; Bouza, E.; Escribano, P.; Guinea, J. Growth kinetics in Candida spp.: Differences between species and potential impact on antifungal susceptibility testing as described by the EUCAST. Med. Mycol. 2018, 57, 601–608. [Google Scholar] [CrossRef]
- Jung, S.I.; Shin, J.H.; Kim, S.H.; Kim, J.; Kim, J.H.; Choi, M.J.; Chung, E.K.; Lee, K.; Koo, S.H.; Chang, H.H.; et al. Comparison of E,E-Farnesol Secretion and the Clinical Characteristics of Candida albicans Bloodstream Isolates from Different Multilocus Sequence Typing Clades. PLoS ONE 2016, 11, e0148400. [Google Scholar] [CrossRef]
- Chandra, J.; Mukherjee, P.K.; Ghannoum, M.A. In vitro growth and analysis of Candida biofilms. Nat. Protoc. 2008, 3, 1909–1924. [Google Scholar] [CrossRef]
- Sherry, L.; Ramage, G.; Kean, R.; Borman, A.; Johnson, E.M.; Richardson, M.D.; Rautemaa-Richardson, R. Biofilm-Forming Capability of Highly Virulent, Multidrug-Resistant Candida auris. Emerg. Infect. Dis. 2017, 23, 328–331. [Google Scholar] [CrossRef] [PubMed]
- Popp, C.; Hampe, I.A.I.; Hertlein, T.; Ohlsen, K.; Rogers, P.D.; Morschhauser, J. Competitive Fitness of Fluconazole-Resistant Clinical Candida albicans Strains. Antimicrob. Agents Chemother. 2017, 61, e00584-17. [Google Scholar] [CrossRef]
- Li, H.; Wang, Q.; Wang, R.; Zhang, Y.; Wang, X.; Wang, H. Global regulator SoxR is a negative regulator of efflux pump gene expression and affects antibiotic resistance and fitness in Acinetobacter baumannii. Medicine 2017, 96, e7188. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Coste, A.T.; Liechti, M.; Bachmann, D.; Sanglard, D.; Lamoth, F. Novel ERG11 and TAC1b mutations associated with azole resistance in Candida auris. Antimicrob. Agents Chemother. 2023, 65, e02663-20. [Google Scholar] [CrossRef]
- Chow, N.A.; Gade, L.; Tsay, S.V.; Forsberg, K.; Greenko, J.A.; Southwick, K.L.; Barrett, P.M.; Kerins, J.L.; Lockhart, S.R.; Chiller, T.M.; et al. Multiple introductions and subsequent transmission of multidrug-resistant Candida auris in the USA: A molecular epidemiological survey. Lancet Infect. Dis. 2018, 18, 1377–1384. [Google Scholar] [CrossRef]
- Chow, N.A.; Munoz, J.F.; Gade, L.; Berkow, E.L.; Li, X.; Welsh, R.M.; Forsberg, K.; Lockhart, S.R.; Adam, R.; Alanio, A.; et al. Tracing the Evolutionary History and Global Expansion of Candida auris Using Population Genomic Analyses. mBio 2020, 11, e03364-19. [Google Scholar] [CrossRef]
- Rybak, J.M.; Cuomo, C.A.; Rogers, P.D. The molecular and genetic basis of antifungal resistance in the emerging fungal pathogen Candida auris. Curr. Opin. Microbiol. 2022, 70, 102208. [Google Scholar] [CrossRef]
- Shastri, P.S.; Shankarnarayan, S.A.; Oberoi, J.; Rudramurthy, S.M.; Wattal, C.; Chakrabarti, A. Candida auris candidaemia in an intensive care unit—Prospective observational study to evaluate epidemiology, risk factors, and outcome. J. Crit. Care 2020, 57, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Sayeed, M.A.; Farooqi, J.; Jabeen, K.; Awan, S.; Mahmood, S.F. Clinical spectrum and factors impacting outcome of Candida auris: A single center study from Pakistan. BMC Infect. Dis. 2019, 19, 384. [Google Scholar] [CrossRef] [PubMed]
- Chowdhary, A.; Tarai, B.; Singh, A.; Sharma, A. Multidrug-Resistant Candida auris Infections in Critically Ill Coronavirus Disease Patients, India, April–July 2020. Emerg. Infect. Dis. 2020, 26, 2694–2696. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Bing, J.; Zheng, Q.; Zhang, F.; Liu, J.; Yue, H.; Tao, L.; Du, H.; Wang, Y.; Wang, H.; et al. The first isolate of Candida auris in China: Clinical and biological aspects. Emerg. Microbes Infect. 2018, 7, 93. [Google Scholar] [CrossRef]
- Tsai, Y.T.; Lu, P.L.; Tang, H.J.; Huang, C.H.; Hung, W.C.; Tseng, Y.T.; Lee, K.M.; Lin, S.Y. The first invasive Candida auris infection in Taiwan. Emerg. Microbes Infect. 2022, 11, 1867–1875. [Google Scholar] [CrossRef] [PubMed]
- Cendejas-Bueno, E.; Kolecka, A.; Alastruey-Izquierdo, A.; Theelen, B.; Groenewald, M.; Kostrzewa, M.; Cuenca-Estrella, M.; Gomez-Lopez, A.; Boekhout, T. Reclassification of the Candida haemulonii complex as Candida haemulonii (C. haemulonii group I), C. duobushaemulonii sp. nov. (C. haemulonii group II), and C. haemulonii var. vulnera var. nov.: Three multiresistant human pathogenic yeasts. J. Clin. Microbiol. 2012, 50, 3641–3651. [Google Scholar] [CrossRef] [PubMed]
- Sipiczki, M.; Tap, R.M. Candida vulturna pro tempore sp. nov., a dimorphic yeast species related to the Candida haemulonis species complex isolated from flowers and clinical sample. Int. J. Syst. Evol. Microbiol. 2016, 66, 4009–4015. [Google Scholar] [CrossRef]
- Sugita, T.; Takashima, M.; Poonwan, N.; Mekha, N. Candida pseudohaemulonii Sp. Nov., an amphotericin B-and azole-resistant yeast species, isolated from the blood of a patient from Thailand. Microbiol. Immunol. 2006, 50, 469–473. [Google Scholar] [CrossRef]
- de Barros Rodrigues, D.K.; Lockhart, S.R.; Berkow, E.L.; Gade, L.; Bonfietti, L.X.; Mazo Favero Gimenes, V.; Silva Ruiz, L.; Bronze Macioni, M.; de Souza Carvalho Melhem, M. Whole-genome sequencing of Candida haemulonii species complex from Brazil and the United States: Genetic diversity and antifungal susceptibility. Med. Mycol. 2023, 61, myad030. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, A.; Kontoyiannis, D.P.; Robert, V. On the Emergence of Candida auris: Climate Change, Azoles, Swamps, and Birds. mBio 2019, 10, e01397-19. [Google Scholar] [CrossRef]
- Casadevall, A.; Kontoyiannis, D.P.; Robert, V. Environmental Candida auris and the Global Warming Emergence Hypothesis. mBio 2021, 12, e00360-21. [Google Scholar] [CrossRef]
- Jung, J.; Kim, M.J.; Kim, J.Y.; Lee, J.Y.; Kwak, S.H.; Hong, M.J.; Chong, Y.P.; Lee, S.O.; Choi, S.H.; Kim, Y.S.; et al. Candida auris colonization or infection of the ear: A single-center study in South Korea from 2016 to 2018. Med. Mycol. 2020, 58, 124–127. [Google Scholar] [CrossRef] [PubMed]
- Rybak, J.M.; Sharma, C.; Doorley, L.A.; Barker, K.S.; Palmer, G.E.; Rogers, P.D. Delineation of the Direct Contribution of Candida auris ERG11 Mutations to Clinical Triazole Resistance. Microbiol. Spectr. 2021, 9, e0158521. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Agent | Species/Clade * | No. Tested | No. of Isolates with the Following MIC (mg/L) ** | No. (%) of Resistant Isolates † | Geometric Mean of MICs | |||||||||||
| ≤0.03 | 0.06 | 0.12 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | ≥64 | |||||
| Antifungals which tentative MIC breakpoints were provided by the US CDC | ||||||||||||||||
| FLU | C. auris clade II | 96 | 1 | 9 | 8 | 5 | 7 | 25 | 41 | 66 (68.8) | 27.697 | |||||
| C. auris clade I | 8 | 3 | 1 | 1 | 1 | 2 | 2 (25.0) | 6.727 | ||||||||
| C. pseudohaemulonii | 11 | 1 | 9 | 1 | 0 (0) | 2.000 | ||||||||||
| C. haemulonii | 7 | 2 | 1 | 4 | 4 (57.1) | 40.317 | ||||||||||
| C. haemulonii var. vulnera | 2 | 1 | 1 | 0 (0) | 5.657 | |||||||||||
| AMB | C. auris clade II | 96 | 4 | 13 | 69 | 10 | 0 (0) | 0.461 | ||||||||
| C. auris clade I | 8 | 2 | 6 | 6 (75) | 1.682 | |||||||||||
| C. pseudohaemulonii | 11 | 4 | 1 | 6 | 11 (100) | 4.537 | ||||||||||
| C. haemulonii | 7 | 1 | 1 | 1 | 4 | 6 (85.7) | 5.040 | |||||||||
| C. haemulonii var. vulnera | 2 | 1 | 1 | 1 (50.0) | 1.414 | |||||||||||
| ANI | C. auris clade II | 96 | 17 | 21 | 50 | 4 | 4 | 0 (0) | 0.086 | |||||||
| C. auris clade I | 8 | 1 | 4 | 1 | 2 | 0 (0) | 0.352 | |||||||||
| C. pseudohaemulonii | 11 | 2 | 3 | 6 | 0 (0) | 0.158 | ||||||||||
| C. haemulonii | 7 | 1 | 4 | 2 | 0 (0) | 0.153 | ||||||||||
| C. haemulonii var. vulnera | 2 | 1 | 1 | 0 (0) | 0.042 | |||||||||||
| CAS | C. auris clade II | 96 | 12 | 59 | 24 | 1 | 0 (0) | 0.066 | ||||||||
| C. auris clade I | 8 | 1 | 5 | 2 | 0 (0) | 0.271 | ||||||||||
| C. pseudohaemulonii | 11 | 5 | 1 | 5 | 0 (0) | 0.184 | ||||||||||
| C. haemulonii | 7 | 3 | 3 | 1 | 0 (0) | 0.144 | ||||||||||
| C. haemulonii var. vulnera | 2 | 1 | 1 | 0 (0) | 0.061 | |||||||||||
| MCF | C. auris clade II | 96 | 33 | 55 | 8 | 0 (0) | 0.050 | |||||||||
| C. auris clade I | 8 | 1 | 4 | 3 | 0 (0) | 0.145 | ||||||||||
| C. pseudohaemulonii | 11 | 4 | 3 | 4 | 0 (0) | 0.060 | ||||||||||
| C. haemulonii | 7 | 3 | 3 | 1 | 0 (0) | 0.071 | ||||||||||
| C. haemulonii var. vulnera | 2 | 1 | 1 | 0 (0) | 0.030 | |||||||||||
| Antifungals which tentative MIC breakpoints were not provided by the US CDC | ||||||||||||||||
| VOR | C. auris clade II | 96 | 20 | 7 | 6 | 26 | 25 | 6 | 5 | 1 | NA | 0.190 | ||||
| FR (n = 66) | 1 | 2 | 26 | 25 | 6 | 5 | 1 | NA | 0.435 | |||||||
| FS (n = 30) | 20 | 6 | 4 | NA | 0.031 | |||||||||||
| C. auris clade I | 8 | 5 | 1 | 1 | 1 | NA | 0.080 | |||||||||
| C. pseudohaemulonii | 11 | 10 | 1 | NA | 0.021 | |||||||||||
| C. haemulonii | 7 | 3 | 1 | 1 | 1 | 1 | NA | 0.349 | ||||||||
| C. haem.var. vulnera | 2 | 1 | 1 | NA | 0.042 | |||||||||||
| ITC | C. auris clade II | 96 | 10 | 23 | 11 | 43 | 8 | 1 | NA | 0.140 | ||||||
| FR (n = 66) | 2 | 6 | 7 | 42 | 8 | 1 | NA | 0.212 | ||||||||
| FS (n = 30) | 8 | 17 | 4 | 1 | NA | 0.056 | ||||||||||
| C. auris clade I | 8 | 4 | 1 | 1 | 1 | 1 | NA | 0.112 | ||||||||
| C. pseudohaemulonii | 11 | 9 | 2 | NA | 0.030 | |||||||||||
| C. haemulonii | 7 | 2 | 2 | 1 | 1 | 1 | NA | 0.436 | ||||||||
| C. haem.var. vulnera | 2 | 1 | 1 | NA | 0.042 | |||||||||||
| POS | C. auris clade II | 96 | 34 | 12 | 32 | 15 | 3 | NA | 0.066 | |||||||
| FR (n = 66) | 6 | 10 | 32 | 15 | 3 | NA | 0.118 | |||||||||
| FS (n = 30) | 28 | 2 | NA | 0.018 | ||||||||||||
| C. auris clade I | 8 | 5 | 2 | 1 | NA | 0.047 | ||||||||||
| C. pseudohaemulonii | 11 | 6 | 5 | NA | 0.021 | |||||||||||
| C. haemulonii | 7 | 2 | 2 | 1 | 1 | 1 | NA | 0.164 | ||||||||
| C. haem.var. vulnera | 2 | 1 | 1 | NA | 0.021 | |||||||||||
| 5-FC | C. auris clade II | 96 | 23 | 17 | 40 | 10 | 6 | NA | 0.139 | |||||||
| FR (n = 66) | 18 | 14 | 26 | 5 | 3 | NA | 0.114 | |||||||||
| FS (n = 30) | 4 | 4 | 14 | 5 | 3 | NA | 0.211 | |||||||||
| C. auris clade I | 8 | 1 | 5 | 1 | 1 | NA | 0.085 | |||||||||
| C. pseudohaemulonii | 11 | 11 | NA | 0.060 | ||||||||||||
| C. haemulonii | 7 | 5 | 1 | 1 | NA | 0.081 | ||||||||||
| C. haem.var. vulnera | 2 | 2 | NA | 0.060 | ||||||||||||
| Source | Species/Clade | No. Tested | No. of Isolates Grown (No. with Growth Degree +++/++/+) at: * | ||||
|---|---|---|---|---|---|---|---|
| 30 °C | 37 °C | 40 °C | 42 °C | 45 °C | |||
| Korean collection | C. auris, clade II | 96 | 96 (96/0/0) | 96 (96/0/0) | 95 (85/2/8) | 85 (18/40/27) | 0 (0/0/0) |
| C. auris, clade I | 8 | 8 (8/0/0) | 8 (8/0/0) | 8 (8/0/0) | 8 (6/0/2) | 6 (0/4/2) | |
| C. pseudohaemulonii | 11 | 11 (11/0/0) | 11 (11/0/0) | 0 (0/0/0) | 0 (0/0/0) | 0 (0/0/0) | |
| C. haemulonii | 7 | 7 (7/0/0) | 7 (7/0/0) | 0 (0/0/0) | 0 (0/0/0) | 0 (0/0/0) | |
| C. haemulonii var. vulnera | 2 | 2 (2/0/0) | 2 (2/0/0) | 0 (0/0/0) | 0 (0/0/0) | 0 (0/0/0) | |
| FDA- CDC collection | C. auris, clade II | 1 | 1 (1/0/0) | 1 (1/0/0) | 1 (1/0/0) | 1 (0/0/1) | 0 (0/0/0) |
| C. auris, clade I | 5 | 5 (5/0/0) | 5 (5/0/0) | 5 (5/0/0) | 5 (3/2/0) | 4 (0/1/3) | |
| C. auris, clade III | 2 | 2 (2/0/0) | 2 (2/0/0) | 2 (2/0/0) | 2 (2/0/0) | 2 (0/0/2) | |
| C. auris, clade IV | 2 | 2 (2/0/0) | 2 (2/0/0) | 2 (2/0/0) | 2 (2/0/0) | 2 (0/2/0) | |
| C. auris Isolates | Clade | FLU-AMB-ANI-CAS-MIC Susceptibilities | Amino Acid Substitutions of * | ||
|---|---|---|---|---|---|
| Erg11p | Tac1Ap | Tac1Bp | |||
| Korean collection (source, year, hospital) | |||||
| 1 (Blood, 2018, E) | I | R-S-S-S-S | K143R | None | A640V |
| 2 (Ear, 2019, F) | I | R-S-S-S-S | None | W188C | G91C |
| 3 (Blood, 2022, D) | I | S-S-S-S-S | None | None | None |
| 4 (Blood, 2022, D) | I | S-S-S-S-S | None | None | None |
| 5 (Blood, 2022, D) | I | S-S-S-S-S | None | None | None |
| 6 (Blood, 2022, D) | I | S-S-S-S-S | None | None | None |
| 7 (Blood, 2022, D) | I | S-S-S-S-S | None | None | None |
| 8 (Urine, 2022, D) | I | S-S-S-S-S | None | None | None |
| FDA-CDC collection (AR No) | |||||
| 9 (AR0381) | II | S-S-S-S-S | None | V145E, K713N | K215R |
| 10 (AR0382) | I | S-S-S-S-S | None | None | None |
| 11 (AR0383) | III | R-S-S-S-S | F126L | V13I, S116A, V145E, G149D, D500E, E560D, E565D, S627G, E758G, S762P, A766T | S36L, S89Y, E200K, K215R, K225N, Q226R, I268V, D278V, Q298K, C331S, C334F, L335S, S339A, T346I, Q503R, F580L, Y608H, Y647C, S754N, L774M, M809I, 772-773dle |
| 12 (AR0384) | III | R-S-S-S-S | F126L | Identical to AR0383 | Identical to AR0383 |
| 13 (AR0385) | IV | R-S-S-S-S | Y132F | V13I, S116A, V145E, A288S, D500E, E560D, S762P | S36L, K215R, Q226R, D278V, L328Q, C331S, C334F, L335S, S339A, Y608H, S754N, M809I, 862-866del |
| 14 (AR0386) | IV | R-S-S-S-S | Y132F | Identical to AR0385 | Identical to AR0385 |
| 15 (AR0387) | I | S-S-S-S-S | None | None | None |
| 16 (AR0388) | I | R-S-S-S-S | K143R | None | A640V |
| 17 (AR0389) | I | R-R-S-S-S | Y132F | None | L582I |
| 18 (AR0390) | I | R-S-S-S-S | K143R | None | A640V |
| Amino Acid Substitutions of * | No. of Isolates with Fluconazole MIC (mg/L): | Total No. | ||||||
|---|---|---|---|---|---|---|---|---|
| Erg11p | Tac1Ap | Tac1Bp | ≥64 (FR) | 32 (FR) | 16 (FS) | 8 (FS) | ≤4 (FS) | |
| None | None | F214S | 16 | 11 | 1 | 28 | ||
| None | None | S201Y, L582I | 1 | 3 | 4 | |||
| K143R | None | None | 2 | 1 | 3 | |||
| K143R | None | S427R | 2 | 1 | 3 | |||
| None | None | L582I | 1 | 1 | 2 | |||
| R157S | None | F214S | 2 | 2 | ||||
| None | None | Q526E | 2 | 2 | ||||
| None | None | W198C | 1 | 1 | 2 | |||
| K143R | None | F214S | 1 | 1 | ||||
| None | I783F | F214S | 1 | 1 | ||||
| A298G | None | F214S | 1 | 1 | ||||
| None | None | N393I, D865N | 1 | 1 | ||||
| Q357K | None | L582I | 1 | 1 | ||||
| I466L | None | S192N, S427R | 1 | 1 | ||||
| None | None | R50L | 1 | 1 | ||||
| None | None | R202M, P595L | 1 | 1 | ||||
| None | None | S531P | 1 | 1 | ||||
| E468V | None | S531P | 1 | 1 | ||||
| None | None | P595A | 1 | 1 | ||||
| None | None | A651V | 1 | 1 | ||||
| Y132F | None | None | 1 | 1 | ||||
| None | None | W198L, R505S | 3 | 3 | ||||
| L43H | None | F214S | 1 | 1 | ||||
| None | None | C168F | 1 | 1 | ||||
| None | None | F191V | 1 | 1 | ||||
| None | None | F649I | 1 | 1 | ||||
| None | E281Q | P860T | 1 | 1 | ||||
| None | None | R202M | 1 | 1 | ||||
| None | None | P600S | 1 | 1 | ||||
| None | None | L704V | 1 | 1 | ||||
| None | None | P860T | 1 | 1 | ||||
| R462K | None | None | 1 | 1 | ||||
| None | None | P213L | 1 | 1 | ||||
| None | None | L759S | 1 | 1 | ||||
| None | None | S192N, D845Y | 1 | 1 | ||||
| None | None | None | 1 | 3 | 3 | 14 | 21 | |
| Total | 41 | 25 | 7 | 5 | 18 | 96 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Byun, S.A.; Kwon, Y.J.; Lee, G.Y.; Choi, M.J.; Jeong, S.H.; Kim, D.; Choi, M.H.; Kee, S.-J.; Kim, S.H.; Shin, M.G.; et al. Virulence Traits and Azole Resistance in Korean Candida auris Isolates. J. Fungi 2023, 9, 979. https://doi.org/10.3390/jof9100979
Byun SA, Kwon YJ, Lee GY, Choi MJ, Jeong SH, Kim D, Choi MH, Kee S-J, Kim SH, Shin MG, et al. Virulence Traits and Azole Resistance in Korean Candida auris Isolates. Journal of Fungi. 2023; 9(10):979. https://doi.org/10.3390/jof9100979
Chicago/Turabian StyleByun, Seung A., Yong Jun Kwon, Ga Yeong Lee, Min Ji Choi, Seok Hoon Jeong, Dokyun Kim, Min Hyuk Choi, Seung-Jung Kee, Soo Hyun Kim, Myung Geun Shin, and et al. 2023. "Virulence Traits and Azole Resistance in Korean Candida auris Isolates" Journal of Fungi 9, no. 10: 979. https://doi.org/10.3390/jof9100979
APA StyleByun, S. A., Kwon, Y. J., Lee, G. Y., Choi, M. J., Jeong, S. H., Kim, D., Choi, M. H., Kee, S.-J., Kim, S. H., Shin, M. G., Won, E. J., & Shin, J. H. (2023). Virulence Traits and Azole Resistance in Korean Candida auris Isolates. Journal of Fungi, 9(10), 979. https://doi.org/10.3390/jof9100979