



Generation, Transfer, and Loss of Alternative Oxidase Paralogues in the Aspergillaceae Family
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mining of Alternative Oxidase Genes, Intron–Exon Structure Conservation, and Gene Synteny
2.2. Maximum-Likelihood Phylogenetic Analysis
2.3. Confirmation of Expression with Extant RNA Sequence Reads
2.4. Expression Verification of aox Paralogous Genes
2.5. Isolation of Total RNA for cDNA Sequence Analyses
2.6. Polymerase Chain Reaction (PCR) and cDNA Sequence Determination
3. Results and Discussion
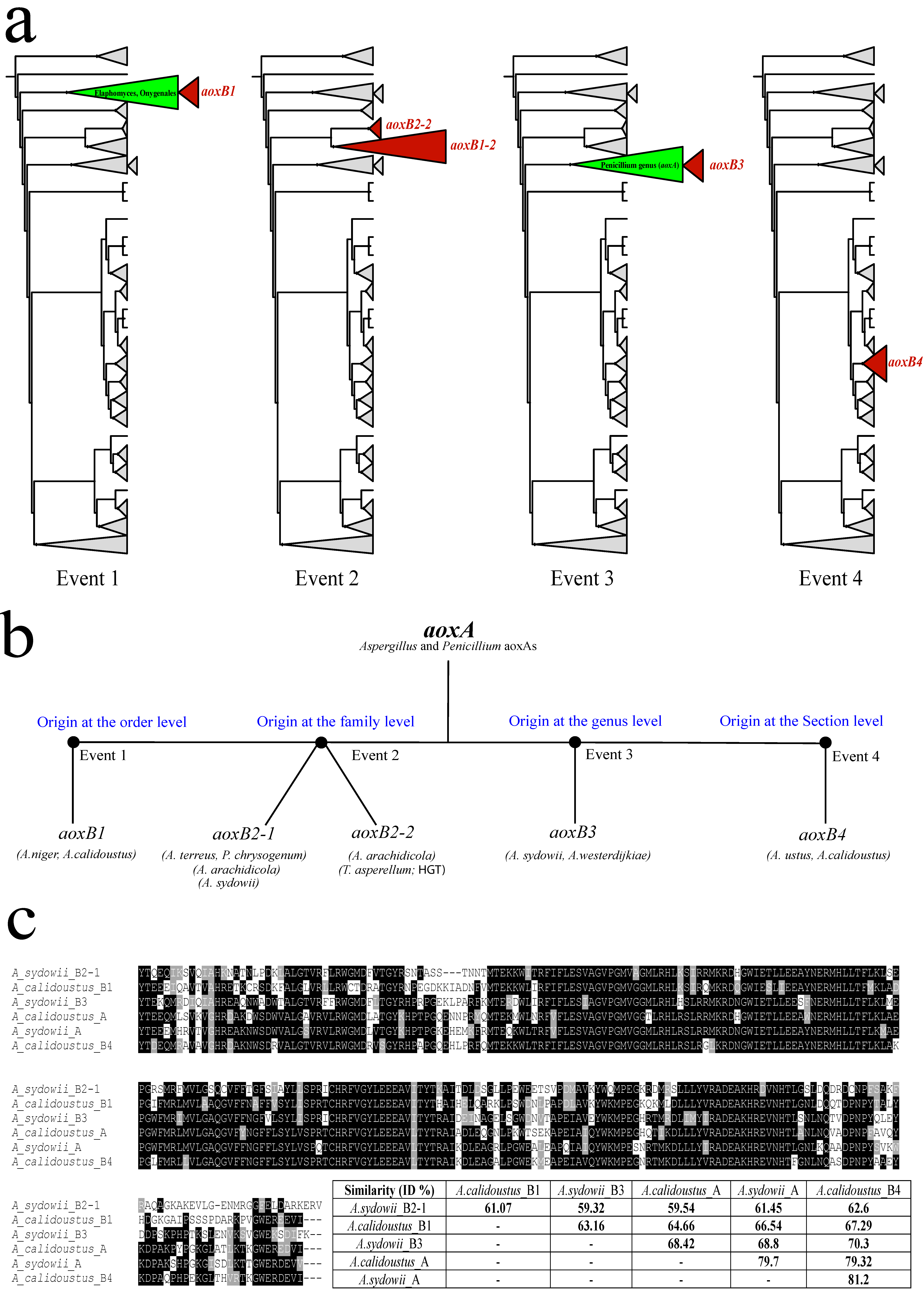
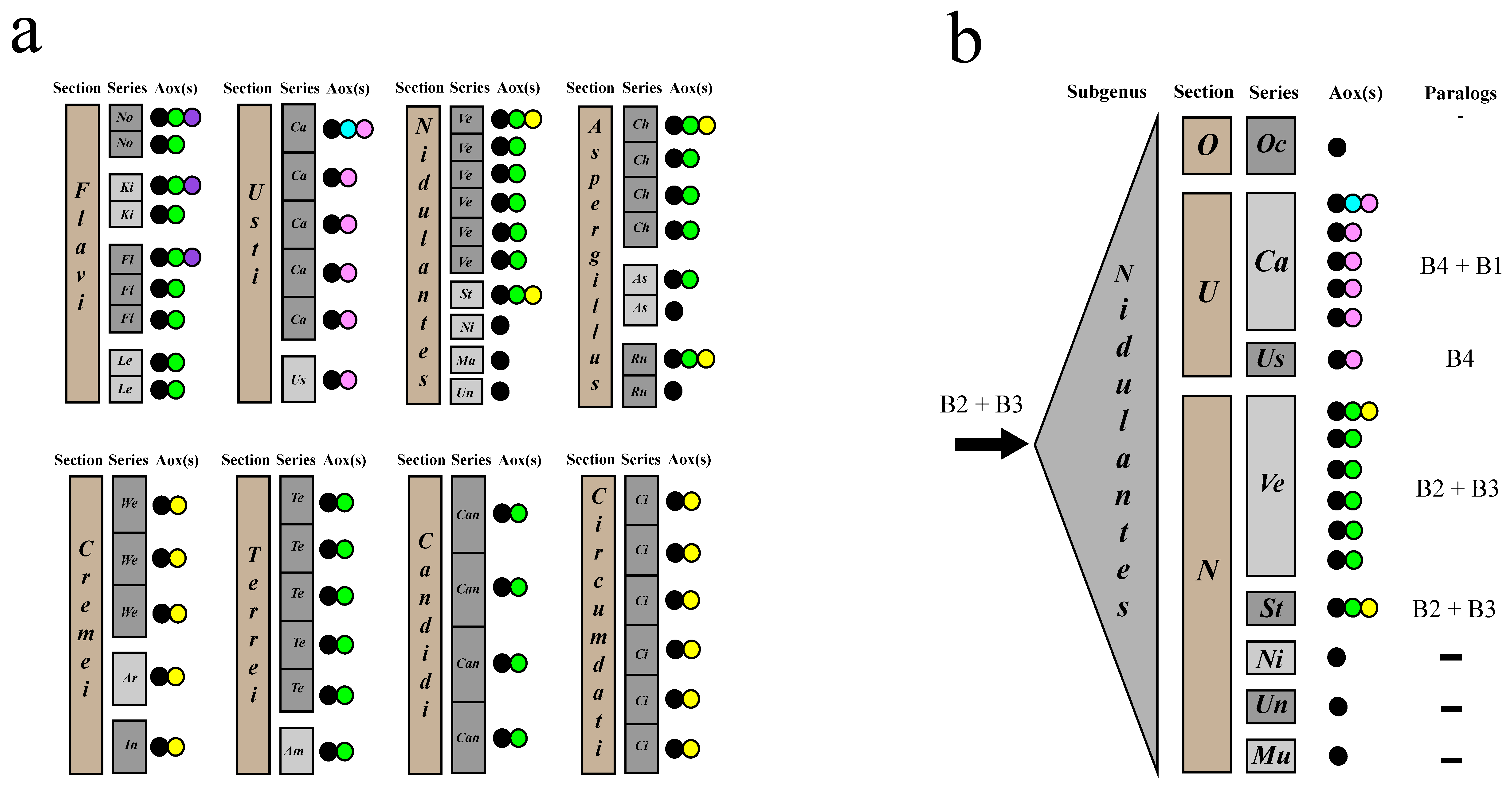
3.1. aox Paralogous Genes Have Been Generated Independently at Four Different Occasions in the Aspergillaceae
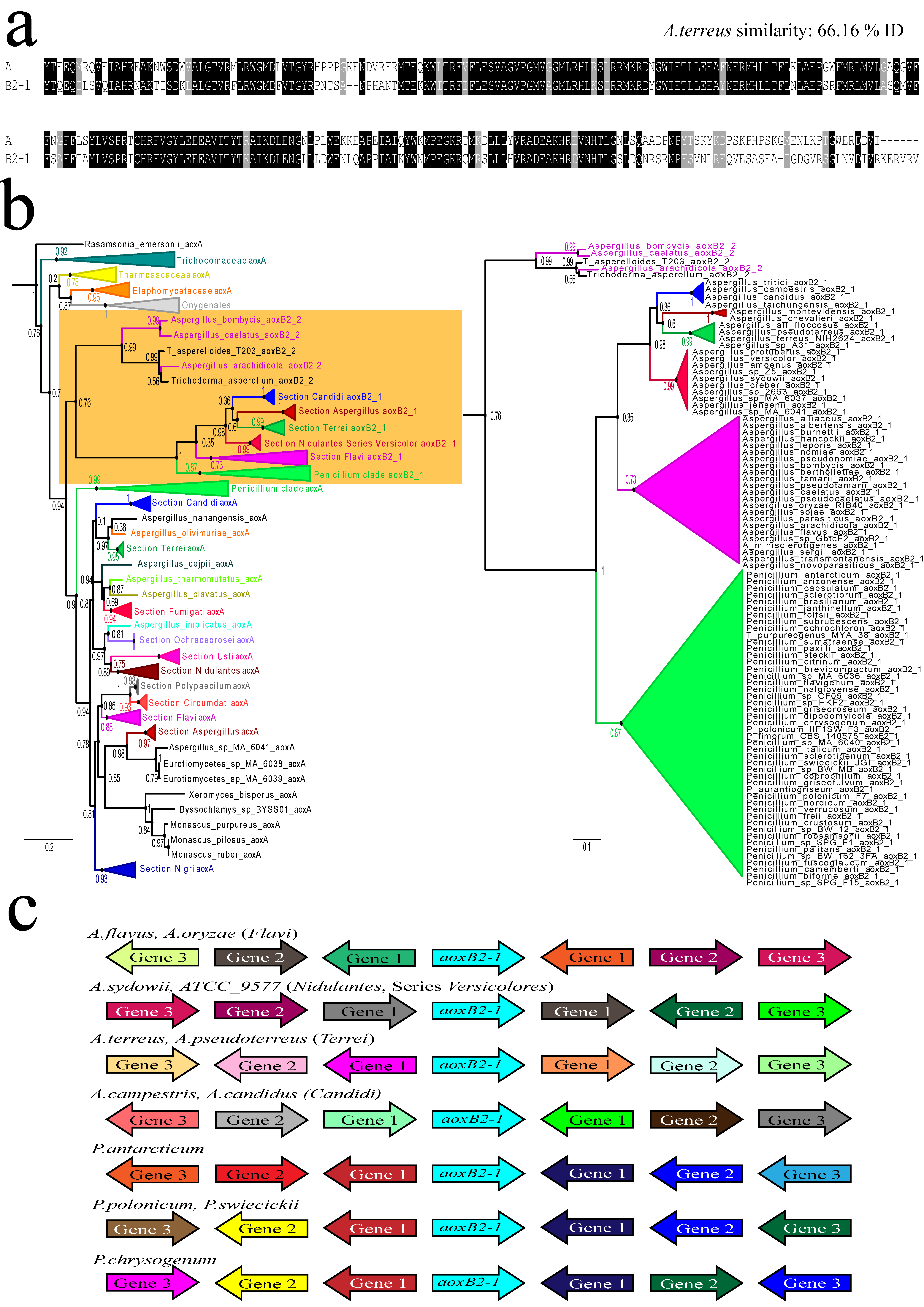
3.2. Ancient Gene Duplication in an Aspergillaceae Ancestor of Aspergillus and Penicillium
3.3. Lateral Transfer of a Fungal aox Gene between Species of Different Taxonomic Classes
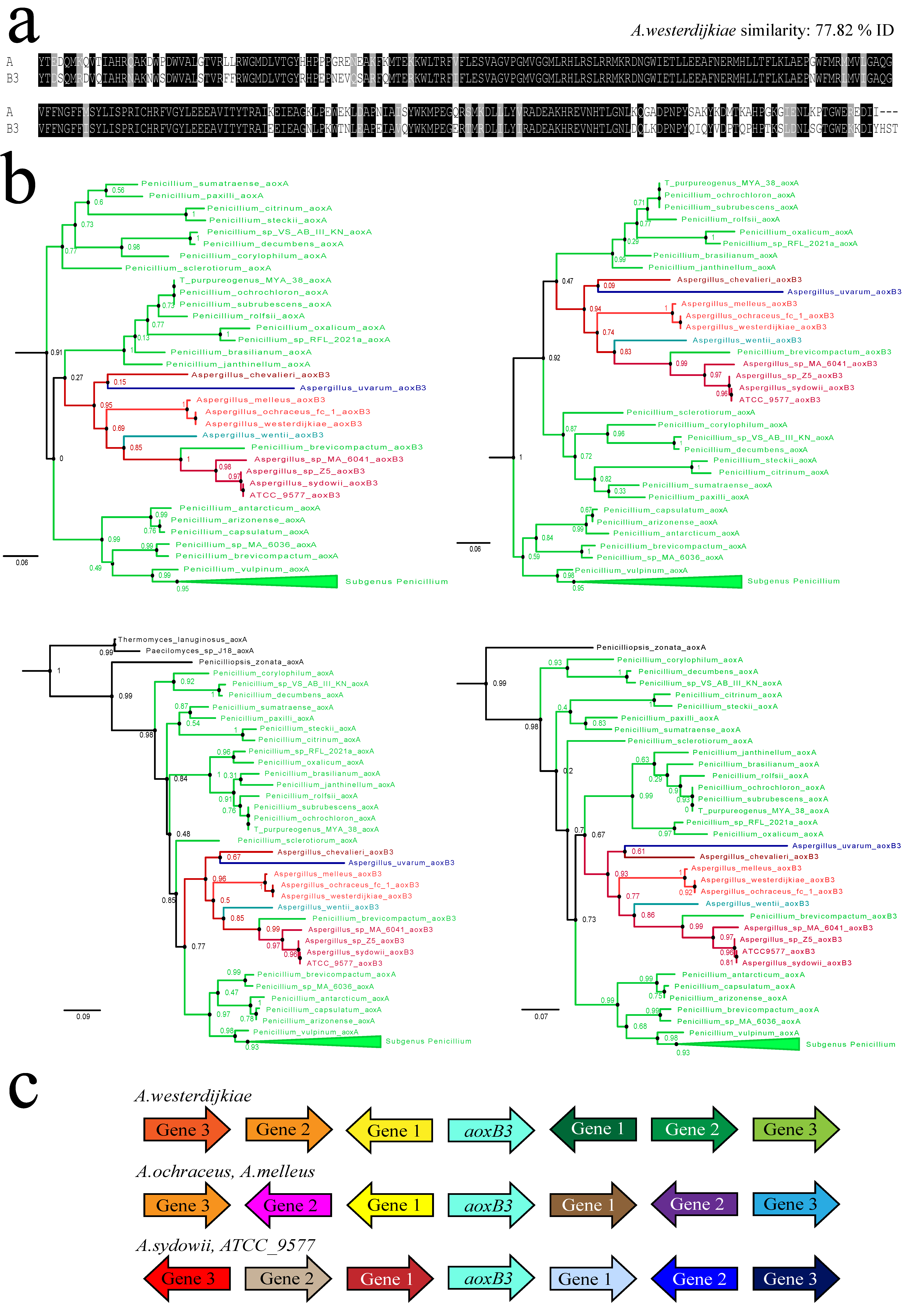
3.4. A Gene Duplication Seemingly Arising from within the Clade of Penicillium aoxA
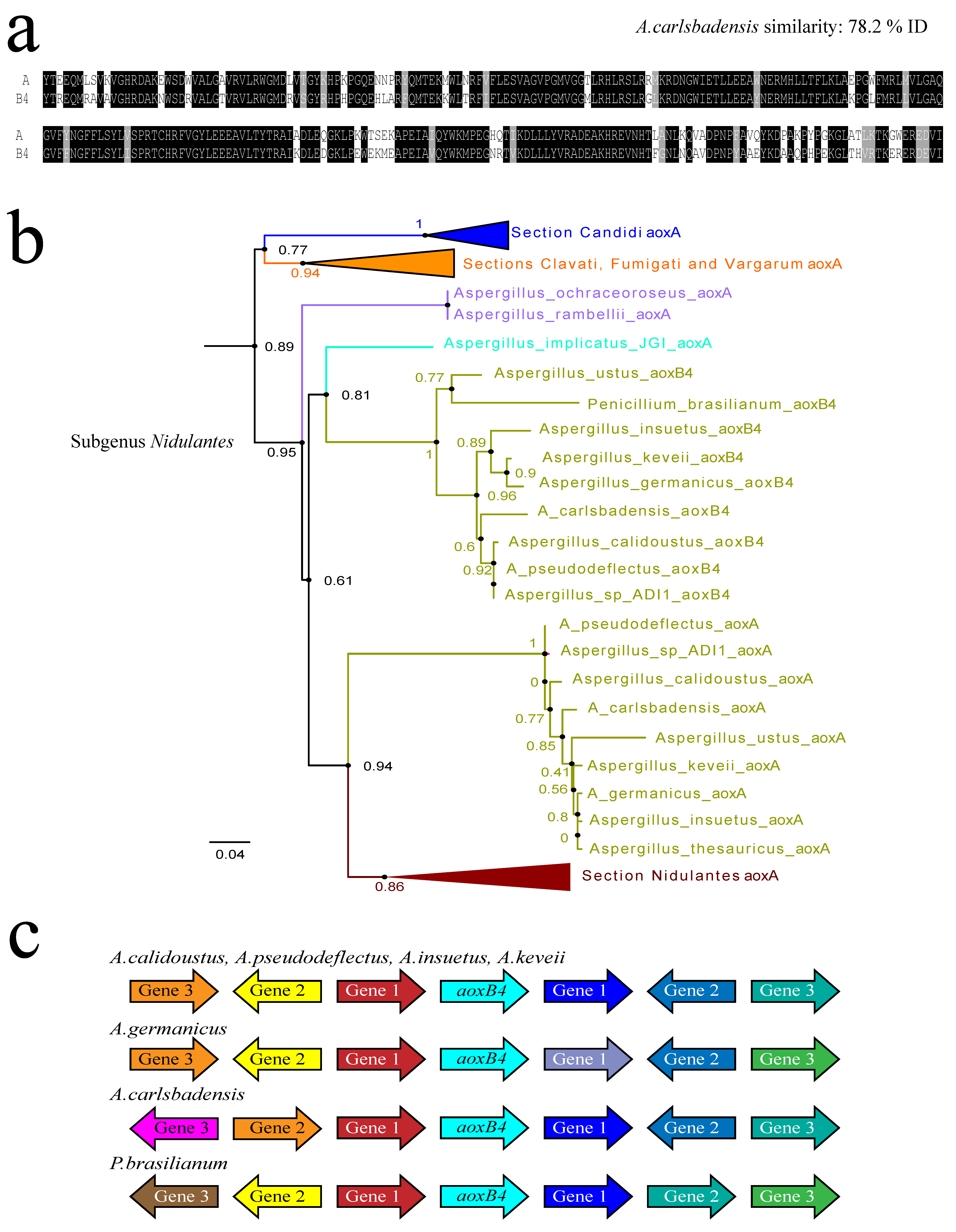
3.5. Recent Gene Duplication at the Basis of Section Usti (Subgenus Nidulantes)
3.6. Verification of the Expression of Alternative Oxidase Paralogous (aoxB) Genes
3.7. Different Patterns of aoxB Gene Loss in the Aspergillus Genus
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lambowitz, A.M.; Sabourin, J.R.; Bertrand, H.; Nickels, R.; McIntosh, L. Immunological identification of the alternative oxidase of Neurospora crassa mitochondria. Mol. Cell. Biol. 1989, 9, 1362–1364. [Google Scholar] [PubMed]
- Siedow, J.N.; Umbach, A.L. Plant mitochondrial electron transfer and molecular biology. Plant Cell 1995, 7, 821–831. [Google Scholar] [CrossRef]
- Joseph-Horne, T.; Hollomon, D.W.; Wood, P.M. Fungal respiration: A fusion of standard and alternative components. Biochim. Biophys. Acta-Bioenerg. 2001, 1504, 179–195. [Google Scholar] [CrossRef]
- Tudella, V.G.; Curti, C.; Soriani, F.M.; Santos, A.C.; Uyemura, S.A. In situ evidence of an alternative oxidase and an uncoupling protein in the respiratory chain of Aspergillus fumigatus. Int. J. Biochem. Cell Biol. 2004, 36, 162–172. [Google Scholar] [CrossRef]
- McDonald, A.E.; Vanlerberghe, G.C.; Staples, J.F. Alternative oxidase in animals: Unique characteristics and taxonomic distribution. J. Exp. Biol. 2009, 212, 2627–2634. [Google Scholar] [CrossRef]
- Vanlerberghe, G.C.; McIntosh, L. Alternative oxidase: From gene to function. Annu. Rev. Plant Biol. 1997, 48, 703–734. [Google Scholar] [CrossRef]
- Kirimura, K.; Yoda, M.; Shimizu, H.; Sugano, S.; Mizuno, M.; Kino, K.; Usami, S. Contribution of cyanide-insensitive respiratory pathway, catalyzed by the alternative oxidase, to citric acid production in Aspergillus niger. Biosci. Biotechnol. Biochem. 2000, 64, 2034–2039. [Google Scholar] [CrossRef]
- Del-Saz, N.F.; Ribas-Carbo, M.; Martorell, G.; Fernie, A.R.; Florez-Sarasa, I. Measurements of electron partitioning between cytochrome and alternative oxidase pathways in plant tissues. Methods Mol. Biol. 2017, 1670, 203–217. [Google Scholar]
- Williams, B.A.P.; Elliot, C.; Burri, L.; Kido, Y.; Kita, K.; Moore, A.L.; Keeling, P.J. A broad distribution of the alternative oxidase in microsporidian parasites. PLoS Pathog. 2010, 6, e1000761. [Google Scholar] [CrossRef] [PubMed]
- James, T.Y.; Pelin, A.; Bonen, L.; Ahrendt, S.; Sain, D.; Corradi, N.; Stajich, J.E. Shared signatures of parasitism and phylogenomics unite Cryptomycota and microsporidia. Curr. Biol. 2013, 23, 1548–1553. [Google Scholar] [CrossRef] [PubMed]
- Huh, W.K.; Kang, S.O. Characterization of the gene family encoding alternative oxidase from Candida albicans. Biochem. J. 2001, 356, 595–604. [Google Scholar] [CrossRef]
- Tanton, L.L.; Nargang, C.E.; Kessler, K.E.; Li, Q.; Nargang, F.E. Alternative oxidase expression in Neurospora crassa. Fungal Genet. Biol. 2003, 39, 176–190. [Google Scholar] [CrossRef] [PubMed]
- Flipphi, M.; Márton, A.; Bíró, V.; Ág, N.; Sándor, E.; Fekete, E.; Karaffa, L. Mutations in the second alternative oxidase gene: A new approach to group Aspergillus niger strains. J. Fungi 2023, 9, 570. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Kirimura, K.; Ogawa, S.; Hattori, T.; Kino, K. Expression analysis of alternative oxidase gene (aox1) with enhanced green fluorescent protein as marker in citric acid-producing Aspergillus niger. J. Biosci. Bioeng. 2006, 102, 210–214. [Google Scholar] [CrossRef] [PubMed]
- Kirimura, K.; Yoda, M.; Usami, S. Cloning and expression of the cDNA encoding an alternative oxidase gene from Aspergillus niger WU-2223L. Curr. Genet. 1999, 34, 472–477. [Google Scholar] [CrossRef] [PubMed]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef] [PubMed]
- Grigoriev, I.V.; Nikitin, R.; Haridas, S.; Kuo, A.; Ohm, R.; Otillar, R.; Riley, R.; Salamov, A.; Zhao, X.; Korzeniewski, F.; et al. MycoCosm portal: Gearing up for 1000 fungal genomes. Nucleic Acids Res. 2014, 42, D699–D704. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2019, 20, 1160–1166. [Google Scholar] [CrossRef]
- Criscuolo, A.; Gribaldo, S. BMGE (Block Mapping and Gathering with Entropy): A new software for selection of phylogenetic informative regions from multiple sequence alignments. BMC Evol. Biol. 2010, 10, 210. [Google Scholar] [CrossRef]
- Guindon, S.; Dufayard, J.-F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef]
- Anisimova, M.; Gascuel, O. Approximate likelihood-ratio test for branches: A fast, accurate, and powerful alternative. Syst. Biol. 2006, 55, 539–552. [Google Scholar] [CrossRef]
- Arnaud, M.B.; Cerqueira, G.C.; Inglis, D.O.; Skrzypek, M.S.; Binkley, J.; Chibucos, M.C.; Crabtree, J.; Howarth, C.; Orvis, J.; Shah, P.; et al. The Aspergillus Genome Database (AspGD): Recent developments in comprehensive multispecies curation, comparative genomics and community resources. Nucleic Acids Res. 2012, 40, D653–D659. [Google Scholar] [CrossRef]
- Machida, M.; Asai, K.; Sano, M.; Tanaka, T.; Kumagai, T.; Terai, G.; Kusumoto, K.I.; Arima, T.; Akita, O.; Kashiwagi, Y.; et al. Genome sequencing and analysis of Aspergillus oryzae. Nature 2005, 438, 1157–1161. [Google Scholar] [CrossRef]
- Petersen, C.; Sørensen, T.; Nielsen, M.R.; Sondergaard, T.E.; Sørensen, J.L.; Fitzpatrick, D.A.; Frisvad, J.C.; Nielsen, K.L. Comparative genomic study of the Penicillium genus elucidates a diverse pangenome and 15 lateral gene transfer events. IMA Fungus 2023, 14, 3. [Google Scholar] [CrossRef]
- de Vries, R.P.; Riley, R.; Wiebenga, A.; Aguilar-Osorio, G.; Amillis, S.; Uchima, C.A.; Anderluh, G.; Asadollahi, M.; Askin, M.; Barry, K.; et al. Comparative genomics reveals high biological diversity and specific adaptations in the industrially and medically important fungal genus Aspergillus. Genome Biol. 2017, 18, 28. [Google Scholar] [CrossRef]
- Horn, F.; Linde, J.; Mattern, D.J.; Walther, G.; Guthke, R.; Scherlach, K.; Martin, K.; Brakhage, A.A.; Petzke, L.; Valiante, V. Draft genome sequences of fungus Aspergillus calidoustus. Genome Announc. 2016, 4, e00102-16. [Google Scholar] [CrossRef]
- Druzhinina, I.S.; Chenthamara, K.; Zhang, J.; Atanasova, L.; Yang, D.; Miao, Y.; Rahimi, M.J.; Grujic, M.; Cai, F.; Pourmehdi, S.; et al. Massive lateral transfer of genes encoding plant cell wall-degrading enzymes to the mycoparasitic fungus Trichoderma from its plant-associated hosts. PLoS Genet. 2018, 14, e1007322. [Google Scholar] [CrossRef]
- Fierro, F.; Vaca, I.; Castillo, N.I.; García-Rico, R.O.; Chávez, R. Penicillium chrysogenum, a vintage model with a cutting-edge profile in biotechnology. Microorganisms 2022, 10, 573. [Google Scholar] [CrossRef]
- Houbraken, J.; Kocsubé, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.-C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.A.; et al. Classification of Aspergillus, Penicillium, Talaromyces and related genera (Eurotiales): An overview of families, genera, subgenera, sections, series and species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef]
- Kjærbølling, I.; Vesth, T.; Frisvad, J.C.; Nybo, J.L.; Theobald, S.; Kildgaard, S.; Petersen, T.I.; Kuo, A.; Sato, A.; Lyhne, E.K.; et al. A comparative genomics study of 23 Aspergillus species from section. Flavi. Nat. Commun. 2020, 11, 1106. [Google Scholar] [CrossRef] [PubMed]
- Samuels, G.J.; Ismaiel, A.; Bon, M.C.; De Respinis, S.; Petrini, O. Trichoderma asperellum sensu lato consists of two cryptic species. Mycologia 2010, 102, 944–966. [Google Scholar] [CrossRef] [PubMed]
- Chorev, M.; Carmel, L. The function of introns. Front. Genet. 2012, 3, 55. [Google Scholar] [CrossRef] [PubMed]
- Bokor, E.; Flipphi, M.; Kocsubé, S.; Ámon, J.; Vágvölgyi, C.; Scazzocchio, C.; Hamari, Z. Genome organization and evolution of a eukaryotic nicotinate co-inducible pathway. Open Biol. 2021, 11, 210099. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Strain | Relevant Master Accession Numbers | Reference(s) to the Genome Sequences | Source of Live Material | GenBank Accession Numbers [cDNA] * [This Work] |
|---|---|---|---|---|---|
| Aspergillus terreus | NIH 2624 | AAJN [GenBank] | [25] | CBS *** | [aoxA] OR702883 [aoxB2-1] OR702884 |
| Aspergillus oryzae | RIB40 | JZJM [GenBank] | [26] [25] | CBS | [aoxA] OR683635 [aoxB2-1] OR683636 |
| Penicillium rubens ** | NRRL 1951 | AM920416–64 [EMBL] JAQKAF [GenBank] | [27] | Antibióticos S.A.(León, Spain) | [aoxA] OR702887 [aoxB2-1] OR702888 |
| Aspergillus wentii | DTO 134E9 | LJSE [GenBank] | [28] | CBS | [aoxA] OR702885 [aoxB3] OR702886 |
| Aspergillus calidoustus | SF006504 | CDMC [GenBank] | [29] | Hans Knöll Institute (Jena, Germany) | [aoxA] OR714815 [aoxB1] OR631741 [aoxB4] OR631740 |
| Aspergillus sydowii | CBS 593.65 | MRCH [GenBank] | [28] | CBS | [aoxA] OR702890 [aoxB2-1] OR702889 [aoxB3] OR702891 |
| Trichoderma asperellum | CBS 433.97 | MBGH [GenBank] | [30] | CBS | [aoxA] OR683637 [aoxB2-2] OR683638 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Flipphi, M.; Márton, A.; Bíró, V.; Ág, N.; Sándor, E.; Fekete, E.; Karaffa, L. Generation, Transfer, and Loss of Alternative Oxidase Paralogues in the Aspergillaceae Family. J. Fungi 2023, 9, 1195. https://doi.org/10.3390/jof9121195
Flipphi M, Márton A, Bíró V, Ág N, Sándor E, Fekete E, Karaffa L. Generation, Transfer, and Loss of Alternative Oxidase Paralogues in the Aspergillaceae Family. Journal of Fungi. 2023; 9(12):1195. https://doi.org/10.3390/jof9121195
Chicago/Turabian StyleFlipphi, Michel, Alexandra Márton, Vivien Bíró, Norbert Ág, Erzsébet Sándor, Erzsébet Fekete, and Levente Karaffa. 2023. "Generation, Transfer, and Loss of Alternative Oxidase Paralogues in the Aspergillaceae Family" Journal of Fungi 9, no. 12: 1195. https://doi.org/10.3390/jof9121195
APA StyleFlipphi, M., Márton, A., Bíró, V., Ág, N., Sándor, E., Fekete, E., & Karaffa, L. (2023). Generation, Transfer, and Loss of Alternative Oxidase Paralogues in the Aspergillaceae Family. Journal of Fungi, 9(12), 1195. https://doi.org/10.3390/jof9121195