
Plant-Associated Neoscytalidium dimidiatum—Taxonomy, Host Range, Epidemiology, Virulence, and Management Strategies: A Comprehensive Review


Abstract
:1. Introduction
2. Taxonomy and Classification of Neoscytalidium dimidiatum
2.1. Historical Changes in Neoscytalidium dimidiatum Taxonomy
2.2. Significance of Resolving Neoscytalidium Species Concepts
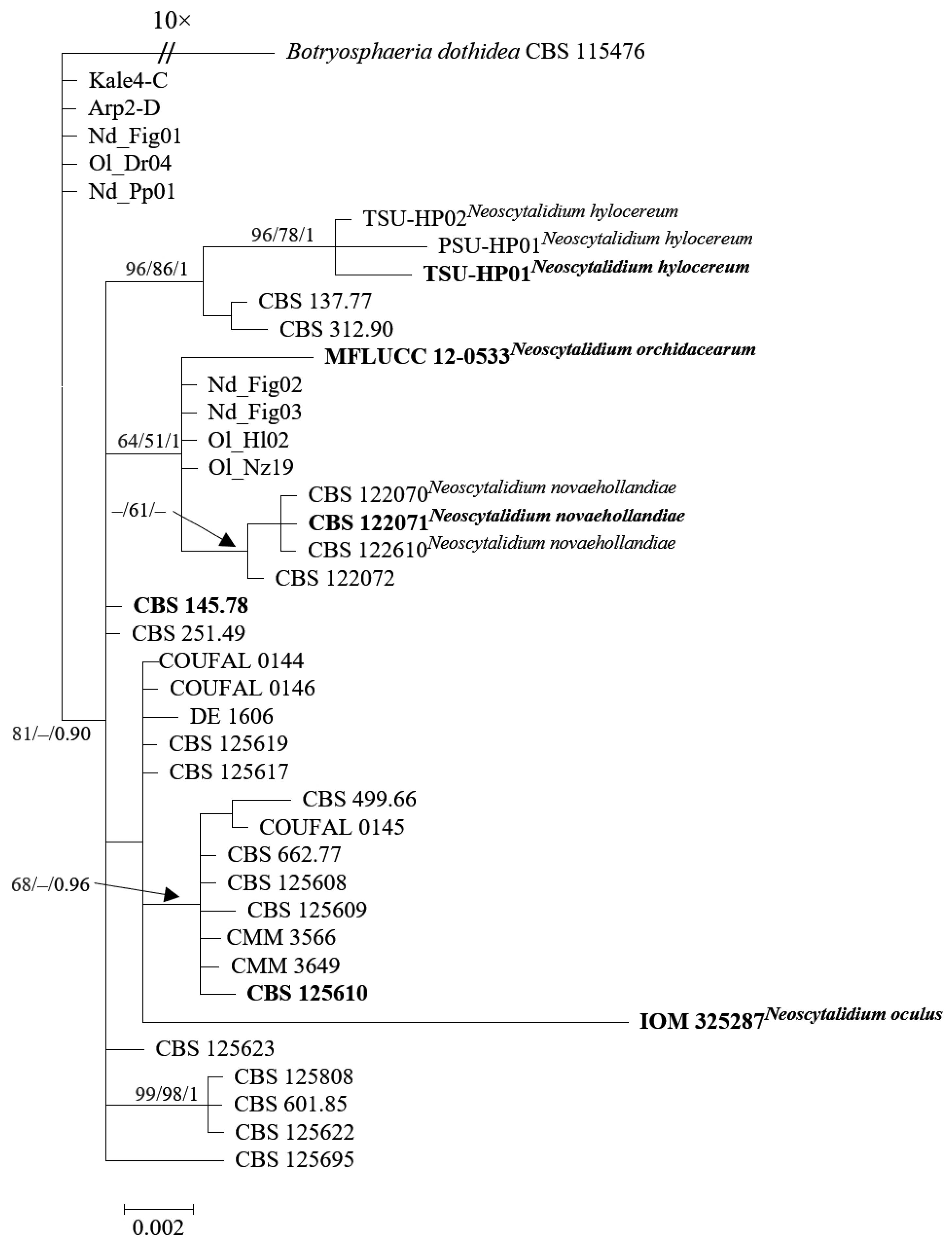
2.3. Molecular Evidence for Synonymy and Intraspecific Variation in Neoscytalidium dimidiatum
2.4. Current Species Name
2.5. Classification
2.6. Synonyms and Basionyms
- 1.
- Torula dimidiata Penz. [59].
- 2.
- Hendersonula toruloidea Nattrass [56].
- 3.
- Exosporina fawcettii E.E. Wilson [60].
- 4.
- Scytalidium hyalinum C.K. Campb. & J.L. Mulder [61].
- 5.
- Scytalidium dimidiatum (Penz.) B. Sutton & Dyko [57].
- 6.
- Fusicoccum dimidiatum (Penz.) D.F. Farr [58].
- 7.
- Neoscytalidium dimidiatum (Penz.) Crous & Slippers [50].
- 8.
- Neoscytalidium novaehollandiae Pavlic, T.I. Burgess & M.J. Wingf. [51].
- 9.
- Neoscytalidium dimidiatum var. hyalinum (C.K. Camp. & J.L. Mulder) Madrid, Cano, Stchigel & Guarro [64].
- 10.
- Neoscytalidium hyalinum (C.K. Campb. & J.L. Mulder) A.J.L. Phillips, Groenewald & Crous [52].
- 11.
- Neoscytalidium orchidacearum S. K. Huang, N. Tangthirasunun, J. C. Kang & K. D. Hyde [53].
- 12.
- Neoscytalidium oculus (Neoscytalidium oculi in MycoBank) J. Mena, Raymundo & Bautista-de-Lucio (the taxonomic status of this species was consolidated as a synonym in the present investigation).
- 13.
- Neoscytalidium hylocereum Kheawleng, Intaraa-nun & Rodkaew (the taxonomic status of this species was consolidated as a synonym in the present investigation).
2.7. Rejected Synonyms
- 1.
- Dothiorella mangiferae Syd. & P. Syd. (1916) [57].
- 2.
- Fusicoccum eucalypti Sousa da Câmara (1929) [57].
- 3.
- Hendersonula cypria Nattrass (1937) [57].
- 4.
- Hendersonula agathidis H.E. Young (1948) [57].
- 5.
- Nattrassia mangiferae (Syd. & P. Syd.) B. Sutton & Dyko [57].
- 6.
- Fusicoccum mangiferae (Syd. & P. Syd.) Johnson, Slippers & M.J. Wingf. [62].
- 7.
- Scytalidium lignicola Pesante [50].
3. Identification
4. Host Range, Symptoms, and Geographic Distribution of Neoscytalidium dimidiatum
4.1. Host Range and Geographical Distribution Diversity of Neoscytalidium dimidiatum

{kind=link}
{kind=link}
| Plant Species Family | Host Species | Common Host Name | Identified Species Name | Target Loci for IDENTIFICATION | Koch’s Postulates | Symptoms | Country | Continent | References |
|---|---|---|---|---|---|---|---|---|---|
| Acanthaceae | Avicennia marina | White mangrove | Nd | ITS | + | Canker and dieback | Iran | Asia | [77] |
| Amaryllidaceae | Hymenocallis littoralis | White spider lily | Nd | ITS | + | Leaf blight | Malaysia | Asia | [78] |
| Clivia miniata | Natal lily | Nd | ITS, tef1 | + | Leaf blight | Iran | Asia | [79] | |
| Anacardiaceae | Anacardium occidentale | Cashew | Nh | ITS, tef1 | + | Dieback and stem and branch cankers | Brazil | South America | [72] |
| Mangifera indica | Mango | Ht | − | + | Leaf spot | India | Asia | [80] | |
| Ht | − | + | Dieback | Niger | Africa | [81] | |||
| Ef | − | NA | Herbarium specimen records | South Africa | Africa | [57] | |||
| Ht | − | NA | NA | Brazil | South America | [82] | |||
| Fd | ITS, tub2 | − | NA | US—California | North America | [58] | |||
| Nd and Nn | ITS, tef1 | + | Dieback | Australia | Australia | [35] | |||
| Nd and Nn | ITS, tef1 | + | Canker | Australia | Australia | [83] | |||
| Nd | ITS, tef1 | + | Dieback and stem-end rot | Brazil | South America | [84] | |||
| Nh | ITS, tef1 | + | Dieback and stem and branch cankers | Brazil | South America | [72] | |||
| Pistacia vera | Pistachio | Nd | ITS, tef1, tub2 | + | Endophytic | Iran | Asia | [85] | |
| Nd | ITS, LSU | + | Canker, shoot blight, and root rot | Turkey | Asia | [42] | |||
| Nn | ITS, tef1 | + | Dieback | Turkey | Asia | [86] | |||
| Rhus typhina | Staghorn sumac | Td | ITS, tub2 | NA | NA | US—West Virginia | North America | [87,88] | |
| Apocynaceae | Nerium oleander | Oleander | Nn | ITS, LSU, tef1 | − | Sooty canker | Iran | Asia | [40] |
| Araceae | Thaumatophyllum bipinnatifidum (Philodendron bipinnatifidum) | Split-leaf philodendron | Ht | − | NA | NA | India | Asia | [89] |
| Araliaceae | Meryta denhamii | Meryta | Nd | ITS, tef1, tub2 | + | Branch canker and dieback | Italy | Europe | [90] |
| Araucariaceae | Agathis robusta (A.palmerstoni) | Kauri | Ef | − | NA | Herbarium specimen records | Australia | Australia | [57] |
| Araucaria sp. | Chilean pine | Ht | − | NA | NA | Malaysia | Asia | [91] | |
| Asparagaceae | Agave sp. | Century plant | Ht | − | NA | NA | Guinea | Africa | [92] |
| Agave americana | Century plant | Ef | − | NA | Herbarium specimen records | India | Asia | [57] | |
| Agave sisalana | Sisal | Nd | ITS, tef1, tub2 | + | Black leaf spot | China | Asia | [93] | |
| Furcraea foetida (F. gigantea) | Green aloe | Ht | − | NA | NA | Malaysia | Asia | [94] | |
| Sansevieria hyacinthoides (S. guineensis) | African Bowstring Hemp | Ht | − | NA | NA | Guinea | Africa | [92] | |
| Sansevieria trifasciata (Dracaena trifasciata) | Dracaena | Nd | ITS | + | Leaf blight | Malaysia | Asia | [95] | |
| Nd | ITS, tef1 | + | Leaf blight | Brazil | South America | [96] | |||
| Asphodeloideae | Aloidendron dichotomum | Quiver tree | Nd | ITS, LSU, tef1, tub2, chs−1 | − | Epiphyte on stems | South Africa | Africa | [97] |
| Berberidaceae | Berberis vulgaris | Barberry | Nd | ITS, tef1, tub2, act | + | Canker and dieback | Iran | Asia | [98] |
| Betulaceae | Alnus glutinosa | Common alder | Nn | ITS, tef1 | + | Branch and trunk cankers | Iran | Asia | [1] |
| Carpinus betulus | Common hornbeam | Nn | ITS, tef1 | + | Branch and trunk cankers | Iran | Asia | [1] | |
| Boraginaceae | Cordia myxa | Assyrian plum | Nn | ITS, LSU, tef1 | − | Canker | Iran | Asia | [40] |
| Bromeliaceae | Ananas comosus (A. sativa) | Pineapple | Ht | − | NA | Leaf spot; fruit rot | Malaysia | Asia | [94] |
| Ht | − | NA | Leaf spot; fruit rot | Tanzania | Africa | [99] | |||
| Ef | − | NA | Herbarium specimen records | Sierra Leone | Africa | [57] | |||
| Ef | − | NA | Herbarium specimen records | Solomon Islands—Rendova | Oceania | [57] | |||
| Nd | ITS, LSU | + | Postharvest stem end rot | Malaysia | Asia | [100] | |||
| Cactaceae | Nopalea cochenillifera | Pickly pear cactus | Nh | ITS, tef1, tub2 | + | Squamous cladode spots | Brazil | South America | [101] |
| Selenicereus (Hylocereus) undatus | Pitahaya (white-fleshed dragon fruit) | Nd | ITS | + | Stem canker | Taiwan | Asia | [5] | |
| Nd | ITS | + | Brown stem-spot-forming canker | China | Asia | [6] | |||
| Nd | ITS, tub2 | + | Internal black rot | Israel | Asia | [8] | |||
| Nd | ITS | + | Canker, internal brown rot | China | Asia | [7,102] | |||
| Nd | ITS | + | Stem and fruit canker | US—Florida | North America | [10] | |||
| Nd | ITS, tub2 | + | Stem and fruit canker | US—Florida | North America | [11] | |||
| Selenicereus (Hylocereus) undatus × S. polyrhizus | Red-fleshed dragon fruit | Nd | ITS, tub2 | + | Stem and fruit canker | US—Florida | North America | [11] | |
| Selenicereus (Hylocereus) polyrhizus | Red-fleshed dragon fruit | Nd | ITS | + | Stem canker | Taiwan | Asia | [5] | |
| Nd | ITS | + | Stem canker | Malaysia | Asia | [9] | |||
| Nd | ITS | + | Stem canker | China | Asia | [12] | |||
| Nd | ITS, LSU, tub2 | + | Stem canker | Thailand | Asia | [15] | |||
| Selenicereus (Hylocereus) megalanthus | Yellow pitahaya (dragon fruit) | Nd | ITS, tef1, tub2 | + | Stem canker | Malaysia | Asia | [17] | |
| Nd | ITS, tef1, tub2 | + | Stem canker | Ecuador | South America | [18] | |||
| Selenicereus (Hylocereus) monacanthus | Dragon fruit | Nd | tub2 | + | Stem and fruit canker | Philippines | Asia | [13] | |
| Selenicereus spp. (Hylocereus spp.) | Dragon fruit | Nd | ITS, tef1 | + | Stem canker | Puerto Rico | North America | [14] | |
| Nd | ITS, tef1, tub2 | + | Stem canker | India | Asia | [16] | |||
| Casuarinaceae | Casuarina sp. | Casuarina | Ht | − | NA | NA | Pakistan | Asia | [103] |
| Combretaceae | Conocarpus erectus | Buttonwood or button mangrove | Nn | ITS, LSU, tef1 | + | Lamination of the trunk bark | Iran | Asia | [40] |
| Convolvulaceae | Ipomoea batatas | Sweet potato | Ht | − | NA | NA | Malaysia | Asia | [94,104] |
| Nd | ITS, tef1 | + | Root rot | Brazil | South America | [105] | |||
| Nd | ITS, tef1 | + | Root and stem rot | Brazil | South America | [106] | |||
| Cucurbitaceae | Cucumis melo | Melon | Nh | ITS | + | Fruit rot | Iran | Asia | [107] |
| Cupressaceae | Cupressus sempervirens | Mediterranean cypress | Nn | ITS, LSU, tef1 | − | Canker and dieback | Iran | Asia | [40] |
| Sequoiadendron giganteum (Sequoia gigantea) | Giant redwood, giant sequoia | Ht | − | NA | NA | US—California | North America | [108] | |
| Dioscoreaceae | Dioscorea esculenta | Lesser yam | Nd | ITS, tef1, tub2 | + | Dieback | China | Asia | [109] |
| Dioscorea rotundata | Yam | Nd | ITS, tub2 | − | Tuber dry rot | Colombia | South America | [110] | |
| Ebenaceae | Diospyros kaki | Japanese persimmon | Nn | ITS, tef1 | + | Branch dieback | Turkey | Asia | [111] |
| Ericaceae | Arbutus menziesii | Madrone | Ht | − | NA | NA | US—Washington | North America | [112,113] |
| Ht | − | NA | NA | Canada | North America | [114] | |||
| Ht | − | NA | Canker | US—California | North America | [115] | |||
| Arbutus unedo | Strawberry tree | Ht | − | + | Leaf spotting and defoliation | Greece | Europe | [116] | |
| Euphorbiaceae | Jatropha curcas | Physic nut, a biofuel plant | Nh | ITS | + | Collar and root rot | Brazil | South America | [117] |
| Nd | ITS, tef1, tub2 | + | Collar and root rot | Brazil | South America | [118] | |||
| Manihot esculenta (M. utilissima) | Cassava | Ht | − | NA | NA | Ghana | Africa | [119] | |
| Ht | − | NA | NA | Kenya | Africa | [120] | |||
| Ef | − | NA | Herbarium specimen records | Ghana | Africa | [57] | |||
| Nh | ITS, tef1, tub2 | + | Black root rot | Brazil | South America | [121] | |||
| Nd | ITS, tef1, rpb2 | + | Black root rot and stem cutting dry rot | Brazil | South America | [122] | |||
| Nd | ITS, tef1 | + | Black stem and root rot | Thailand | Asia | [123] | |||
| Fabaceae | Acacia auriculiformis | Earleaf acacia | Ht | − | NA | NA | India | Asia | [89,124] |
| Ef | − | NA | Herbarium specimen records | India | Asia | [57] | |||
| Acacia melanoxylon | Australian blackwood | Ht | − | NA | NA | India | Asia | [124,125] | |
| Acacia synchronicia | Bardi bush | Nn | ITS, tef1 | − | Asymptomatic branches (sapwood) | Australia | Australia | [51] | |
| Nh | ITS, tef1 | + | Endophyte as a potential pathogen | Australia | Australia | [126] | |||
| Albizia lebbeck | Siris tree | Sd | − | − | Dieback | Oman | Asia | [34] | |
| Nn | ITS, LSU, tef1 | − | Sooty canker | Iran | Asia | [40] | |||
| Bauhinia purpurea | Orchid tree | Nn | ITS, LSU, tef1 | + | Lamination of the trunk bark | Iran | Asia | [40] | |
| Cassia fistula | Golden shower tree | Nn | ITS, LSU, tef1 | − | Canker | Iran | Asia | [40] | |
| Cassia floribunda | Arsenic bush | Nn | ITS, LSU, tef1 | − | Canker | Iran | Asia | [40] | |
| Cicer arietinum | Chickpea | Nd | ITS, tef1, tub2 | + | Blight and root rot | Turkey | Asia | [49] | |
| Crotalaria medicaginea | Rattlepods | Nn | ITS, tef1 | − | Asymptomatic branches (sapwood) | Australia | Australia | [51] | |
| Nd | ITS, tef1 | + | Endophyte as a potential pathogen | Australia | Australia | [126] | |||
| Delonix regia | Royal poinciana | Sd | − | − | Dieback | Oman | Asia | [34] | |
| Nd | ITS, LSU, tef1, tub2 | + | Stem canker | United Arab Emirates | Asia | [127] | |||
| Lysiphyllum cunninghamii | Kimberley bauhinia or jigal tree | Nn | ITS, tef1 | + | Endophyte as a potential pathogen | Australia | Australia | [126] | |
| Parkinsonia aculeata | Palo verde | Nn | − | − | Dieback (when used as a bipherbicide) | Australia | Australia | [128] | |
| Peltophorum petrocarpum | Copperpod | Sd | − | − | Dieback | Oman | Asia | [34] | |
| Fagaceae | Castanea sativa | Sweet chestnut | Ef | − | NA | Canker | US—California | North America | [129] |
| Ht | − | NA | Canker | US—California | North America | [115] | |||
| Fagus orientalis | Oriental beech | Nn | ITS, tef1 | + | Branch and trunk cankers | Iran | Asia | [1] | |
| Quercus brantii | Persian oak | Nn | ITS, LSU, tef1 | + | Dieback | Iran | Asia | [74] | |
| Nd | ITS, LSU, SSU | + | Decline and sooty canker | Iran | Asia | [130] | |||
| Iridaceae | Gladiolus sp. | Gladiolus | Ht | − | NA | NA | US—California | North America | [115] |
| Juglandaceae | Juglans californica | California black walnut | Ht | − | NA | On twigs | US—California | North America | [131] |
| Juglans regia | English walnut | Ef | − | NA | Branch wilt | US—California | North America | [60,129,132] | |
| Ht | − | NA | NA | US—California | North America | [115,133] | |||
| Fd | ITS, tub2 | − | NA | US—California | North America | [58,134] | |||
| Nd | ITS, tef1, tub2 | + | Black canker and death of graft union | US—California | North America | [135,136] | |||
| Nd | ITS, LSU, tef1 | + | Black canker, root rot, decline | Turkey | Asia | [137,138] | |||
| Nd | Decline | Iran | Asia | [139] | |||||
| Nn | ITS, LSU, tef1 | − | Sooty canker | Iran | Asia | [40] | |||
| Lamiaceae | Lavandula angustifolia | Lavender | Nd | ITS, tef1 | + | Foliar and stem blight | Turkey | Asia | [140] |
| Melissa officinalis | Lemon balm | Nd | ITS, tef1 | + | Blight | Turkey | Asia | [141] | |
| Origanum onites | Turkish oregano | Nd | ITS, tef1 | + | Leaf blight | Turkey | Asia | [142] | |
| Salvia officinalis | Common sage | Nn | ITS, tef1 | + | Root rot and foliar blight | Turkey | Asia | [143] | |
| Lythraceae | Punica granatum | Pomegranate | Nd | ITS, tef1, tub2, act | + | Necrotic wood tissues | Iran | Asia | [144] |
| Nn | ITS, LSU, tef1 | − | Sooty canker | Iran | Asia | [40] | |||
| Malvaceae | Adansonia gibbosa | Baobab | Nn | ITS, tef1 | − | Asymptomatic branches (sapwood) | Australia | Australia | [51] |
| Adansonia gregorii | Boab | Nh | ITS, tef1 | + | Endophyte as a potential pathogen, canker | Australia | Australia | [126] | |
| Hibiscus rosa-sinensis | Chinese hibiscus | Nn | ITS, LSU, tef1 | − | Canker and dieback | Iran | Asia | [40] | |
| Thespesia populnea | Portia tree | Sd | − | − | Dieback | Oman | Asia | [34] | |
| Meliaceae | Azadirachta indica | Neem | Nh | ITS, tef1, tub2, act | + | Decline | Iran | Asia | [145] |
| Melia azedarach | Chinaberry | Ht | − | − | NA | Pakistan | Asia | [146] | |
| Nd | ITS, LSU, tub2 | − | Canker and dieback | Iraq | Asia | [147] | |||
| Moraceae | Ficus benghalensis | Indian banyan | Sd | − | − | Dieback | Oman | Asia | [34] |
| Nd | Sooty canker | Iran | Asia | [39] | |||||
| Nn | ITS, LSU, tef1 | − | Dieback and sooty canker | Iran | Asia | [40] | |||
| Ficus benjamina | Weeping fig | Nd | ITS, LSU, rpb2 | Branch dieback | Mexico | North America | [36] | ||
| Nd | ITS, LSU, SSU | + | Sooty canker | Egypt | Africa | [37] | |||
| Ficus carica | Common fig | Ht | − | − | NA | Cyprus | Europe | [148] | |
| Ht | − | NA | NA | US—California | North America | [115,131,149] | |||
| Ht | − | NA | Herbarium specimen records | Cyprus | Europe | [57] | |||
| Sd | − | − | Dieback | Oman | Asia | [34] | |||
| Nd | ITS, tef1 | + | Dieback | Australia | Australia | [35] | |||
| Nd | ITS, tef1, tub2 | + | Dieback and canker | Turkey | Asia | [38] | |||
| Nd | − | Limb dieback | US—California | North America | [41] | ||||
| Ficus nitida | Chinese banyan | Nd | ITS, LSU, SSU | + | Sooty canker | Egypt | Africa | [37] | |
| Ficus religiosa | Bodhi tree | Nn | ITS, LSU, tef1 | + | Dieback and sooty canker | Iran | Asia | [40] | |
| Ficus retusa | Banyan tree | Sd | − | − | Dieback | Oman | Asia | [34] | |
| Morus alba | White mulberry | Ef | − | NA | Herbarium specimen records | Pakistan | Asia | [57] | |
| Ef | − | NA | Herbarium specimen records | US | North America | [57] | |||
| Ht | − | − | NA | Pakistan | Asia | [146] | |||
| Nn | ITS, LSU, tef1 | + | Shoot and branch deaths | Turkey | Asia | [150] | |||
| Morus bombycis (Morus australis) | Korean mulberry | Nn | ITS, LSU, tef1 | + | Branch necrosis | Turkey | Asia | [150] | |
| Morus nigra | Black mulberry | Nn | ITS, LSU, tef1 | + | Shoot and branch deaths | Turkey | Asia | [150] | |
| Nn | ITS, LSU, tef1 | + | Sooty canker | Iran | Asia | [40] | |||
| Musaceae | Musa nana | Banana (dwarf) | Ht | − | − | Tip rot | Jamaica | North America | [151] |
| Musa spp. | Banana | Ht | − | − | Tip rot | Hawaii | North America | [88,152] | |
| Musa acuminata | Banana | Ht | − | − | Leaf spot | Hawaii | North America | [153] | |
| Myrtaceae | Callistemon viminalis | Weeping bottlebrush | Nn | ITS, LSU, tef1 | − | Canker | Iran | Asia | [40] |
| Eucalyptus camaldulensis | River red gum | Nd | ITS | + | Sooty canker | Iraq | Asia | [154] | |
| Nn | ITS, LSU, tef1 | + | Dieback, lamination of the trunk bark, and sooty canker | Iran | Asia | [40] | |||
| Eucalyptus sp. | Eucalyptus | Ef | − | NA | Herbarium specimen records | Portugal | Europe | [57] | |
| Nn | ITS, tef1 | + | Endophyte as a potential pathogen | Australia | Australia | [126] | |||
| Eucalyptus spp. | Eucalyptus | Nd | ITS, LSU, tub2 | − | Canker and dieback | Iraq | Asia | [147] | |
| Psidium guajava | Guava | Ht | − | NA | NA | India | Asia | [104] | |
| Nh | ITS, tef1 | + | Dieback and stem and branch cankers | Brazil | South America | [72] | |||
| Nd | ITS, tef1 | + | Postharvest fruit rot | Malaysia | Asia | [155] | |||
| Syzygium cumini | Java plum | Nh | ITS, tef1, tub2, act | + | Cankers and wedge-shaped wood necrosis | Iran | Asia | [156] | |
| Nh | ITS, tef1, tub2, act | + | Asymptomatic wood tissue | Iran | Asia | [156] | |||
| Nn | ITS, LSU, tef1 | + | Lamination of the trunk bark | Iran | Asia | [40] | |||
| Oleaceae | Olea europaea | Olive | Nd | ITS, tef1, tub2 | + | Canker and leaf scorch | Turkey | Asia | [76] |
| Orchidaceae | Arachnis sp. | Scorpion orchid | Ht | − | − | NA | Malaysia | Asia | [157] |
| NA | Orchid | No | ITS, LSU | − | From a fallen orchid leaf | Thailand | Asia | [53] | |
| Cattleya lueddemanniana var. lueddemanniana | Cattleya orchid | No | ITS, LSU | + | Leaf spot | Thailand | Asia | [158] | |
| Cattleya × hybrid | Orchids | Nd | ITS | + | Leaf blight | Taiwan | Asia | [159] | |
| Pinaceae | Picea pungens | Blue spruce | Nd | ITS, tef1, tub2 | + | Needle blight | Turkey | Asia | [160] |
| Pinus eldarica | Afghan pine | Nd | ITS, LSU | + | Shoot and needle blight | Turkey | Asia | [27] | |
| Nn | ITS, LSU, tef1 | + | Dieback | Iran | Asia | [28] | |||
| Pinus nigra | European black pine | Nd | ITS, LSU | + | Shoot and needle blight | Turkey | Asia | [27] | |
| Pinus sylvestris | Scots pine | Nd | ITS, LSU | + | Shoot and needle blight | Turkey | Asia | [27] | |
| Proteaceae | Grevillia agrifolia | Blue grevillea | Nn | ITS, tef1 | − | Asymptomatic branches (sapwood) | Australia | Australia | [51] |
| Nn | ITS, tef1 | + | Endophyte as a potential pathogen | Australia | Australia | [126] | |||
| Rhamnaceae | Ziziphus spina-christi | Christ’s thorn jujube | Nn | ITS, LSU, tef1 | − | Canker | Iran | Asia | [40] |
| Rhizophoraceae | Rhizophora mucronata | Red mangrove | Nd | ITS | + | Canker and dieback | Iran | Asia | [77] |
| Rosaceae | Malus domestica (M. pumila) | Apple | Ht | − | NA | Gummosis and dieback | Egypt | Africa | [56] |
| Ht | − | NA | NA | India | Asia | [89] | |||
| Ef | − | NA | Herbarium specimen records | India | Asia | [57] | |||
| Ef | − | NA | Herbarium specimen records | Iraq | Asia | [57] | |||
| Nd | Branch canker | Iran | Asia | [161] | |||||
| Nd | ITS, tef1 | + | Branch dieback and canker | Turkey | Asia | [32] | |||
| Nd | Cankers | China | Asia | [162] | |||||
| Prunus armeniaca | Apricot | Ht | − | NA | Gummosis and dieback | Egypt | Africa | [56] | |
| Ht | − | − | NA | Cyprus | Europe | [148] | |||
| Ht | − | NA | Herbarium specimen records | Cyprus | Europe | [57] | |||
| Ht | − | NA | Canker | US—California | North America | [115,131] | |||
| Nd | ITS, LSU, tef1, tub2 | + | Shoot blight, branch dieback, and canker | Turkey | Asia | [31] | |||
| Prunus avium | Cherry | Nn | ITS, tef1 | + | Canker and branch dieback | Turkey | Asia | [163] | |
| Prunus domestica | Plum | Ht | − | NA | Gummosis and dieback | Egypt | Africa | [56] | |
| Nd | ITS | + | Decline and dieback | Tunisia | Africa | [29] | |||
| Nn | ITS, tef1 | + | Branch dieback and stem cankers | Turkey | Asia | [164] | |||
| Prunus dulcis (P. amygdalus) | Almond | Ht | − | + | Secondary canker infection | US—California | North America | [165] | |
| Ht | − | NA | Canker | US—California | North America | [115,131] | |||
| Nd | ITS, tef1, tub2 | + | Trunk and branch cankers, spur and shoot blight, fruit rot | US—California | North America | [30] | |||
| Nn | ITS, tef1 | + | Stem canker and branch dieback | Turkey | Asia | [32] | |||
| Nd | ITS, tef1, tub2, GPD | + | Trunk and branch cankers | US—California | North America | [33] | |||
| Prunus persica | Peach | Ht | − | − | Canker | US—California | North America | [115,131] | |
| Nn | ITS, LSU, tef1 | − | Sooty canker | Iran | Asia | [40] | |||
| Prunus sp. | Prunus | Fd | ITS, tef1, tub2 | + | NA | Egypt | Africa | [58,134,166] | |
| Pyrus communis | Pear | Nn | ITS, LSU, tef1 | + | Shoot blight and branch canker | Turkey | Asia | [167] | |
| Rutaceae | Citrus aurantifolia | Acid lime | Ht | − | NA | NA | US—California | North America | [115,131] |
| Nd | ITS | + | Root rot | Oman | Asia | [46] | |||
| Nh | ITS, tef1, tub2 | + | Canker and dieback | Iran | Asia | [168] | |||
| Nn | ITS, LSU, tef1 | − | Canker and dieback | Iran | Asia | [40] | |||
| Citrus clementina | Clementine | Nd | ITS, tub2 | + | Shoot blight | Jordan | Asia | [169] | |
| Citrus latifolia | Persian lime | Ht | − | NA | NA | US—California | North America | [115,131] | |
| Citrus limetta | Sweet limetta | Nh | ITS, tef1, tub2 | + | Canker and dieback | Iran | Asia | [168] | |
| Nn | ITS, LSU, tef1 | − | Dieback | Iran | Asia | [40] | |||
| Citrus limettioides | Sweet lime | Nd | ITS | + | Root rot | Oman | Asia | [46] | |
| Citrus limon (C. limonium) | Lemon | Ht | − | − | NA | Cyprus | Europe | [148] | |
| Ef | − | NA | NA | US—California | North America | [129] | |||
| Ht | − | NA | NA | US—California | North America | [115,131] | |||
| Nh | ITS, tef1, tub2 | + | Branch canker | US—California | North America | [20] | |||
| Citrus maxima (C. grandis) | Pomelo | Ht | − | NA | NA | US—California | North America | [131] | |
| Ht | − | NA | NA | US—California | North America | [115] | |||
| Nd | ITS, tub2 | + | Shoot blight | Jordan | Asia | [169] | |||
| Citrus meyerii | Meyer lemon | Ht | − | NA | NA | US—California | North America | [131] | |
| Citrus paradisi | Grapefruit | Ht | − | NA | Canker, dieback | US—California | North America | [115,131,170] | |
| Ef | − | NA | NA | US—California | North America | [129] | |||
| Nd | ITS, tef1, tub2 | + | Bot gummosis | US—California | North America | [171] | |||
| Nh | ITS, tef1, tub2 | + | Branch canker | US—California | North America | [20] | |||
| Nd | ITS, tub2 | + | Shoot blight | Jordan | Asia | [169] | |||
| Citrus reticulata | Mandarin | Ht | − | NA | NA | US—California | North America | [115,131] | |
| Citrus sinensis | Sweet orange | Ht | − | NA | NA | South Africa | Africa | [172] | |
| Ht | − | NA | NA | US—California | North America | [115,131] | |||
| Ef | − | NA | Herbarium specimen records | Pakistan | Asia | [57] | |||
| Nd | ITS | + | Blight, canker, and gummosis | Italy | Europe | [19] | |||
| Nn | ITS, LSU, tef1 | − | Dieback | Iran | Asia | [40] | |||
| Citrus sp. | Citrus | Td | − | NA | NA | US—California | North America | [129] | |
| Ht | − | NA | NA | US—California | North America | [115,129] | |||
| Citrus tangelo | Tangelo | Ht | − | NA | NA | US—California | North America | [115,131] | |
| Salicaceae | Populus alba | Silver poplar | Ht | − | − | NA | Cyprus | Europe | [148] |
| Ht | − | NA | Herbarium specimen records | Cyprus | Europe | [57] | |||
| Populus fremontii | Frémont’s cottonwood | Ht | − | NA | NA | US—California | North America | [173] | |
| Populus nigra | Black poplar | Nh | tef1 | + | Decline | Iran | Asia | [43] | |
| Salix alba | White willow | Nh | tef1 | + | Decline, irregular and central wood necrosis | Iran | Asia | [43] | |
| Nd | ITS, LSU | + | Dieback, shoot blight, and branch canker | Turkey | Asia | [44] | |||
| Solanaceae | Capsicum annuum | Pepper | Ht | − | NA | NA | Tanzania | Africa | [99] |
| Solanum lycopersicum | Tomato | Nd | ITS, LSU, tef1 | + | Blight and root rot | Turkey | Asia | [47] | |
| Tomato | Nn | ITS, LSU, tef1 | + | Stem blight | Turkey | Asia | [174] | ||
| Solanum tuberosum | Potato | Nd | ITS, LSU, tef1 | + | Tuber rot | Turkey | Asia | [175] | |
| Ulmaceae | Ulmus sp. | Elm | Nh | ITS, tef1 | + | Decline | Iran | Asia | [176] |
| Vitaceae | Vitis vinifera | Grapevine | Ht | − | NA | Drying | India | Asia | [89,177,178] |
| Ht | − | NA | Branch wilt | Iraq | Asia | [179] | |||
| Nd | ITS | + | Dieback | Iraq | Asia | [21] | |||
| Nd | ITS, tub2 | + | Wood canker and decline | US—California | North America | [22] | |||
| Nh | ITS, tef1 | + | Dieback | Brazil | South America | [23] | |||
| Nd | ITS, LSU, tef1, tub2 | + | Canker and dieback | Turkey | Asia | [24] | |||
| Nn | ITS, tef1 | + | Wood canker | Turkey | Asia | [25] | |||
| Nh | ITS | + | Wood necrosis | Iran | Asia | [26] |
4.2. Diversity of Host Responses and Geographic Distribution Patterns of Neoscytalidium dimidiatum across Plant Families and Countries
5. Understanding the Epidemiological Aspects of Neoscytalidium dimidiatum
5.1. Life Cycle of Neoscytalidium dimidiatum
5.2. Host Infection in a Unique Context of Dragon Fruit Canker
5.3. Factors Influencing Neoscytalidium dimidiatum Infection
5.3.1. Factors Influencing Host Response
5.3.2. Climatic and Edaphic Factors
5.4. Mode of Transmission
6. Insights into the Virulence and Genomic Characteristics of Neoscytalidium dimidiatum
6.1. Host–Pathogen Interaction at Molecular Level
6.2. Neoscytalidium dimidiatum Virulence in Diverse Host Plants
7. Management Strategies for Neoscytalidium dimidiatum
7.1. Antifungal Activity of Different Applications
7.1.1. Fungicides
7.1.2. Essential Oils and Biopesticides
7.1.3. Biocontrol Agents
7.1.4. Nanoparticles
7.2. Strategies for Control
8. Future Challenges
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kazemzadeh, C.M.; Mohammadi, H.; Khodaparast, S.A. Diversity and pathogenicity of Botryosphaeriaceae species on forest trees in the north of Iran. Eur. J. For. Res. 2019, 138, 685–704. [Google Scholar] [CrossRef]
- Zhang, W.; Groenewald, J.Z.; Lombard, L.; Schumacher, R.K.; Phillips, A.J.L.; Crous, P.W. Evaluating species in Botryosphaeriales. Persoonia 2021, 46, 63–115. [Google Scholar] [CrossRef] [PubMed]
- Heidari, S.; Gheisari, M.; Abastabar, M.; Pourabdollah, M.; Mirenayat, M.S.; Basharzad, N.; Seifi, S.; Tavakoli, M.; Jafarzadeh, J.; Ansari, S.; et al. Genotyping and ın vitro antifungal susceptibility profile of Neoscytalidium species isolates from respiratory tract. Mycopathologia 2021, 186, 833–845. [Google Scholar] [CrossRef] [PubMed]
- Dolatabadi, S.; Nasirharandi, S.; Pourahmad, M.; Ahmadikia, K.; Mokhtari, M.; Najafzadeh, M.J.; Mohammadi, R. Phaeohyphomycosis caused by Neoscytalidium dimidiatum in a COVID-19 patient. Rev. Iberoam. Micol. 2022, 39, 72–76. [Google Scholar] [CrossRef]
- Chuang, M.F.; Ni, H.F.; Yang, H.R.; Shu, S.L.; Lai, S.Y.; Jiang, Y.L. First report of stem canker disease of pitaya (Hylocereus undatus and H. polyrhizus) caused by Neoscytalidium dimidiatum in Taiwan. Plant Dis. 2012, 96, 906. [Google Scholar] [CrossRef]
- Lan, G.B.; He, Z.F.; Xi, P.G.; Jiang, Z.D. First report of brown spot disease caused by Neoscytalidium dimidiatum on Hylocereus undatus in Guangdong, Chinese Mainland. Plant Dis. 2012, 96, 1702. [Google Scholar] [CrossRef]
- Yi, R.H.; Gan, L.J.; Yan, D.H.; Wu, Z.J.; Tong, Y.T.; Wu, F.F. Identification and biological characteristics of Neoscytalidium dimidiatum causing pitaya canker disease. Acta Phytophylacica Sin. 2013, 40, 102–108. [Google Scholar] [CrossRef]
- Ezra, D.; Liarzi, O.; Gat, T.; Hershcovich, M.; Dudai, M. First report of internal black rot caused by Neoscytalidium dimidiatum on Hylocereus undatus (pitahaya) fruit in Israel. Plant Dis. 2013, 97, 1513. [Google Scholar] [CrossRef]
- Mohd, M.H.; Salleh, B.; Zakaria, L. Identification and molecular characterizations of Neoscytalidium dimidiatum causing stem canker of red-fleshed dragon fruit (Hylocereus polyrhizus) in Malaysia. J. Phytopathol. 2013, 161, 841–849. [Google Scholar] [CrossRef]
- Sanahuja, G.; Lopez, P.; Palmateer, A.J. First report of Neoscytalidium dimidiatum causing stem and fruit canker of Hylocereus undatus in Florida. Plant Dis. 2016, 100, 1499. [Google Scholar] [CrossRef]
- Hong, C.F.; Gazis, R.; Crane, J.H.; Zhang, S. Prevalence and epidemics of Neoscytalidium stem and fruit canker on pitahaya (Hylocereus spp.) in South Florida. Plant Dis. 2020, 104, 1433–1438. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Peng, Y.; Qi, Z.; Yan, Z.; Yang, L.; He, M.D.; Li, Q.X.; Liu, C.L.; Ruan, Y.Z.; Wei, S.S.; et al. Identification of Neoscytalidium dimidiatum causing canker disease of pitaya in Hainan, China. Australas. Plant Pathol. 2018, 47, 547–553. [Google Scholar] [CrossRef]
- Balendres, M.A.; Taguiam, J.D.; Evallo, E.; Estigoy, J.; Cortaga, C. Fruit Brown Rot Caused by Neoscytalidium dimidiatum on Selenicereus monacanthus in the Philippines; MycoAsia: Visakhapatnam, India, 2022. [Google Scholar] [CrossRef]
- Serrato-Diaz, L.M.; Goenaga, R. First report of Neoscytalidium dimidiatum causing stem canker on dragon fruit (Hylocereus spp.) in Puerto Rico. Plant Dis. 2021, 105, 2728. [Google Scholar] [CrossRef] [PubMed]
- Dy, K.S.; Wonglom, P.; Pornsuriya, C.; Sunpapao, A. Morphological, molecular ıdentification and pathogenicity of Neoscytalidium dimidiatum causing stem canker of Hylocereus polyrhizus in Southern Thailand. Plants 2022, 11, 504. [Google Scholar] [CrossRef]
- Salunkhe, V.N.; Bhagat, Y.S.; Chavan, S.B.; Lonkar, S.G.; Kakade, V.D. First report of Neoscytalidium dimidiatum causing stem canker of dragon fruit (Hylocereus spp.) in India. Plant Dis. 2023, 107, 1222. [Google Scholar] [CrossRef]
- Khoo, Y.W.; Tan, H.T.; Khaw, Y.S.; Li, S.F.; Chong, K.P. First report of Neoscytalidium dimidiatum causing stem canker on Selenicereus megalanthus in Malaysia. Plant Dis. 2023, 107, 222. [Google Scholar] [CrossRef]
- Espinoza-Lozano, L.; Sumba, M.; Calero, A.; Jiménez, M.I.; Quito-Avila, D.F. First report of Neoscytalidium dimidiatum causing stem canker on yellow dragon fruit (Hylocereus megalantus) in Ecuador. Plant Dis. 2023, 107, 1949. [Google Scholar] [CrossRef]
- Polizzi, G.; Aiello, D.; Vitale, A.; Giuffrida, F.; Groenewald, J.Z.; Crous, P.W. First Report of shoot blight, canker, and gummosis caused by Neoscytalidium dimidiatum on citrus in Italy. Plant Dis. 2009, 93, 1215. [Google Scholar] [CrossRef]
- Mayorquin, J.S.; Wang, D.H.; Twizeyimana, M.; Eskalen, A. Identification, distribution, and pathogenicity of Diatrypaceae and Botryosphaeriaceae associated with citrus branch canker in the Southern California desert. Plant Dis. 2016, 100, 2402–2413. [Google Scholar] [CrossRef]
- Al-Saadoon, A.H.; Ameen, M.K.M.; Hameed, M.A.; Al-Badran, A.; Ali, Z. First report of grapevine dieback caused by Lasiodiplodia theobromae and Neoscytalidium dimidiatum in Basrah, Southern Iraq. Afr. J. Biotechnol. 2012, 11, 16165–16171. [Google Scholar] [CrossRef]
- Rolshausen, P.E.; Akgul, D.S.; Perez, R.; Eskalen, A.; Gispert, C. First report of wood canker caused by Neoscytalidium dimidiatum on grapevine in California. Plant Dis. 2013, 97, 1511. [Google Scholar] [CrossRef] [PubMed]
- Correia, K.C.; Silva, M.A.; Netto, M.S.B.; Vieira, W.A.S.; Camara, M.P.S.; Michereff, S.J. First report of grapevine dieback caused by Neoscytalidium hyalinum in Brazil. Plant Dis. 2016, 100, 213. [Google Scholar] [CrossRef]
- Oksal, E.; Çelik, Y.; Özer, G. Neoscytalidium dimidiatum causes canker and dieback on grapevine in Turkey. Australas. Plant Dis. Notes 2019, 14, 33. [Google Scholar] [CrossRef]
- Akgül, D.S.; Savaş, N.G.; Özarslandan, M. First report of wood canker caused by Lasiodiplodia exigua and Neoscytalidium novaehollandiae on grapevine in Turkey. Plant Dis. 2019, 103, 1036–1037. [Google Scholar] [CrossRef]
- Bahmani, Z.; Abdollahzadeh, J.; Amini, J.; Evidente, A. Biscogniauxia rosacearum the charcoal canker agent as a pathogen associated with grapevine trunk diseases in Zagros region of Iran. Sci. Rep. 2021, 11, 14098. [Google Scholar] [CrossRef] [PubMed]
- Türkölmez, Ş.; Derviş, S.; Çiftçi, O.; Dikilitas, M. First report of Neoscytalidium dimidiatum causing shoot and needle blight of pines (Pinus spp.) in Turkey. Plant Dis. 2019, 103, 2960–2961. [Google Scholar] [CrossRef]
- Alizadeh, M.; Safaie, N.; Shams-Bakhsh, M.; Mehrabadi, M. Neoscytalidium novaehollandiae causes dieback on Pinus eldarica and its potential for infection of urban forest trees. Sci. Rep. 2022, 12, 9337. [Google Scholar] [CrossRef]
- Hajlaoui, M.R.; Nouri, M.T.; Hamrouni, N.; Trouillas, F.P.; Ben Yahmed, N.; Eddouzi, J.; Mnari-Hattab, M. First record of dieback and decline of plum caused by Neoscytalidium dimidiatum in Tunisia. New Dis. Rep. 2018, 38, 20. [Google Scholar] [CrossRef]
- Nouri, M.T.; Lawrence, D.P.; Yaghmour, M.A.; Michailides, T.J.; Trouillas, F.P. Neoscytalidium dimidiatum causing canker, shoot blight and fruit rot of almond in California. Plant Dis. 2018, 102, 1638–1647. [Google Scholar] [CrossRef]
- Oksal, E.; Yiğit, T.; Özer, G. First report of Neoscytalidium dimidiatum causing shoot blight, dieback and canker of apricot in Turkey. J. Plant Pathol. 2020, 102, 579–580. [Google Scholar] [CrossRef]
- Ören, E.; Koca, G.; Gencer, R.; Bayraktar, H. First report of Neoscytalidium novaehollandiae associated with stem canker and branch dieback of almond trees. Australas. Plant Dis. Notes 2020, 15, 17. [Google Scholar] [CrossRef]
- Holland, L.A.; Trouillas, F.P.; Nouri, M.T.; Lawrence, D.P.; Crespo, M.; Doll, D.A.; Duncan, R.A.; Holtz, B.A.; Culumber, C.M.; Yaghmour, M.A.; et al. Fungal pathogens associated with canker diseases of almond in California. Plant Dis. 2021, 105, 346–360. [Google Scholar] [CrossRef] [PubMed]
- Elshafie, A.E.; Ba-Omar, T. First report of Albizia lebbeck dieback caused by Scytalidium dimidiatum in Oman. Mycopathologia 2002, 154, 37–40. [Google Scholar] [CrossRef] [PubMed]
- Ray, J.D.; Burgess, T.; Lanoiselet, V.M. First record of Neoscytalidium dimidiatum and N. novaehollandiae on Mangifera indica and N. dimidiatum on Ficus carica in Australia. Australas. Plant Dis. Notes 2010, 5, 48–50. [Google Scholar] [CrossRef]
- Fernández-Herrera, E.; Moreno-Salazar, S.F.; Rentería-Martínez, M.E.; Arratia-Castro, A.A.; Villar-Luna, E. Neoscytalidium dimidiatum: Causal agent of dieback in Ficus benjamina L. in Mexico. Rev. Chapingo Ser. Hortic. 2017, 23, 203–210. [Google Scholar] [CrossRef]
- Al-Bedak, O.A.; Mohamed, R.A.; Seddek, N.H. First detection of Neoscytalidium dimidiatum associated with canker disease in Egyptian Ficus trees. For. Pathol. 2018, 48, e12411. [Google Scholar] [CrossRef]
- Güney, İ.G.; Bozoğlu, T.; Özer, G.; Türkölmez, Ş.; Derviş, S. First report of Neoscytalidium dimidiatum associated with dieback and canker of common fig (Ficus carica L.) in Turkey. J. Plant Dis. Prot. 2022, 129, 701–705. [Google Scholar] [CrossRef]
- Yeganeh, S.; Mohammadi, H. Sooty canker, a destructive disease of banyan (Ficus benghalensis L.) trees in landscapes of Kish Island (Iran). Urban For. Urban Green. 2022, 72, 127573. [Google Scholar] [CrossRef]
- Ahmadpour, S.A.; Mehrabi-Koushki, M.; Farokhinejad, R.; Mirsoleymani, Z. Characterization and pathogenicity of Neoscytalidium novaehollandiae causing dieback and sooty canker in Iran. Trop. Plant Pathol. 2023. [Google Scholar] [CrossRef]
- Gusella, G.; Fiore, G.; Vitale, A.; Felts, D.G.; Michailides, T.J. New findings on the effects of different factors involved in fig limb dieback caused by Neoscytalidium dimidiatum in California. Eur. J. Plant Pathol. 2023, 167, 89–97. [Google Scholar] [CrossRef]
- Derviş, S.; Türkölmez, Ş.; Çiftçi, O.; Ulubaş Serçe, Ç.; Dikilitas, M. First report of Neoscytalidium dimidiatum causing canker, shoot blight, and root rot of pistachio in Turkey. Plant Dis. 2019, 103, 1411. [Google Scholar] [CrossRef]
- Hashemi, H.; Mohammadi, H. Identification and characterization of fungi associated with internal wood lesions and decline disease of willow and poplar trees in Iran. For. Pathol. 2016, 46, 341–352. [Google Scholar] [CrossRef]
- Türkölmez, Ş.; Derviş, S.; Çiftçi, O.; Ulubaş Serçe, Ç.; Türkölmez, C.G.; Dikilitas, M. First report of Neoscytalidium dimidiatum causing dieback, shoot blight, and branch canker of willow trees in Turkey. Plant Dis. 2019, 103, 2139. [Google Scholar] [CrossRef]
- Fullerton, R.A.; Sutherland, P.A.; Rebstock, R.S.; Hieu, N.T.; Thu, N.N.A.; Linh, D.T.; Thanh, N.T.K.; Van Hoa, N. The life cycle of dragon fruit canker caused by Neoscytalidium dimidiatum and implications for control. In Proceedings of the Dragon Fruit Regional Network Initiation Workshop, Taipei, Taiwan, 23–24 April 2018; pp. 71–80. [Google Scholar]
- Al-Sadi, A.M.; Al-Ghaithi, A.G.; Al-Fahdi, N.; Al-Yahyai, R. Characterization and pathogenicity of fungal pathogens associated with root diseases of citrus in Oman. Int. J. Agric. Biol. 2014, 16, 371–376. [Google Scholar]
- Türkölmez, Ş.; Derviş, S.; Çiftçi, O.; Serçe, Ç.U.; Dikilitas, M. New disease caused by Neoscytalidium dimidiatum devastates tomatoes (Solanum lycopersicum) in Turkey. Crop Prot. 2019, 118, 21–30. [Google Scholar] [CrossRef]
- Jiménez Luna, I.; Doll, D.; Ashworth, V.; Trouillas, F.P.; Rolshausen, P.E. Comparative profiling of wood canker pathogens from spore traps and symptomatic plant samples within California almond and walnut orchards. Plant Dis. 2022, 106, 2182–2190. [Google Scholar] [CrossRef] [PubMed]
- Güney, İ.G.; Bozoğlu, T.; Özer, G.; Derviş, S. A novel blight and root rot of chickpea: A new host for Neoscytalidium dimidiatum. Crop Prot. 2023, 172, 106326. [Google Scholar] [CrossRef]
- Crous, P.W.; Slippers, B.; Wingfield, M.J.; Rheeder, J.; Marasas, W.F.; Philips, A.J.; Alves, A.; Burgess, T.; Barber, P.; Groenewald, J.Z. Phylogenetic lineages in the Botryosphaeriaceae. Stud. Mycol. 2006, 55, 235–253. [Google Scholar] [CrossRef]
- Pavlic, D.; Wingfield, M.J.; Barber, P.; Slippers, B.; Hardy, G.E.; Burgess, T.I. Seven new species of the Botryosphaeriaceae from baobab and other native trees in Western Australia. Mycologia 2008, 100, 851–866. [Google Scholar] [CrossRef]
- Phillips, A.J.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.Z.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef]
- Huang, S.K.; Tangthirasunun, N.; Phillips, A.J.; Dai, D.Q.; Wanasinghe, D.N.; Wen, T.C.; Bahkali, A.H.; Hyde, K.D.; Kang, J.C. Morphology and phylogeny of Neoscytalidium orchidacearum sp. nov. (Botryosphaeriaceae). Mycobiology 2016, 44, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Calvillo-Medina, R.P.; Martinez-Neria, M.; Mena-Portales, J.; Barba-Escoto, L.; Raymundo, T.; Campos-Guillen, J.; Jones, G.H.; Reyes-Grajeda, J.P.; Gonzalez, Y.M.J.A.; Bautista-de Lucio, V.M. Identification and biofilm development by a new fungal keratitis aetiologic agent. Mycoses 2019, 62, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Wonglom, P.; Pornsuriya, C.; Sunpapao, A. A new species of Neoscytalidium hylocereum sp. nov. causing canker on red-fleshed dragon fruit (Hylocereus polyrhizus) in Southern Thailand. J. Fungi 2023, 9, 197. [Google Scholar] [CrossRef] [PubMed]
- Nattrass, R.M. A new species of Hendersonula (H. toruloidea) on deciduous trees in Egypt. Trans. Br. Mycol. Soc. 1933, 18, 189–198. [Google Scholar] [CrossRef]
- Sutton, B.C.; Dyko, B.J. Revision of Hendersonula. Mycol. Res. 1989, 93, 466–488. [Google Scholar] [CrossRef]
- Farr, D.F.; Elliott, M.; Rossman, A.Y.; Edmonds, R.L. Fusicoccum arbuti sp. nov. causing cankers on Pacific madrone in western North America with notes on Fusicoccum dimidiatum, the correct name for Scytalidium dimidiatum and Natrassia mangiferae. Mycologia 2005, 97, 730–741. [Google Scholar] [CrossRef]
- Penzig, O. Funghi agrumicoli. Contribuzione allo studio dei funghi parassiti degli agrumi. Michelia 1882, 2, 385–508. [Google Scholar]
- Wilson, E.E. The branch wilt of Persian walnut trees and its cause. Hilgardia 1947, 17, 413–436. [Google Scholar] [CrossRef]
- Campbell, C.K.; Mulder, J.L. Skin and nail infection by Scytalidium hyalinum sp. nov. Sabouraudia 1977, 15, 161–166. [Google Scholar] [CrossRef]
- Slippers, B.; Johnson, G.I.; Crous, P.W.; Coutinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Phylogenetic and morphological re-evaluation of the Botryosphaeria species causing diseases of Mangifera indica. Mycologia 2005, 97, 99–110. [Google Scholar] [CrossRef]
- Sydow, H.; Sydow, P.; Butler, E.J. Fungi Indiae Orientalis. V. Ann. Mycol. 1916, 14, 177–220. [Google Scholar]
- Madrid, H.; Ruiz-Cendoya, M.; Cano, J.; Stchigel, A.; Orofino, R.; Guarro, J. Genotyping and in vitro antifungal susceptibility of Neoscytalidium dimidiatum isolates from different origins. Int. J. Antimicrob. Agents 2009, 34, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Xu, J. Fungal species concepts in the genomics era. Genome 2020, 63, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Maharachchikumbura, S.S.N.; Chen, Y.; Ariyawansa, H.A.; Hyde, K.D.; Haelewaters, D.; Perera, R.H.; Samarakoon, M.C.; Wanasinghe, D.N.; Bustamante, D.E.; Liu, J.; et al. Integrative approaches for species delimitation in Ascomycota. Fungal Divers. 2021, 109, 155–179. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP*: Phylogenetic Analysis Using Parsimony. (* and Other Methods); Version 4.0b10; Sinauer Associates: Sunderland, UK, 2003. [Google Scholar]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Coutinho, I.B.L.; Cardoso, J.E.; Lima, C.S.; Lima, J.S.; Goncalves, F.J.T.; Silva, A.M.S.; Freire, F.C.O. An emended description of Neofusicoccum brasiliense and characterization of Neoscytalidium and Pseudofusicoccum species associated with tropical fruit plants in northeastern Brazil. Phytotaxa 2018, 358, 251–264. [Google Scholar] [CrossRef]
- Hohenfeld, C.S.; Santana, M.P.; Junior, L.R.C.; de Oliveira, E.J.; de Oliveira, S.A.S. Modelling growth characteristics and aggressiveness of Neoscytalidium hyalinum and Fusarium solani associated with black and dry root rot diseases on cassava. Trop. Plant Pathol. 2018, 43, 422–432. [Google Scholar] [CrossRef]
- Sabernasab, M.; Jamali, S.; Marefat, A.; Abbasi, S. Morphological and molecular characterization of Neoscytalidium novaehollandiae, the cause of Quercus brantii dieback in Iran. Phytopathol. Mediterr. 2019, 58, 347–357. [Google Scholar] [CrossRef]
- Gusella, G.; Morgan, D.P.; Michailides, T.J. Further ınvestigation on limb dieback of fig (Ficus carica) Caused by Neoscytalidium dimidiatum in California. Plant Dis. 2021, 105, 324–330. [Google Scholar] [CrossRef] [PubMed]
- Güney, İ.G.; Özer, G.; Türkölmez, Ş.; Derviş, S. Canker and leaf scorch on olive (Olea europaea L.) caused by Neoscytalidium dimidiatum in Turkey. Crop Prot. 2022, 157, 105985. [Google Scholar] [CrossRef]
- Goudarzi, A.; Moslehi, M. Distribution of a devastating fungal pathogen in mangrove forests of southern Iran. Crop Prot. 2020, 128, 104987. [Google Scholar] [CrossRef]
- Nurul Nadiah, M.A.; Mohamed Nor, N.M.I.; Latiffah, Z.; Masratul Hawa, M. First report of leaf blight on white spider lily caused by Neoscytalidium dimidiatum in Malaysia. New Dis. Rep. 2017, 35, 16. [Google Scholar] [CrossRef]
- Zaeimian, Z.; Fotouhifar, K.B. First report of Neoscytalidium dimidiatum as the causal agent of leaf blight on Clivia miniata. Sci. Rep. 2023, 13, 16110. [Google Scholar] [CrossRef]
- Pandey, R.S.; Bhargava, S.N.; Shukla, D.N.; Khati, D.V.S. A new leaf spot disease of mango. Plant Dis. 1981, 65, 441–442. [Google Scholar] [CrossRef]
- Reckhaus, P.; Adamous, I. Hendersonula dieback of mango in Niger. Plant Dis. 1987, 71, 1045. [Google Scholar] [CrossRef]
- Mendes, M.A.S.; da Silva, V.L.; Dianese, J.C.; Ferreira, M.A.S.V.; dos Santos, C.E.N.; Urben, A.F.; Castro, C.; Gomes Neto, E. Fungos em Plants no Brasil; Embrapa-SPI/Embrapa-Cenargen: Brasilia, Brazil, 1998; 555p. [Google Scholar]
- Sakalidis, M.L.; Ray, J.D.; Lanoiselet, V.; St, J.; Hardy, G.E.; Burgess, T.I. Pathogenic Botryosphaeriaceae associated with Mangifera indica in the Kimberley Region of Western Australia. Eur. J. Plant Pathol. 2011, 130, 379–391. [Google Scholar] [CrossRef]
- Marques, M.W.; Lima, N.B.; de Morais, M.A.; Michereff, S.J.; Phillips, A.J.; Câmara, M.P. Botryosphaeria, Neofusicoccum, Neoscytalidium and Pseudofusicoccum species associated with mango in Brazil. Fungal Divers. 2013, 61, 195–208. [Google Scholar] [CrossRef]
- Dolatabad, H.K.; Javan-Nikkhah, M.; Shier, W.T. Evaluation of antifungal, phosphate solubilisation, and siderophore and chitinase release activities of endophytic fungi from Pistacia vera. Mycol. Prog. 2017, 16, 777–790. [Google Scholar] [CrossRef]
- Kurt, Ş.; Uysal, A.; Soylu, E.M.; Kara, M.; Soylu, S. First record of Neoscytalidium novaehollandiae associated with pistachio dieback in the Southeastern Anatolia region of Turkey. Mycol. Iran 2019, 6, 55–57. [Google Scholar] [CrossRef]
- Williams, L.; Hayne, S.C. Index of Plant Diseases in West Virginia; West Virginia Department of Agriculture Plant Industries Division: Ripley, WI, USA, 1982; 115p. [Google Scholar]
- Farr, D.F.; Bills, G.F.; Chamuris, G.P.; Rossman, A.Y. Fungi on Plants and Plant Products in the United States; The American Phytopathological Society: St. Paul, MN, USA, 1989. [Google Scholar]
- Mathur, R.S. The Coelomycetes of India; Bishen Singh Mahendra Pal Singh: Delhi, India, 1979; 460p. [Google Scholar]
- Gusella, G.; Di Pietro, C.; Vecchio, L.; Campo, G.; Polizzi, G. Branch canker and dieback of Meryta denhamii caused by Neofusicoccum parvum and Neoscytalidium dimidiatum in Italy. Australas. Plant Dis. Notes 2023, 18, 31. [Google Scholar] [CrossRef]
- Peregrine, W.T.H.; Ahmad, K.B. Brunei: A first annotated list of plant diseases and associated organisms. Phytopathol. Pap. 1982, 27, 1–87. [Google Scholar]
- Kranz, J. Fungi collected in the Republic of Guinea, Collections from the Kindia area in 1962. Sydowia 1963, 17, 174–185. [Google Scholar]
- Xie, H.H.; Long, L.; Huang, S.; Mao, L.; Huang, Q.; Wang, L.; Li, J. First Report of Black Spot Caused by Neoscytalidium dimidiatum on Sisal in Guangxi, China. Plant Dis. 2021, 105, 701. [Google Scholar] [CrossRef]
- Johnston, A. A supplement to a host list of plant diseases in Malaya. Mycol. Pap. 1960, 77, 1–30. [Google Scholar]
- Kee, Y.J.; Suhaimi, N.N.; Zakaria, L.; Mohd, M.H. Characterisation of Neoscytalidium dimidiatum causing leaf blight on Sansevieria trifasciata in Malaysia. Australas. Plant Dis. Notes 2017, 12, 60. [Google Scholar] [CrossRef]
- Monteles, R.P.; Sousa, E.S.; da Silva Matos, K.; de Brito, V.S.T.; de Melo, M.P.; Beserra, J.E.A. Neoscytalidium dimidiatum causes leaf blight on Sansevieria trifasciata in Brazil. Australas. Plant Dis. Notes 2020, 15, 19. [Google Scholar] [CrossRef]
- Crous, P.W.; Hernández-Restrepo, M.; Schumacher, R.K.; Cowan, D.A.; Maggs-Kölling, G.; Marais, E.; Groenewald, J.Z. New and interesting fungi. Fungal Syst. Evol. 2021, 7, 255–343. [Google Scholar] [CrossRef]
- Esmaeili, N.; Mohammadi, H.; Sohrabi, M. Barberry (Berberis vulgaris L.) as an alternative host of grapevine fungal trunk pathogens. Eur. J. Plant Pathol. 2023. [Google Scholar] [CrossRef]
- Ebbels, D.L.; Allen, D.J. A supplementary and annotated list of plant diseases, pathogens and associated fungi in Tanzania. Phytopathol. Pap. 1979, 22, 1–89. [Google Scholar]
- Kuruppu, M.; Siddiqui, Y.; Ahmad, K.; Ali, A. First report of postharvest stem end rot disease on MD2 pineapple fruits caused by Neoscytalidium dimidiatum in Malaysia. Plant Dis. 2021, 105, 1564. [Google Scholar] [CrossRef] [PubMed]
- Feijo, F.M.; Silva, M.J.S.; Nascimento, A.D.; Infante, N.B.; Ramos-Sobrinho, R.; Assuncao, I.P.; Lima, G.S.A. Botryosphaeriaceae species associated with the pickly pear cactus, Nopalea cochenillifera. Trop. Plant Pathol. 2019, 44, 452–459. [Google Scholar] [CrossRef]
- Yi, R.H.; Lin, Q.L.; Mo, J.J.; Wu, F.F.; Chen, J. Fruit internal brown rot caused by Neoscytalidium dimidiatum on pitahaya in Guangdong Province, China. Australas. Plant Dis. Notes 2015, 10, 13. [Google Scholar] [CrossRef]
- Boa, E.; Lenné, J. Diseases of Nitrogen Fixing Trees in Developing Countries. An Annotated List; Natural Resources Institute: Kent, UK, 1994; 82p. [Google Scholar]
- Watson, A.J. Foreign Bacterial and Fungus Diseases of Food, Forage, and Fiber Crops; U.S.D.A. Agricultural Research Service: Washington, DC, USA, 1971; Volume 111.
- de Mello, J.F.; Brito, A.C.Q.; Motta, C.M.S.; Vieira, J.C.B.; Michereff, S.J.; Machado, A.R. First report of Neoscytalidium dimidiatum causing root rot in sweet potato in Brazil. Plant Dis. 2019, 103, 373. [Google Scholar] [CrossRef]
- de Mello, J.F.; Brito, A.C.Q.; Vieira, J.C.B.; Camara, M.P.S.; Michereff, S.J.; de Souza-Motta, C.M.; Machado, A.R. Identification and pathogenicity of Botryosphaeriaceae species associated with root and stem rot of sweet potato in Brazil. Plant Pathol. 2021, 70, 1601–1615. [Google Scholar] [CrossRef]
- Mirtalebi, M.; Sabahi, F.; Banihashemi, Z. Fruit rot caused by Neoscytalidium hyalinum on melon in Iran. Australas. Plant Dis. Notes 2019, 14, 8. [Google Scholar] [CrossRef]
- Anonymous. California Fungi. Nos. 1-1325. Exsiccati Set 1931–1970, N/A Pages. Available online: https://data.nal.usda.gov/dataset/united-states-national-fungus-collections-fungus-host-dataset (accessed on 22 August 2023).
- Lin, C.H.; Chen, Y.X.; Liu, W.B.; Wu, W.Q.; Miao, W.G.; Zheng, F.C. First report of Dioscorea esculenta dieback caused by Neoscytalidium dimidiatum in China. Plant Dis. 2017, 101, 1320. [Google Scholar] [CrossRef]
- Arrieta-Guerra, J.J.; Díaz-Cabadiaz, A.T.; Pérez-Pazos, J.V.; Cadena-Torres, J.; Sánchez-López, D.B. Fungi associated with dry rot disease of yam (Dioscorea rotundata Poir.) tubers in Cordoba, Colombia. Agron. Mesoam. 2021, 32, 790–807. [Google Scholar] [CrossRef]
- Ören, E.; Koca, G.; Bayraktar, H. First report of Neoscytalidium novaehollandiae associated with branch dieback on Japanese persimmon in Turkey. J. Plant Pathol. 2020, 102, 1311–1312. [Google Scholar] [CrossRef]
- Davison, A.D. Factors affecting development of madrone canker. Plant Dis. Rep. 1972, 56, 50–52. [Google Scholar]
- Shaw, C.G. Host Fungus Index for the Pacific Northwest-I. Hosts; Washington Agricultural Experiment Station: Washington, DC, USA, 1973; Volume 765, pp. 1–121. [Google Scholar]
- Ginns, J.H. Compendium of plant disease and decay fungi in Canada 1960–1980. Res. Br. Can. Agric. Publ. 1986, 1813, 416. [Google Scholar]
- French, A.M. California Plant Disease Host Index; California Department of Food and Agriculture: Sacramento, CA, USA, 1989; 394p.
- Tsahouridou, P.C.; Thanassoulopoulos, C.C. First report of Hendersonula toruloidea as a foliar pathogen of strawberry-tree (Arbutus unedo) in Europe. Plant Dis. 2000, 84, 487. [Google Scholar] [CrossRef]
- Machado, A.R.; Pinho, D.B.; Dutra, D.C.; Pereira, O.L. First report of collar and root rot of physic nut (Jatropha curcas) caused by Neoscytalidium dimidiatum in Brazil. Plant Dis. 2012, 96, 1697. [Google Scholar] [CrossRef] [PubMed]
- Machado, A.R.; Pinho, D.B.; Pereira, O.L. Phylogeny, identification and pathogenicity of the Botryosphaeriaceae associated with collar and root rot of the biofuel plant Jatropha curcas in Brazil, with a description of new species of Lasiodiplodia. Fungal Divers. 2014, 67, 231–247. [Google Scholar] [CrossRef]
- Hughes, S.J. Fungi From the Gold Coast. I. Mycol. Pap. 1952, 48, 1–91. [Google Scholar]
- Nattrass, R.M. Host lists of Kenya fungi and bacteria. Mycol. Pap. 1961, 81, 1–46. [Google Scholar]
- Machado, A.R.; Pinho, D.B.; Oliveira, S.A.S.; Pereira, O.L. New occurrences of Botryosphaeriaceae causing black root rot of cassava in Brazil. Trop. Plant Pathol. 2014, 39, 464–470. [Google Scholar] [CrossRef]
- Brito, A.C.Q.; de Mello, J.F.; Camara, M.P.S.; Vieira, J.C.B.; Michereff, S.J.; Souza-Motta, C.M.; Machado, A.R. Diversity and pathogenicity of Botryosphaeriaceae species associated with black root rot and stem cutting dry rot in Manihot esculenta in Brazil. Eur. J. Plant Pathol. 2020, 157, 583–598. [Google Scholar] [CrossRef]
- Sangpueak, R.; Duchanee, S.; Saengchan, C.; Papathoti, N.K.; Hoang, N.H.; Thanh, T.L.; Phansak, P.; Buensanteai, N. Identification of cassava black stem and root rot agents in Thailand. Chil. J. Agric. Res. 2023, 83, 70–82. [Google Scholar] [CrossRef]
- Pande, A.; Rao, V.G. A Compendium Fungi on Legumes from India; Scientific Publishers: Jodhpur, India, 1998; 188p. [Google Scholar]
- Chandra, S. Some new leaf-spot diseases from Allahabad (India). Beih. Nova Hedwig. 1974, 47, 35–102. [Google Scholar]
- Sakalidis, M.L.; Hardy, G.E.S.; Burgess, T.I. Endophytes as potential pathogens of the baobab species Adansonia gregorii: A focus on the Botryosphaeriaceae. Fungal Ecol. 2011, 4, 1–14. [Google Scholar] [CrossRef]
- Al Raish, S.M.; Saeed, E.E.; Sham, A.; Alblooshi, K.; El-Tarabily, K.A.; AbuQamar, S.F. Molecular characterization and disease control of stem canker on royal poinciana (Delonix regia) caused by Neoscytalidium dimidiatum in the United Arab Emirates. Int. J. Mol. Sci. 2020, 21, 1033. [Google Scholar] [CrossRef] [PubMed]
- Galea, V.J. Use of stem ımplanted bioherbicide capsules to manage an ınfestation of Parkinsonia aculeata in Northern Australia. Plants 2021, 10, 1909. [Google Scholar] [CrossRef]
- Anonymous. Index of Plant Diseases in the United States. In U.S.D.A. Agriculture Handbook; U.S. Department of Agriculture: Washington, DC, USA, 1960; Volume 165, pp. 1–531. [Google Scholar]
- Alidadi, A.; Kowsari, M.; Javan-Nikkhah, M.; Jouzani, G.R.S.; Rastaghi, M.E. New pathogenic and endophytic fungal species associated with Persian oak in Iran. Eur. J. Plant Pathol. 2019, 155, 1017–1032. [Google Scholar] [CrossRef]
- French, A.M. California Plant Disease Host Index. Part 1: Fruit and Nuts; California Department of Food and Agriculture: Sacramento, CA, USA, 1987; 39p.
- Wilson, E.E. The pycnidial stage of the walnut branch wilt fungus, Exosporina fawcettii. Phytopathology 1949, 53, 705–712. [Google Scholar]
- Sommer, N.F. Sunburn predisposes walnut trees to branch-wilt. Phytopathology 1955, 45, 607–613. [Google Scholar]
- Phillips, A.J.L.; Oudemans, P.V.; Correia, A.; Alves, A. Characterisation and epitypification of Botryosphaeria corticis, the cause of blueberry cane canker. Fung Divers. 2006, 21, 141–155. [Google Scholar]
- Chen, S.F.; Fichtner, E.; Morgan, D.P.; Michailides, T.J. First report of Lasiodiplodia citricola and Neoscytalidium dimidiatum causing death of graft union of English walnut in California. Plant Dis. 2013, 97, 993. [Google Scholar] [CrossRef]
- Chen, S.; Morgan, D.P.; Hasey, J.K.; Anderson, K.; Michailides, T.J. Phylogeny, morphology, distribution, and pathogenicity of Botryosphaeriaceae and Diaporthaceae from English walnut in California. Plant Dis. 2014, 98, 636–652. [Google Scholar] [CrossRef] [PubMed]
- Derviş, S.; Türkölmez, Ş.; Çiftçi, O.; Ulubaş Serçe, Ç.; Dikilitas, M. First report of Neoscytalidium dimidiatum causing black canker and root rot of walnut in Turkey. Plant Dis. 2019, 103, 2129. [Google Scholar] [CrossRef]
- Çiftçi, O.; Özer, G.; Türkölmez, Ş.; Derviş, S. Lasiodiplodia theobromae and Neoscytalidium dimidiatum associated with grafted walnut (Juglans regia L.) decline in Turkey. J. Plant Dis. Prot. 2023, 130, 1117–1128. [Google Scholar] [CrossRef]
- Bagherabadi, S.; Zafari, D.; Maharachchikumbura, S.S. Neoscytalidium dimidiatum as one of the fungal agents associated with walnut decline in Iran. J. Nuts 2022, 13, 31–39. [Google Scholar] [CrossRef]
- Güney, İ.G.; Özer, G.; Turan, İ.; Koşar, İ.; Derviş, S. First report of Neoscytalidium dimidiatum causing foliar and stem blight of lavender in Turkey. J. Plant Pathol. 2021, 103, 1347–1348. [Google Scholar] [CrossRef]
- Özer, G.; Günen, T.U.; Koşar, İ.; Güler, İ.G.; Derviş, S. First report of Neoscytalidium dimidiatum causing blight of Melissa officinalis in Turkey. J. Plant Dis. Prot. 2022, 129, 197–199. [Google Scholar] [CrossRef]
- Alkan, M.; Özer, G.; Koşar, I.; Güney, I.G.; Derviş, S. First report of leaf blight of Turkish oregano (Origanum onites) caused by Neoscytalidium dimidiatum in Turkey. J. Plant Pathol. 2022, 104, 471. [Google Scholar] [CrossRef]
- Derviş, S.; Güney, İ.G.; Koşar, İ.; Bozoğlu, T.; Özer, G. First report of Neoscytalidium novaehollandiae on common sage (Salvia officinalis). Australas. Plant Dis. 2021, 16, 19. [Google Scholar] [CrossRef]
- Kadkhoda-Hematabadi, S.; Mohammadi, H.; Sohrabi, M. Morphological and molecular identification of plant pathogenic fungi associated with necrotic wood tissues of pomegranate trees in Iran. J. Plant Pathol. 2023, 105, 465–479. [Google Scholar] [CrossRef]
- Ghasemi-Sardareh, R.; Mohammadi, H. Characterization and pathogenicity of fungal trunk pathogens associated with declining of neem (Azadirachta indica A. Juss) trees in Iran. J. Plant Pathol. 2020, 102, 1159–1171. [Google Scholar] [CrossRef]
- Ahmad, S.; Iqbal, S.H.; Khalid, A.N. Fungi of Pakistan. Sultan Ahmad Mycological Society of Pakistan; Department of Botany, University of the Punjab: Lahore, Pakistan, 1997; 248p. [Google Scholar]
- Abdulrahman, D.N.; Haleem, R.A. Morphological and molecular characterization of Neoscytalidium isolates that cause canker and dieback in eucalyptus and chinaberry trees in Iraq. Plant Prot. Sci. 2023, 59, 92–105. [Google Scholar] [CrossRef]
- Georghiou, G.P.; Papadopoulos, C. A Second List of Cyprus Fungi; Government of Cyprus, Department of Agriculture: Nicosia, Cyprus, 1957; 38p.
- Paxton, J.D.; Wilson, E.E.; Davis, J.R. Branch wilt disease of fig caused by Hendersonula toruloidea. Plant Dis. Rep. 1964, 48, 142. [Google Scholar]
- Oksal, E. Prevalence, molecular characterization, and variety reactions of Neoscytalidium novaehollandiae on mulberries in Turkey. Not. Bot. Horti Agrobot. Cluj-Napoca 2022, 50, 12716. [Google Scholar] [CrossRef]
- Meredith, D.S. Tip rot of banana fruits in Jamaica. Trans. Br. Mycol. Soc. 1963, 46, 473–481. [Google Scholar] [CrossRef]
- Meredith, D.S. Fungal diseases of bananas in Hawaii. Plant Dis. Rep. 1969, 53, 63–66. [Google Scholar]
- Raabe, R.D.; Conners, I.L.; Martinez, A.P. Checklist of Plant Diseases in Hawaii; Information Text Series No. 22; College of Tropical Agriculture and Human Resources, University of Hawaii: Honolulu, HI, USA, 1981; 313p. [Google Scholar]
- Al-Tememe, Z.A.M.; Lahuf, A.; Abdalmoohsin, R.G.; Al-Amirry, A.T. Occurrence, identification, pathogenicity and control of Neoscytalidium dimidiatum fungus, the causal agent of sooty canker on Eucalyptus camaldulensis in Kerbala Province of Iraq. Plant Arch. 2019, 19, 31–38. [Google Scholar]
- Ismail, S.I.; Ahmad Dahlan, K.; Abdullah, S.; Zulperi, D. First Report of Neoscytalidium dimidiatum causing fruit rot on guava (Psidium guajava L.) in Malaysia. Plant Dis. 2021, 105, 220. [Google Scholar] [CrossRef]
- Panahandeh, S.; Mohammadi, H.; Gramaje, D. Trunk disease fungi associated with Syzygium cumini in Iran. Plant Dis. 2019, 103, 711–720. [Google Scholar] [CrossRef]
- Williams, T.H.; Liu, P.S.W. A host list of plant diseases in Sabah, Malaysia. Phytopathol. Pap. 1976, 19, 1–67. [Google Scholar]
- Suwannarach, N.; Kumla, J.; Lumyong, S. Leaf spot on cattleya orchid caused by Neoscytalidium orchidacearum in Thailand. Can. J. Plant Pathol. 2018, 40, 109–114. [Google Scholar] [CrossRef]
- Chang, C.W.; Chen, C.Y.; Wang, C.L.; Chung, W.H. First report of leaf blight on Cattleya × hybrid caused by Neoscytalidium dimidiatum in Taiwan. J. Plant Pathol. 2020, 102, 921. [Google Scholar] [CrossRef]
- Derviş, S.; Türkölmez, Ş.; Güney, İ.G.; Alkan, M.; Ozer, G. First report of needle blight of blue spruce (Picea pungens) caused by Neoscytalidium dimidiatum in Turkey. J. Plant Pathol. 2023, 105, 1195–1196. [Google Scholar] [CrossRef]
- Nourian, A.; Salehi, M.; Safaie, N.; Khelghatibana, F.; Abdollahzadeh, J. Fungal canker agents in apple production hubs of Iran. Sci. Rep. 2021, 11, 22646. [Google Scholar] [CrossRef] [PubMed]
- Sha, S.; Wang, Z.; Hao, H.; Wang, L.; Feng, H. First report of Neoscytalidium dimidiatum inducing canker disease on apple trees in China. J. Plant Pathol. 2022, 104, 1149–1150. [Google Scholar] [CrossRef]
- Ören, E.; Palacıoğlu, G.; Ozan, G.N.; Çelik, K.; Bayraktar, H. First report of Neoscytalidium novaehollandiae associated with canker and branch dieback on cherry trees in Turkey. J. Plant Pathol. 2022, 104, 391. [Google Scholar] [CrossRef]
- Ören, E.; Palacıoğlu, G.; Ozan, G.N.; Bayraktar, H. First report of Neoscytalidium novaehollandiae associated with branch dieback and stem cankers on plum in Turkey. J. Plant Pathol. 2022, 104, 1167. [Google Scholar] [CrossRef]
- English, H.; Davis, J.R.; Devay, J.E. Relationship of Botryosphaeria dothidea and Hendersonula toruloidea to a canker disease of almond. Phytopathology 1975, 65, 114–122. [Google Scholar] [CrossRef]
- Burgess, T.I.; Barber, P.A.; St., J.; Hardy, G.E. Botryosphaeria spp. associated with eucalypts in Western Australia, including the description of Fusicoccum macroclavatum sp. nov. Australas. Plant Pathol. 2005, 34, 557–567. [Google Scholar] [CrossRef]
- Oksal, E.; Özer, G. First report of shoot blight and branch canker of Pyrus communis by Neoscytalidium novaehollandiae in Turkey. J. Plant Pathol. 2021, 103, 673–674. [Google Scholar] [CrossRef]
- Espargham, N.; Mohammadi, H.; Gramaje, D. A Survey of Trunk Disease Pathogens within Citrus Trees in Iran. Plants 2020, 9, 754. [Google Scholar] [CrossRef]
- Alananbeh, K.M.; Al-Qasim, M.; Gharaibeh, A.; Al-Hiary, H.A. First report of shoot blight caused by Neoscytalidium dimidiatum on citrus in Jordan. Plant Dis. 2020, 104, 571. [Google Scholar] [CrossRef]
- Calavan, E.C.; Wallace, J.M. Hendersonula toruloidea Nattrass on Citrus in California. Phytopathology 1954, 44, 635–639. [Google Scholar]
- Adesemoye, A.O.; Mayorquin, J.S.; Wang, D.H.; Twizeyimana, M.; Lynch, S.C.; Eskalen, A. Identification of species of Botryosphaeriaceae causing bot gummosis in citrus in California. Plant Dis. 2014, 98, 55–61. [Google Scholar] [CrossRef]
- Doidge, E.M. The South African fungi and lichens to the end of 1945. Bothalia 1950, 5, 1–1094. [Google Scholar]
- Ogawa, J.M. The occurrence of Hendersonula toruloidea Nattrass on Populus species in California. Plant Dis. Rep. 1954, 38, 238. [Google Scholar]
- Derviş, S.; Özer, G.; Türkölmez, Ş. First report of Neoscytalidium novaehollandiae causing stem blight on tomato in Turkey. J. Plant Pathol. 2020, 102, 1339–1340. [Google Scholar] [CrossRef]
- Derviş, S.; Özer, G.; Türkölmez, Ş. First report of Neoscytalidium dimidiatum causing tuber rot of potato in Turkey. J. Plant Pathol. 2020, 102, 1295–1296. [Google Scholar] [CrossRef]
- Hashemi, H.; Mohammadi, H.; Abdollahzadeh, J. Symptoms and fungi associated with elm trees decline in Iran. Eur. J. For. Res. 2017, 136, 857–879. [Google Scholar] [CrossRef]
- Wangikar, P.D.; Raut, J.G.; Gopalkrishna, N. Drying of grape vines caused by Hendersonula toruloidea. Indian Phytopathol. 1969, 22, 403–404. [Google Scholar]
- Sarbhoy, A.K.; Lal, G.; Varshney, J.L. Fungi of India (1967–1971); Navyug Traders: New Delhi, India, 1971; 148p. [Google Scholar]
- Natour, R.M.; Ahmed, J.M. Control of branch wilt disease of grape. Plant Dis. Rep. 1969, 53, 152–153. [Google Scholar]
- Ören, E.; Palacıoğlu, G.; Koca, G.; Ozan, G.N.; Bayraktar, H. First report of Neoscytalidium dimidiatum causing branch dieback and canker on apple in Turkey. J. Plant Pathol. 2022, 104, 429. [Google Scholar] [CrossRef]
- Xu, M.; Liu, C.L.; Fu, Y.; Liao, Z.W.; Guo, P.Y.; Xiong, R.; Cheng, Y.; Wei, S.S.; Huang, J.Q.; Tang, H. Molecular characterization and expression analysis of pitaya (Hylocereus polyrhizus) HpLRR genes in response to Neoscytalidium dimidiatum infection. BMC Plant Biol. 2020, 20, 160. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.F.; Lawrence, D.P.; Morales-Cruz, A.; Travadon, R.; Minio, A.; Hernandez-Martinez, R.; Rolshausen, P.E.; Baumgartner, K.; Cantu, D. Phylogenomics of plant-associated Botryosphaeriaceae Species. Front. Microbiol. 2021, 12, 652802. [Google Scholar] [CrossRef]
- Türkölmez, Ş.; Eren, A.; Özer, G.; Derviş, S. The effect of Talaromyces funiculosus ST976 isolated from pistacia vera rhizosphere on phosphorus solubility in soil samples with different physicochemical properties. J. Agric. Nat. 2022, 25, 1077–1085. [Google Scholar] [CrossRef]
- Kuan, C.S.; Ng, K.P.; Yew, S.M.; Umar Meleh, H.; Seow, H.F.; How, K.N.; Yeo, S.K.; Jee, J.M.; Tan, Y.C.; Yee, W.Y.; et al. Comparative genomic and phenotypic analyses of pathogenic fungi Neoscytalidium dimidiatum and Bipolaris papendorfii isolated from human skin scraping. Braz. J. Microbiol. 2023, 54, 1351–1372. [Google Scholar] [CrossRef] [PubMed]
- Slippers, B.; Wingfield, M.J. Botryosphaeriaceae as endophytes and latent pathogens of woody plants: Diversity, ecology and impact. Fungal Biol. Rev. 2007, 21, 90–106. [Google Scholar] [CrossRef]
- Khan, A.H. Some new diseases of Citrus in Pakistan. Mycopathol. Mycol. Appl. 1959, 10, 289–302. [Google Scholar] [CrossRef]
- Paxton, J.D.; Wilson, E.E. Anatomical and physiological aspects of branch wilt disease of Persian walnut. Phytopathology 1965, 55, 21. [Google Scholar]
- Hassan, W.A.; Haleem, R.A.; Hassan, P.H. Effect of heat-stress predisposition on the development of sooty canker caused by Neoscytalidium dimidiatum (Penz.) Crous and Slippers. Acta Agrobot. 2011, 64, 207–212. [Google Scholar] [CrossRef]
- Zhu, X.M.; Liu, X.F. A new species and genus distribution record from China: Neoscytalidium novaehollandiae. Indian. J. Microbiol. 2012, 52, 565–568. [Google Scholar] [CrossRef]
- Corlett, R.T. Frugivory and seed dispersal by vertebrates in tropical and subtropical Asia: An update. Glob. Ecol. Conserv. 2017, 11, 1–22. [Google Scholar] [CrossRef]
- Belair, M.; Restrepo-Leal, J.D.; Praz, C.; Fontaine, F.; Remond, C.; Fernandez, O.; Besaury, L. Botryosphaeriaceae gene machinery: Correlation between diversity and virulence. Fungal Biol. 2023, 127, 1010–1031. [Google Scholar] [CrossRef] [PubMed]
- Nagel, J.H.; Wingfield, M.J.; Slippers, B. Increased abundance of secreted hydrolytic enzymes and secondary metabolite gene clusters define the genomes of latent plant pathogens in the Botryosphaeriaceae. BMC Genom. 2021, 22, 589. [Google Scholar] [CrossRef] [PubMed]
- Reveglia, M.; Masi, A.; Cimmino, T.; Cinelli, L.; Mugnai, A. Evidente phytotoxins produced by Lasiodiplodia laeliocattleyae involved in Botryosphaeria dieback of grapevines in Brazil. Phytopathol Mediterr. 2019, 58, 207–211. [Google Scholar] [CrossRef]
- Trotel-Aziz, P.; Robert-Siegwald, G.; Fernandez, O.; Leal, C.; Villaume, S.; Guise, J.F.; Abou-Mansour, E.; Lebrun, M.H.; Fontaine, F. Diversity of Neofusicoccum parvum for the production of the phytotoxic metabolites (-)-terremutin and (R)-mellein. J. Fungi 2022, 8, 319. [Google Scholar] [CrossRef]
- Bénard-Gellon, M.; Farine, S.; Goddard, M.L.; Schmitt, M.; Stempien, E.; Pensec, F.; Laloue, H.; Mazet-Kieffer, F.; Fontaine, F.; Larignon, P.; et al. Toxicity of extracellular proteins from Diplodia seriata and Neofusicoccum parvum involved in grapevine Botryosphaeria dieback. Protoplasma 2015, 252, 679–687. [Google Scholar] [CrossRef]
- Cimmino, A.; Cinelli, T.; Evidente, M.; Masi, M.; Mugnai, L.; Silva, M.A.; Michereff, S.J.; Surico, G.; Evidente, A. Phytotoxic fungal exopolysaccharides produced by fungi involved in grapevine trunk diseases. Nat. Prod. Commun. 2016, 10, 1481–1484. [Google Scholar] [CrossRef]
- Andolfi, A.; Maddau, L.; Cimmino, A.; Linaldeddu, B.T.; Basso, S.; Deidda, A.; Serra, S.; Evidente, A. Lasiojasmonates A–C, three jasmonic acid esters produced by Lasiodiplodia sp., a grapevine pathogen. Phytochemistry 2014, 103, 145–153. [Google Scholar] [CrossRef]
- Yu, C.; Diao, Y.; Lu, Q.; Zhao, J.; Cui, S.; Xiong, X.; Lu, A.; Zhang, X.; Liu, H. Comparative genomics reveals evolutionary traits, mating strategies, and pathogenicity-related genes variation of Botryosphaeriaceae. Front. Microbiol. 2022, 13, 800981. [Google Scholar] [CrossRef]
- Drula, E.; Garron, M.L.; Dogan, S.; Lombard, V.; Henrissat, B.; Terrapon, N. The carbohydrate-active enzyme database: Functions and literature. Nucleic Acids Res. 2022, 50, D571–D577. [Google Scholar] [CrossRef]
- Xu, M.; Liu, C.L.; Luo, J.; Qi, Z.; Yan, Z.; Fu, Y.; Wei, S.S.; Tang, H. Transcriptomic de novo analysis of pitaya (Hylocereus polyrhizus) canker disease caused by Neoscytalidium dimidiatum. BMC Genom. 2019, 20, 10. [Google Scholar] [CrossRef]
- Wang, M.; Wang, Z.; Wei, S.; Xie, J.; Huang, J.; Li, D.; Hu, W.; Li, H.; Tang, H. Identification of RT-qPCR reference genes suitable for gene function studies in the pitaya canker disease pathogen Neoscytalidium dimidiatum. Sci. Rep. 2022, 12, 22357. [Google Scholar] [CrossRef]
- Arkam, M.; Alves, A.; Lopes, A.; Cechova, J.; Pokluda, R.; Eichmeier, A.; Zitouni, A.; Mahamedi, A.E.; Berraf-Tebbal, A. Diversity of Botryosphaeriaceae causing grapevine trunk diseases and their spatial distribution under different climatic conditions in Algeria. Eur. J. Plant Pathol. 2021, 161, 933–952. [Google Scholar] [CrossRef]
- Xian, X.; Lin, S.; Zhu, G.; Wei, X.; Qin, W.; Zhong, Y. Indoor virulence and field effects of fungicides on pitaya canker. J. South. Agric. 2018, 49, 1338–1345. [Google Scholar]
- Noegrahati, S.; Sulasmi, S.; Hemadi, E.; Asviastuti, S. Dissipation pattern of azoxystrobin and difenoconazole in red dragon fruit (Hylocereus polyrhizus) cultivated in Indonesian highland (West Java) and coastal area (D.I. Jogyakarta) and its implication for dietary risk assessment. Food Qual. Saf. 2019, 3, 99–106. [Google Scholar] [CrossRef]
- Eraslan Sür, A.; Oksal, E. In vitro efficiency of some fungicides against Neoscytalidium dimidiatum (Penz.) Crous and Slippers causing sudden shoot dry on apricot trees. Turk. J. Agric.-Food Sci. Technol. 2021, 9, 797–802. [Google Scholar] [CrossRef]
- Riska, J.; Budiyanti, T.; Husada, E.D.; Indriyani, N.L.P.; Hadiati, S.; Muas, I.; Mansyah, E. Stem canker of dragon fruit (Hylocereus polyrhizus): Neoscytalidium sp. is a pathogen of the disease and its control using sodium salt. Plant Protect. Sci. 2023, 59, 245–255. [Google Scholar] [CrossRef]
- Campos-Rivero, G.; Sánchez-Teyer, L.F.; De la Cruz-Arguijo, E.A.; Ramírez-González, M.S.; Larralde-Corona, C.P.; Narváez-Zapata, J.A. Bioprospecting for fungi with potential pathogenic activity on leaves of Agave tequilana Weber var. Azul. J. Phytopathol. 2019, 167, 283–294. [Google Scholar] [CrossRef]
- Taguiam, J.D.; Evallo, E.; Bengoa, J.; Maghirang, R.; Balendres, M.A. Susceptibility of the three dragon fruit species to stem canker and growth inhibition of Neoscytalidium dimidiatum by chemicals. J. Plant Pathol. 2020, 102, 1077–1084. [Google Scholar] [CrossRef]
- Ratanaprom, S.; Nakkanong, K.; Nualsri, C.; Jiwanit, P.; Rongsawat, T.; Woraathakorn, N. Overcoming encouragement of dragon fruit plant (Hylocereus undatus) against stem brown spot disease caused by Neoscytalidium dimidiatum using Bacillus subtilis combined with sodium bicarbonate. Plant Pathol. J. 2021, 37, 205–214. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, R.; Yuan, Z.; Chen, P. Biological prevention and control of pitaya fruit canker disease using endophytic fungi isolated from papaya. Arch. Microbiol. 2021, 203, 4033–4040. [Google Scholar] [CrossRef]
- Al Hamad, B.M.; Al Raish, S.M.; Ramadan, G.A.; Saeed, E.E.; Alameri, S.S.A.; Al Senaani, S.S.; AbuQamar, S.F.; El-Tarabily, K.A. Effectiveness of augmentative biological control of Streptomyces griseorubens UAE2 depends on 1-aminocyclopropane-1-carboxylic acid deaminase activity against Neoscytalidium dimidiatum. J. Fungi 2021, 7, 885. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Chen, X.; Xie, L.; Zhang, Y.; Zeng, F.; Long, Y.; Ren, L.; Qi, X.; Wei, J. Biocontrol potential of lipopeptides produced by Paenibacillus polymyxa AF01 against Neoscytalidium dimidiatum in pitaya. Front. Microbiol. 2023, 14, 1188722. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Gao, Y.; Li, H.; Wang, D.; Li, J.; Hu, X.; Huang, X.; Lin, M.; Tang, Y.; Liu, Z. Identification and characterization of biocontrol agent Lysinibacillus boronitolerans P42 against Cerrena unicolor that causes root rot of arecanut palm. Arch. Microbiol. 2023, 205, 157. [Google Scholar] [CrossRef] [PubMed]
- Travadon, R.; Lawrence, D.P.; Li, S.; Trouillas, F. Evaluation of biological control agents for the protection of almond pruning wounds against infection by fungal canker pathogens. Phytopathology 2023, 113, 1417–1427. [Google Scholar] [CrossRef]
- Du, B.D.; Ngoc, D.T.B.; Thang, N.D.; Tuan, L.N.A.; Thach, B.D.; Hien, N.Q. Synthesis and in vitro antifungal efficiency of alginate-stabilized Cu2O-Cu nanoparticles against Neoscytalidium dimidiatum causing brown spot disease on dragon fruit plants (Hylocereus undatus). Vietnam J. Chem. 2019, 57, 318–323. [Google Scholar] [CrossRef]
- Tuan, L.N.A.; Du, B.D.; Ha, L.D.T.; Dzung, L.T.K.; Van Phu, D.; Hien, N.Q. Induction of chitinase and brown spot disease resistance by oligochitosan and nanosilica–oligochitosan in dragon fruit plants. Agric. Res. 2019, 8, 184–190. [Google Scholar] [CrossRef]
- Acay, H.; Yildirim, A.; Güney, I.G.; Derviş, S. Morchella esculenta-based chitosan bionanocomposites: Evaluation as an antifungal agent. J. Food Process. Preserv. 2022, 46, e17117. [Google Scholar] [CrossRef]
- Hashem, A.H.; Abdelaziz, A.M.; Hassanin, M.M.H.; Al-Askar, A.A.; AbdElgawad, H.; Attia, M.S. Potential impacts of clove essential oil nanoemulsion as bio fungicides against Neoscytalidium blight disease of Carum carvi L. Agronomy 2023, 13, 1114. [Google Scholar] [CrossRef]
- Duong, N.L.; Nguyen, V.M.; Tran, T.A.N.; Phan, T.D.T.; Tran, T.B.Y.; Do, B.L.; Phung Anh, N.; Nguyen, T.A.T.; Ho, T.G.; Nguyen, T. Durian shell-mediated simple green synthesis of nanocopper against plant pathogenic fungi. ACS Omega 2023, 8, 10968–10979. [Google Scholar] [CrossRef]
- Bragard, C.; Baptista, P.; Chatzivassiliou, E.; Di Serio, F.; Gonthier, P.; Jaques Miret, J.A.; Justesen, A.F.; MacLeod, A.; Magnusson, C.S.; Milonas, P. Pest categorisation of Neoscytalidium dimidiatum. EFSA J. 2023, 21, e08001. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Derviş, S.; Özer, G. Plant-Associated Neoscytalidium dimidiatum—Taxonomy, Host Range, Epidemiology, Virulence, and Management Strategies: A Comprehensive Review. J. Fungi 2023, 9, 1048. https://doi.org/10.3390/jof9111048
Derviş S, Özer G. Plant-Associated Neoscytalidium dimidiatum—Taxonomy, Host Range, Epidemiology, Virulence, and Management Strategies: A Comprehensive Review. Journal of Fungi. 2023; 9(11):1048. https://doi.org/10.3390/jof9111048
Chicago/Turabian StyleDerviş, Sibel, and Göksel Özer. 2023. "Plant-Associated Neoscytalidium dimidiatum—Taxonomy, Host Range, Epidemiology, Virulence, and Management Strategies: A Comprehensive Review" Journal of Fungi 9, no. 11: 1048. https://doi.org/10.3390/jof9111048
APA StyleDerviş, S., & Özer, G. (2023). Plant-Associated Neoscytalidium dimidiatum—Taxonomy, Host Range, Epidemiology, Virulence, and Management Strategies: A Comprehensive Review. Journal of Fungi, 9(11), 1048. https://doi.org/10.3390/jof9111048