

Detection of Pneumocystis and Morphological Description of Fungal Distribution and Severity of Infection in Thirty-Six Mammal Species

 ,
,  , ,
, ,  ,
,  , , ,
, , , 
Abstract
:
1. Introduction
2. Materials and Methods
3. Results
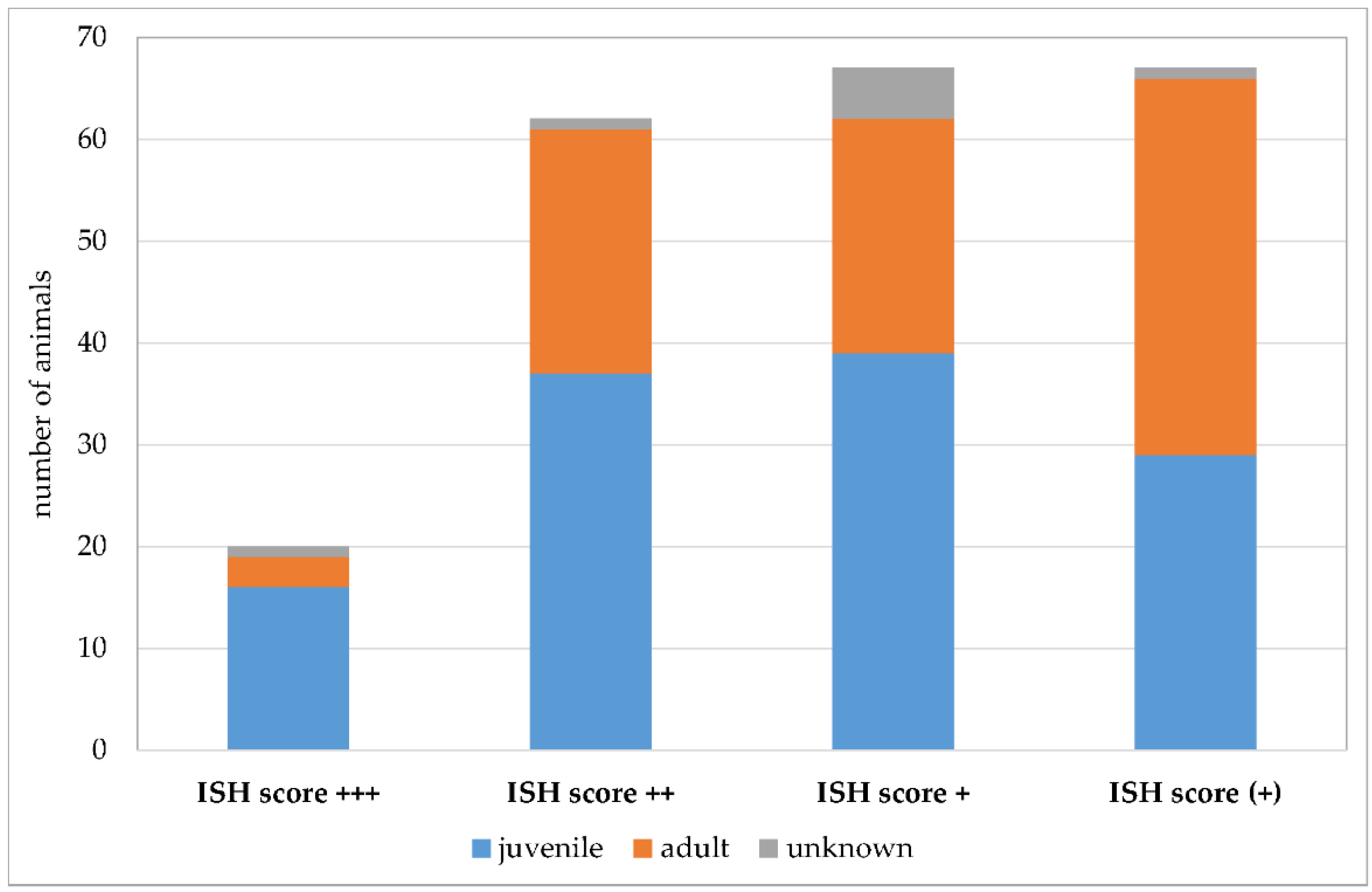
3.1. Many Mammal Species Are Colonized with Pneumocystis spp. though Severe Infections Are Rare
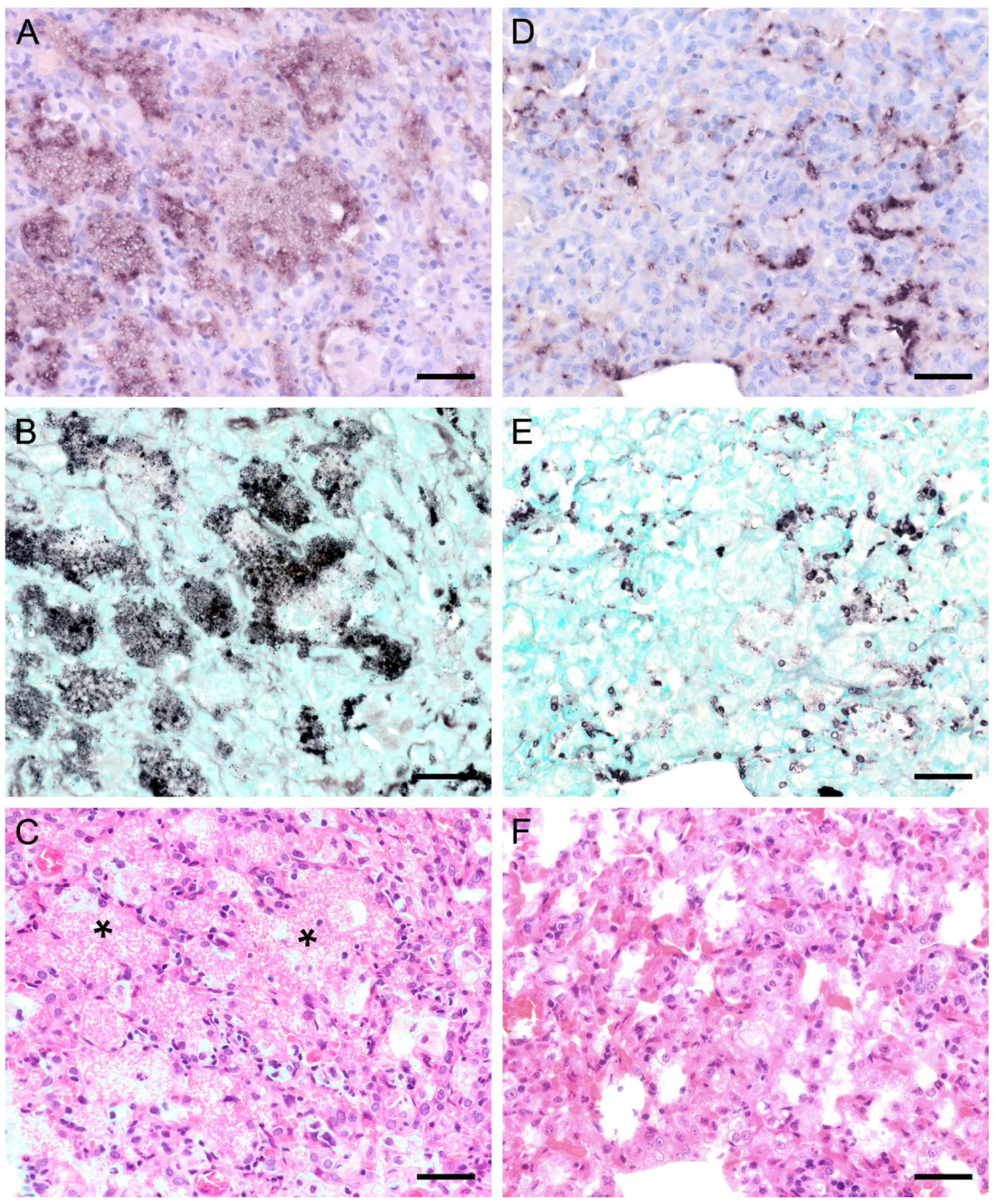
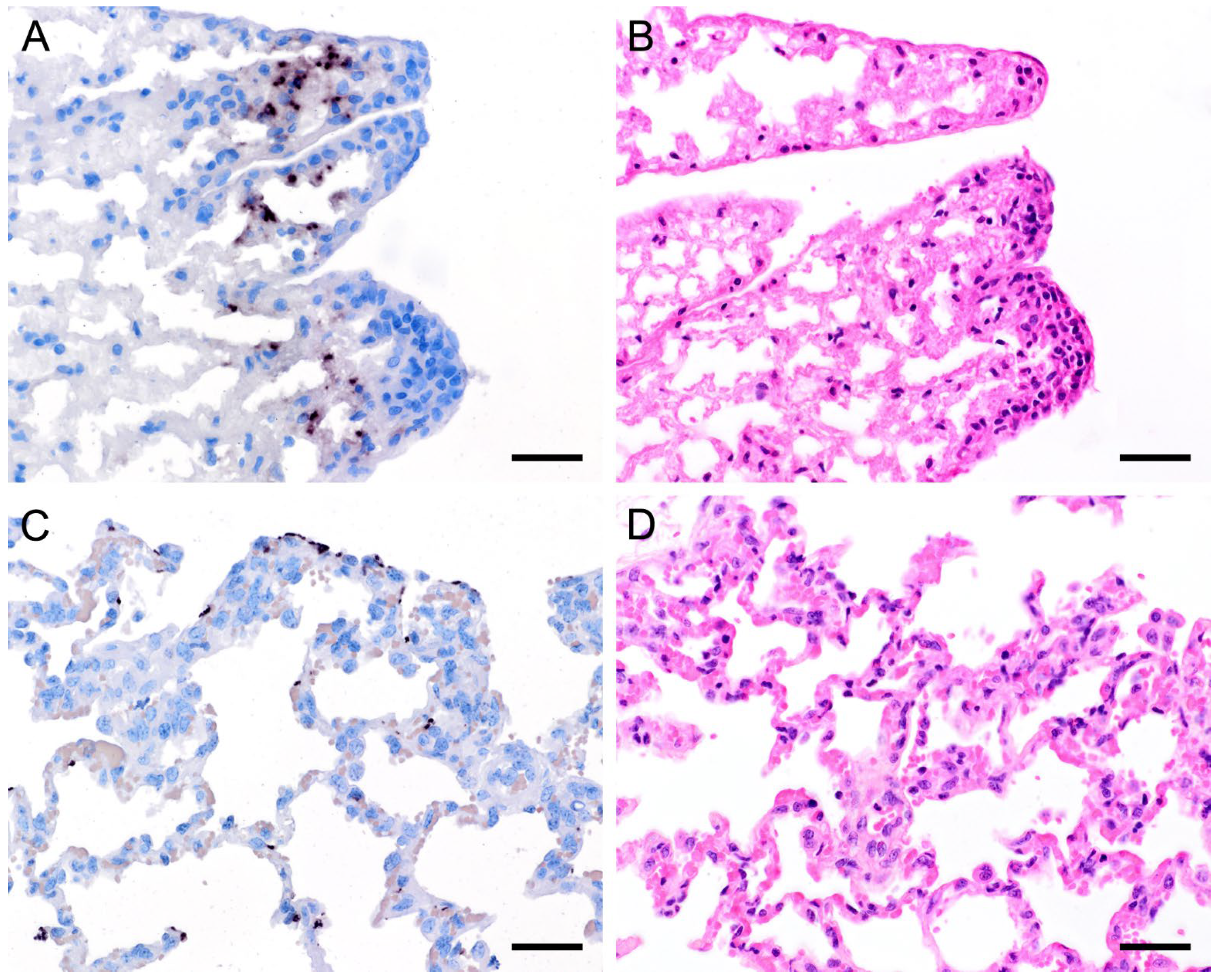
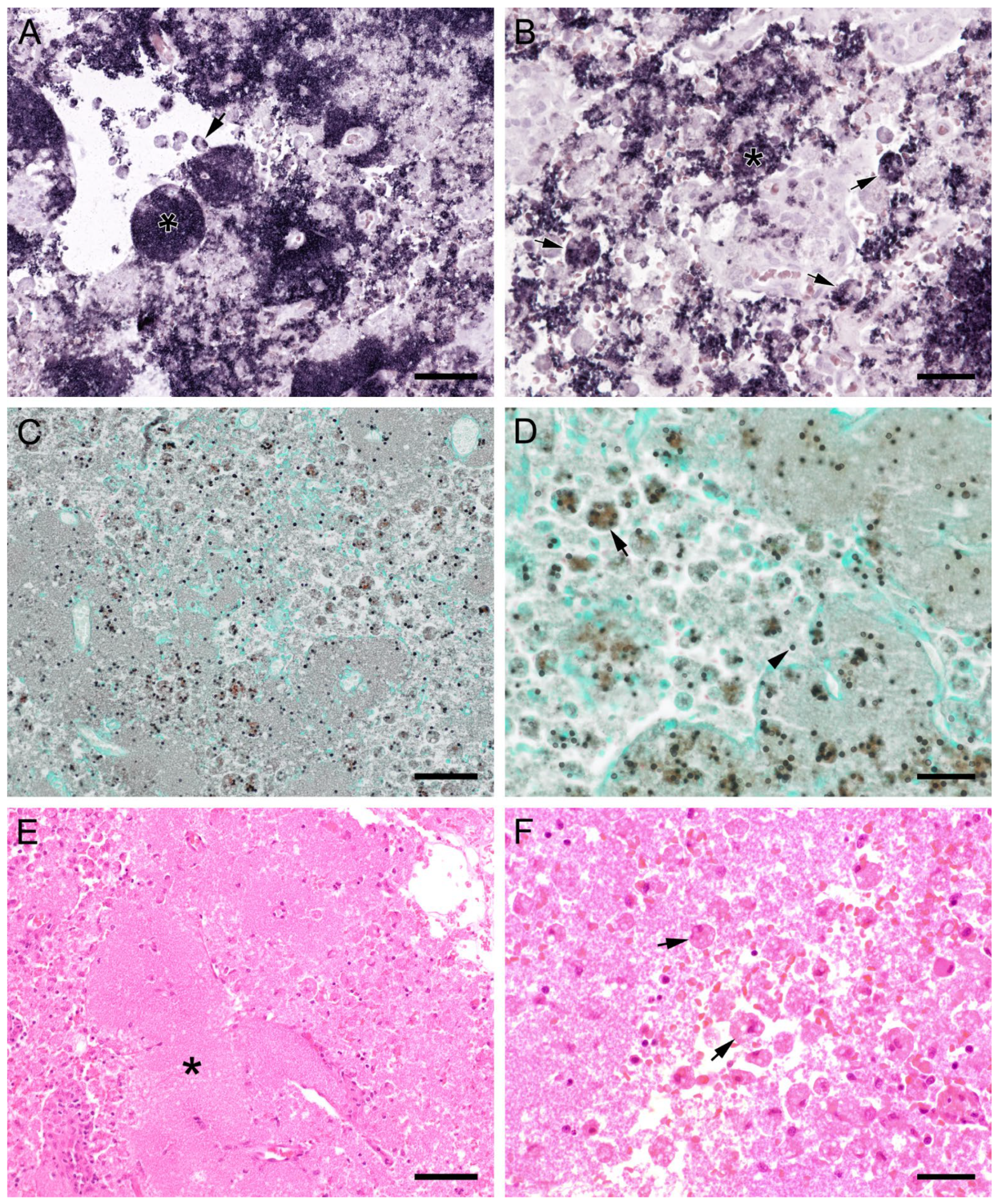
3.2. Pneumocystis Colonizes the Lung with Different Distribution Patterns but the Lung Tissue Is Only Marginally Impaired by the Infection
3.3. Co-Infections and Non-Infectious Concomitant Diseases and Factors
4. Discussion
4.1. Pneumocystis spp. in the Artiodactyla Order
4.2. Pneumocystis spp. in the Carnivora Order
4.3. Pneumocystis spp. in the Chiroptera, Eulipotyphla, Perissodactyla, and Primates Orders
4.4. Pneumocystis spp. in the Lagomorpha and Rodentia Orders (Glires Clade)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aliouat-Denis, C.M.; Chabé, M.; Demanche, C.; Aliouat, E.M.; Viscogliosi, E.; Guillot, J.; Delhaes, L.; Dei-Cas, E. Pneumocystis species, co-evolution and pathogenic power. Infect. Genet. Evol. 2008, 8, 708–726. [Google Scholar] [CrossRef] [PubMed]
- Weyant, R.B.; Kabbani, D.; Doucette, K.; Lau, C.; Cervera, C. Pneumocystis jirovecii: A review with a focus on prevention and treatment. Expert Opin. Pharmacother. 2021, 22, 1579–1592. [Google Scholar] [CrossRef] [PubMed]
- Morris, A.; Wei, K.; Afshar, K.; Huang, L. Epidemiology and clinical significance of Pneumocystis colonization. J. Infect. Dis. 2008, 197, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Guillot, J.; Demanche, C.; Hugot, J.-P.; Berthelemy, M.; Wakefield, A.E.; Dei-Cas, E.; Chermette, R. Parallel phylogenies of Pneumocystis species and their mammalian hosts. J. Eukaryot. Microbiol. 2001, 48, 113S–115S. [Google Scholar] [CrossRef]
- Settnes, O.P.; Henriksen, S.A. Pneumocystis carinii in large domestic animals in Denmark. A preliminary report. Acta Vet. Scand. 1989, 30, 437–440. [Google Scholar] [CrossRef]
- Underwood, W.J.; Morin, D.E.; Mirsky, M.L.; Haschek, W.M.; Zuckermann, F.A.; Petersen, G.C.; Scherba, G. Apparent retrovirus-induced immunosuppression in a yearling llama. J. Am. Vet. Med. Assoc. 1992, 200, 358–362. [Google Scholar]
- Settnes, O.P.; Elvestad, K.; Clausen, B. Pneumocystis carinii Delanoe & Delanoe, 1912 found in lungs of freeliving animals in Denmark at autopsy. Nord. Vet. Med. 1986, 38, 11–15. [Google Scholar]
- Weissenbacher-Lang, C.; Kureljušić, B.; Nedorost, N.; Matula, B.; Schießl, W.; Stixenberger, D.; Weissenböck, H. Retrospective analysis of bacterial and viral co-infections in Pneumocystis spp. positive lung samples of Austrian pigs with pneumonia. PLoS ONE 2016, 11, e0158479. [Google Scholar] [CrossRef]
- Borba, M.R.; Sanches, E.M.C.; Corrêa, A.M.R.; Spanamberg, A.; de Souza Leal, J.; Soares, M.P.; Guillot, J.; Driemeier, D.; Ferreiro, L. Immunohistochemical and ultra-structural detection of Pneumocystis in wild boars (Sus scrofa) co-infected with porcine circovirus type 2 (PCV2) in Southern Brazil. Med. Mycol. 2011, 49, 172–175. [Google Scholar] [CrossRef]
- Weissenbacher-Lang, C.; Fuchs-Baumgartinger, A.; Guija-De-Arespacochaga, A.; Klang, A.; Weissenböck, H.; Künzel, F. Pneumocystosis in dogs: Meta-analysis of 43 published cases including clinical signs, diagnostic procedures, and treatment. J. Vet. Diagn. Investig. 2018, 30, 26–35. [Google Scholar] [CrossRef]
- Riebold, D.; Lubig, J.; Wolf, P.; Wolf, C.; Russow, K.; Loebermann, M.; Slevogt, H.; Mohr, E.; Feldhusen, F.; Reisinger, E.C. First molecular detection of Pneumocystis spp. in red foxes (Vulpes vulpes LINNAEUS, 1758) and raccoon dogs (Nyctereutes procyonoides GRAY, 1834). Comp. Immunol. Microbiol. Infect. Dis. 2020, 73, 101531. [Google Scholar] [CrossRef]
- Danesi, P.; Corrò, M.; Falcaro, C.; Carminato, A.; Furlanello, T.; Cocchi, M.; Krockenberger, M.B.; Meyer, W.; Capelli, G.; Malik, R. Molecular detection of Pneumocystis in the lungs of cats. Med. Mycol. 2019, 57, 813–824. [Google Scholar] [CrossRef]
- Dias, S.; Planellas, M.; Canturri, A.; Martorell, J. Extreme tetralogy of fallot with polycythemia in a ferret (Mustela putorius furo). Top. Companion Anim. Med. 2017, 32, 80–85. [Google Scholar] [CrossRef]
- Sanches, E.M.C.; Pacheco, S.M.; Cericatto, A.S.; Melo, R.M.; Colodel, E.M.; Hummel, J.; Bianchi, S.P.; Spanamberg, A.; Santurio, J.M.; Ferreiro, L. Detection of Pneumocystis in lungs of bats from Brazil by PCR amplification. Pesqui. Vet. Bras. 2009, 29, 469–473. [Google Scholar] [CrossRef]
- Laakkonen, J.; Soveri, T. Characterization of Pneumocystis carinii infection in Sorex araneus from southern Finland. J. Wildl. Dis. 1995, 31, 228–232. [Google Scholar] [CrossRef]
- Mazars, E.; Guyot, K.; Fourmaintraux, S.; Renaud, F.; Petavy, F.; Camus, D.; Dei-Cas, E. Detection of Pneumocystis in European wild animals. J. Eukaryot. Microbiol. 1997, 44, 39S. [Google Scholar] [CrossRef]
- Punsmann, S.; Hoppe, J.; Klopfleisch, R.; Venner, M. Acute interstitial pneumonia in foals: A severe, multifactorial syndrome with lung tissue recovery in surviving foals. Equine Vet. J. 2021, 53, 718–726. [Google Scholar] [CrossRef]
- Poelma, F.G. Pneumocystis carinii infections in zoo animals. Z. Parasitenkd. 1975, 46, 61–68. [Google Scholar] [CrossRef]
- Rothenburger, J.L.; Himsworth, C.G.; Clifford, C.B.; Ellis, J.; Treuting, P.M.; Leighton, F.A. Respiratory pathology and pathogens in wild urban rats (Rattus norvegicus and Rattus rattus). Vet. Pathol. 2015, 52, 1210–1219. [Google Scholar] [CrossRef]
- Hayashimoto, N.; Morita, H.; Ishida, T.; Uchida, R.; Tanaka, M.; Ozawa, M.; Yasuda, M.; Itoh, T. Microbiological survey of mice (Mus musculus) purchased from commercial pet shops in Kanagawa and Tokyo, Japan. Exp. Anim. 2014, 64, 155–160. [Google Scholar] [CrossRef]
- Binanti, D.; Mostegl, M.M.; Weissenbacher-Lang, C.; Nedorost, N.; Weissenböck, H. Detection of Pneumocystis infections by in situ hybridization in lung samples of Austrian pigs with interstitial pneumonia. Med. Mycol. 2014, 52, 196–201. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. 1999, 41, 95–98. [Google Scholar]
- Weissenbacher-Lang, C.; Fuchs-Baumgartinger, A.; Klang, A.; Kneissl, S.; Pirker, A.; Shibly, S.; von Ritgen, S.; Weissenböck, H.; Künzel, F. Pneumocystis carinii infection with severe pneumomediastinum and lymph node involvement in a Whippet mixed-breed dog. J. Vet. Diagn. Investig. 2017, 29, 757–762. [Google Scholar] [CrossRef] [PubMed]
- Kucera, K.; Vanek, J.; Jirovec, O. Pneumozystose [Pneumocystosis]. In Leitfaden der Zooanthroponosen; Toppich, E., Kriiger, W., Eds.; Volk und Gesundheit: Berlin/Heidelberg, Germany, 1971; pp. 279–294. [Google Scholar]
- Jecny, V. Pneumocystis carinii findings in young ones of some farm animals. Epidemiol. Mikrobiol. Immunol. 1973, 22, 135–140. [Google Scholar]
- Shimizu, A.; Kimura, F.; Kimura, S. Occurrence of Pneumocystis carinii in animals in Japan. Jpn. J. Vet. Sci. 1985, 47, 309–311. [Google Scholar] [CrossRef]
- McConnell, E.E.; Basson, P.A.; Pienaar, J.G. Pneumocystosis in a domestic goat. J. Vet. Res. 1971, 38, 117–124. [Google Scholar]
- Sakakibara, M.; Shimizu, C.; Kadota, K.; Hatama, S. Pneumocystis carinii infection in a domestic goat (Capra hircus domesticus) with multibacillary paratuberculosis. J. Vet. Med. Sci. 2013, 75, 671–674. [Google Scholar] [CrossRef]
- Rice, J.A.; Carrasco-Medina, L.; Hodgins, D.C.; Shewen, P.E. Mannheimia haemolytica and bovine respiratory disease. Anim. Health Res. Rev. 2008, 8, 117–128. [Google Scholar] [CrossRef]
- Cushion, M.T.; Linke, M.J.; Ashbaugh, A.; Sesterhenn, T.M.; Collins, M.S.; Lynch, K.; Brubaker, R.; Walzer, P.D. Echinocandin treatment of Pneumocystis pneumonia in rodent models depletes cysts leaving trophic burdens that cannot transmit the infection. PLoS ONE 2010, 5, e8524. [Google Scholar] [CrossRef]
- Bondy, R. Pneumocystis carinii in human and veterinary medicine. Sborn. Ces. Akad. Zemed. Ved. 1958, 3, 199–210. [Google Scholar]
- Nikolskij, S.N.; Shchetinin, A.N. Pneumocystis in swine. Veterinariia 1967, 44, 65. [Google Scholar]
- Kucera, K.; Slesinger, L.; Kadlec, A. Pneumocystosis in pigs. Folia Parasitol. 1968, 15, 75–78. [Google Scholar]
- Seibold, H.R.; Munnell, J.F. Pneumocystis carinii in a pig. Vet. Pathol. 1977, 14, 89–91. [Google Scholar] [CrossRef]
- Fujita, M.; Furuta, T.; Nakajima, T.; Kurita, F.; Kaneuchi, C.; Ueda, K.; Ogata, M. Prevalence of Pneumocystis carinii in slaughtered pigs. Jpn. J. Vet. Sci. 1989, 51, 200–202. [Google Scholar] [CrossRef]
- Bille-Hansen, V.; Jorsal, S.E.; Henriksen, S.A.; Settnes, O.P. Pneumocystis carinii pneumonia in Danish piglets. Vet. Rec. 1990, 127, 407–408. [Google Scholar]
- Sanches, E.M.C.; Borba, M.R.; Spanamberg, A.; Pescador, C.; Corbellini, L.G.; Ravazzolo, A.P.; Driemeier, D.; Ferreiro, L. Co-infection of Pneumocystis carinii f. sp. suis and porcine circovirus-2 (PCV2) in pig lungs obtained from slaughterhouses in southern and midwestern regions of Brazil. J. Eukaryot. Microbiol. 2006, 53, S92–S94. [Google Scholar] [CrossRef]
- Kim, K.S.; Jung, J.Y.; Kim, J.H.; Kang, S.C.; Hwang, E.K.; Park, B.K.; Kim, D.Y.; Kim, J.H. Epidemiological characteristics of pulmonary pneumocystosis and concurrent infections in pigs in Jeju Island, Korea. J. Vet. Sci. 2011, 12, 15–19. [Google Scholar] [CrossRef]
- Zlotowski, P.; de Almeida, P.R.; Boos, G.S.; Sanches, E.M.C.; Ferreiro, L.; Spanamberg, A.; Ravazzolo, A.P.; Driemeier, D. Mixed mycotic rhinitis and pneumonia in wild boars. Acta Sci. Vet. 2011, 39, 1008. [Google Scholar]
- Esgalhado, R.; Esteves, F.; Antunes, F.; Matos, O. Study of the epidemiology of Pneumocystis carinii f. sp. suis in abattoir swine in Portugal. Med. Mycol. 2013, 51, 66–71. [Google Scholar] [CrossRef]
- Blasi, B.; Sipos, W.; Knecht, C.; Dürlinger, S.; Ma, L.; Cissé, O.H.; Nedorost, N.; Matt, J.; Weissenböck, H.; Weissenbacher-Lang, C. Pneumocystis spp. in pigs: A longitudinal quantitative study and co-infection assessment in Austrian farms. J. Fungi 2022, 8, 43. [Google Scholar] [CrossRef]
- Kureljušić, B.; Weissenbacher-Lang, C.; Nedorost, N.; Stixenberger, D.; Weissenböck, H. Association between Pneumocystis spp. and co-infections with Bordetella bronchiseptica, Mycoplasma hyopneumoniae and Pasteurella multocida in Austrian pigs with pneumonia. Vet. J. 2016, 207, 177–179. [Google Scholar] [CrossRef] [PubMed]
- Weissenbacher-Lang, C.; Nedorost, N.; Knecht, C.; Hennig-Pauka, I.; Huber, M.; Voglmayr, T.; Weissenböck, H. Comparison of Pneumocystis nucleic acid and antibody profiles and their associations with other respiratory pathogens in two Austrian pig herds. PLoS ONE 2017, 12, e0185387. [Google Scholar] [CrossRef] [PubMed]
- Sedlmeier, H.; Dahme, E. Pneumocystis carinii infection in a dog. Zentralblatt Allg. Pathol. 1955, 93, 150–155. [Google Scholar]
- Van den Akker, S.; Goedbloed, E. Pneumonia caused by Pneumocystis carinii in a dog. Trop. Geogr. Med. 1960, 12, 54–58. [Google Scholar]
- Avalos, A.D. Latent infection produced by Pneumocystis carinii in domestic animals in Mexico City. Salud Publica Mex. 1963, 5, 975. [Google Scholar]
- Zavala, V.; Rosado, E.R. Pneumocystis carinii in domestic animals of the city of Mérida, Yucatán. Salud Publica Mex. 1972, 14, 103–106. [Google Scholar]
- Farrow, B.R.H.; Watson, A.D.J.; Hartley, W.J.; Huxtable, C.R.R. Pneumocystis pneumonia in the dog. J. Comp. Pathol. 1972, 82, 447–452. [Google Scholar] [CrossRef]
- Sheldon, W.H. Experimental pulmonary Pneumocystis carinii infection in rabbits. J. Exp. Med. 1959, 110, 147–160. [Google Scholar] [CrossRef]
- Copland, J.W. Canine pneumonia caused by Pneumocystis carinii. Aust. Vet. J. 1974, 50, 515–518. [Google Scholar] [CrossRef]
- Kirberger, R.M.; Lobetti, R.G. Radiographic aspects of Pneumocystis carinii pneumonia in the miniature Dachshund. Vet. Radiol. Ultrasound 1998, 39, 313–317. [Google Scholar] [CrossRef]
- Lobetti, R.G. Common variable immunodeficiency in miniature dachshunds affected with Pneumonocystis carinii pneumonia. J. Vet. Diagn. Investig. 2000, 12, 39–45. [Google Scholar] [CrossRef]
- Lobetti, R.G.; Leisewitz, A.L.; Spencer, J.A. Pneumocystis carinii in the miniature dachshund: Case report and literature review. J. Small Anim. Pract. 1996, 37, 280–285. [Google Scholar] [CrossRef]
- McCully, R.M.; Lloyd, J.; Kuys, D.; Schneider, D.J. Canine Pneumocystis pneumonia. J. S. Afr. Vet. Assoc. 1979, 50, 207–209. [Google Scholar]
- Botha, W.S.; van Rensburg, I.B. Pneumocystosis: A chronic respiratory distress syndrome in the dog. J. S. Afr. Vet. Assoc. 1979, 50, 173–179. [Google Scholar]
- Brownlie, S.E. A retrospective study of diagnosis in 109 cases of canine lower respiratory disease. J. Small Anim. Pract. 1990, 31, 371–376. [Google Scholar] [CrossRef]
- Hagiwara, Y.; Fujiwara, S.; Takai, H.; Ohno, K.; Masuda, K. Pneumocystis carinii pneumonia in a Cavalier King Charles Spaniel. J. Vet. Med. Sci. 2001, 63, 349–351. [Google Scholar] [CrossRef]
- Meffert, F.J. Pneumocystis pneumonia in two Cavalier King Charles Spaniel littermates. Aust. Vet. Pract. 2009, 39, 2–9. [Google Scholar]
- Okine, A.A.K.; Chapman, S.; Hostutler, R.A.; Livingston, R.S. Diagnosis of Pneumocystis pneumonia in a 2-year-old King Charles Cavalier Spaniel using the polymerase chain reaction. Vet. Clin. Pathol. 2018, 47, 146–149. [Google Scholar] [CrossRef]
- Ramsey, I.K.; Foster, A.; McKay, J.; Herrtage, M.E. Pneumocystis carinii pneumonia in two Cavalier King Charles spaniels. Vet. Rec. 1997, 140, 372–373. [Google Scholar] [CrossRef]
- Sukura, A.; Saari, S.; Järvinen, A.K.; Olsson, M.; Marjatta, K.; Timo, I. Pneumocystis carinii pneumonia in dogs—A diagnostic challenge. J. Vet. Diagn. Investig. 1996, 32, 124–130. [Google Scholar] [CrossRef]
- Watson, P.J.; Wotton, P.; Eastwood, J.; Swift, S.T.; Jones, B.; Day, M.J. Immunoglobulin deficiency in Cavalier King Charles Spaniels with Pneumocystis pneumonia. J. Vet. Intern. Med. 2006, 20, 523–527. [Google Scholar] [CrossRef] [PubMed]
- Petini, M.; Furlanello, T.; Danesi, P.; Zoia, A. Nested–polymerase chain reaction detection of Pneumocystis carinii f. sp. canis in a suspected immunocompromised Cavalier King Charles spaniel with multiple infections. SAGE Open Med. Case Rep. 2019, 7, 2050313X1984116. [Google Scholar] [CrossRef]
- Merrill, K.; Coffey, E.; Furrow, E.; Masseau, I.; Rindt, H.; Reinero, C. X-linked CD40 ligand deficiency in a 1-year-old male Shih Tzu with secondary Pneumocystis pneumonia. J. Vet. Intern. Med. 2021, 35, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Cabañes, F.J.; Roura, X.; Majö, N.; Bragulat, M.R.; Domingo, M. Pneumocystis carinii pneumonia in a Yorkshire terrier dog. Med. Mycol. 2000, 38, 451–453. [Google Scholar] [CrossRef]
- Best, M.P.; Boyd, S.P.; Danesi, P. Confirmed case of Pneumocystis pneumonia in a Maltese Terrier × Papillon dog being treated with toceranib phosphate. Aust. Vet. J. 2019, 97, 162–165. [Google Scholar] [CrossRef]
- Danesi, P.; Ravagnan, S.; Johnson, L.R.; Furlanello, T.; Milani, A.; Martin, P.; Boyd, S.P.; Best, M.P.; Galgut, B.; Irwin, P.; et al. Molecular diagnosis of Pneumocystis pneumonia in dogs. Med. Mycol. 2017, 55, 828–842. [Google Scholar] [CrossRef]
- Furuta, T.; Nogami, S.; Kojima, S.; Fujita, M.; Kamata, H.; Kuwabara, M.; Ohba, S.; Yoshida, H. Spontaneous Pneumocystis carinii infection in the dog with naturally acquired generalised demodicosis. Vet. Rec. 1994, 134, 423–424. [Google Scholar] [CrossRef]
- Kanemoto, H.; Morikawa, R.; Chambers, J.K.; Kasahara, K.; Hanafusa, Y.; Uchida, K.; Ohno, K.; Nakayama, H. Common variable immune deficiency in a Pomeranian with Pneumocystis carinii pneumonia. J. Vet. Med. Sci. 2015, 77, 715–719. [Google Scholar] [CrossRef]
- Sukura, A.; Laakkonen, J.; Rudbäck, E. Occurrence of Pneumocystis carinii in canine distemper. Acta Vet. Scand. 1997, 38, 201–205. [Google Scholar] [CrossRef]
- Giannoulopoulos, G.; Lobetti, R. Pneumocystosis in a lurcher puppy. Med. Mycol. Case Rep. 2020, 30, 15–18. [Google Scholar] [CrossRef]
- Sakashita, T.; Kaneko, Y.; Izzati, U.Z.; Hirai, T.; Fuke, N.; Torisu, S.; Yamaguchi, R. Disseminated pneumocystosis in a toy poodle. J. Comp. Pathol. 2020, 175, 85–89. [Google Scholar] [CrossRef]
- Danesi, P.; Petini, M.; Falcaro, C.; Bertola, M.; Mazzotta, E.; Furlanello, T.; Krockenberger, M.; Malik, R. Pneumocystis colonization in dogs is as in humans. Int. J. Environ. Res. Public Health 2022, 19, 3192. [Google Scholar] [CrossRef]
- Settnes, O.P.; Hasselager, E. Occurrence of Pneumocystis carinii Delanoë & Delanoë, 1912 in dogs and cats in Denmark. Nord. Vet. Med. 1984, 36, 179–181. [Google Scholar]
- Wöhrer, D.; Spergser, J.; Bagrinovschi, G.; Möstl, K.; Weissenböck, H. Age-related presence of selected viral and bacterial pathogens in paraffin-embedded lung samples of dogs with pneumonia. Acta Vet. Hung. 2016, 64, 103–115. [Google Scholar] [CrossRef]
- Shiota, T.; Shimada, Y.; Kurimoto, H.; Oikawa, H. Pneumocystis carinii infection in corticosteroid-treated cats. J. Parasitol. 1990, 76, 441–445. [Google Scholar] [CrossRef]
- Hagler, D.N.; Kim, C.K.; Walzer, P.D. Feline leukemia virus and Pneumocystis carinii infection. J. Parasitol. 1987, 73, 1284–1286. [Google Scholar] [CrossRef]
- Klang, A.; Loncaric, I.; Spergser, J.; Eigelsreiter, S.; Weissenböck, H. Disseminated histoplasmosis in a domestic cat imported from the USA to Austria. Med. Mycol. Case Rep. 2013, 2, 108–112. [Google Scholar] [CrossRef]
- Dyer, N.W.; Schamber, G.J. Pneumocystosis associated with canine distemper virus infection in a mink. Can. Vet. J. 1999, 40, 577–578. [Google Scholar]
- Lainson, R.; Shaw, J.J. Pneumocystis and Histoplasma infections in wild animals from the Amazon region of Brazil. Trans. R. Soc. Trop. Med. Hyg. 1975, 69, 505–508. [Google Scholar] [CrossRef]
- Akbar, H.; Pinçon, C.; Aliouat-Denis, C.M.; Derouiche, S.; Taylor, M.L.; Pottier, M.; Carreto-Binaghi, L.H.; González-González, A.E.; Courpon, A.; Barriel, V.; et al. Characterizing Pneumocystis in the lungs of bats: Understanding Pneumocystis evolution and the spread of Pneumocystis organisms in mammal populations. Appl. Environ. Microbiol. 2012, 78, 8122–8136. [Google Scholar] [CrossRef]
- González-González, A.E.; Aliouat-Denis, C.M.; Ramírez-Bárcenas, J.A.; Demanche, C.; Pottier, M.; Carreto-Binaghi, L.E.; Akbar, H.; Derouiche, S.; Chabé, M.; Aliouat, E.M.; et al. Histoplasma capsulatum and Pneumocystis spp. co-infection in wild bats from Argentina, French Guyana, and Mexico. BMC Microbiol. 2014, 14, 23. [Google Scholar] [CrossRef] [PubMed]
- Sanches, E.M.C.; Ferreiro, L.; Andrade, C.P.; Pacheco, S.M.; Almeida, L.L.; Spanamberg, A.; Wissmann, G. Pneumocystis spp. in bats evaluated by qPCR. J. Mycol. Med. 2013, 23, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Veloso, S.S.C.; Ferreiro, L.; Pacheco, S.M.; da Silva, R.R.P.; Souza, E.d.C.; Machado, G.; Wissmann, G.; Spanamberg, A.; Sanches, E.M.C. Pneumocystis spp. and Histoplasma capsulatum in bats lungs in Southern and Midwestern regions of Brazil. Acta Sci. Vet. 2014, 42, 1252. [Google Scholar]
- Derouiche, S.; Deville, M.; Taylor, M.L.; Akbar, H.; Guillot, J.; Carreto-Binaghi, L.H.; Pottier, M.; Aliouat, E.M.; Aliouat-Denis, C.M.; Dei-Cas, E.; et al. Pneumocystis diversity as a phylogeographic tool. Mem. Inst. Oswaldo Cruz 2009, 104, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Danesi, P.; da Rold, G.; Rizzoli, A.; Hauffe, H.C.; Marangon, S.; Samerpitak, K.; Demanche, C.; Guillot, J.; Capelli, G.; de Hoog, S.G. Barcoding markers for Pneumocystis species in wildlife. Fungal Biol. 2016, 120, 191–206. [Google Scholar] [CrossRef]
- Bishop, R.; Gurnell, J.; Laakkonen, J.; Whithwell, K.; Peters, S.E. Detection of Pneumocystis DNA in the lungs of several species of wild mammal. J. Eukaryot. Microbiol. 1997, 44, 57S. [Google Scholar] [CrossRef]
- Laakkonen, J.; Fisher, R.N.; Case, T.J. Pneumocystosis in wild small mammals from California. J. Wildl. Dis. 2001, 37, 408–412. [Google Scholar] [CrossRef]
- Laakkonen, J.; Sukura, A.; Haukisalmi, V.; Henttonen, H. Pneumocystis carinii and helminth parasitism in shrews Sorex araneus and Sorex caecutiens. J. Wildl. Dis. 1993, 29, 273–277. [Google Scholar] [CrossRef]
- Laakkonen, J.; Kallio, E.R.; Kallio-Kokko, H.; Vapalahti, O.; Vaheri, A.; Henttonen, H. Is there an association of Pneumocystis infection with the presence of arena-, hanta-, and poxvirus antibodies in wild mice and shrews in Finland? Parasitology 2006, 132, 461–466. [Google Scholar] [CrossRef]
- Ainsworth, D.M.; Weldon, A.D.; Beck, K.A.; Rowland, P.H. Recognition of Pneumocystis carinii in foals with respiratory distress. Equine Vet. J. 1993, 25, 103–108. [Google Scholar] [CrossRef]
- Clark-Price, S.C.; Cox, J.H.; Bartoe, J.T.; Davis, E.G. Use of dapsone in the treatment of Pneumocystis carinii pneumonia in a foal. J. Am. Vet. Med. Assoc. 2004, 224, 407–410+371. [Google Scholar] [CrossRef]
- Ewing, P.J.; Cowell, R.L.; Tyler, R.D.; MacAllister, C.G.; Meinkoth, J.H. Pneumocystis carinii pneumonia in foals. J. Am. Vet. Med. Assoc. 1994, 204, 929–933. [Google Scholar]
- Franklin, R.P.; Long, M.T.; MacNeill, A.; Alleman, A.R.; Giguère, S.; Uhl, E.; López-Martinez, A.; Wilkerson, M. Proliferative interstitial pneumonia, Pneumocystis carinii infection, and immunodeficiency in an adult Paso Fino horse. J. Vet. Intern. Med. 2002, 16, 607–611. [Google Scholar] [CrossRef]
- MacNeill, A.L.; Alleman, A.R.; Franklin, R.P.; Long, M.; Giguère, S.; Uhl, E.; López-Martinez, A.; Wilkerson, M. Pneumonia in a Paso-Fino mare. Vet. Clin. Pathol. 2003, 32, 73–76. [Google Scholar] [CrossRef]
- Perron Lepage, M.F.; Gerber, V.; Suter, M.M. A case of interstitial pneumonia associated with Pneumocystis carinii in a foal. Vet. Pathol. 1999, 36, 621–624. [Google Scholar] [CrossRef]
- Prescott, J.F.; Wilcock, B.P.; Carman, P.S.; Hoffman, A.M. Sporadic, severe bronchointerstitial pneumonia of foals. Can. Vet. J. 1991, 32, 421–425. [Google Scholar]
- Perryman, L.E.; McGuire, T.C.; Crawford, T.B. Maintenance of foals with combined immunodeficiency: Causes and control of secondary infections. Am. J. Vet. Res. 1978, 39, 1043–1047. [Google Scholar]
- Tanaka, S.; Ochiai, K.; Itakura, C.; Kaji, Y.; Taniyama, H.; Matsukawa, K. Pneumocystis carinii pneumonia in a thoroughbred foal. J. Vet. Med. Sci. 1994, 56, 135–137. [Google Scholar] [CrossRef]
- Jensen, T.K.; Boye, M.; Bille-Hansen, V. Application of fluorescent in situ hybridization for specific diagnosis of Pneumocystis carinii pneumonia in foals and pigs. Vet. Pathol. 2001, 38, 269–274. [Google Scholar] [CrossRef]
- Hugot, J.-P.; Demanche, C.; Barriel, V.; Dei-Cas, E.; Guillot, J. Phylogenetic systematics and evolution of primate-derived Pneumocystis based on mitochondrial or nuclear DNA sequence comparison. Syst. Biol. 2003, 52, 735–744. [Google Scholar] [CrossRef]
- Demanche, C.; Berthelemy, M.; Petit, T.; Polack, B.; Wakefield, A.E.; Dei-Cas, E.; Guillot, J. Phylogeny of Pneumocystis carinii from 18 primate species confirms host specificity and suggests coevolution. J. Clin. Microbiol. 2001, 39, 2126–2133. [Google Scholar] [CrossRef] [PubMed]
- Icenhour, C.R.; Rebholz, S.L.; Collins, M.S.; Cushion, M.T. Early acquisition of Pneumocystis carinii in neonatal rats as evidenced by PCR and oral swabs. Eukaryot. Cell 2002, 1, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Cushion, M.T.; Zhang, J.; Kaselis, M.; Giuntoli, D.; Stringer, S.L.; Stringer, J.R. Evidence for two genetic variants of Pneumocystis carinii coinfecting laboratory rats. J. Clin. Microbiol. 1993, 31, 1217–1223. [Google Scholar] [CrossRef] [PubMed]
- Latinne, A.; Bezé, F.; Delhaes, L.; Pottier, M.; Gantois, N.; Nguyen, J.; Blasdell, K.; Dei-Cas, E.; Morand, S.; Chabé, M. Genetic diversity and evolution of Pneumocystis fungi infecting wild Southeast Asian murid rodents. Parasitology 2018, 145, 885–900. [Google Scholar] [CrossRef]
- Latinne, A.; Chen, H.W.; Kuo, C.C.; Lorica, R.; Singleton, G.; Stuart, A.; Malbas, F.F.; Demanche, C.; Chabé, M.; Michaux, J.; et al. Revisiting the Pneumocystis host specificity paradigm and transmission ecology in wild Southeast Asian rodents. Infect. Genet. Evol. 2021, 93, 104978. [Google Scholar] [CrossRef]
- Chabé, M.; Herbreteau, V.; Hugot, J.-P.; Bouzard, N.; Deruyter, L.; Morand, S.; Dei-Cas, E. Pneumocystis carinii and Pneumocystis wakefieldiae in wild Rattus norvegicus trapped in Thailand. J. Eukaryot. Microbiol. 2010, 57, 213–217. [Google Scholar] [CrossRef]
- Palmer, R.J.; Settnes, O.P.; Lodal, J.; Wakefield, A.E. Population structure of rat-derived Pneumocystis carinii in Danish wild rats. Appl. Environ. Microbiol. 2000, 66, 4954–4961. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host Order | Host Family | Number of Animals | ||||
|---|---|---|---|---|---|---|
| Positive | ISH Score +++ | ISH Score ++ | ISH Score + | ISH Score (+) | ||
| Even-toed ungulates (Artiodactyla) (n = 380) | Bovidae (n = 96) | 25 | 2 | 0 | 10 | 13 |
| Camelidae (n = 23) | 2 | 0 | 0 | 1 | 1 | |
| Cervidae (n = 21) | 3 | 0 | 0 | 1 | 2 | |
| Suidae (n = 240) | 114 | 13 | 51 | 38 | 12 | |
| Carnivores (Carnivora) (n = 156) | Canidae (n = 31) | 19 | 1 | 1 | 5 | 12 |
| Felidae (n = 76) | 5 | 0 | 0 | 1 | 4 | |
| Mustelidae (n = 47) | 26 | 1 | 8 | 5 | 12 | |
| Procyonidae (n = 2) | 1 | 0 | 0 | 0 | 1 | |
| Bats (Chiroptera) (n = 18) | Pteropodidae (n = 6) | 0 | 0 | 0 | 0 | 0 |
| Rhinolophidae (n = 1) | 0 | 0 | 0 | 0 | 0 | |
| Vespertilionidae (n = 11) | 1 | 0 | 0 | 0 | 1 | |
| Insectivores (Eulipotyphla) (n = 41) | Erinaceidae (n = 17) | 0 | 0 | 0 | 0 | 0 |
| Soricidae (n = 19) | 1 | 0 | 0 | 1 | 0 | |
| Talpidae (n = 3) | 0 | 0 | 0 | 0 | 0 | |
| Tupaiidae (n = 2) | 0 | 0 | 0 | 0 | 0 | |
| Lagomorphs (Lagomorpha) (n = 14) | Leporidae (n = 14) | 3 | 1 | 0 | 0 | 2 |
| Odd-toed ungulates (Perissodactyla) (n = 36) | Equidae (n = 36) | 5 | 0 | 0 | 3 | 2 |
| Primates (Primates) (n = 47) | Aotidae (n = 1) | 0 | 0 | 0 | 0 | 0 |
| Callitrichidae (n = 19) | 0 | 0 | 0 | 0 | 0 | |
| Cebidae (n = 10) | 0 | 0 | 0 | 0 | 0 | |
| Cercopithecidae (n = 14) | 0 | 0 | 0 | 0 | 0 | |
| Hominidae (n = 2) | 0 | 0 | 0 | 0 | 0 | |
| Pitheciidae (n = 1) | 0 | 0 | 0 | 0 | 0 | |
| Rodents (Rodentia) (n = 153) | Castoridae (n = 1) | 0 | 0 | 0 | 0 | 0 |
| Caviidae (n = 12) | 2 | 0 | 1 | 0 | 1 | |
| Chinchillidae (n = 6) | 1 | 0 | 0 | 0 | 1 | |
| Cricetidae (n = 10) | 1 | 0 | 0 | 0 | 1 | |
| Echimyidae (n = 8) | 0 | 0 | 0 | 0 | 0 | |
| Muridae (n = 107) | 7 | 2 | 2 | 1 | 2 | |
| Octodontidae (n = 4) | 0 | 0 | 0 | 0 | 0 | |
| Sciuridae (n = 5) | 0 | 0 | 0 | 0 | 0 | |
| Host Family | Exclusively Positive | Exclusively Negative | Positive or Negative |
|---|---|---|---|
| Bovidae | Chamois (Rupicapra rupicapra), water buffalo (Bubalus bubalis) | Cattle (Bos taurus), sheep (Ovis aries), Goat (Capra hircus), Blackbuck (Antilope cervicapra), Bison (Bos bonasus) | |
| Camelidae | Bactrian camel (Camelus bactrianus) | Llama (Lama glama), Arabian camel (Camelus dromedarius) | Alpaca (Vicugna pacos) |
| Cervidae | Red deer (Cervus elaphus), Sika deer (Cervus nippon), Visayan spotted deer (Cervus alfredi), Reindeer (Rangifer tarandus), Pere David’s deer (Elaphurus davidianus) | Western roe deer (Capreolus capreolus), Deer (Cervus) 1 | |
| Suidae | Domestic pig (Sus scrofa domesticus), Wild boar (Sus scrofa) | ||
| Canidae | Red fox (Vulpes vulpes) | Dog (Canis lupus familiaris), Gray wolf (Canis lupus), Eastern Canadian wolf (Canis lupus lycaon) | |
| Felidae | Wild cat (Felis silvestris), Eurasian lynx (Lynx lynx), Lion (Panthero leo), Tiger (Panthera tigris), Puma (Puma concolor), leopard (Panthera pardus) | Cat (Felis catus) | |
| Mustelidae | Oriental small-clawed otter (Aonyx cinereus), Northern American river otter (Lontra canadensis), European mink (Mustela lutreola) | Marten (Martes) 1 | Beach marten (Martes foina), Eurasian badger (Meles meles), Ferret (Mustela putorius furo), Eurasian river otter (Lutra lutra), Striped skunk (Mephitis mephitis) |
| Procyonidae | Racoon (Procyon lotor) | ||
| Pteropodidae | Indian flying fox (Pteropus giganteus), Flying fox (Pteropus) 1, short-nosed fruit bat (Cynopterus) 1 | ||
| Rhinolophidae | Lesser horseshoe bat (Rhinolophus hipposidenus) | ||
| Vespertilionidae | Bat (Vespertilio) 1, Noctule (Nyctalus noctula), Savi’s pipistrelle (Hypsugo savii), Common serotine (Eptesicus serotinus), Whiskered bat (Myotis mystacinus) | Particolored bat (Vespertilio murinus) | |
| Erinaceidae | Western European hedgehog (Erinaceus europaeus), Middle-African hedgehog (Atelerix albiventris), Northern white-breasted hedgehog (Erinaceus roumanicus) | ||
| Soricidae | European shrew (Sorex araneus) | Bicolored shrew (Crocidura leucodon), Lesser white-toothed shrew (Crocidura suaveolens), Eurasian pygmy shrew (Sorex minutus) | |
| Talpidae | European mole (Talpa europaea) | ||
| Tupaiidae | Common tree shrew (Tupaia glis) | ||
| Leporidae | European brown hare (Lepus europaeus), Rabbit (Oryctolagus cuniculus) | ||
| Equidae | Donkey (Equus asinus), Pony (Equus caballus), Plains zebra (Equus quagga) | Horse (Equus caballus) | |
| Aotidae | Night monkey (Aotus trivirgatus) | ||
| Callitrichidae | Brown-headed tamarin (Leontocebus fuscicollis), Cotton-top tamarin (Saguinus oedipus), Golden lion tamarin (Leontopithecus rosalia), Common marmoset (Callithrix jacchus) | ||
| Cebidae | White-faced sapajou (Cebus capucinus), Common squirrel monkey (Saimiri sciureus) | ||
| Cercopithecidae | King colobus (Colobus polykomos), Barbary ape (Macaca sylvanus), Gelada (Theropithecus gelada), Mantled guereza (Colobus guereza), Long-tailed monkey (Cercopithecus, Miopithecus) 1, Rhesus monkey (Macaca mulatta) | ||
| Hominidae | Orang-utan (Pongo) 1 | ||
| Pitheciidae | White-faced saki (Pithecia pithecia) | ||
| Castoridae | Eurasian beaver (Castor fiber) | ||
| Caviidae | Rock cavy (Kerodon rupestris) | Guinea pig (Cavia porcellus) | |
| Chinchillidae | Long-tailed chinchilla (Chinchilla lanigera) | ||
| Cricetidae | Dzhungarian hamster (Phodopus sungorus), Golden hamster (Mesocricetus auratus) | Black-bellied hamster (Cricetus cricetus) | |
| Echimyidae | Nutria (Myocastor coypus) | ||
| Muridae | Mouse (Mus musculus), Spiny mouse (Mus saxicola), Mongolian gerbil (Meriones unguiculatus) | Rat (Rattus norvegicus, Rattus rattus) | |
| Octodontidae | Degu (Octodon degus) | ||
| Sciuridae | Swinehoe’s striped squirrel (Tamiops swinhoei), Eurasian red squirrel (Sciurus vulgaris), chipmunk (Tamias) 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weissenbacher-Lang, C.; Blasi, B.; Bauer, P.; Binanti, D.; Bittermann, K.; Ergin, L.; Högler, C.; Högler, T.; Klier, M.; Matt, J.; et al. Detection of Pneumocystis and Morphological Description of Fungal Distribution and Severity of Infection in Thirty-Six Mammal Species. J. Fungi 2023, 9, 220. https://doi.org/10.3390/jof9020220
Weissenbacher-Lang C, Blasi B, Bauer P, Binanti D, Bittermann K, Ergin L, Högler C, Högler T, Klier M, Matt J, et al. Detection of Pneumocystis and Morphological Description of Fungal Distribution and Severity of Infection in Thirty-Six Mammal Species. Journal of Fungi. 2023; 9(2):220. https://doi.org/10.3390/jof9020220
Chicago/Turabian StyleWeissenbacher-Lang, Christiane, Barbara Blasi, Patricia Bauer, Diana Binanti, Karin Bittermann, Lara Ergin, Carmen Högler, Tanja Högler, Magdalena Klier, Julia Matt, and et al. 2023. "Detection of Pneumocystis and Morphological Description of Fungal Distribution and Severity of Infection in Thirty-Six Mammal Species" Journal of Fungi 9, no. 2: 220. https://doi.org/10.3390/jof9020220