
Isolation, Identification, and Evaluation of the Predatory Activity of Chinese Arthrobotrys Species towards Economically Important Plant-Parasitic Nematodes

 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Nematode Cultures
2.2. Sample Collection and Isolation of Strains
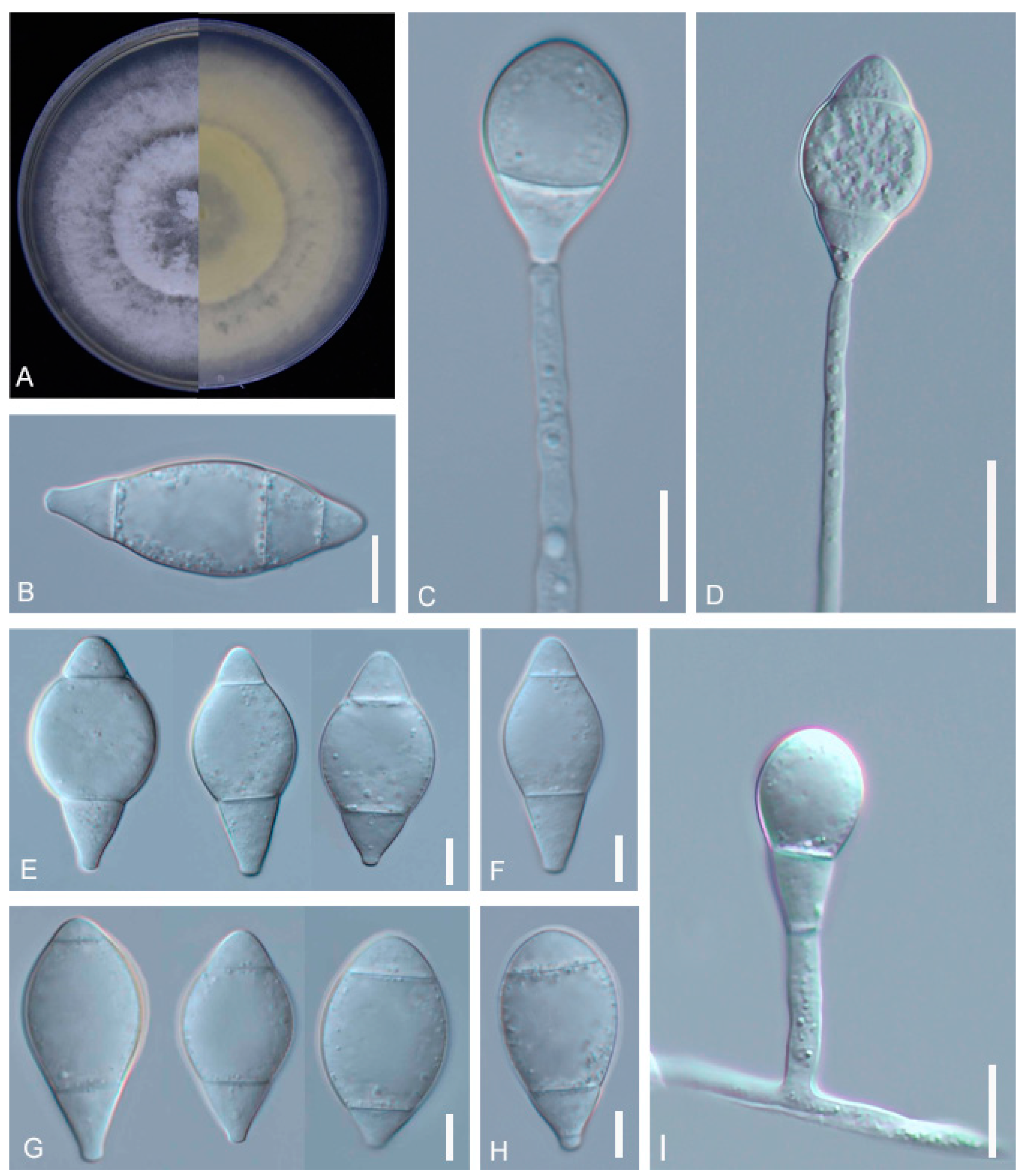
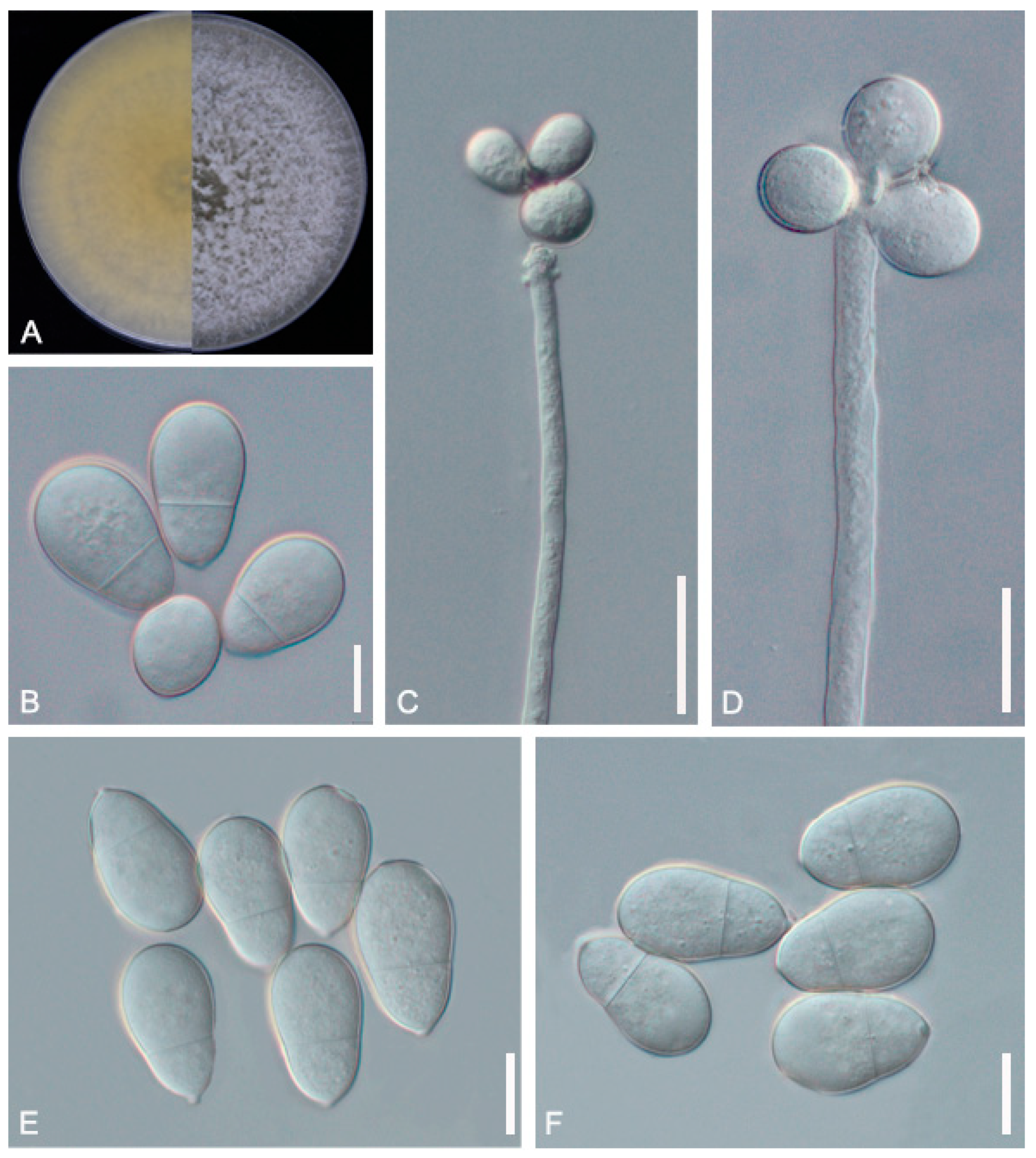
2.3. Morphological Observations
2.4. DNA Extraction, PCR, and Sequencing
2.5. Phylogenetic Analysis
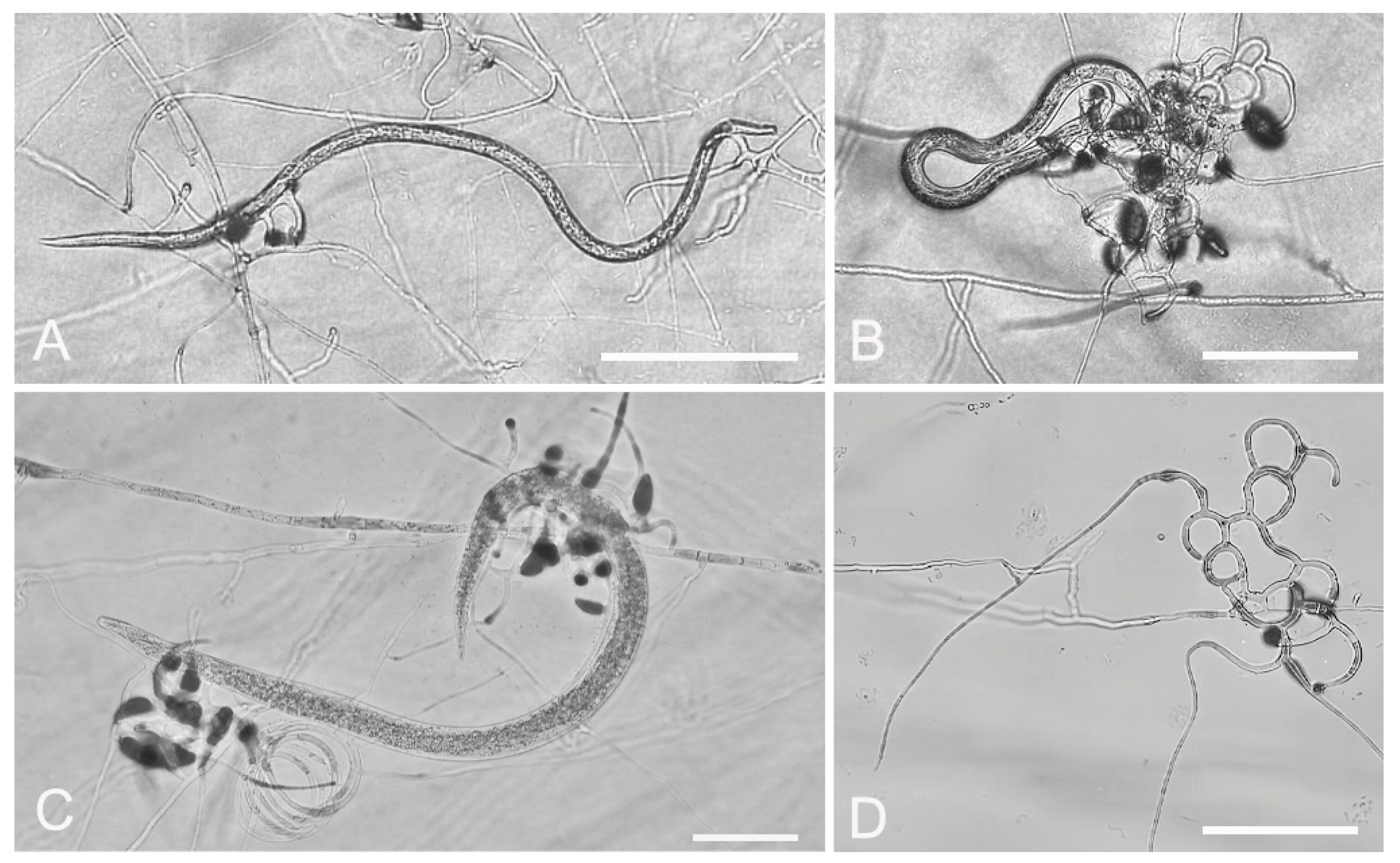
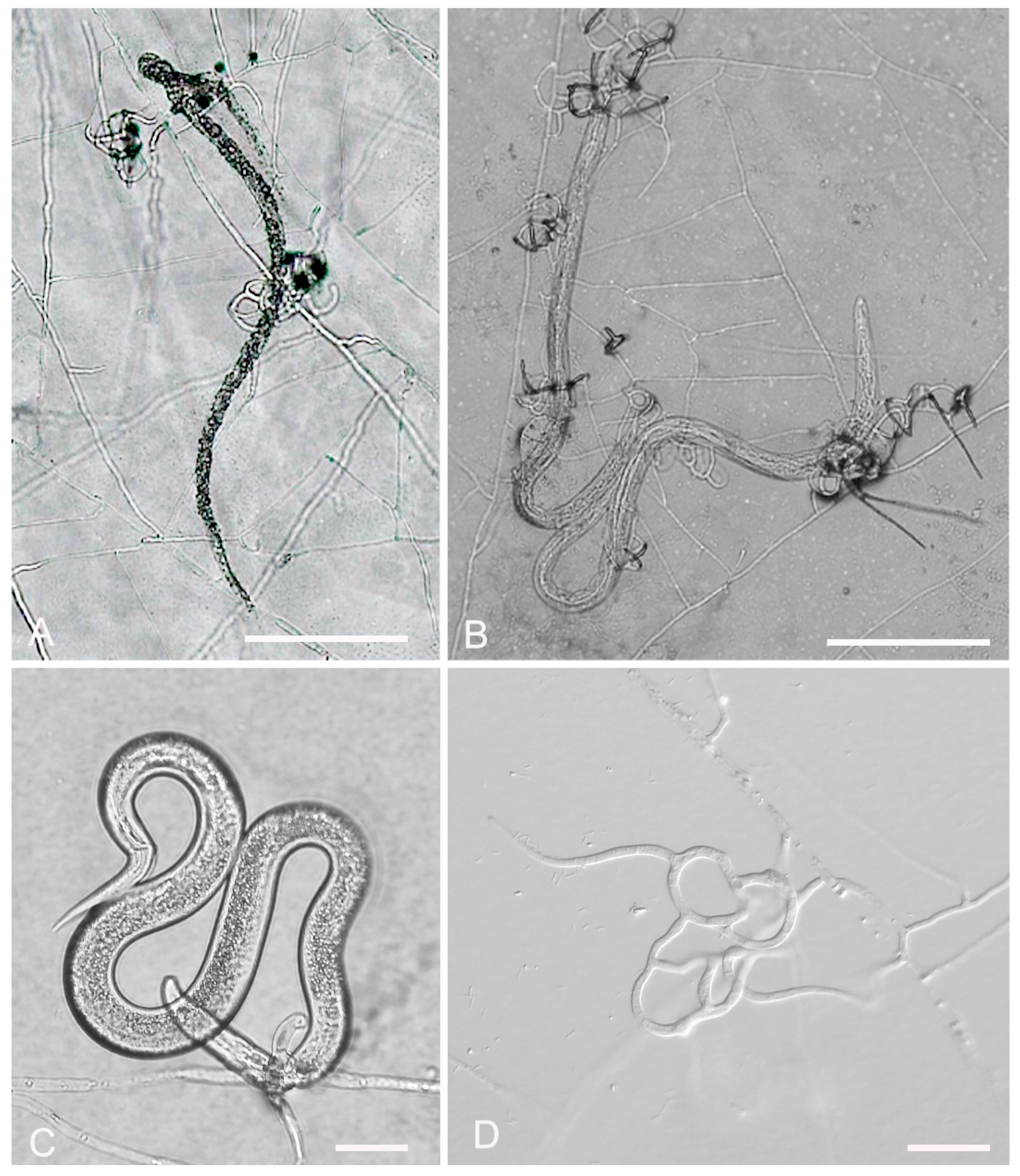
2.6. In Vitro Predatory Activity of the Fungal Isolates against Nematodes
3. Results
3.1. Identification of the Fungal Isolates
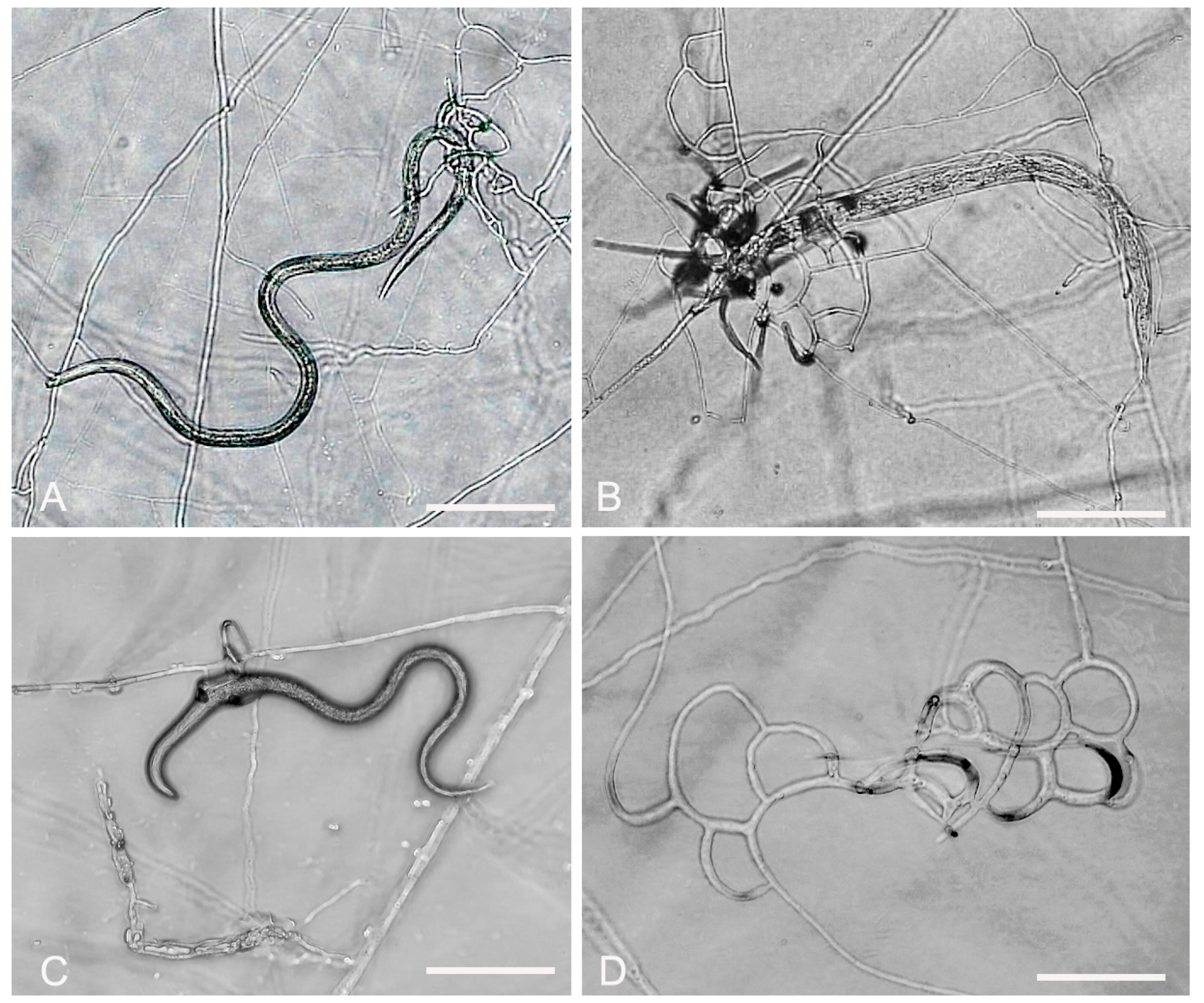
3.2. Evaluation of Fungal Predatory Activity vs. Plant Parasitic Nematodes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fuller, V.L.; Lilley, C.J.; Urwin, P.E. Nematode resistance. New Phytol. 2008, 180, 27–44. [Google Scholar] [CrossRef]
- Kumar, K.K.; Arthurs, S. Recent advances in the biological control of citrus nematodes: A review. Biol. Control 2021, 157, 104593. [Google Scholar] [CrossRef]
- Mesa-Valle, C.M.; Garrido-Cardenas, J.A.; Cebrian-Carmona, J.; Talavera, M.; Manzano-Agugliaro, F. Global Research on Plant Nematodes. Agronomy 2020, 10, 1148. [Google Scholar] [CrossRef]
- Ali, M.A.; Abbas, A.; Azeem, F.; Javed, N.; Bohlmann, H. Plant-Nematode Interactions: From Genomics to Metabolomics. Int. J. Agric. Biol. 2015, 17, 1071–1082. [Google Scholar] [CrossRef]
- Webster, J.M. The Host-Parasite Relationships of Plant-Parasitic Nematodes. In Advances in Parasitology; Dawes, B., Ed.; Academic Press: Cambridge, MA, USA, 1969; Volume 7, pp. 1–40. [Google Scholar]
- Hu, W.; Strom, N.; Haarith, D.; Chen, S.; Bushley, K.E. Mycobiome of Cysts of the Soybean Cyst Nematode Under Long Term Crop Rotation. Front. Microbiol. 2018, 9, 386. [Google Scholar] [CrossRef]
- Roeber, F.; Jex, A.R.; Gasser, R.B. Advances in the diagnosis of key gastrointestinal nematode infections of livestock, with an emphasis on small ruminants. Biotechnol. Adv. 2013, 31, 1135–1152. [Google Scholar] [CrossRef]
- Ploeg, A.; Stoddard, S.; Becker, J. Control of Meloidogyne incognita in sweetpotato with fluensulfone. J. Nematol. 2019, 51, e2019-18. [Google Scholar] [CrossRef]
- Dong, L.; Zhang, K. Microbial control of plant-parasitic nematodes: A five-party interaction. Plant Soil. 2006, 288, 31–45. [Google Scholar] [CrossRef]
- Fan, M.; Shen, J.; Yuan, L.; Jiang, R.; Chen, X.; Davies, W.J.; Zhang, F. Improving crop productivity and resource use efficiency to ensure food security and environmental quality in China. J. Exp. 2012, 63, 13–24. [Google Scholar] [CrossRef]
- Tang, X.; Zhu, B.; Katou, H. A review of rapid transport of pesticides from sloping farmland to surface waters: Processes and mitigation strategies. J. Environ. Sci. 2012, 24, 351–361. [Google Scholar] [CrossRef]
- Niciura, S.C.M.; Cruvinel, G.G.; Moraes, C.V.; Chagas, A.C.S.; Esteves, S.N.; Benavides, M.V.; Amarante, A.F.T. In vivo selection for Haemonchus contortus resistance to monepantel. J. Helminthol. 2020, 94, e46. [Google Scholar] [CrossRef]
- Prado, A.; Airoldi, C. Effect of the pesticide 2, 4-D on microbial activity of the soil monitored by microcalorimetry. Thermochim. Acta 2000, 349, 17–22. [Google Scholar] [CrossRef]
- Singh, B.; Devindrappa; Hazra, K.K.; Singh, U.; Gupta, S. Eco–friendly management of Meloidogyne javanica in chickpea (Cicer arietinum L.) using organic amendments and bio–control agent. J. Clean. 2020, 257, 120542. [Google Scholar] [CrossRef]
- Leng, P.; Zhang, Z.; Pan, G.; Zhao, M. Applications and development trends in biopesticides. Afr. J. Biotechnol. 2011, 10, 19864–19873. [Google Scholar] [CrossRef]
- Wilschut, R.A.; Magnee, K.J.H.; Geisen, S.; van der Putten, W.H.; Kostenko, O. Plant population and soil origin effects on rhizosphere nematode community composition of a range-expanding plant species and a native congener. Oecologia 2020, 194, 237–250. [Google Scholar] [CrossRef]
- Doolotkeldieva, T.; Bobushova, S.; Muratbekova, A.; Schuster, C.; Leclerque, A. Isolation, Identification, and Characterization of the Nematophagous Fungus Arthrobotrys oligospora from Kyrgyzstan. Acta Parasitol. 2021, 66, 1349–1365. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, K.Q.; Hyde, K. The Ecology of Nematophagous Fungi in Natural Environments. In Nematode-Trapping Fungi; Zhang, K.-Q., Hyde, K.D., Eds.; Springer: Amsterdam, The Netherlands, 2014; Volume 23, pp. 211–229. [Google Scholar] [CrossRef]
- Schouten, A. Mechanisms Involved in Nematode Control by Endophytic Fungi. Annu. Rev. Phytopathol. 2016, 54, 121–142. [Google Scholar] [CrossRef]
- Zhang, K.Q.; Hyde, K.D. Nematode-Trapping Fungi; Springer: Amsterdam, The Netherlands, 2014; Volume 23. [Google Scholar] [CrossRef]
- Herrera-Estrella, A.; Casas-Flores, S.; Kubicek, C.P. 13 Nematophagous Fungi. In Environmental and Microbial Relationships; Druzhinina, I.S., Kubicek, C.P., Eds.; Springer International Publishing: Berlin/Heidelberg, Germany, 2016; pp. 247–267. [Google Scholar] [CrossRef]
- Zopf, W.F. Zur Kenntniss der Infections-Krankheiten niederer Thiere und Pflanzen. Acad. Nat. 1888, 52, 314–376. [Google Scholar]
- Niu, X.M.; Zhang, K.Q. Arthrobotrys oligospora: A model organism for understanding the interaction between fungi and nematodes. Mycology 2011, 2, 59–78. [Google Scholar] [CrossRef]
- Nordbring-Hertz, B. Morphogenesis in the nematode-trapping fungus Arthrobotrys oligospora–An extensive plasticity of infection structures. Mycologist 2004, 18, 125–133. [Google Scholar] [CrossRef]
- Pires, D.; Vicente, C.S.L.; Menéndez, E.; Faria, J.M.S.; Rusinque, L.; Camacho, M.J.; Inácio, M.L. The Fight against Plant-Parasitic Nematodes: Current Status of Bacterial and Fungal Biocontrol Agents. Pathogens 2022, 11, 1178. [Google Scholar] [CrossRef]
- Futai, K. Pine wood nematode, Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 2013, 51, 61–83. [Google Scholar] [CrossRef]
- Tülek, A.; Kepenekci, İ.; Çobanoğlu, S.; Hekimhan, H.; Devran, Z.; Melik, B.; Elekcioğlu, H. A new culturing method for the rice white tip nematode, Aphelenchoides besseyi Christie, 1942, on carrot discs. Russ. J. Nematol 2009, 17, 143–144. [Google Scholar]
- Farrell, F.C.; Jaffee, B.A.; Strong, D.R. The nematode-trapping fungus Arthrobotrys oligospora in soil of the Bodega marine reserve: Distribution and dependence on nematode-parasitized moth larvae. Soil Biol. Biochem. 2006, 38, 1422–1429. [Google Scholar] [CrossRef]
- Xue, Y.J.; Li, E.L.; Jing, C.X.; Ma, L.; Cai, K.Z. Isolation, Identification and Characterization of the Nematophagous Fungus Arthrobotrys (Monacrosporium) Sinense from China. Acta Parasit. 2018, 63, 325–332. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, J.; Innis, M.; Gelfand, D.; Sninsky, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In Pcr Protocols: A Guide to Methods and Applications; Academic Press: Cambridge, MA, USA, 1990; Volume 31, pp. 315–322. [Google Scholar]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef]
- Yang, Y.; Yang, E.; An, Z.; Liu, X. Evolution of nematode-trapping cells of predatory fungi of the Orbiliaceae based on evidence from rRNA-encoding DNA and multiprotein sequences. Proc. Natl. Acad. Sci. USA 2007, 104, 8379–8384. [Google Scholar] [CrossRef]
- Vu, D.; Groenewald, M.; Vries, M.; Gehrmann, T.; Stielow, B.; Eberhardt, U.; Al-Hatmi, A.; Groenewald, J.Z.; Cardinali, G.; Houbraken, J.; et al. Large-scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 2019, 92, 135–154. [Google Scholar] [CrossRef]
- Zhang, F.; Boonmee, S.; Bhat, J.D.; Xiao, W.; Yang, X.Y. New Arthrobotrys Nematode-Trapping Species (Orbiliaceae) from Terrestrial Soils and Freshwater Sediments in China. J. Fungi 2022, 8, 671. [Google Scholar] [CrossRef]
- Li, J.; Liu, Y.; Zhu, H.; Zhang, K.Q. Phylogenic analysis of adhesion related genes Mad1 revealed a positive selection for the evolution of trapping devices of nematode-trapping fungi. Sci. Rep. 2016, 6, 22609. [Google Scholar] [CrossRef]
- Kano, S.; Aimi, T.; Masumoto, S.; Kitamoto, Y.; Morinaga, T. Physiology and Molecular Characteristics of a Pine Wilt Nematode-Trapping Fungus, Monacrosporium megalosporum. Curr. Microbiol. 2004, 49, 158–164. [Google Scholar] [CrossRef]
- Ocampo-Gutierrez, Y.; Hernandez-Velazquez, M.; Aguilar-Marcelino, L.; Cardoso-Taketa, A.; Zamilpa, A.; Lopez-Arellano, M.E.; Gonzalez-Cortazar, M.; Hernandez-Romano, J.; Reyes-Estebanez, M.; Mendoza-de, G.P. Morphological and molecular characterization, predatory behaviour and effect of organic extracts of four nematophagous fungi from Mexico. Fungal Ecol. 2021, 49, 101004. [Google Scholar] [CrossRef]
- Hastuti, L.; Berliani, K.; Budi Mulya, M.; Hartanto, A.; Pahlevi, S. Arthrobotrys Sinensis (Orbiliaceae Orbiliales), a New Record of Nematode-Trapping Fungal Species for Sumatra, Indonesia. In Proceedings of the 2nd Universitas Kuningan International Conference on System, Engineering, and Technology, UNISET 2022, Kuningan, Indonesia, 2 December 2022. [Google Scholar] [CrossRef]
- Zhang, F.; Boonmee, S.; Yang, Y.-Q.; Zhou, F.-P.; Xiao, W.; Yang, X.-Y. Arthrobotrys Blastospora Sp. Nov. (Orbiliomycetes): A Living Fossil Displaying Morphological Traits of Mesozoic Carnivorous Fungi. J. Fungi 2023, 9, 451. [Google Scholar] [CrossRef]
- Zhang, H.; Wei, Z.; Liu, X.; Zhang, J.; Diao, G. Growth and decline of arboreal fungi that prey on Bursaphelenchus xylophilus and their predation rate. J. For. Res. 2022, 33, 699–709. [Google Scholar] [CrossRef]
- Soliman, M.S.; El-Deriny, M.M.; Ibrahim, D.S.S.; Zakaria, H.; Ahmed, Y. Suppression of root-knot nematode Meloidogyne incognita on tomato plants using the nematode trapping fungus Arthrobotrys oligospora Fresenius. J. Appl. Microbiol. 2021, 131, 2402–2415. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Strain Number | GenBank Accession Number | Reference | |
|---|---|---|---|---|
| ITS | TEF | |||
| A. arthrobotryoides | CBS 119.54 | MH857262 | _ | [33] |
| A. conoides | 670 | AY773455 | AY773397 | [32] |
| A. dianchiensis | 1.00571 | MH179720 | _ | [34] |
| A. elegans | 1.00027 | MH179688 | _ | Unpublished |
| A. eryuanensis | YXY45 | ON808616 | ON809547 | [34] |
| A. flagrans | 1.01471 | MH179741 | MH179583 | [34] |
| A. indica | YMF1.01845 | KT932086 | _ | [35] |
| A. jinpingensis | CGMCC3.20896 | OM855569 | OM850311 | [34] |
| A. megalosporus | TWF800 | MN013995 | _ | [34] |
| A. microscaphoides | YMF1.00028 | MF948395 | MF948552 | Unpublished |
| A. musiformis | 1.03481 | MH179783 | MH179607 | [34] |
| A. oligospora | 920 | AY773462 | AY773404 | [32] |
| A. pseudoclavata | 1130 | AY773446 | AY773388 | [32] |
| A. shuifuensis | CGMCC3.19716 | MT612334 | OM850306 | [34] |
| A. sinensis | 105-1 | AY773445 | AY773387 | [32] |
| A. xiangyunensis | YXY10-1 | MK537299 | _ | [34] |
| A. zhaoyangensis | CGMCC3.20944 | OM855568 | OM850310 | [34] |
| A. zhaoyangensis | YXY86 | ON808620 | ON809551 | [34] |
| A. thaumasia | 917 | AY773461 | AY773403 | [32] |
| A. reticulata | CBS 550.63 | MH858355 | _ | [32] |
| Dactylellina mammillata | CBS229.54 | AY902794 | DQ999843 | [34] |
| D. yushanensis | CGMCC3.19713 | MK372061 | MN915113 | Unpublished |
| Genus/Species | Conidia Size | Reference | |
|---|---|---|---|
| Mean (μm) | Range (μm) | ||
| A. megalosporus | Length: — | 40–75 | [36] |
| Width: — | 18–35 | ||
| A. megalosporus | Length: 29.4 | 24.32–41.1 | GUCC220044 |
| Width: 15 | 11.69–17.5 | ||
| A. oligospora | Length: 21.4 | 17.5–25.6 | [37] |
| Width: 11.8 | 11.4–12.5 | ||
| A. oligospora | Length: 23.5 | 20.92–27.3 | GUCC220045 |
| Width: 12.8 | 11.88–14.4 | ||
| A. sinense | Length: 25.5 | 20.4–30 | [38] |
| Width: 20 | 18–22 | ||
| A. sinense | Length: 25.9 | 20.50–33.1 | GUCC220046 |
| Width: 14.4 | 12.14–17.3 | ||
| Nematode | Treatment | 24 h Mortality (%) | 48 h Mortality (%) |
|---|---|---|---|
| A. besseyi | A. megalosporus | 83.3 ± 6.7 a | 99.7 ± 0.7 a |
| A. oligospora | 84.5 ± 6.5 a | 97.3 ± 0.9 a | |
| A. sinensis | 69 ± 6.7 b | 85.1 ± 5.5 b | |
| control | 0 | 0 | |
| B. xylophilus | A. megalosporus | 24 ± 3.2 b | 83 ± 5.3 b |
| A. oligospora | 51.1 ± 2.7 a | 97.3 ± 1.26 a | |
| A. sinensis | 52.3 ± 1.3 a | 68.3 ± 6.7 c | |
| control | 0 | 0 | |
| D. destructor | A. megalosporus | 3.4 ± 0.7 c | 21.1 ± 4.3 b |
| A. oligospora | 23.8 ± 5.5 a | 54.6 ± 5.0 a | |
| A. sinensis | 10.5 ± 2.8 b | 19.0 ± 0.7 ab | |
| control | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Yang, Z.; Jiang, Z.; Nizamani, M.M.; Zhang, H.; Liu, M.; Wei, S.; Wang, Y.; Li, K. Isolation, Identification, and Evaluation of the Predatory Activity of Chinese Arthrobotrys Species towards Economically Important Plant-Parasitic Nematodes. J. Fungi 2023, 9, 1125. https://doi.org/10.3390/jof9121125
Wu Y, Yang Z, Jiang Z, Nizamani MM, Zhang H, Liu M, Wei S, Wang Y, Li K. Isolation, Identification, and Evaluation of the Predatory Activity of Chinese Arthrobotrys Species towards Economically Important Plant-Parasitic Nematodes. Journal of Fungi. 2023; 9(12):1125. https://doi.org/10.3390/jof9121125
Chicago/Turabian StyleWu, Yan, Zaifu Yang, Zhaochun Jiang, Mir Muhammad Nizamani, Hui Zhang, Mingrui Liu, Shan Wei, Yong Wang, and Kaihuai Li. 2023. "Isolation, Identification, and Evaluation of the Predatory Activity of Chinese Arthrobotrys Species towards Economically Important Plant-Parasitic Nematodes" Journal of Fungi 9, no. 12: 1125. https://doi.org/10.3390/jof9121125
APA StyleWu, Y., Yang, Z., Jiang, Z., Nizamani, M. M., Zhang, H., Liu, M., Wei, S., Wang, Y., & Li, K. (2023). Isolation, Identification, and Evaluation of the Predatory Activity of Chinese Arthrobotrys Species towards Economically Important Plant-Parasitic Nematodes. Journal of Fungi, 9(12), 1125. https://doi.org/10.3390/jof9121125